0022-538X/91/084334-07$02.00/0
CopyrightC) 1991, AmericanSociety forMicrobiology
Rotavirus
Spike Structure and Polypeptide Composition
INDUMATHY D. ANTHONY, STANLEYBULLIVANT, SHOBHNADAYAL, A.RICHARD BELLAMY,* ANDJOHN A. BERRIMANt
Departmentof Cellular and Molecular Biology, University of Auckland, Auckland, New Zealand Received 29 November1990/Accepted 10 May 1991
Negativelystainedpreparationsof rotavirusimagedwithalowdose of electronsprovidesufficientcontrast toreveal surfaceprojectionsorspikes.The number ofspikesfoundprojectingfrom differentparticles indicates thatnot all 60 peripentonalsites are occupied. Treatment atpH 11.2 with 250 mM ammonium hydroxide specificallyremovesthe spikes, yieldingsmooth double-shelled particles of the samediameterasthat ofthe native virus. Protein analysis confirmsthat the released spikesarecomposed ofpolypeptide VP4(orits two cleavageproducts VP5* and VP8*)and that thesmooth particle retains theothermajoroutershell protein VP7.Spikeless particles canbedecoratedby amonoclonalantibody specific for themajorimmunodominant
neutralizingdomainof VP7, implyingthat removalofthespikesdoesnotdenature the VP7that is retainedon
thesurfaceof the smoothparticle.
The outershell of thedouble-layered rotavirus particle is constructed fromtwo viral proteins. The most abundant of these is VP7, a 38-kDa glycoprotein which is the
type-specific antigen (9, 12). VP7 is the translational product of genomic segment 9 (3, 17). The second protein, VP4 (88 kDa), is encoded by genomic segment 4 and is the viral hemagglutinin, a protein which also specifies cell tropism (9). Both VP7 and VP4 are assembled on the surface of the
single-shelled rotavirus particle at some stage during the maturation of the virus in thelumen ofthe rough
endoplas-mic reticulum (20, 21). However, when trypsin is incorpo-ratedintotheculture medium, VP4is cleaved proteolytically to yieldVP5* (60 kDa)and VP8* (28kDa), anevent which greatly enhances infectivity in vitro (4, 6, 7). The cleavage sitesprobably resideatArg-241and Arg-247, residues which areconserved in most strains of rotavirusfor whichcDNAs have been sequenced (2).
The capsomeres of the inner shell have been shown by platinum shadowing to be arranged in a T = 13L configura-tion (19, 24). Prasadetal. (23) appliedcryoelectron micros-copy and image-averaging techniques to construct a model for therotavirus particle with aresolution of 4.0nm. Their work revealed that the outershell is also arranged in aT =
13L symmetry and that there are 60 surface projections
(spikes) approximately4.5 nmlong and 3.5nmwide. They
estimatedfrom their model that eachspike was one molecule of VP4. In a subsequent analysis (22), the spike was
de-scribed as athin structure extending to a height of 4.5 nm, withafurther well-defined globular domain of approximately 5.5nmindiameter, acrosswhich lies abilobedstructure4.0 nmwide and 7.0nmacross. The estimated mass of the spike and the fact that two Fab fragments appeared to bind to each spike led these researchers to infer that the spike was a dimer of VP4.
Yeager et al. (26) have also reconstructed images of rotavirus from cryoelectron micrographs and similarly de-scribe the spike as having a complex bilobed morphology. The spikes were found to be shorter by the reconstruction method than could be measured on the micrograph, and they
* Correspondingauthor.
tPresent address: Laboratory of Molecular Biology, Medical ResearchCouncil, CambridgeCB22QH, England.
suggested that morphological variability would be lostas a result of the averaging implicit in the reconstruction proce-dure. Indiscussing the oligomeric nature of the spike, they drew attention to the large disparity of stoichiometry be-tween VP4 and VP7 from densitometer measurements of stainedpolyacrylamide gels (15) and suggested thatpurified viruses may well not all carry the full complement of 60 spikes. In a study of rotavirus by the freeze-etch method (la), it was found that while spikes on the surface of the intact virus could beclearlydemonstrated, occupancy of the peripentonal sites was low.
