h ttp://ww w . n a t u r e z a e c o n s e r v a c a o . c o m . b r
Natureza
&
Conservação
Brazilian
Journal
of
Nature
Conservation
SupportedbyBoticárioGroupFoundationforNatureProtection
Essays
and
Perspectives
Synthesis
of
the
first
10
years
of
long-term
ecological
research
in
Amazonian
Forest
ecosystem
–
implications
for
conservation
and
management
Fernanda
V.
Costa
a,∗,
Flávia
R.C.
Costa
a,
William
E.
Magnusson
a,
Elizabeth
Franklin
a,
Jansen
Zuanon
a,
Renato
Cintra
a,
Flávio
Luizão
a,
José
Luís
C.
Camargo
a,b,
Ana
Andrade
b,
William
F.
Laurance
b,c,
Fabricio
Baccaro
e,a,
Jorge
Luiz
Pereira
Souza
a,
Helder
Espírito-Santo
d aInstitutoNacionaldePesquisasdaAmazônia(INPA),Manaus,AM,BrazilbBiologicalDynamicsofForestFragmentsProject,InstitutoNacionaldePesquisasdaAmazônia(INPA),Manaus,AM,Brazil
cCentreforTropicalEnvironmentalandSustainabilityScience&CollegeofMarineandEnvironmentalSciences,JamesCookUniversity,
Cairns,Australia
dPostgraduateEcologyProgram,InstitutoNacionaldePesquisasdaAmazônia(INPA),Manaus,AM,Brazil
eUniversidadeFederaldoAmazonas(UFAM),Manaus,Amazonas,Brazil
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received26November2014 Accepted11March2015 Availableonline5May2015
Keywords: Globalchanges Habitatloss Long-termstudies Spatial-temporalvariations Selectivelogging
a
b
s
t
r
a
c
t
Wepresentasynthesisofthefirst10yearsofLongTermEcologicalResearchprojectin AmazonianForest.Weelucidatethenaturaldynamicsofforestecosystemprocessesand associatedbiota,anditschangescausedbydistinctpressuresofselectivetimberextraction andforestfragmentation.Wefoundthat,forbothplantsandanimals,densitiesof individ-ualsanddistributionofspeciesassemblagesarespatiallyheterogeneousatthemesoscale, eveninrelativelyundisturbedforests,andthatassociationswithtopo-edaphicvariables allowpredictionofaconsiderablepartofthisvariation.Forbiologicalgroupswhose dynam-icswerestudiedintheshort-term,levelsofchangeinspeciescompositionanddensities wererelativelyhigh,andthesechangesweregenerallyintunewithspatial environmen-talvariation.Theimpactofselectiveloggingonassemblagesandecosystemprocesseswas normallymoderate,andaround19yearswereneededforrecoveringforestbiomassandtree sizedistribution.Continuedstudiesareneededtodeterminethetimerequiredfor recuper-ationoftimberstocksandpre-loggingfloristiccomposition.Selectiveloggingappearstobe compatiblewiththebiodiversityconservation,butreductionandbetterplanningofroad accessmaybemoreimportantthanplannedloggingintensities.Habitat-loss’impacton organismsandecosystemprocessesislargeandlong-lasting,sinceitinducesthelossof manytaxonomicgroupsandspecies,highertreemortalityandacceleratedforestdynamics. Therewasanegativesynergybetweentheimpactsofhabitatlossandclimaticchanges,and abetterunderstandingoftheseprocessescanonlybeobtainedthroughlong-termresearch. ©2015Associac¸ãoBrasileiradeCiênciaEcológicaeConservac¸ão.PublishedbyElsevier EditoraLtda.Allrightsreserved.
∗ Correspondingauthor.
E-mailaddresses:[email protected],[email protected](F.V.Costa). http://dx.doi.org/10.1016/j.ncon.2015.03.002
Introduction
Growth ofhuman populationsand economicactivities are transformingtropicalforestsinto amosaic ofaltered envi-ronments, pastures and isolated forest fragments, with severeconsequencesforbiodiversity(Bierregaardetal.,1992;
Lauranceetal.,2011).ThedestructionofAmazonianforests
grew exponentially during the 1970sand 1980s (Fearnside,
1987,1990),andcontinuestoimpactdynamicsoffauna,flora
andecosystemprocesses.
Humanactivitiesintropicalforestareasrangefrom selec-tivetimberextractionwithmoderateenvironmentalimpacts to complete deforestation for agro-pastoral and industrial activities (Laurance and Vasconcelos,2009). Understanding howandtowhatextenttropicalforestecosystemsrespond todifferentanthropogenicactivitiesprovidesusanimportant baseforenvironmentalplanning,managementand monitor-ing,allessentialforthepromotionofsustainabledevelopment oftropicalenvironments,suchasAmazonia.
Inresponsetothesechallenges,researchersoftheNational Institute for Amazonian Research (INPA) included Central AmazoniaintheBrazilianprogramforLongTermEcological Research–LTER(hereafterreferencedasLTER-Site#1) estab-lishedbytheBrazilianNationalResearchCouncil(CNPq).The aimoftheprogramistocollectandorganizedataonthe struc-ture and functioning ofthe principal Brazilianecosystems inorder togenerate aknowledge basetoevaluate biologi-caldiversity,andsubsequentlyproposeplansforsustainable useandconservation.TheLTER-Site#1(Fig.1)istheoldest
AmazoniansiteintheBrazilianLTERprogram,andincludes threeforestreservesmanagedbyINPAthatdifferinlanduse intensity.Allsitesarecoveredbyevergreendenseterra-firme forest.
TheDuckeForestReserve(RFD)islocated26kmnorthof Manaus(2◦5547.80S;59◦5830.34W),andcovers10.000ha (10×10km).Thisareadoesnotsufferenvironmentalimpact withinitsborders,butissurroundedbyurbansprawlfrom Manauscity.Allstudieswereconductedinpermanentplots regularly distributed acrossthe landscape. The Experimen-tal ForestManagementStation(ZF2)islocatedabout 90km northeastofManaus(2◦38S,60◦11W),andwassubmittedto selectiveloggingtreatmentsin4-haplots,arrangedinthree blocks.Eachblockhasonecontrolunloggedplot,threeplots loggedin1987,andoneplotloggedin1993toassesstheeffect oftimeaftertimberextraction.Theloggedplotsexperienced differentintensitiesoftimberextraction.Alltheseplotswere establishedintheuplandflatterrain(“plateau”),tominimize topographiceffects.ThereservesoftheBiologicalDynamicsof theForestsFragmentsProject(PDBFF)aresituated80kmfrom Manaus andspanabout1.000km2.Thestudyareaincludes 11 forest fragments (fiveof1ha, four of10ha, and twoof 100ha),andexpansesofnearbycontinuousforestthatserveas experimentalcontrols.Intheearly1980s,thefragmentswere isolated from nearbyintactforestbydistancesof80–650m byclearingandburningthesurroundingforest.Moststudies performedinPDBFFassessedtheeffectoffragmentation com-ponents–sizeoffragmentsandedgeeffect–onbiodiversity, ratherthanevaluatingfragmentationasalandscapeprocess asproposedbyFahrig(2003,2013).
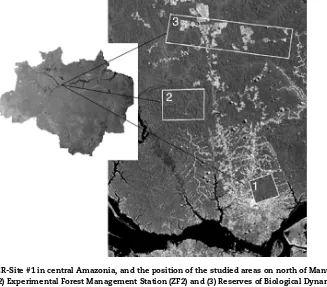
Fig.1–LocationofLTER-Site#1incentralAmazonia,andthepositionofthestudiedareasonnorthofManuscity.(1)Ducke
ForestReserve(RFD),(2)ExperimentalForestManagementStation(ZF2)and(3)ReservesofBiologicalDynamicsofForest
[image:2.612.132.459.425.712.2]Table1–TablesummarizingLTER-Site#1areaswithitsrespectivestudiedgroupsandenvironmentalcharacteristics affectedbyeachpredictorofchangeevaluated.
