Baculovirus
Per Os
Infectivity Factor Complex: Components
and Assembly
Xi Wang,
a,bYu Shang,
aCheng Chen,
a,bShurui Liu,
a,bMeng Chang,
a,bNan Zhang,
a,bHengrui Hu,
a,bFenghua Zhang,
aTao Zhang,
aZhiying Wang,
aXijia Liu,
aZhe Lin,
cFei Deng,
aHualin Wang,
aZhen Zou,
cJust M. Vlak,
dManli Wang,
aZhihong Hu
aaState Key Laboratory of Virology, Wuhan Institute of Virology, Chinese Academy of Sciences, Wuhan, China
bUniversity of Chinese Academy of Sciences, Beijing, China
cState Key Laboratory of Integrated Management of Pest Insects and Rodents, Institute of Zoology, Chinese Academy of Sciences, Beijing, China
dLaboratory of Virology, Wageningen University and Research, Wageningen, the Netherlands
ABSTRACT
Baculovirus entry into insect midgut cells is dependent on a multiprotein
complex of
per os
infectivity factors (PIFs) on the envelopes of occlusion-derived virions
(ODVs). The structure and assembly of the PIF complex are largely unknown. To reveal
the complete members of the complex, a combination of blue native polyacrylamide gel
electrophoresis, liquid chromatography-tandem mass spectrometry, and Western
blot-ting was conducted on three different baculoviruses. The results showed that the PIF
complex has a molecular mass of
⬃
500 kDa and consists of nine PIFs, including a newly
discovered member (PIF9). To decipher the assembly process, each
pif
gene was knocked
out from the
Autographa californica
multicapsid nucleopolyhedrovirus (AcMNPV)
ge-nome individually by use of synthetic baculovirus technology, and the impact on PIF
complex formation was investigated. Deletion of
pif8
resulted in the formation of an
⬃
400-kDa subcomplex. Deletion of
pif0
, -
4
, -
6
, -
7
, or -
9
resulted in a subcomplex of
⬃
230 kDa, but deletion of
pif1
, -
2
, or -
3
abolished formation of any complex. Taken
to-gether, our data identified a core complex of
⬃
230 kDa, consisting of PIF1, -2, and -3.
This revised the previous knowledge that the core complex was about 170 kDa and
con-tained PIF1 to -4. Analysis of the PIF complex in cellular fractions suggested that it is
as-sembled in the cytoplasm before being transported to the nucleus and subsequently
in-corporated into the envelopes of ODVs. Only the full complex, not the subcomplex, is
resistant to proteolytic attack, indicating the essentiality of correct complex assembly for
oral infection.
IMPORTANCE
Entry of baculovirus into host insects is mediated by a
per os
infectivity
factor (PIF) complex on the envelopes of occlusion-derived viruses (ODVs). Knowledge of
the composition and structure of the PIF complex is fundamental to understanding its
mode of action. By using multiple approaches, we determined the complete list of
pro-teins (nine) in the PIF complex. In contrast to previous knowledge in the field, the core
complex is revised to
⬃
230 kDa and consists of PIF1 to -3 but not PIF4. Interestingly, our
results suggest that the PIF complex is formed in the cytoplasm prior to its transport to
the nucleus and subsequent incorporation into ODVs. Only the full complex is resistant
to proteolytic degradation in the insect midgut, implying the critical role of the entire
complex. These findings provide the baseline for future studies on the ODV entry
mech-anism mediated by the multiprotein complex.
KEYWORDS
baculovirus, PIF complex, PIF9, entry,
per os
infectivity factor
B
aculoviruses are large, rod-shaped double-stranded DNA (dsDNA) viruses that
infect insects from the families Lepidoptera, Hymenoptera, and Diptera. The
Bacu-loviridae
family contains four genera:
Alphabaculovirus
,
Betabaculovirus
,
Gammabacu-CitationWang X, Shang Y, Chen C, Liu S,
Chang M, Zhang N, Hu H, Zhang F, Zhang T, Wang Z, Liu X, Lin Z, Deng F, Wang H, Zou Z, Vlak JM, Wang M, Hu Z. 2019. Baculovirusper os infectivity factor complex: components and assembly. J Virol 93:e02053-18.https://doi.org/ 10.1128/JVI.02053-18.
EditorRozanne M. Sandri-Goldin, University of
California, Irvine
Copyright© 2019 American Society for
Microbiology.All Rights Reserved.
Address correspondence to Manli Wang, [email protected], or Zhihong Hu, [email protected].