Here we present new evidence on the nature of thespikes by using negatively stained preparations examined with a low dose (26) of electrons (low-dose images). This method has enabled us totest many different extraction conditions for their effect on the virus. We have found that treatment of theintact virion with the weak base ammoniumhydroxideat pH 11.2 removes the spikes, releases VP4 (or its cleavage products VP5* and VP8*) into the supernatant, and yields smoothparticles that are otherwise intact.
MATERIALSANDMETHODS
Cellsand virus. The SAl1 strain ofrotavirus was
propa-gated in 1,585-cm2 roller bottle cultures of MA104 cells as described previously (25), usingculture mediumcontaining 25 pLgoftrypsin (Hazleton Biologics Inc., Lenexa, Kans.) per ml. For the preparation of virus which contained the uncleaved form of VP4, this medium was removed 6 h
postinfection and the monolayerswereextensivelywashed andreincubated with mediumlacking trypsinandcontaining 1%fetal calfserum. Theinfected cells were then harvested 40 h postinfection. Double-shelled virus was purified by
banding on CsCl gradients that contained 10mM Tris HCl (pH 7.5) and 10 mM CaCl2 (25) and stored at 4°C in CsCl priorto use.
Trypsin cleavage ofVP4 was achieved by first dialyzing thevirusagainstTBS (140mM NaCl,0.7 mM Na2HPO4,5 mM KCI, 20mMTris HCl [pH 7.5]), followed bydigestion
with1-,ug/mltolylsulfonylphenylalanylchloromethyl ketone (TPCK)-treated trypsin (Sigma type XIII) for 30 minat37°C.
Aprotinin (100 ,uglml; Serva GmbH & Co., Heidelberg,
Germany)wasaddedtoinhibitfurthertrypsindigestion,the viruswasrebandedon apreformed CsCl gradient, and then 4334
on November 10, 2019 by guest
http://jvi.asm.org/
CsCl was removed by dialysis against TBS. Samples were prepared for microscopy at concentrations of about 2 mg of viral protein per ml.
NH40H treatment of virus. Virus at 0.5mg/ml in 4 M CsCl was dialyzed against a solution containing 25 mM NaCl, 1 mM Tris HCl (pH 7.5), and 1 mM CaCl2. Aprotinin was added to a final concentration of 10,ug/ml. NH40H (1.25 M) (prepared from a 25% NH3 solution [analytical grade]; BDH, Poole, England) was added to the virus preparation to give a final concentration of 250 mMNH40H, and thepreparation was incubated at room temperature for 25 min. The treated virus (100,ul)was then overlaid onto a 20-,ul cushion of 30% sucrose prepared in TBS and contained in an airfuge tube (5 by 20 mm; Beckman Instruments, Inc., Palo Alto, Calif.). The mixture was then centrifuged at 22 lb/in2 for 90 s in a Beckman Airfuge with an A-100/30 rotor to pellet thesmooth particles which were resuspended in TBS for electron mi-croscopy. For protein analysis by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (14), the supernatant (80pul) was first neutralized with acetic acid, and the protein was concentrated by precipitation with 9 vol-umes of ethanol at -20°Cfor 1 h.
Electron microscopy. Specimens were examined by using the low-dose facility of a Philips CM12 electronmicroscope operating at 80 kV. Suitable areas were recorded onKodak SO-163 film at a magnification of x28,000 and a dose of about 10 electrons per 0.01 nm2. The dose was calibrated against the speed of the film, which was developed in full-strength D19 for 12 min. The magnification was verified with catalase crystal spacings and maintained by using a constant (eucentric) objective lens current.
Cryoelectron microscopy. Films with perforations were made by dipping cleaned microscope slides into a 0.5% solution of collodion in acetone. The dried films were examined by phase-contrast microscopy to assess size and distribution of holes. Suitable films were floated ontowater, and 400-mesh copper grids were applied to the surface. These grids were picked up, coated with platinum-carbon (2.0 nm) toimprove electrical conductivity, and then coated with carbon (10.0 nm) using afreeze-fracture coating source. The plastic layer was dissolved away by rinsing individual grids in amyl acetate, and after drying the grids were examined once more. The very high contrast of theplatinum layer made it possible to identify false holes or bounded depressions. A
5-pI
sample of virus was applied to thegrid,which was then blotted andplunged into liquid ethane (1). Without warming, the grid wastransferred toliquidnitrogen
and inserted into a cold stage(Gatan Inc.,Warrendale, Pa.) designed to maintain the specimen in amorphousice (5). To minimize contamination with ice in the microscope, a blade-type anticontaminator (10) was used in addition to the standard cold trap.