Area Predictorofchange Studiedgrouporcharacteristicaffected
DUCKE
Soiltexture/topography Plantbiomass,treemortality,wooddecomposition,nutrientcontent Plantcomposition(palms,herbs,shrubs,treesandlianas)
Fungirichnessandabundance(leaflitterfungi)
Animalcomposition–vertebrates(birds,primates,lizards,snakesandamphibians)and soilinvertebrates(ants,pseudoscorpions,mitesandtermites)
Distancefromriparianzone Plant(palms,herbs,shrubs,treesandlianas)andbirdcomposition Vegetationstructure
WaterandWatershed properties
Animalcomposition(birds,snakes,frogs) Wooddecomposition
Compositionofaquaticfauna(fishandinvertebrates) Floristiccomposition
Time Treebiomass,treemortality
Fungirichnessandabundance(leaflitterfungi) Vertebrates(fish)andinvertebrates(pseudoscorpions)
ZF2
Skiddertrails Richnessandcompositionofunderstoryherbs
Loggingintensity Treeregeneration,biomass,treespeciesrichness,volumeoftimberspecies Nutrients
Timeafterlogging PopulationabundancesofVertebrates(lizards,frogs)andinvertebrates(antsand termites)
Treespeciesrichness,treebiomass,deadbiomass,understoryherbs Vertebrates(understorybirds)
Soilproperties/topography Treebiomass
Plantcomposition(understoryandtreespecies,lianasandpalms)
PDBFF
Plantlitter,arborealbiomass,treemortality GlobalClimatechange
Edgeeffects
Climaticattributes(light,desiccation,heatandwinds)
Ecosystemprocess(germination,pollination,dispersionandherbivory) Habitat
loss
Micro-climaticregime,waterflow
Forestdynamics(plantmortality,recruitment,andgrowth,gapformation) CompositionandrichnessofVertebrates(birds,primates,mammals,fishandfrogs) andinvertebrates(buterfliesandaquaticinsects)
Plantcomposition(palms,trees)
Since these areas are similar in terms of experimental designs (samples collected with standardized protocols in permanentnonfloodedevergreen rainforestplots)wewere encouraged tosynthesize the information carriedout dur-ingthelast10yearsofresearchbytheBrazilianLTER-Site#1 (1999–2009).Here,wepresentasynthesisofthemain stud-iesundertakeninthisperiod,coveringrelationshipsbetween ecosystemprocessesandassociatedbiotawithdifferentland uses,suchaschangesinnaturalforestlandscapes,impacts causedbyselectivelogging,andintenseforestfragmentation. Thebiologicalgroupsstudiedandenvironmental character-isticsaffectedbyeachpredictorofchangeevaluatedineach areaaresummarizedinTable1.
Biodiversity
and
ecosystem
processes
vary
within
the
mesoscale
topography
Thetropicalforestischaracterizedbyamplenaturalvariation generatedbydifferences insoilcharacteristics,topography, climateandvegetationstructure.StudiesinRFDatLTER-Site #1,Central Amazonia,have shownthat withinmesoscales (10.000ha), soil and topography gradients are the princi-pal factors responsible for differences between landscape elements.
Thelocaltopography,consistingofplateaus, slopesand stream valleys,clearly influencessoil texture and nutrient
distribution. Soilclaycontent ishighonplateaus and low-lyingareashavesandysoils(Araújoetal.,2002;Luizãoetal., 2004).Thenitrogenconcentration(N)inleaves,plantlitterand theuppersoilstratumwerefoundtobelowerinvalleysthan onslopesandplateaus.Thecarbonconcentration(C)didnot varywithtopography,resultinginhighC/Nratiosinvalley bot-toms.ThelowquantitiesofNcirculatinginvalleysarelikely attributabletosandysoilsandseasonalflooding(Araújoetal.,
2002;Luizãoetal.,2004).
Levels of nitrogen mineralization and nitrification were significantly lower in soils of valleys, with N immobiliza-tionmainlyobservedduringtherainyseason(Luizãoetal., 2004). The production of fine plant litter(maximum stem width≤2cm)anditsdecompositionwereslightlygreaterin plateausascomparedtovalleys(Luizãoetal.,2004).Therefore, athicklayerofleavesaccumulatesoversandysoilsduetothe scarcityofnutrientsfoundinthesesoils,whichthenresults inrelativelyslowerdecompositionrates.Thelocaltopography (plateau,slopeandvalley)clearlyinfluencesthelitterandsoil nutrientdistribution.Thesedifferencesshouldbeconsidered intheconstructionofmodelspredictingcarbonemissionrates thatusedataondecompositionintheirestimates.
[image:3.612.52.552.79.424.2]inclay-richsoilsofplateauswhilepalmbiomassisgreater insandysoilsofslopesandvalleys.InthePDBFFareas,tree biomassincreaseswiththequantityofclayinsoilsaswell assoilfertility,mainlyduetonitrogen(Lauranceetal.,1999). However,thecorrelationbetweenbiomassandnitrogenmay be due to the contribution of leaves to soil nitrogen, and notcausedbysoilnitrogencontent.Sincesoilvariablesare markedlyinfluencedbylocaltopography,thesestudiesshow theimportanceoftakingintoaccounttopographytoimprove ourcurrentestimatesofcarbonstocksoverlargeareasinthe Amazon,anditcanbeeasilyassessedusingsatelliteimagery. Theprocessofwooddecompositionislargelyindependent ofsoilcharacteristics,topography,and vegetationstructure
inRFD (Toledoet al., 2009). The absence of a relationship
betweenwooddecompositionandvariablesinfluencingtree biomassanddecompositionofplantlitterindicatesan asym-metrybetweenproductivityanddecomposition,whichshould intheorybringaboutagreateraccumulationofdeadwood invalleys,whereratesoftreemortalitywerereportedtobe greater(Toledoetal.,2012).AccordingtoToledoetal.(2012), thegreaterproportionoftreedeathduetouprootinginvalleys waslikelytoberelatedtorestrictionsonrootestablishment duetoanoxicsupersaturatedsoils,andthereducedneedfor finerootsbecauseofthegreateravailabilityofphosphorous invalleysoils.Together,thesefactorsfavorlowinvestmentin subterraneanbiomassinvalleys.
Biodiversitydistributionfollowssimilarpatternstothose observedforecosystemprocesses.Studiesdemonstratethat thecompositionofplantsinmanyfunctionalgroupsstudied inRFD(herbs,palms,shrubs,treesandlianas),aswellasfungi, isstronglyassociatedwithedaphicconditions,changing grad-uallyalongthetopographicgradientthatextendsfromsandy, poorandpoorly-drainedsoilsofvalleystomoreclay-richand well-drainedsoilsofplateaus(Costaetal.,2005,2009;Kinupp
andMagnusson,2005;Braga-Netoetal.,2008;Druckeretal.,
2008;Schiettiet al., 2013; Rodrigueset al., 2014).Similarly,
studies from non-fragmented areas of the PDBFF reserves show that topography and soilfertility affect the distribu-tionofunderstoryplants(Harmsetal.,2004),lianas(Laurance et al., 2001), arboreal successional species(Lauranceet al.,
2006a),andthecompositionofthetreecommunity(Bohlman
etal.,2008;Lauranceetal.,2010).
Someanimal groups, especiallythose assumed to have limiteddispersal–suchasamphibians(Meninetal.,2008)and soilinvertebrates(Araújo,2007;Franklinetal.,2007;Oliveira
etal.,2009;Dambros,2010;Souzaetal.,2012)–displayed
dis-tributionstosomedegreeassociatedwithtopographyandsoil characteristics.Thereisevidencethatabundanceand diver-sityofedaphicarthropodsareassociatedwithdifferentlevels ofC,P,Na andCainlitterand/orinsoil.Thispattern sug-geststhatqualityofsubstratemaylimitthesoilmesofauna inDuckeReserve.Approximatelyhalfoftheantspecieswere foundsolelyinvalleysor onlyonplateaus, suggestingthat themajorityare habitatspecialistsand dependofsoil tex-ture,whichisrelatedtomoisturecontent(Oliveiraetal.,2009). Pseudoscorpions,however,werenotfoundtobesensitiveto environmentalfactors,suchasterraintopography, percent-ageofclay insoils, quantityofplant litterorsoilpH, and assuch,canbeconsideredhabitatgeneralists(Aguiaretal.,
2006).
Groupswithamplegeographicdistribution,suchaslizards
(Pinto,2008a)andsnakes(Fragaetal.,2011)werealsofound
tobelittleaffectedbytopographicandsoilvariablespresent inthemesoscalecontextatRFD.Similarly,VidalandCintra
(2006)showedthatthetopographicgradientcharacterizedby
plateaus–slopes–lowlands(60–140mofaltitude)hadlittle effectontheoccurrence,sizeandgroupdensityofthe pri-mateSaguinusbicolor(Sauim-de-coleira,anendangeredspecies). However,owlsandbirdspeciescompositionchangesalongthe topographicgradient,andalsoinrelationtoleaflitterdepth and distance to forest streams. Compositional differences werealsofoundintheaviancommunityintheeasternand westernwaterbasinsthatcomposethereserve.Theseresults suggestthatalthoughmostbirdspeciesoccurthroughoutthe mesoscale,manyspeciestrackdifferencesinthelandscape andforeststructure(BarrosandCintra,2009;CintraandNaka,
2012).