Received16 November 2018
Accepted17 December 2018
Accepted manuscript posted online2
January 2019
Published
crossm
5 March 2019
on November 6, 2019 by guest
http://jvi.asm.org/
lovirus
, and
Deltabaculovirus
(1).
Autographa californica
multicapsid
nucleopolyhedro-virus (AcMNPV) is the type member of the alphabaculonucleopolyhedro-viruses and the most
well-studied baculovirus (2). There are two types of progeny viruses produced during a
typical baculovirus life cycle, namely, the budded virus (BV) and the occlusion-derived
virus (ODV). ODVs are embedded in occlusion bodies (OBs) and are responsible for oral
infection within the insect midgut, while BVs cause systemic infection in other larval
tissues.
In nature, baculovirus infection begins when OBs are ingested and dissolved in the
highly alkaline and protease-rich midgut. The released ODV particles pass through the
peritrophic membrane lining the gut and initiate infection in midgut epithelial cells.
Successful oral infection depends on a group of viral proteins, called
per os
infectivity
factors (PIFs), on the ODV envelope (3, 4). PIF0 (originally called P74 because the
molecular weight is
⬃
74 kDa) was the first PIF found to be essential for oral infection.
The deletion of PIF0 has no impact on infectious BV production but totally abolishes
oral infectivity of ODVs (5). Eight additional PIFs were identified later and were named
PIF1 (Ac119;
⬃
60 kDa), PIF2 (Ac22;
⬃
44 kDa), PIF3 (Ac115;
⬃
23 kDa), PIF4 (Ac96;
⬃
20 kDa), PIF5 (ODV-E56 or Ac148;
⬃
41 kDa), PIF6 (Ac68;
⬃
16 kDa), PIF7 (Ac110;
⬃
7 kDa), and PIF8 (Ac83;
⬃
96 kDa) (6–13).
All the PIF proteins are expressed in the late stage of virus infection, and they are
envelope proteins of ODVs, which are assembled in the nuclei of the infected cells.
Many PIFs contain the inner nuclear membrane sorting motif (INM-SM), which is
believed to guide the synthesized polypeptides into the nucleus (14). Another
common feature of PIFs is that all their genes are conserved in
Baculoviridae
.
Homologs of
pif
genes are also present in a wide range of invertebrate large dsDNA
viruses, such as white spot syndrome virus (family
Nimaviridae
) (15), salivary gland
hypertrophy virus (family
Hytrosaviridae
) (16),
Apis mellifera
filamentous virus (17),
nudivirus (family
Nudiviridae
) (18), and bracovirus (family
Polydnaviridae
) (19).
A stable PIF complex of
⬎
170 kDa was first identified which could withstand 2%
SDS–5%

-mercaptoethanol and heating at 50°C for 5 min. When this partial
denatur-ation method was applied to
pif
-knockout viruses, it was demonstrated that the
complex contained PIF1, -2, and -3 as a stable core, while PIF0 was loosely associated
with the core (20). Later on, by use of blue native PAGE (BN-PAGE), a preferred method
for isolating multiprotein complexes in a native state (21), the PIF complex was found
to be
⬃
480 kDa and to contain PIF0, PIF1 to -4, and PIF8, while a stable core complex
of
⬃
170 kDa, containing PIF1, -2, -3, and -4, was revealed by the partial denaturation
method (22). A proteomic study showed that PIF6, Ac5, and Ac108 were associated with
PIF1 (22). Later, PIF6 was confirmed to be a component of the PIF complex (23), while
Ac5 was not (24). Recently, PIF7 was also identified as a component of the complex (3).
However, it remains unknown if the current list of complex members is complete. All
PIFs except PIF5 are associated with the PIF complex, which highlights the pivotal role
of the complex for oral baculovirus infection (25). The intact complex was required to
protect PIFs from proteolytic degradation by OB endogenous proteinases during ODV
release from OBs (23). Understanding the structure and function of this unique complex
calls for a detailed study, particularly in light of the ubiquitous nature of
pif
genes in a
wide range of invertebrate large dsDNA viruses.
The mechanism of PIF complex assembly is still largely unknown. Study of the core
complex should help us to understand the structure and formation of the entire
complex. However, so far, the published information on the core complex has not been
consistent, especially regarding the role of PIF4. While PIF1, -2, and -3 are essential
for core complex formation, deletion of
pif4
did not completely impair the stable core
complex but resulted in a smaller stable complex of
⬃
150 kDa (22). This
⬃
150-kDa
complex was later found to be sensitive to proteolytic degradation, and the apparent
inconsistency is due to the fact that the partial denaturation method used did not
completely inhibit proteolytic degradation by the OB’s endogenous proteinase (23).