Negative staining. Optimal results were obtained using freshly prepared 10% uranylformate. One-milliliteraliquots of 1% uranyl acetate weredispensed into Eppendorf tubes. Then 0.1 ml of 1.0 M NaOH was added,and thetubes were sealed and shaken. The precipitate was sedimented by centrifugation, and the supernatant was discarded. The pellet was dissolved in 100 p1 of5% formic acid byvigorous
mixing and used immediately. A carbon-coated grid was floated on 5
p,l
ofvirus on a Parafilmsurface (AmericanCan Co., Greenwich, Conn.). After about 1 min, the grid was sequentially washed and stained bytransfer across 350-pd
drops of TBS and 2 drops of stain before itwasblotteddry
with filter paper held to the side of the grid.
Hemagglutination assays. Virus samples in TBS were
diluted in microtitration dishes
(Behring
Institute,
Behring-werke, Marburg, Germany),
yielding
dilutedsamples
of100-,ulvolume. Anequalvolumeofa
freshly
prepared
0.5% suspension of human group 0erythrocytes
(suspended
in thesamebuffer)
wasaddedtoeachwell,
and the assaywasread after 3 hat roomtemperature.
RESULTS
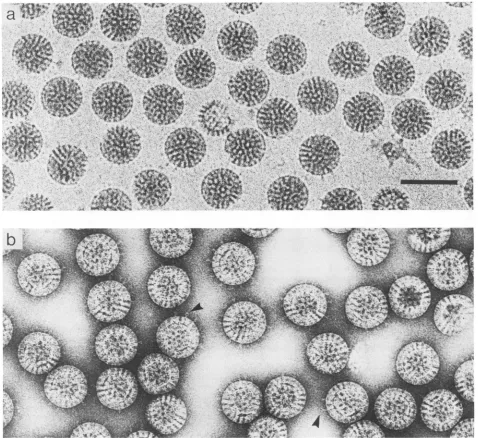
Rotavirus spike morphology.
Images
of rotavirus in ice show low-contrastspikes
extending
from the surface of the intact virions (Fig. la) as describedpreviously
(23, 27).
Low-dose uranyl formate-stained
images
(Fig.
lb)
also re-veal these structures, which were radiation sensitive and whichwerelostusing
conventionalimaging
methods.Figure
lb reveals that different numbers of
spikes
project
as a coronafrom individualparticles.
Thenumberofspikes
isso low for some virions that the variation cannot be due to chance orientation but rather must be due to a reduced occupancy of the 60 sites in thecapsid.
Anotherfeature of the negatively stainedspikes
is the presence ofapparent Y andlooped conformations(indicated
by
arrowheads inFig.
lb). However, while
initially
thiscould be takenassupport-ive evidence that the
spike
isdimeric,
this may be due toimagesof
neighboring
spikes
being
superimposed.
Virus that had not beenexposed
totrypsin
showed the same widevariability in numbers of
spikes
(Fig. 2a),
and these were morefrequently
seen assingle
straight
structures. The absenceofYandlooped
conformations inundigested
prep-arationscould be
interpreted
asindicating
thedependence
of these structures onprior
trypsin
treatment, but itmight
merely be the consequence of
variability
instaining
condi-tionsbetweenpreparations.
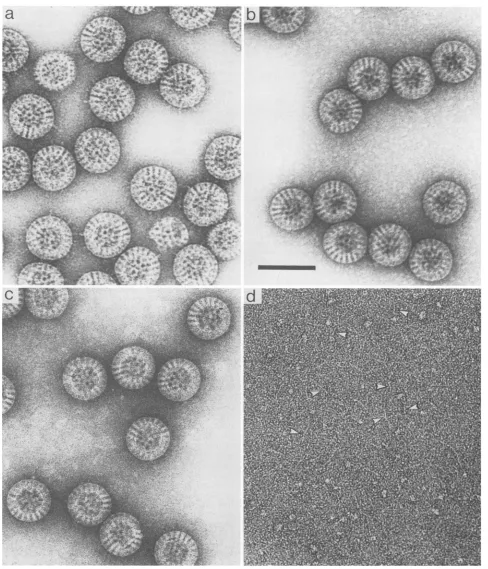
Removal of spikes by treatment at elevated
pH.