Regarding aquaticfauna ofRFD, Mendonc¸a et al. (2005) showedthatfishspeciescompositionvariesbetweendrainage basins,andthatthis differenceisassociatedwithvariation inwaterchemistry,currentvelocitiesand substrate proper-ties.TheridgethatrunsdownthemiddleofRFDseparates thesetwomajorwaterbasins,withstreamstotheeast flow-ingtotributariesoftheAmazonRiverandthosetothewestto theNegroRiver(Mendonc¸aetal.,2005).Someterrestrialplant groups alsohadspecies restrictedtooneor anotherwater basin(Costaetal.,2005;Garcia,2005;KinuppandMagnusson, 2005).However,itisdifficulttoidentifywhichfactorsaccount forthis difference sincethereislittle variationin soiland other environmentalfactors betweendrainage basins.The onlyimportantdifferencebetweenwatershedsisthe avail-ablephosphoruscontent,whichmaybelinkedtothedrainage patterns.Sincethesewatershedsareseparatedbylowridges, theydonotrepresentbarrierstodispersalforsomegroups
(Costaetal.,2005).
Studieshaveshownthat,whileextremelyinterestingfor somequestions,itisnotgenerallyeconomicallyviabletofocus on exhaustivelists ofmegadiversegroups ofinvertebrates, suchasoribatidmitesandants.Forstudiesdesignedtodetect environmentalchanges,itisbettertoevaluatechangesinthe presence ofmorecommon species (Moraes et al., 2011) or evenuse reduced-samplingtechniques (Souza et al.,2009). For example,not sorting the less frequentspecies didnot severelyaffecttheobservedpatternofsimilarityamong sam-pleplots,indicatingthatmoreabundantspeciescanbeused assubstitutesforlessabundantspeciesinbiomonitoring stud-ies.Inaddition,thesamplingeffortreductiondidnotaffectthe capacitytodetecttheeffectofsoilpHforthecommunityof antgenera(Souzaetal.,2009).
10% of species turnover
Water table level 60
50
40
30
20
10
0
All life forms Trees
Shrubs 90% of species
turnover
Saturated zone
Horizontal distance from nearest drainage (m)
Height abo
v
e
the nearest dr
ainage (m)
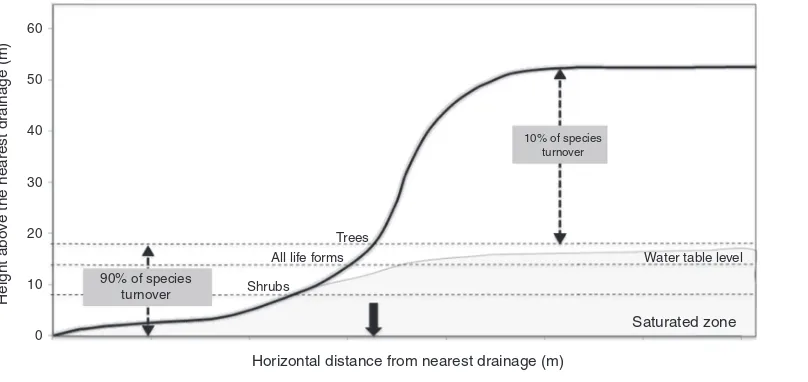
Fig.2–Representationofthepositionswheremostchangesinplantcompositionoccursinrelationtothesmallforest
streams.Watertablefluctuationzoneisshowningray.Theblackarrowhighlightedisthemaximumextensionofthe
riparianarea(350m),givenlimitsdetectedinplantspeciescomposition.
AdaptedfromSchiettietal.(2013).
touselocalmicroclimatesmightbemorerelevantforfuture survivalthantheirgeneralassociationswithclimateatlarge scales(Antonellietal.,2010).
The
importance
of
the
riparian
zone
DiversestudiesinLTER-Site#1haveshownthatspecies com-position ofdistinct assemblages such as plants, frogs and tadpoles,snakesandbirds,variesinrelationtodistancefrom waterbodies(Druckeretal.,2008;Rodriguesetal.,2010;Fraga
etal.,2011;Buenoetal., 2012;Schiettietal., 2013).InRFD,
Druckeretal.(2008)showedthatthefloristiccompositionof
understoryherbsisdistinctfrom thatfoundupto approxi-mately100mhorizontallyfromtheedgeofstreams,orabove about70mabovedrainages,varyingslightlywiththesizeof thestreamvalley.Asaresult,theonlydistincthabitat recog-nizedbymostspeciesappearstoberestrictedtoareasaround streams.Furtheraway,differencesaresubtleandthepattern variesamongspecies.
Schiettietal.(2013)showedthatthefloristiccomposition
ofherbs,shrubs,palms, treesand lianasvary considerably betweenvalleys,butisrelativelyconstantinareasofhigher elevation(Fig.2).Mostchangesinspeciescomposition(about 90%)occur up todistancesbetween 8and 18min vertical drainageheight,dependingonthegroupofplants.This ver-tical interval is equivalent to a distance of approximately 60–250mhorizontally from waterways. Similarly,frogs and theirtadpoles(Rodriguesetal.,2010),understorybirds(Bueno etal.,2012)andsnakes(Fragaetal.,2011)studiedinRFDalso appeartorecognizeariparianzonesimilarinwidth (approxi-mately100–200m)tothatestimatedforunderstoryherbsand theotherplantgroups.
UnderBrazilianlaw,thewidthofprotectedriparian vegeta-tiondependsonthewidthofthewaterbody.Thislegislation mandatesprotectionofareaswithin30mforstreamsupto
10mwide, suchasthose that occurin theDuckeReserve. Our studiesshowed that this riparianarea issmallerthan thatrecognizedbymostanimalandplantgroups,andthat awidthof100–250mwouldbemoreappropriatefor conser-vationpurposes. Thestudies alsoshowed thataminimum verticaldistancefrom theclosestdrainageareawouldbea bettercriteriontoprotectbiodiversitythanprotectionbased onlyonahorizontaldistance.Thecriticalverticaldistance from drainage areas for conservations purposesshould be defined by the zone of groundwater fluctuation, and this willvaryamongAmazonianlandscapesdependingon topo-graphic variation, precipitation regime and soil properties. Thedifferencesinspeciescompositionbetweenriparianareas inadjacent smallwatershedsalsoindicate thateach small watershedhasanovelcontributionintermsofspeciestothe regionaldiversity.Thesefindingssupportourclaimthat nar-rowprotectedareas,asBrazilianlawprescribes,willnotbe sufficienttoprotectthemajorityofspecies.
Thefindingthatareasofhighestcompositionalchangeand mostdistinct fromothers inthe environment(about200m horizontallyoneachsideofstream,and8–18mverticallyfrom the riparianzone)arethose directlyaffected by groundwa-terfluctuationalsohasimplicationsforpredictingtheeffects ofclimate change.If the climatebecomesdrier in Amazo-nia, water storage in the soil and groundwater levels will decrease.Thiswillreducethesizeofriparianareasandcause changesinspeciescompositionsduetodifferencesindrought tolerance(Engelbrechtetal.,2007).Themajorityof tropical-forestspeciesarenotcapableofdispersingoverlongdistances
(Clarketal.,2005;Colwelletal.,2008;Terborgh etal.,2011)
and plantsalreadyconfinedtonarrowzonesneardrainage areaswillhavelittlerecoursetomigratetosimilar environ-ments.Thesespeciesmayalsobecomethreatenedbypressure fromspeciesofhigherareasthatwillbeabletoinvadeareas aroundstreamsasthegroundwaterleveldrops(Schiettietal.,
[image:5.612.100.504.63.254.2]Spatial
and
temporal
variation
in
biodiversity
and
ecosystem
processes
Biodiversityandecosystemprocessesvarynotonlyinspace, butalsointime,andthistemporaldynamiccanalsoberelated toenvironmentalcharacteristics.Understandingthevariation oftemporaldynamicsinthelandscapeisimportantfor mod-elingscenariosforclimatechangeandconservationplanning. Astudy offish assemblies inRFD streams showedthat speciescomposition and fish abundance vary betweenthe dryandwetseason,withrichness(20%)andabundance(30%) being higher in the dry season. However, the composition tendstoreturntoitsoriginalstateattheonsetofthenextdry season,duetothecommonestspeciesthataccountforthe sta-bilityoffishassembliesoverthelongterm.Changesinspecies composition observedbetween seasonsappeartobe asso-ciatedwithlateralmigrationoffishtotemporarypoolsand othermicro-habitatsduringthefloodperiod(Espírito-Santo
etal.,2009,2013).Theseobservationsaresimilartolateralfish
migrationpatterns associatedwith largeAmazonianrivers (Junketal.,1989).
Braga-Netoandcollaborators(2008)foundthatspeciesof
leaf-litterfungirespondtotheinteractionbetweenedaphic conditions,whichvaryinspace,andtoprecipitation,which variesintime.Thenumberoffruitingmorphospeciesvaries alonganedaphicgradientonlyduringthedriestperiods,being lowerinclay-richanddriersoils.Duringtherainiestperiods oftheyear,humidity islesslimiting,resultinginasimilar numberofspeciesfruitingalongtheedaphicgradient.