In this study, we systematically investigated the composition, assembly, and
func-tion of the baculovirus PIF complex to understand its role in virus entry. Multiple
on November 6, 2019 by guest
http://jvi.asm.org/
approaches, including BN-PAGE, liquid chromatography-tandem mass spectrometry
(LC-MS/MS), and Western blotting, were used to obtain a complete list of components
of the PIF complex. The impact of each component on the formation of the AcMNPV
complex under natural conditions was then analyzed by BN-PAGE. We also addressed
the initial intracellular assembly of the complex. Our results provide a likely complete
list of PIF complex components, revise the previous understanding of the core complex,
and shed light on the process of complex assembly and intracellular transport.
RESULTS
Disclosure of components of the PIF complex.
To date, all the known PIFs have
been identified individually, but it is still not clear if there are hitherto unidentified
members of the PIF complex. To reveal all components of the PIF complex, we chose
three baculoviruses, AcMNPV,
Bombyx mori
NPV (BmNPV), and
Helicoverpa armigera
NPV (HearNPV), and analyzed the composition of their PIF complexes systematically. An
integrative investigation by BN-PAGE, LC-MS/MS, and Western blot analysis was
con-ducted (Fig. 1). Specific antibodies were raised against individual PIFs of AcMNPV and
HearNPV to signal their presence in their native PIF complexes. Because PIF4, -6, and -7
failed to yield effective antibodies, AcMNPV recombinants with hemagglutinin
(HA)-tagged PIF4, -6, and -7 were generated using bacmid technology (Fig. 2A), and anti-HA
antibody was used instead.
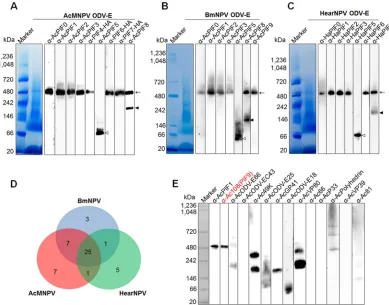
FIG 1Identification of the components of the baculovirus PIF complex. (A to C) Identification of PIF complexes in AcMNPV (A), BmNPV (B), and HearNPV (C). The envelope fractions of wt AcMNPV, BmNPV, and HearNPV ODVs were subjected to BN-PAGE and analyzed by Western blotting with the indicated antibodies against PIFs. The⬃500-kDa PIF complexes are indicated by arrows. PIF8 dimers are indicated by closed arrowheads and PIF5 by open arrowheads. (D) Summary of the LC-MS/MS data on the PIF complexes. The numbers of identified proteins are shown. (E) Verification of other proteins in the AcMNPV PIF complex. BN-PAGE and Western analysis were conducted using the indicated antibodies, and anti-AcPIF1 was used as a positive control for identification of the PIF complex. Except for Ac108, none of the other proteins were identified as components of the PIF complex. ODV envelope proteins derived from appropriate OB concentrations (2⫻107OBs for detection of PIF5 and PIF8 and
1⫻108OBs for the other PIFs) were loaded into each lane for Western blot analysis. For staining, proteins from 3⫻109OBs
were loaded.
on November 6, 2019 by guest
http://jvi.asm.org/
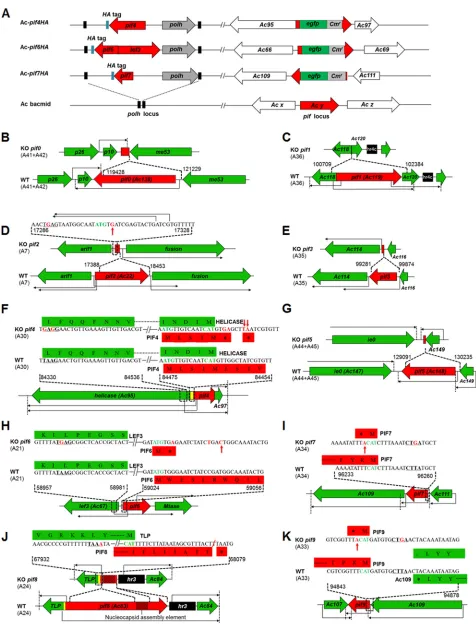
[image:3.585.44.433.71.376.2]FIG 2Construction of recombinant AcMNPVs. (A) Schematic diagrams for construction of recombinant AcMNPVs with HA-tagged PIF4, -6, and -7 by use of the Bac-to-Bac system. (B to K) Strategies for knocking out 10pifgenes. The A fragments containingpifgenes and their modification strategies are shown. Transcriptional start sites and orientations of genes are indicated by long arrows; translational start codons (ATG or complementary CAT) are shown in green, the mutated or inserted nucleotides are shown in red, and translation was abolished by incorporating a stop codon as indicated.