A wide rangeofexperimental
conditions wereinvestigated
for theirabilityto remove
spikes
from the intact virion as observedbylow-dose electron
microscopy
andgel
electrophoresis.
Asanticipated,
thespikes
wereresistant tosonication,
isopyc-nic
banding
onCsClgradients,
and other methodstradition-allyused invirus
purification.
Digestion
withawide rangeofproteolytic
enzymes,including
chymotrypsin,
thermolysin,
papain,
trypsin,
subtilisin,
pepsin,
and V8 protease, was foundtobeineffective.Treatmentwith heat and exposureto urea, methods whichsuccessfully
release reovirusspikes
(8), also were ineffectual. The nonionic
detergents
TritonX-100, Nonidet
P-40,
and,-octyl
glucoside
also failed todisrupt orsolubilize theviral
proteins.
While
investigating
the effects ofpH,
the best method whichreproducibly
removed thespikes
yetyielded
other-wise intactparticles
was found to be treatment with dilute ammoniumhydroxide.
Higher
concentrations of this weak base were found to release anincreasing
proportion
ofspikes, withaconcentration of 250 mM
being
optimal (Fig.
2b) atpH 11.2. Factors other than
pH
must be involved in this processasNaOH-phosphate
andNaOH-glycine
bufferscausedreleaseataround
pH
13 butalsodisrupted
thevirus. The material in thebackground
ofFig.
2b is taken to be releasedspike
protein.
Whenseparated
by
centrifugation,
the
resuspended
pellet
brought
topH
7.4(Fig.
2c)
gaveimages of intact
particles
with agood
circular outline and evidence forgood
structuralintegrity.
Intheseviruses,
itcanbe seen that the stain-filled channels
running through
the viral coat are morehighly
contrasted where thespikes
areabsent.
The supernatantderived from the
NH40H
treatment(Fig.
2d) was free of intact
particles
and viral capsomeres buton November 10, 2019 by guest
http://jvi.asm.org/
vu
~~
AFIG. 1. Rotavirus(isolatedin thepresence oftrypsin) imaged by cryoelectron microscopy inamorphous iceandby low-dosenegative stainingwithuranyl formate.(a) Animage ofthe unstainedpreparation,underfocusedby3,um, shows therandomly orientedintactparticles in the thinlayer (about100 nmthick) of ice. Spikescanbe seenprojectingfrom thesurfacewithanindication ofaterminal domain.(b)In
astainedpreparation imagedcloser tofocus,theprojectionsexhibitloopedand bilobed conformations(indicated by arrowheads)but these maybethe resultof overlap betweenpairs or groups of spikes. Bar, 100 nm.
contained material exhibiting globular and short fibrous structures. Althoughthis material could not be identified by microscopy, SDS-PAGE (Fig. 3) showed that the released material (lanes4and8) wasalmostexclusivelyVP4(orVP5* and VP8* in trypsin-treated virus). The spike protein was almost completely released by alkaline treatment while the major external protein VP7wasretained (Fig. 3, lanes 3 and 7). Minor amounts of VP6 present on the gel must derive from a small fraction of particles that are either disrupted
during centrifugation or inefficiently sedimented, because this proteinwasalso present in the controls (Fig. 3, lanes 2 and 6).