Comparisonsbetween older studies (Adis and Mahnert,
1990,1993)andmorerecentones(Aguiaretal.,2006)on
pseu-doscorpionsshowedthatthesepredatorshaveaconsistent patternofabundanceanddiversityoverdecades.Thisresult suggeststhat distributionsofspeciesintheseassemblages arestableoverthelongterm,contrastingwiththevariations observedintheshorttermforotherorganisms.
Castilhoet al.(2010)showedthatthe short-termrateof
biomasschangeinRFDwaspredominatelypositiveinplotson clay-richandmorefertilesoils,andpredominatelynegativein plotswithsandyandinfertilesoilsduringthefirstmonitoring period(2001–2003),butnotinthesecond(2002–2004)(Castilho etal.,2010).Therefore,incontrasttoresultsofseveralrecent studies,thatdidnottakeintoaccounttheinteractionbetween climateandsoils,thatstudysuggeststhatlargeareasof Ama-zoniawithsandysoilsmaylosebiomassundertheconditions predictedbymostclimate-changemodels.Thepersistenceof thiseffectthroughtimestillneedstobeverified.
Theeffectsofsoilandtopographyontreemortalityalso vary temporally.Soil and topographyexplainabout 25% of thespatialvariationinsmall-tree(<30cmDBH)mortality dur-ing a period with heavy rain storms, while no effect was detectedduringaperiodoffewerstorms(Toledoetal.,2012). Themagnitudeofthe effectofsoilandtopographyontree mortalityalsoincreasedafterstorms.Ingeneral,plotswith lessfertilesoilsonsteepslopesorsandysoilsinvalleyshad greatertreemortalitythanthoseonwell-drainedclayeysoils ontheplateaus(Toledoetal.,2012).Infact,thehigher propor-tionofuprooteddeadtreesinvalleysinAmazoniahasbeen attributedtopoorsoildrainage,sincewaterexcessinthese
areasrestrictsrootestablishment,productivityoffineroots androotingdepth(EspeletaandClark,2007).
The dynamics of populations, species assemblages and ecosystem processesvary spatiallyon the landscapescale. Mostofthosestudiesreportedthatdynamicsdifferbetween differentlandscapeelements,suchasclayplateausandsandy valleys.Knowledgeofthesevariationsinnaturaldynamics acrossspaceiscrucialtointerpretchangesindynamicsdueto anthropogenicdisturbances.Wedemonstratethatlocal con-trolplotsthatdonotcoverthelandscapevariationwillfailto providethebaselinetowhichperturbationsshouldbe com-pared.
The
effects
of
selective
timber
extraction
on
biodiversity
and
ecosystem
processes
are
moderate
ExperimentaltimberextractioninZF2wasconductedinplots thatunderwentdifferenttimbercuttingtreatments:low(25%), moderate(50%),high(75%)andacontroltreatmentgroupthat correspondstotheprimaryforestswithoutwoodextraction
(HiguchiandBiot,1997).Theintensityofloggingineachplot
corresponded tothe reductioninvolumeofall commercial treesspecieswithDBH≥10cm.
Thestudiesintheareaofselectiveloggingindicatethat mostofthetreebiomasshadrecoveredby19yearsafter cut-ting. However,inthisperiod,commercialspeciesrecovered only66–88%oftheoriginalwoodvolume,suggestingthatmore time is needed forrecuperation ofthe timber stocksafter exploration(Pinto,2008b).Thecompositionoftheforeststill differsmuchfromtheoriginal,withcommercialspecies hav-inglower basalarea.Theextractionoflargetreesmaylead tochangesinthelongterm,becauseestablishmentofmany commercial hardwoodspeciesisslow.Logging significantly increasedrichnessofthetreeregeneratinglayerinrelationto controlplots(Magnussonetal.,1999)andaffectedtheoverall similarityinspeciescompositionamongplots,butthe mag-nitude of the effect was small comparedto the difference betweenonecontrolplotandallotherplots(Magnussonetal.,
1999).
Pinto (2008b) suggested that a cycle of30 years is
fea-sible for the forests to be managed again, if no external or internaldisturbanceoccurs.A compilationofstudiesin the BrazilianAtlanticforest(Liebschetal., 2008)estimated atleastahundredyears torecoverpre-disturbance propor-tions ofanimal-dispersedtreespecies(80%ofthespecies), non-pioneertreespecies(90%)andunderstoryspecies(50%). ConsideringthatthestudyatZF2wasfocusedoncommercial treespeciesandwascarriedoutundercontrolledconditions ofdisturbance,weemphasizethatthis estimateofatleast 30yearsisapplicableonlytocommercialplantsandactions fortimber management.Morestudies onlong termeffects ofloggingshouldbedonetopredictthetimenecessaryfor recuperationoftheinitialtreespeciescomposition.
accumulated vegetative residues that decomposed in the yearsfollowingcutting,contributingwithcarbonand nutri-ents to the soil, as well as CO2 to the atmosphere. The liberationofnutrientsfromcoarseleaflitterandwood decom-positionincreasedtheconcentrationofpotassium,calcium and magnesium available in the soils in the two years followinglogging.Necromassincreased,andthereforethe res-pirationratealsoincreasedinthefirstfiveyearsanduptoat least15yearsaftercutting.Pinto(2008c)comparedthe nutri-entcontentandstocksintreesoftwosecondaryforestsof differentagesandhistoryoflanduse.Treesinthesecondary growthafterlogging(forestof23yearsage)hadhigher nutri-entcontents(N,K,Ca,Mg,Na)thantreesinsecondaryforests developedafterclearcutting(forestof14yearage),showing theimportanceofusehistoryandtimeafterdisturbancefor theecosystemprocesses.
CostaandMagnusson(2002) examinedthe effectof
dif-ferentloggingintensities,timesinceloggingandtheskidder tracksonthecompositionanddiversityofunderstoryherbs. Nooveralldifferenceswerefoundinthecompositionofherbs betweenthedifferentloggingintensities,buttherewerelocal differencesinplantcompositionassociatedwitholdclearings andalong skiddertracks.Thecompositionand richness of herbsdidnotvarywithtimesinceloggingupto10years.Costa
and Magnusson (2002,2003) showed that selective cutting
affectsthereproductionofunderstoryplants,buttheseeffects werenotnecessarilynegative.Theabundanceand number offloweringplantspecieswasgreater 4yearsafterlogging andreturnedtocontrollevelsafter11years.Theintensityof logginghadnoimpactonthereproductionofthe understory-plantassemblages as a whole. However,for somespecies, reproductionwashigherinareaswithgreaterlogging inten-sity. Thesestudies demonstratethat managedforests with similarselectiveloggingintensitiescanbeusedforthe conser-vationofunderstoryherbspecies.Additionally,skiddertrails shouldbeminimizedsincetheycause greatestimpactover thelongtermandpermitcolonizationbyinvasivespecies.
Low-impactselectiveloggingcausedmoderateeffectson bothvertebrateandinvertebrateassemblages.Guilhermeand
Cintra(2001)showedthatthediversity,speciescomposition
andfrequencyofusebyunderstorybirdswerenotaffectedby differentloggingintensities,butwereaffectedbytimeafter logging.Theseauthorsshowedthatselectiveloggingdoesnot causegreatimpactsonbirdassemblages,andthat,insome situations,managedareascouldbeabetteroptionfor conser-vationinrelationtootherformsoflanduse.Incontrast,Lima
etal.(2001)foundthatselectiveloggingaffectedthreelizard
speciesdifferently.Theabundanceofonespeciesincreased withselectiveloggingwhileanotherspeciesdecreased.In con-trasttotheresultsforvertebrates,selectiveloggingdidnot affectthenumberofspeciesortotalabundanceofantsonthe forestground(Vasconcelosetal.,2000).However,differences inloggingintensitycausedchangesinthedensityofvarious speciesofants(Vasconcelosetal.,2000)andleaf-littertermites (Limaetal.,2000),andtheseeffectslastedformorethan10 yearsaftercutting.
Ingeneral,theeffectsofselectiveloggingweremoderate, and no species loss was recorded, even though the abun-danceofsomespeciesincreasedordecreased.Howeveritis importanttonotethat,inthesestudies,selectiveloggingwas
undertakenbyaresearchorganization,didnotexperienced forestfiresorotherdisturbancestypicallyassociatedwith ille-gallogging,anddidnotincludeanevaluationoftheimpacts ofaccessroads.Thisformofselectiveloggingappearstobe compatiblewithbiodiversityconservation,butreductionand betterplanningofaccessroadsmaybemoreimportantthan plannedloggingintensities,aconclusionalsoreachedinan areawithcommerciallogging(Diasetal.,2010).