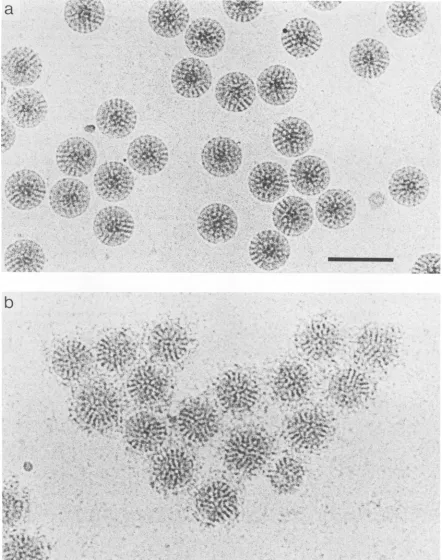
To confirm that the proteins of the virus have not been denatured by the ammonium hydroxide treatment, the
treatedparticleswereincubated withamonoclonalantibody
that recognizes the serotype-specific epitope present on
region A of the single large immunodominant neutralizing
domain of VP7 (18). Cryoelectron microscopy (Fig. 4) re-vealed that the monoclonal antibody bound to the smooth
particle, implying that alkaline treatment had not
grossly
denatured theVP7proteinwhich remainedonthesurface of thesmoothparticle.
Hemagglutination.Inview of theassignmentof VP4asthe viralhemagglutinin(11), thehemagglutinatingability (HA)of virus treatedtoeither cleaveorremoveVP4wasdetermined by using human erythrocytes. Table 1 shows that virus
prepared in the presence oftrypsinexhibited aconsistently
lowerhemagglutinationtiter(approximatelytwo- tofourfold
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.79.557.77.515.2]FIG. 2. Effect of ammonium hydroxide treatment on rotavirus as revealed by negative staining and low-dose microscopy. (a) The uncleaved (VP4) preparationof virus (cf. with Fig. lb) shows manysingle spikeprojections. (b) Virus preparation followingadditionof ammonium hydroxide and incubation at pH 11.2. Note the complete release ofthe spikes from the virus surface. The material in the backgroundmay be releasedspikeproteinwhich appearsaggregatedwhenstainedatthishighpH. (c) Particles separated bycentrifugation and brought to neutral pH have a smooth circularprofile and show very clear stain-filled channels radiating through the capsid. The background is free of released protein. (d)ThesupernatantbroughttoneutralpHyields images(white arrowheads) ofshort(20-to40-nm) linear moleculeswhich may be the releasedspike protein. Small globular material may be either denaturedprotein fromthespikeorthe inner capsidVP6. Bar, 100nm.
0, fc.
b
'41..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.56.544.75.644.2]Trypsin
pH
11.2 + +P S P S P S PS
1 1 1 A C A< 7
VPI -VP2,3 -VP4f VP5* -VP6 VP7 -VP8* -20
FIG. 3. The spikeis composedofpolypeptideVP4orits cleav-age productsVP5* and VP8*. 12.5% SDS-PAGEanalysisof rota-viruspolypeptidesderived from virusbearingintact VP4(lanes1 to
4)ortrypsin-cleaved VP4 (VP5* andVP8*, lanes 5 to 8). Control virus (lanes 1, 2, 5, and6) was centrifugedto yield pellet(P) and
supernatant(S)fractions. Pellets andsupernatants generatedby pH
11.2treatmentareinlanes 3 and 4(uncleavedvirus)andlanes7 and
8(cleaved virus).Lane9 contains molecularmassmarkers(sizesin
kilodaltons). LargearrowsindicateVP4;smallarrowsindicate VP5*
and(faint)VP8* bands.
less for an equivalent number ofparticles) than virus
pre-pared inthe absence oftrypsin (preparation 1 versus 2 and
preparation 3 versus 4). Preparation 5,whichwas grown in the absence oftrypsin but then subsequently digested with
trypsin, also showed a reductionin HAtiter. Thus, trypsin cleavage consistently yielded virus that exhibited a lower HAtiter. Followingalkaline treatmenttoremoveVP4,both cleaved and uncleaved viruslost all detectable HA and no
HAcould be recovered from thesupernatant.
DISCUSSION
In their study of tobacco mosaic virus, Williams and
Fisher(26) appliedlow-dose methods toreveal finedetail of negativelystainedpreparations. Cryoelectronmicroscopyis
dependentonthesetechniquesbecauseof thehighradiation sensitivity of unstained biological molecules in ice. The
contrast in suchan imageismuch lower thanthatproduced by stained specimens, but the preparation is free of the compressionanddistortion causedbydrying. Consequently, computational averaging methods are used to overcome
noise and generate athree-dimensionalmodel. Prasad et al. (23) and Yeager et al. (27) applied icosahedral averaging
methods to images of rotavirus produced by cryoelectron microscopyandderived the structureof theaveraged spike.