Forest
fragmentation
strongly
affects
ecosystem
processes
and
associated
biota
Studiesonthecompositionanddynamicsofthetree assem-blagesinfragmentedareasofPDBFFatLTER-Site#1areoneof thelargestintermsofspatialextent(1.000km2)and tempo-ralscale(approximately30years)anywhereintheworld.The impactofforestfragmentationontreeassemblageshasa pro-foundinfluenceonthelong-termpersistenceofanimaland plantpopulationsinfragmentedforests.Themicro-climatic regimeinfragmentsdiffersfromintactareas(Camargoand
Kapos,1995).Pastureoragriculturalareassurrounding
frag-mentshavelowerratesofevapotranspiration,andarehotter and drier. Theeffects ofdryingcan penetrate from 100to 200mintofragments,notnecessarilyfollowingalinear
pat-tern(CamargoandKapos,1995;Malcolm,1998;Didhamand
Lawton,1999).Waterbodiesinfragmentedlandscapespresent
greater temporal variationinflow than dothose in forest, duetothelowerratesofevapotranspiration(Trancoso,2008). Thesedifferencespromotelocalizedfloodinginthewet sea-son and stream failure in the dry season. Together, these climaticandhydrologicalmodificationshavelargeeffectson organisms and ecosystem processes, especially nearforest edges.
Edge effects in fragments are variable. They include increaseindesiccation,heat,light,andturbulencefromwinds amongotherclimatic factorsthatdirectly affectecosystem processes(Lauranceetal.,1997,2004a),suchasseed germi-nation andsurvivalofseedlings(Daviesetal.,1998;Bruna, 1999),pollination,seeddispersion,andherbivory(Harmsand
Dalling,1997;CordeiroandHowe,2001,2003;Terborghetal.,
2001),seedfall(DaviesandSemui, 2006;Nascimentoet al.,
2006),andnutrientcycling(Klein,1989).Fragmentationalso altersforestdynamics,increasingmortalityrates,tree dam-age and clearings formation (Ferreira and Laurance, 1997;
Lauranceetal.,1998a).Oneofthemostimportantimplications
abouttheincreaseintreemortalityisthelossoflivebiomass inthefirst100–300mofedge(Lauranceetal., 1997,1998a). Notonlydomoretreesdieinfragments,butthegreatest pro-portionofthesedeathsisalsoconstitutedbylargetreesthat makeupthelargestfractionofforestbiomass(Lauranceetal., 2000).Smallincreasesinbiomassoflianasandsmalltrees, andcoarseandfineplantlitterdonotcompensatefortheloss ofbiomasscausedbyincreasedtreemortality(Nascimento
and Laurance,2004).Mathematical modelspredictthat the
lossofbiomassincreasesabruptly infragmentswithareas between100and400ha,dependingontheshapeofthe
frag-ment(Lauranceetal.,1997,1998a).Thesemodelsalsosuggest
areascould beresponsibleforanannualemissionofmore than150milliontonsofcarbonintotheatmosphere(Laurance
etal.,1998b).
Areareductioniscommonlyassociatedwiththe impover-ishmentofbiodiversityinforestfragments.Reductionsinthe numberofspecieswithfragmentationwereobservedinalarge numberoftaxonomicgroupsincludingprimates(Gilbertand
Setz,2001;BoyleandSmith,2010),insectivorousunderstory
birds,(StratfordandStouffer,1999;Ferrazetal.,2007),palms
(Scariot,1999),andlargeherbivorousmammals(Timo,2003),
amongothers.However,smallerfragmentsdonotalwayshave fewerspeciesthanlargerfragments(Laurance,2008).Forsome taxonomicgroups,suchasbutterflies(BrownandHutchings, 1997),frogs(Tocheret al.,1997)andleaf-litterinvertebrates
(Didham,1997),speciesnumbersincreasewithfragmentation.
IncentralAmazonia,thenumberoffrogspeciesincreasedas aresultoftheresilience toedgeeffectsbythe majorityof forestspecies,andduetotheinfluxofspeciescomingfrom modifiedhabitatsurroundingfragmentedareas(Tocheretal., 1997).Similarly,thenumberofbutterflyspeciesalsoincreased aftertheisolationoffragmentsduetotheinvasionofspecies typicalofopenareas(BrownandHutchings,1997).Insmall fragments(plotsof1ha),thenumberoftreespeciesdidnot declinewithinforest fragments, dueto the recruitment of speciesfrom initialsuccessional stagesthatnormally have lowfrequencyincontinuousforests.Thisgroupofplantsis favoredbytheprocessofforestfragmentationand prolifer-atesinmany fragmentsduetothe mortalityoflarge trees
(Lauranceetal.,2006b;Nascimentoetal.,2006).The
magni-tudeoffloristicchangesinseedlingsisfasterandmoreintense thanthatobservedforadults(Benitez-Malvido,1998;Sizerand
Tanner,1999),whichmayaffectfuturegenerations.
Overall,the responsesofdifferentspeciesor taxonomic groupsto fragmentation variedconsiderably. Some species werefavoredbyforestfragmentation,suchasspecialistson clearingsformedbynaturaltree falls,which becomemore abundantinfragment borders.On theother hand, species sensitivetotheeffectsoffragmentationgenerallydonot tol-eratehabitatconditionssurroundingfragments(Gasconetal., 1999), astheyare vulnerable toedgeeffectsand incapable ofcrossingdeforestedareas(StoufferandBierregaard,1995;
Lauranceet al.,2004a),orhavelargehomeranges(Rylands
andKeuroghlian,1988;Harper,1989).
The main lessons learned are that small fragments (<100ha)are extremelysensitiveand tend todegradewith timedue tohigh treemortality,but if protectedfrom fires andothermajordisturbances,canbegintorecoverinjusta decadeortwo(Lauranceetal.,2011).Unfortunately,thisisnot theprevailingscenarioinBrazilianecosystems.Inthisway, wereinforcetheimportanceofthelong-termstudies, includ-ingareasisolatedformanydecades,sincefragmentationhas long-lastingeffectsonbiodiversity.
Anthropogenic
disturbances
and
global
changes
interact
synergistically
on
ecosystem
processes
Severalhypothesesontheeffectsofglobalclimatechangeon forestbiomassandspeciessubstitutionarebeingtested(Lewis
etal.,2004a).Itisstillunclearifincreasesintemperature,CO2
concentrations,nitrogendepositionorchangesinintensityof elNinoeventsarethemostimportantdriversandwhatisthe sizeoftheirimpacts.
Laurance and Williamson (2001) showed that tree
mor-talityand the decayof plantlitterfrom treesunder stress duringperiodsofextremedroughtinEl-Ni ˜noyearsincrease dramaticallyinalreadydisturbedareasoffragmentborders, ascomparedtoareasinsidefragments.Theaccumulationof combustiblematerialandthegreaterpenetrationoflightin borderscouldbethesourceofforestfiresindrieryears, espe-ciallyunderthescenariospredictedbymostclimate-change models.
ArborealbiomassincreasedovertimeinCentralAmazonia, perhapsasaresultofdirectclimatechangesorthe fertiliza-tionoftheatmospherewithCO2(Castilhoetal.,2010).Biomass change averaged +5.6Mgha−1ano−1, a level in accordance withstudiesinotherAmazonianareas.Biomasschangewas foundtobenegativelyassociatedwithprecipitation, suggest-ing thatmortalityratesincreaseinyearswithlargestorms andaffectstandingcarbonstocks(Castilhoetal.,2010). Simi-larresultsoveramuchlargerareabasedonplotsthatinclude those fromPDBFFatLTER-Site#1were obtainedbyPhillips
etal.(1998,2004),Bakeretal.(2004),Lewisetal.(2004b)and
Lauranceetal.(2004b),andthosestudiesindicatedthatareas
ofintactforestscouldbesignificantcarbonsinks.
Incontrast,physiologicalmodelsdevelopedbyChambers
et al.(2004)suggestthatupland (terrafirme)forestsarenot
efficientintheiruseofcarbon.Basedontheirmodels, car-bonemissionsviatreerespirationandfromorganicmaterial shouldbegreaterthanthephotosyntheticabsorptionbytrees. Thisfactsuggeststhattheforestgrowthprobablydoesnot respondtotheincreaseinatmosphericCO2concentration.If thisisthecase,theforestwouldnotbebehavingasacarbon sink.Thesedifferencesbetweenmodelsandresults regard-ingvegetationcomponentshowtheimportanceoflong-term investigationtotheinterpretationofresultsfromshort-term studiesandphysiologicalmodels.
Conclusions
Theresultsobtainedinthefirst10yearsofAmazonianLTER (1999–2009)showedthatdensitiesofindividualsandspecies assemblagesdistributionsarespatiallyheterogeneous,even inrelativelyundisturbedforests,andthatassociationswith topo-edaphicvariablesallowpredictionofaconsiderablepart ofthisvariation.For thebiologicalgroupswhosedynamics were studiedintheshort-term, levelsofchangeinspecies composition and density were relatively high, and in gen-eral changed in tune with the environmental variation in space.Therefore,inclusionofenvironmentalpredictorssuch astopography,soilcontentandwaterattributesallows devel-opmentofmoreprecisemodelsoffuturedynamics.
recoveryofpre-loggingassemblages.Incontrast,theimpactof forestfragmentationonorganismsandecosystemprocesses isgreatandlong-lasting.Studiesindicatethatthereisnegative synergybetweentheimpactsoffragmentationandclimatic changes,andthatabetterunderstandingoftheseprocesses canonlybeobtainedthroughlong-termresearch.
TheresultsoftheresearchinLTER-Site#1 indicatethat conservationdecisionsregardingtheAmazonforestarenot goingtobeeasy.Interventions,suchasselectivelogging,are likelytoaffectdifferentgroups,suchascommerciallyvaluable treesandinvertebrates,differently.Fragmentationhas imme-diate obvious effects onbiodiversity, but even inrelatively undisturbedareas,subtledifferencesinlandscapeelements makelargeareassuitableorunsuitableforsometaxa,which mean that it will often be necessary to undertake in situ studies to effectively plan reserve boundaries, at least for mesoscalereserves.Possiblesourcesoffundingforreserves, suchascarbontrading,arebasedonstaticmodelsofbiomass distribution, but theinteractions betweenedaphicfeatures and climatecurrentlymakepredictionsimprecise,and the insitumeasurementsarenotinaccordwiththeoretical mod-els.Systematicconservationplanningwillonlybeeffectiveif wecangeneratethedatanecessaryformodels,andthe expe-riencefromLTER-Site#1indicatesthatwearefarfromhaving adequateunderstandingofthelong-termprocesses.
Conflicts
of
interest
Theauthorsdeclarenoconflictsofinterest.
Acknowledgments
Weare gratefultofinancialandlogisticsupportgrantedby LongTermEcologicalResearchProgram(ProgramadePesquisa Ecológicade LongaDurac¸ão, PELD)through CNPq, the Pro-gramforBiodiversityResearch(PPBio)throughMinistériode Ciência,Tecnologia eInivac¸ão(MCTI), Coordinationforthe ImprovementofHigherEducationPersonnel(CAPES), Biologi-calDynamicsoftheForestsFragmentsProject(PDBFF-ST669) andSmithsonianTropicalResearchInstitute(STRI).CNPQand CAPESprovidedmanyscholarshipstothestudentsinvolved. Wealsothankalltheresearchers,students,fieldandlab assis-tantsinvolvedinthestudies,whoaretoonumeroustoname here.
r
e
f
e
r
e
n
c
e
s
Adis,J.,Mahnert,V.,1990.Verticaldistributionandabundanceof
Pseudoscorpionspecies(Arachnida)inthesoilofa
Neotropicalsecondaryforestduringthedryandtherainy
season.ActaZool.190,11–16.
Adis,J.,Mahnert,V.,1993.Verticaldistributionandabundanceof
Pseudoscorpions(Arachnida)inthesoiloftwodifferent
neotropicalprimaryforestsduringthedryandrainyseasons.
Mem.Qld.Mus.33,431–440.
Aguiar,N.,Gualberto,T.L.,Franklin,E.,2006.Amedium-spatial
scaledistributionpatternofPseudoscorpionida(Arachnida)in
agradientoftopography(altitudeandinclination),soil
factors,litterincentralAmazonianforestreserve,Brazil.Braz.
J.Biol.66,791–802.
Antonelli,A.,etal.,2010.Molecularstudiesandphylogeography
ofAmazoniantetrapodsandtheirrelationtogeologicaland
climaticmodels.In:Hoorn,C.,Wesselingh,F.(Eds.),
Amazonia,LandscapeandSpeciesEvolution:ALookIntothe
Past.BlackwellPublishing,pp.386–404.
Araújo,J.S.,(M.Sc.thesis)2007.Métodosdeamostragem,
influênciadosfatoresambientaiseguiadeidentificac¸ãodos
escorpiões(Chelicerata,Scorpiones)daReservaDucke,
Manaus,Amazonas,Brasil.NationalInstituteforAmazonian
Research(INPA),Manaus,Brazil,pp.82.
Araújo,A.C.,etal.,2002.Comparativemeasurementsofcarbon
dioxidefluxesfromtwonearbytowersinacentralAmazonian
rainforest:theManausLBAsite.J.Geophys.Res.107,8090.
Baker,T.R.,etal.,2004.IncreasingbiomassinAmazonianforest
plots.R.Soc.Lond.359,353–365.
Barros,O.C.,Cintra,R.,2009.Theeffectsofforeststructureon
occurrenceandabundanceonthreeowlspecies(Aves:
Strigidae)inthecentralAmazonForest.Zoologia26,85–96.
Benitez-Malvido,J.,1998.Impactofforestfragmentationon
seedlingabundanceinatropicalrainforest.Conserv.Biol.12,
380–389.
Bierregaard,R.O.,etal.,1992.Thebiologicaldynamicsoftropical
rainforestfragments.Bioscience42,859–866.
Bohlman,S.A.,etal.,2008.Importanceofsoils,topographyand
geographicdistanceinstructuringcentralAmazoniantree
communities.J.Veg.Sci.19,863–874.
Boyle,S.A.,Smith,A.T.,2010.Behavioralmodificationsin
northernbeardedsakimonkeys(Chiropotessatanas
chiropotes)inforestfragmentsofcentralAmazonia.Primates
51,43–51.
Braga-Neto,etal.,2008.LeaflitterfungiinaCentralAmazonian
forest:theinfluenceofrainfall,soilandtopographyonthe
distributionoffruitingbodies.Biodivers.Conserv.17,
2701–2712.
Brown,K.S.,Hutchings,R.W.,1997.Disturbance,fragmentation,
andthedynamicsofdiversityinAmazonianforestbutterflies.
In:Laurance,W.F.,Bierregaard,R.O.(Eds.),TropicalForest
Remnants:Ecology,Management,andConservationof
FragmentedCommunities.UniversityofChicagoPress,
Chicago,pp.91–110.
Bruna,E.M.,1999.Seedgerminationinrainforestfragments.
Nature402,139.
Bueno,A.S.,etal.,2012.Thewidthofriparianhabitatsfor
understorybirdsinanAmazonianforest.Ecol.Appl.22,
722–734.
Camargo,J.L.C.,Kapos,V.,1995.Complexedgeeffectsonsoil
moistureandmicroclimateincentralAmazonianforest.J.
Trop.Ecol.11,205–221.
Castilho,C.V.,etal.,2006.Variationinabovegroundtreelive
biomassinacentralAmazonianforest:effectsofsoiland
topography.For.Ecol.Manag.234,85–96.
Castilho,C.V.,etal.,2010.Short-termtemporalchangesintree
livebiomassinaCentralAmazonianForest,Brazil.Biotropica
42,95–103.
Cintra,R.,Naka,L.N.,2012.Spatialvariationinbirdcommunity
compositioninrelationtotopographicgradientandforest
heterogeneityinacentralAmazonianrainforest.Int.J.Ecol.
2012,ID435671,25p.
Chambers,J.Q.,etal.,2004.Responseoftreebiomassandwood
littertodisturbanceinaCentralAmazonForest.Oecologia
141,596–614.
Clark,C.J.,Poulsen,J.R.,Bolker,B.M.,Connor,E.F.,Parker,V.T., 2005.Comparativeseedshadowsofbird-,monkey-,and
wind-dispersedtrees.Ecology86,2684–2694.
Cordeiro,N.J.,Howe,H.F.,2001.Lowrecruitmentoftrees
dispersedbyanimalsinAfricanForestfragments.Conserv.
Cordeiro,N.J.,Howe,H.F.,2003.Forestfragmentationsevers
mutualismbetweenseeddispersersandanendemicAfrican
tree.Proc.Natl.Acad.Sci.100,14052–14056.
Costa,F.R.C.,Magnusson,W.E.,2002.Selectiveloggingeffectson
abundance,diversity,andcompositionoftropicalunderstory
herbs.Ecol.Appl.12,807–819.
Costa,F.R.C.,Magnusson,W.E.,2003.Effectsofselectivelogging
onthediversityandabundanceoffloweringandfruiting
understoryplantsinaCentralAmazonianforest.Biotropica
35,103–114.
Costa,F.R.C.,Magnusson,W.E.,Luizão,R.C.,2005.Mesoscale
distributionpatternsofAmazonianunderstoreyherbsin
relationtotopography,soilandwatersheds.J.Ecol.93,
863–878.
Costa,F.R.C.,etal.,2009.Gradientswithingradients:the
mesoscaledistributionpatternsofpalmsinacentral
Amazonianforest.J.Veg.Sci.20,69–78.