Their work revealed the spike to have a complex bilobed
morphologyandFabdecoration led to theinferencethat the spike is adimerof VP4(22).
Ourfindingsshowthat thesurfaceprojectionsof rotavirus canbe imagedin stainbylow-dose methods. In the proteo-lyzed state, the Y andlooped conformations observed sug-gestthe opening up ofa dimer, in agreement with the Fab
decoration data (22). The spike images, however, could be superimpositions, because inthe polarprojection,
contribu-tions totheimagemaybe madefromboth aboveandbelow theequator. A full interpretationof the structuresobserved
would be dependenton thegeneration ofacomplete three-dimensional model; such ananalysis isbeyondthe scope of this report and in anyeventcould bemisleadingbecause of the distorting effects created by specimen
dehydration.
Neithercanpossible variations in conditionsduringstaining
be excludedas apossible explanationforourfindingthat the spike is in a straight conformation when unproteolyzed.
However, ourresults dosuggest thatarigorous analysis of the effects oftrypsinon spike morphology should be under-takenby cryoelectron microscopyandimage analysis.
By specifically releasing the spikes from the virus, we have shown that the constituent protein is VP4 (VP5* and VP8* aftertrypsin treatment). This observation agrees with the results of Fab decoration(22) which also localized VP4 tothespike. We have also shown that VP7 doesnotform a part of thespikeand thatfollowingreleaseofVP4,the
major
neutralizing epitope of VP7 is correctly presented on the surface of the virus. Consequently, it is reasonable to conclude that the presence of VP4 is not necessary fora structurally sound outershell, indicatingthatVP4 and VP7 probably do not formjoint domains on the surface of the virus. The lack of expected stoichiometry found between these proteins (15) and the frequently observed "bald" regions in virus images (both in ice and in stain) could be explained in the following two ways. (i) VP4, which lacks a signal sequence (16) and is cytoplasmic, associates rather randomly with the virus as it buds through the membrane (20, 21). (ii) Some of the spikes could be lost during isolation of the virus. Since the virus does not encounterahighpH
environmentduring the purification proto ol and the spikes
on the virus areotherwise very refractory to extraction, loss ofspikes during purification appears unlikely.
The closely related reovirus also has surface projections
and these have been shown to be composed of a single
protein, sigma 1 (8). There is no sequence homology be-tween VP4 and sigma 1, and there is no evidence for a coiled-coil alpha-helical domain in VP4. However, as both theseproteinsareviralhemagglutinins, the appearanceofan apical globular domain on VP4 may be linked with this activity. The fact that reduced HA was found for trypsin-treated rotavirus is enigmatic but confirms the observations of Kitaoka et al. (13); trypsin treatment is known to enhance infectivity (4, 6, 7) and might be expected to increase, rather than reduce, the interaction of virus with cell membranes. We have shown(Fig. 3) that trypsin does not release either of the two cleavage products (VP5* and VP8*) from the virus. Thechange in HA may be linked to a possible change inshape of the spike. Two reconstructions of rotavirus have shown spikes with bilobed distal domains. Yeageretal. (27) point out the effect low occupancy of spikes on the virus surface would have on the averaged model. In light ofour findings, we believe that it is important to determine the extentofproteolysis in preparations studied by cryoelectron
microscopy so that any structural alterations which occur
following cleavage can be defined.
Finally, this work has demonstrated that the smooth
(spikeless) particles which lack VP4 retain their structural integrity. They havean undistortedcircularprofile andgive every expectation of being suitable candidates for image reconstructionusing cryoelectron microscopy (2a). By
com-parisonwith nativevirus,itshould bepossible to determine thestructureof theanchoringpartofVP4within the capsid. Further investigation of the smooth particle therefore may
helpto unraveltheinteractions of the inner and outershell
proteins of rotavirus and the structural features that sur-round the base of the spike.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.79.303.72.252.2]IL "
's.
.I;:t '*
I* I>..,I
)*#~
.*.r,tN4. "I,'
*4.¶ r .' .
a '; S A
rw ees.
$4;)e *
FIG. 4. Smooth particles retain the epitopefor theVP7-specificmonoclonalantibody 159. (a) Smooth particles preparedasforFig. 2c but unstained inamorphous ice andunderfocused by3,um.(b)Smoothparticles preparedasin panelabutdecoratedwith monoclonal antibody 159(18) by mixing thealkali-treated virus with antibody and incubatingatroomtemperaturefor2 hpriortofreezing. Bar, 100nm.
I*I. t:
*.y,i X
*,W,r *
a
4'.
S.
- 't.r
$7'L4r.
vt,p ,t_
iC.1X Wf
Ae
H,4 -,4.v
b
*0*.
1-4
't lb.
.
.0""
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.77.519.100.660.2]TABLE 1. Hemagglutinationby rotavirus particlesfollowing trypsin cleavage or pH11.2treatment
Virus Cleavage Alkali HAtiter
prepn' stateb treatment (U/mg ofvirus)
1 - - 11,428
2 + - 5,818
3 - - 22,068
4 + - 7,655
5 - - 22,068
+ - 11,034
6 + - 27,526
+ + <100
7 - - 102,400
+ <200
a Seven individuallypurified viruspreparationsyielded differentHAtiters.
bCleavage of>98% (+) and <10% (-), as determined by SDS-PAGE
analysis.
ACKNOWLEDGMENTS
This workwas supported bygrants fromthe Medical Research Council ofNewZealand and theNew ZealandChildren's Health ResearchFoundation.J.A.B. was a NewZealandUniversity Grants Committee Research Fellow.
Wethank AlasdairSteven forhelpful criticismof themanuscript. HarryGreenberggenerouslysuppliedtheVP7-specific monoclonal antibody159 andprovidedhelpful suggestions forthepreparation of uncleaved virus.WethankBarryBenning fortheconstruction ofa supplementary coldtrapfor the electronmicroscope.
REFERENCES
1. Adrian, M., J. Dubochet, J. Lepault, and A. McDowell. 1984. Cryo-electron microscopy of viruses.Nature(London) 308:32-36.
la.Bailey, C.I. 1988.M.S. thesis. UniversityofAuckland, Auck-land, NewZealand.
2. Bellamy, A. R., and G. W. Both. 1990. Molecular biology of rotaviruses. Adv. Virus Res. 38:1-43.
2a.Berriman,J. A., M. Yeager, and A. R.Bellamy. Unpublished data.
3. Both, G. W., J. S. Mattick, and A. R. Bellamy. 1983. Serotype-specific glycoprotein of simian-11 rotavirus: coding assignment andgenesequence. Proc. Natl.Acad. Sci. USA 80:3091-3095. 4. Clark,S.M., J. R.Roth,M. L.Clark,B. B.Barnett,and R.S. Spendlove. 1981. Trypsinenhancementof rotavirusinfectivity: mechanisms ofenhancement. J.Virol.39:816-822.
5. Dubochet, J., J. Lepault, R. Freeman, J. A. Berriman, andJ.-C. Homo.1982. Electron microscopy of frozenwaterandaqueous solutions.J. Microsc. 128:219-237.
6. Espejo,R.T.,S. Lopez, and C. Arias. 1981. Structural polypep-tides of simian rotavirusSAl and the effect oftrypsin. J.Virol. 37:156-160.
7. Estes, M. K., D. Y. Graham, and B. B. Mason. 1981.Proteolytic enhancementof rotavirusinfectivity: molecularmechanisms. J. Virol.39:879-888.
8. Furlong, D.B., M. L. Nibert, and B. N. Fields. 1988. Sigma 1 protein of mammalian reoviruses extends from the surfaces of viralparticles.J. Virol. 62:246-256.
9. Greenberg,H.B., J.Valdesuso,K. van Wyke, K. Midthun, M.
Walsh, V.McAuliffe, R.G.Wyatt, A. R.Kalica,J.Flores,and Y. Hoshino. 1983. Production andpreliminarycharacterization of monoclonal antibodies directed at two surface proteins of
rhesusrotavirus. J. Virol. 47:267-275.