Colwell,R.K.,etal.,2008.Globalwarming,elevationalrange
shifts,andlowlandbioticattritioninthewettropics.Science
322,258–261.
Dambros,C.S.,(M.Sc.thesis)2010.Efeitodoambientena
composic¸ãodeespéciesdecupinsesuficiênciaamostralem
umaflorestaprimáriadeterra-firmenaAmazôniacentral.
NationalInstituteforAmazonianResearch(INPA),Manaus,
Brazil.
Davies,S.J.,etal.,1998.Comparativeecologyof11sympatric
speciesofMacarangainBorneo:treedistributioninrelationto
horizontalandverticalresourceheterogeneity.J.Ecol.86,
662–673.
Davies,S.J.,Semui,H.,2006.Competitivedominanceina
secondarysuccessionalrainforestcommunityinBorneo.J.
Trop.Ecol.22,53–64,24.
Dias,M.S.,Magnusson,W.E.,Zuanon,J.,2010.Effectsof
reduced-impactloggingonfishassemblagesincentral
Amazonia.Conserv.Biol.24,278–286.
Didham,R.K.,1997.Theinfluenceofedgeeffectsandforest
fragmentationonleaf-litterinvertebratesincentral
Amazonia.In:Laurance,W.F.,Bierregaard,R.O.(Eds.),Tropical
ForestRemnants:Ecology,Management,andConservationof
FragmentedCommunities.UniversityofChicagoPress,
Chicago,pp.55–70.
Didham,R.K.,Lawton,J.H.,1999.Edgestructuredeterminesthe
magnitudeofchangesinmicroclimateandvegetation
structureintropicalforestfragments.Biotropica31,
17–30.
Drucker,D.P.,Costa,F.R.C.,Magnusson,W.E.,2008.Howwideis
theriparianzoneofsmallstreamsintropicalforests?Atest
withterrestrialherbs.J.Trop.Ecol.24,65–74.
Engelbrecht,B.M.J.,etal.,2007.Droughtsensitivityshapesspecies
distributionpatternsintropicalforests.Nature447,80–82.
Espeleta,J.F.,Clark,D.A.,2007.Multi-scalevariationinfine-root
biomassinatropicalrainforest:aseven-yearstudy.Ecol.
Monogr.77,377–404.
Espírito-Santo,H.M.V.,etal.,2009.Seasonalvariationinthe
compositionoffishassemblagesinsmallAmazonianforest
streams:evidenceforpredictablechanges.Freshw.Biol.54,
536–548.
Espírito-Santo,H.M.V.,Rodríguez,M.A.,Zuanon,J.,2013. ReproductivestrategiesofAmazonianstreamfishesandtheir fine-scaleuseofhabitatareorderedalongahydrological gradient.Freshw.Biol.58,2494–2504,
http://dx.doi.org/10.1111/fwb.12225.
Fahrig,L.,2003.Effectsofhabitatfragmentationonbiodiversity.
Annu.Rev.Ecol.Evol.Syst.34,487–515.
Fahrig,L.,2013.Rethinkingpatchsizeandisolationeffects:the
habitatamountHypothesis.J.Biogeogr.40,1649–1663.
Fearnside,P.M.,1987.CausesofdeforestationintheBrazilian
Amazon.In:Dickinson,R.F.(Ed.),TheGeophysiologyof
Amazonia:VegetationandClimateInteractions.JohnWiley,
SanFrancisco,pp.37–61.
Fearnside,P.M.,1990.DeforestationinBrazilianAmazonia.In:
Woodwell,G.M.(Ed.),TheEarthinTransition:Patternsand
ProcessesofBioticImpoverishment.CambridgeUniversity
Press,NewYork,pp.211–238.
Ferraz,G.,etal.,2007.Alarge-scaledeforestationexperiment:
effectsofpatchareaandisolationonAmazonbirds.Science
315,238–241.
Ferreira,L.V.,Laurance,W.F.,1997.Effectsofforestfragmentation
onmortalityanddamageofselectedtreesincentral
Amazonia.Conserv.Biol.11,797–801.
Fraga,R.,Lima,A.P.,Magnusson,W.E.,2011.Mesoscalespatial
ecologyofatropicalsnakeassemblage:thewidthofriparian
corridorsincentralAmazonia.Herpetol.J.21,51–57.
Franklin,E.,Santos,E.M.R.,Albuquerque,M.I.C.,2007.Edaphic
andarboricolousoribatidmites(Acari:Oribatida)intropical
environments:changesinthedistributionofhigher
taxonomiclevelinthecommunityofspecies.Braz.J.Biol.67,
631–637.
Garcia,A.R.M.,(M.Sc.thesis)2005.Distribuic¸ãoeabundânciade
arbustosdafamíliaPiperaceaenaReservaFlorestalAdolpho
Ducke,Manaus,Amazonas–Brasil.NationalInstitutefor
AmazonianResearch(INPA),Manaus,Brazil.
Gascon,C.,etal.,1999.Matrixhabitatandspeciesrichnessin
tropicalforestremnants.Biol.Conserv.91,223–230.
Gilbert,K.A.,Setz,E.Z.F.,2001.Primatesinafragmented
landscape:sixspeciesincentralAmazonia.In:Laurance,W.F.,
Bierregaard,R.O.(Eds.),TropicalForestRemnants:Ecology,
ManagementandConservationofFragmentedCommunities.
UniversityofChicagoPress,Chicago,pp.207–221.
Guilherme,E.,Cintra,R.,2001.Effectsofintensityandageof
selectiveloggingandtreegirdlingonanunderstorybird
communitycompositioninCentralAmazonia,Brazil.
Ecotropica7,77–92.
Harms,K.E.,Dalling,J.W.,1997.Damageandherbivorytolerance
throughresproutingasanadvantageoflargeseedsizein
tropicaltreesandlianas.J.Trop.Ecol.13,617–621.
Harms,K.E.,Powers,J.,Montgomery,R.,2004.Variationinsapling
density,understorycoverandresourceavailabilityinfour
Neotropicalforests.Biotropica36,40–51.
Harper,L.H.,1989.Thepersistenceofant-followingbirdsinsmall
Amazonianforestfragments.ActaAmaz.19,249–263.
Higuchi,N.,Biot,Y.,1997.ProjetoBionte:biomassaenutrientes
florestais.FinalReport.Manaus-AM:INPA/DFID.,pp.345.
Junk,W.J.,Bayley,J.B.,Sparks,R.E.,1989.Thefloodpulseconcept
inriver-floodplainsystems.In:Dodge,D.P.(Ed.),Proceedings
oftheInternationalLargeRiverSymposium.CanadianSpecial
PublicationsonFisheriesandAquacultureScience,Ottawa,
ON,pp.110–127.
Klein,B.C.,1989.Effectsofforestfragmentationondungand
carrionbeetlecommunitiesinCentralAmazonia.Ecology70,
1715–1725.
Kinupp,V.F.,Magnusson,W.E.,2005.Spatialpatternsinthe
understoreyshrubgenusPsychotriaincentralAmazonia:
effectsofdistanceandtopography.J.Trop.Ecol.21,363–374.
Laurance,W.F.,etal.,1997.BiomasscollapseinAmazonianForest
fragments.Science278,1117–1118.
Laurance,W.F.,etal.,1998a.Effectsofforestfragmentationon
recruitmentpatternsincentralAmazonia.Conserv.Biol.12,
460–464.
Laurance,W.F.,etal.,1998b.Biomasslossinforestfragments.
Science282,1610–1611.
Laurance,W.F.,etal.,1999.Relationshipbetweensoilsand
Amazonforestbiomass:alandscape-scalestudy.For.Ecol.
Manag.118,127–138.
Laurance,W.F.,etal.,2000.Rainforestfragmentationkillsbig
Laurance,W.F.,etal.,2001.Rainforestfragmentationandthe
structureofAmazonianlianacommunities.Ecology82,
105–116.
Laurance,W.F.,Williamson,G.B.,2001.Positivefeedbacksamong
forestfragmentation,drought,andclimatechangeinthe
Amazon.Conserv.Biol.15,1529–1535.
Laurance,W.F.,etal.,2004a.InferredlongevityofAmazonian
rainforesttreesbasedonalong-termdemographicstudy.For.
Ecol.Manag.190,131–143.
Laurance,W.F.,etal.,2004b.Pervasivealterationoftree
communitiesinundisturbedAmazonianforests.Nature428,
171–175.
Laurance,W.F.,etal.,2006a.Rainforestfragmentationandthe
proliferationofsuccessionaltrees.Ecology87,469–482.
Laurance,W.F.,etal.,2006b.Rapiddecayoftree-community
compositioninAmazonianforestfragments.Proc.Natl.Acad.
Sci.U.S.A.103,19010–19014.