10. Homo, J.-C.,F. Booy, P. Labouesse,J. Lepault,andJ. Dubo-chet. 1984. Improved anticontaminator for cryo-electron mi-croscopywithaPhilipsEM400. J. Microsc. 136:337-341. 11. Kalica,A.R.,J.Flores,and H. B. Greenberg.1983.
Identifica-tion of the rotaviralgene that codes forhemagglutination and
protease-enhanced plaqueformation.Virology 125:194-205. 12. Kalica, A.R., H. B.Greenberg,R.G.Wyatt,J. Flores,M. M.
Sereno, A. Z. Kapikian, and R. M. Chanock. 1981. Genes of human(strainWa)andbovine
(strain
UK)rotaviruses that code forneutralizationandsubgroupantigens.Virology
112:385-390. 13. Kitaoka, S., H. Suzuki, T. Numazaki, T. Sato, T. Konno, T. Ebina,N. Ishida,0. Nakagomi, and T. Nakagomi. 1984.Hem-agglutinationbyhumanrotavirus strains.J.Med.Virol. 13:215-222.
14. Laemmli,U. K.1970.Cleavageof structural
proteins during
theassembly ofthe head of
bacteriophage
T4. Nature(London)
227:680-685.
15. Lui, M.,P.A.Offit,and M.K. Estes.1988. Identification ofthe simian rotavirus SAl1genome segment 3
product.
Virology
163:26-32.
16. Mackow, E. R., R. D. Shaw, S. M. Matsui, P. T. Vo, M.-N. Dang, and H. B. Greenberg. 1988. The rhesus rotavirus gene
encoding protein VP3: location of amino acids involved in
homologousandheterologousrotavirusneutralisation and iden-tification of a putative fusion region. Proc. Natl. Acad. Sci. USA85:645-649.
17. Mason,B.B.,D. Y.Graham,and M. K.Estes.1983. Biochem-ical mapping of the simian rotavirus
SAl1
genome. J. Virol. 46:413-423.18. Matsui, S. M., E. R. Mackow, and H. B. Greenberg. 1989. Molecular determinants ofrotavirusneutralization and protec-tion.Adv. Virus Res. 36:181-214.
19. Metcalf,P.1982.Thesymmetry of reovirus.J.Ultrastruct.Res. 78:292-301.
20. Petrie,B.L.,D.Y.Graham,H.Hansson,and M. K. Estes.1982. Localisation of rotavirusantigensininfected cellsby ultrastruc-turalimmunocytochemistry. J. Gen. Virol. 63:457-467. 21. Poruchynsky, M. S., C. Tyndall, G. W. Both, F. Sato, A. R.
Bellamy,and P. H. Atkinson.1985.Deletionsinan
NH2-terminal
hydrophobic domain result in secretion of rotavirus VP7, a residentendoplasmicreticulummembraneglycoprotein.J. Cell Biol. 101:2199-2209.22. Prasad,B. V.V.,J.W. Burns,E.Marietta,M. K.Estes,and W. Chiu. 1990.Localisation ofVP4neutralisation sites in rotavirus
by three-dimensional cryo-electron microscopy. Nature (Lon-don)343:476-479.
23. Prasad, B. V. V., G. J. Wang, J. P. M. Clerx, and W. Chiu. 1988. Three-dimensional structure of rotavirus. J. Mol. Biol. 199:269-275.
24. Roseto, A.,J.Escaig,E.Delain,J.Cohen,andR. Scherrer.1979. Structure of rotaviruses as studiedby the
freeze-drying
tech-nique. Virology98:471-475.
25. Street, J. E., M. C. Croxson, W. F. Chadderton, and A. R. Bellamy. 1982. Sequence diversity ofhuman rotavirus strains
investigated by Northern blot hybridizationanalysis. J. Virol. 43:369-378.
26. Williams,R.C.,and H. W. Fisher.1970. Electronmicroscopyof tobacco mosaicvirus underconditions of minimalbeam
expo-sure. J. Mol. Biol. 52:121-123.
27. Yeager,M., K. A.Dryden, N.H.Olsen, H. B.Greenberg, and
T.S. Baker. 1990.Three-dimensional structureof rhesus
rota-virusbycryo-electronmicroscopyandimagereconstruction. J. CellBiol. 110:2133-2144.