Laurance,W.F.,2008.Theorymeetsreality:howhabitat
fragmentationresearchhastranscendedislandbiogeographic
theory.Biol.Conserv.141,1731–1744.
Laurance,W.F.,Vasconcelos,H.L.,2009.ConseqüênciasEcológicas
daFragmentac¸ãoFlorestalnaAmazônia.Oecol.Bras.13,
434–451.
Laurance,S.G.,Andrade,A.,Laurance,W.F.,2010.Unanticipated
effectsofstanddynamismonAmazoniantreediversity.
Biotropica42,429–434.
Laurance,W.F.,etal.,2011.ThefateofAmazonianforest
fragments:a32-yearinvestigation.Biol.Conserv.144,56–67.
Lewis,S.L.,Malhi,Y.,Phillips,O.L.,2004a.Fingerprintingthe
impactsofglobalchangeontropicalforests.R.Soc.Lond.359,
437–462.
Lewis,S.L.,etal.,2004b.Concertedchangesintropicalforest
structureanddynamics:evidencefrom50SouthAmerican
long-termplots.R.Soc.Lond.359,421–436.
Liebsch,D.,Marques,M.C.M.,Goldenberg,R.,2008.Howlongdoes
theAtlanticRainForesttaketorecoverafteradisturbance?
Changesinspeciescompositionandecologicalfeatures
duringsecondarysuccession.Biol.Conserv.141,1717–1725.
Lima,A.P.,etal.,2000.Effectofselectiveloggingintensityontwo
termitespeciesofthegenusSyntermesinCentralAmazonia.
For.Ecol.Manag.137,151–154.
Lima,A.P.,Suárez,F.I.O.,Higuchi,N.,2001.Theeffectsofselective
loggingonthelizardsKentropyxcalcarata,Ameivaameivaand
Mabuyanigropunctata.J.Amphib.–Reptil.22,209–216. Luizão,R.C.C.,etal.,2004.Variationofcarbonandnitrogen
cyclingprocessesalongatopographicgradientinacentral
Amazonianforest.GlobalChangeBiol.10,592–600.
Magnusson,W.E.,etal.,1999.Loggingactivityandtree
regenerationinanAmazonianforest.For.Ecol.Manag.113,
67–74.
Malcolm,J.R.,1998.Amodelofconductiveheatflowinforest
edgesandfragmentedlandscapes.Clim.Change39,487–502.
Mendonc¸a,F.P.,Magnusson,W.E.,Zuanon,J.,2005.Relationships
betweenhabitatcharacteristicsandfishassemblagesinsmall
streamsofCentralAmazonia.Copeia4,751–764.
Menin,M.,Waldez,F.,Lima,A.P.,2008.Temporalvariationinthe
abundanceandnumberofspeciesoffrogsin10,000haofa
forestincentralAmazonia,Brazil.S.Am.J.Herpetol.3,68–81.
Moraes,J.,etal.,2011.Speciesdiversityofedaphicmites(Acari:
Oribatida)andeffectsofthetopography,soilpropertiesand
littergradientsontheirqualitativeandquantitative
compositionin64km2offorestinAmazonia.Exp.Appl.
Acarol.55,39–63.
Nascimento,H.,Laurance,W.F.,2004.Biomassdynamicsin
Amazonianforestfragments.Ecol.Appl.14,S127–S138.
Nascimento,H.,etal.,2006.Effectsofthesurroundingmatrixon
treerecruitmentinAmazonianforestfragments.Conserv.
Biol.20,853–860.
Oliveira,P.Y.,etal.,2009.Antspeciesdistributionalonga
topographicgradientina“terra-firme”forestreservein
CentralAmazonia.Pesqui.Agropecu.Bras.44,852–860.
Phillips,O.L.,etal.,1998.Changesinthecarbonbalanceof
tropicalforest:evidencefromlong-termplots.Science282,
439–442.
Phillips,O.L.,etal.,2004.PatternandprocessinAmazontree
turnover.R.Soc.Lond.359,381–407.
Pinto,M.G.M.,(Ph.D.thesis)2008a.Diversidadebeta,métodosde
amostragemeinfluênciadefatoresambientaissobreuma
comunidadedelagartosnaAmazôniaCentral.National
InstituteforAmazonianResearch(INPA),Manaus,Brazil,pp.
91.
Pinto,A.C.M.,(Ph.D.thesis)2008b.Dinâmicadeumaflorestade
terrafirmemanejadaexperimentalmentenaregiãode
Manaus(AM).NationalInstituteforAmazonianResearch
(INPA),Manaus,Brazil,pp.91.
Pinto,F.R.,(M.Sc.thesis)2008c.Estimativasdosestoquesde
biomassaenutrientesemflorestassecundáriasnaAmazônia
Central.NationalInstituteforAmazonianResearch(INPA),
Manaus,Brazil,pp.161.
Rodrigues,D.,etal.,2010.Temporarypondavailabilityand
tadpolespeciescompositionincentralAmazonia.
Herpetologica66,124–130.
Rodrigues,L.F.,etal.,2014.Influencesofforeststructureand
landscapefeaturesonspatialvariationinspecies
compositioninapalmcommunityincentralAmazonia.J.
Trop.Ecol.30,565–578.
Rylands,A.B.,Keuroghlian,A.,1988.Primatepopulationsin
continuousforestandforestfragmentsincentralAmazonia.
ActaAmaz.18,291–307.
Scariot,A.,1999.Forestfragmentationeffectsondiversityofthe
palmcommunityincentralAmazonia.J.Ecol.87,66–76.
Schietti,J.,etal.,2013.Verticaldistancefromdrainagedrives
floristiccompositionchangesinanAmazonianterra-firme
rainforest.PlantEcol.Divers.7,1:13.
Sizer,N.,Tanner,E.V.J.,1999.Responsesofwoodyplantseedlings
toedgeformationinalowlandtropicalrainforest,Amazonia.
Biol.Conserv.91,135–142.
Stouffer,P.C.,Bierregaard,R.O.,1995.Effectsofforest
fragmentationonunderstoryhummingbirdsinAmazonian
Brazil.Conserv.Biol.9,1085–1094.
Souza,J.L.P.,etal.,2009.Efficiencyininventoriesofantsina
forestreserveinCentralAmazonia.Pesqui.Agropecu.Bras.
44,940–948.
Souza,J.L.P.,etal.,2012.Trade-offsbetweencomplementarityand
redundancyintheuseofdifferentsamplingtechniquesfor
ground-dwellingantassemblages.Appl.SoilEcol.56,63–73.
Stratford,J.A.,Stouffer,P.C.,1999.Localextinctionsofterrestrial
insectivorousbirdsinafragmentedlandscapenearManaus,
Brazil.Conserv.Biol.13,1416–1423.
Terborgh,J.,etal.,2001.Ecologicalmeltdowninpredator-free
forestfragments.Science294,1923–1925.
Terborgh,J.,etal.,2011.Decomposingdispersallimitation:limits
onfecundityorseeddistribution?J.Ecol.99,935–944.
Timo,T.P.C.,(M.Sc.thesis)2003.Influênciadefragmentac¸ãoe
matrizsobreacomunidadedemamíferosdemédioegrande
porteemumaflorestadeterrafirmenaAmazôniacentral.
NationalInstituteforAmazonianResearch(INPA),Manaus,
Brazil.
Toledo,J.J.,Magnusson,W.E.,Castilho,C.V.,2009.Influenceofsoil,
topographyandsubstratesondifferencesinwood
decompositionbetweenone-hectareplotsinlowlandtropical
moistforestinCentralAmazonia.J.Trop.Ecol.25,649–656.
Toledo,J.J.,etal.,2012.TreemodeofdeathinCentralAmazonia:
effectsofsoilandtopographyontreemortalityassociated
withstormdisturbances.For.Ecol.Manag.263,
Tocher,M.,Gascon,C.,Zimmerman,B.L.,1997.Fragmentation
effectsonaCentralAmazonianfrogcommunity:aten-year
study.In:Laurance,W.F.,Bierregaard,R.O.(Eds.),Tropical
ForestRemnants:Ecology,Management,andConservationof
FragmentedCommunities.UniversityofChicagoPress,
Chicago,pp.124–137.
Trancoso,(M.Sc.thesis)2008.HydrologicalImpactsof
DeforestationinSmallCatchmentsinBrazilianAmazonia.
NationalInstituteforAmazonianResearch(INPA),Manaus,
Brazil.
Vasconcelos,H.L.,Vilhena,J.M.S.,Caliri,G.J.A.,2000.Responsesof
antstoselectiveloggingofacentralAmazonianforest.J.
Appl.Ecol.37,508–514.
Vidal,M.D.,Cintra,R.,2006.Effectsofforeststructure
componentsontheoccurrence,groupsizeanddensityof
groupsofbare-facetamarin(Saguinusbicolor–Primates:
Callitrichinae)inCentralAmazonia.ActaAmaz.36,