on November 6, 2019 by guest
http://jvi.asm.org/
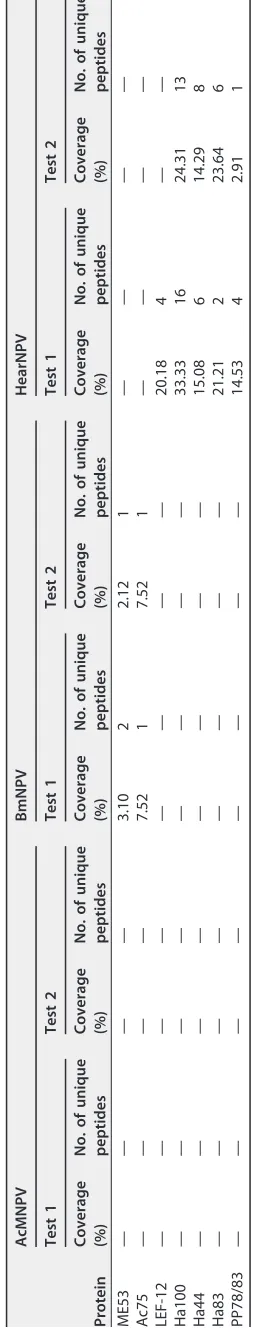
[image:4.585.40.516.66.690.2]TABLE 1 Viral proteins identified in the PIF complex band of AcMNPV, BmNPV, or HearNPV by LC-MS/MS a Protein AcMNPV BmNPV HearNPV Test 1 Test 2 Test 1 Test 2 Test 1 Test 2 Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides PIF0 47.75 30 47.91 30 55.81 37 55.81 37 38.08 24 40.26 29 PIF1 64.91 27 57.92 24 63.57 31 63.95 31 42.99 18 57.20 20 PIF2 56.02 23 56.02 23 55.26 21 66.72 23 59.53 20 56.02 23 PIF3 48.04 7 48.04 7 50.98 8 50.95 8 31.16 4 31.16 7 PIF4 9.25 1 25.43 2 21.43 3 21.43 3 28.90 6 28.90 6 PIF5 55.05 13 44.62 12 46.13 15 45.33 15 27.4 11 49.44 11 PIF6 36.46 7 34.38 5 55.22 7 55.22 7 54.89 8 58.65 7 PIF7 37.50 1 37.50 1 30.51 1 30.51 1 36.21 2 37.93 3 PIF8 63.99 39 65.76 42 52.21 34 53.87 32 53.55 34 60.17 38 Ac108 (PIF9) 49.52 4 49.52 4 49.52 4 49.52 4 53.19 4 51.06 4 ODV-E66 54.69 23 59.8 22 16.10 8 5.13 2 48.07 28 63.84 32 ODV-EC43 77.18 28 77.18 26 70.59 28 69.57 25 56.51 17 56.51 19 49K 69.60 35 74.18 34 68.27 37 66.39 31 61.97 31 57.48 27 ODV-E25 64.47 14 55.70 9 49.12 10 49.12 10 47.39 12 55.22 10 GP41 69.93 23 69.93 21 57.32 21 58.56 20 64.6 14 50.93 17 ODV-E18 16.13 1 16.13 1 33.66 2 33.66 2 45.68 3 51.85 3 VP80 60.93 45 59.77 45 57.95 50 58.81 48 29.09 16 20.50 12 Ac66 33.79 23 38.99 26 35.03 29 38.14 28 58.09 43 57.32 44 P33 32.82 10 40.15 10 27.03 8 22.78 8 42.52 13 49.21 18 Polyhedrin 37.96 9 43.67 11 46.12 10 57.14 12 48.78 13 46.75 14 VP39 19.31 5 25.65 5 10 3 10.29 3 12.29 3 15.02 9 Ac81 40.34 11 40.35 11 31.62 8 31.62 8 36.93 8 26.56 8 Viral ubiquitin 28.57 2 7.79 1 68.83 5 49.35 3 18.07 2 18.07 1 P24 50.51 9 57.58 9 14.87 3 57.58 9 14.92 3 3.63 2 Ac26 21.71 3 21.71 3 15.50 2 9.3 1 15.04 2 — — Ac73 69.70 5 43.35 11 18.18 2 18.18 2 — — — — Ac114 55.90 21 49.76 21 53.54 19 49.06 18 — — — — Ac78 33.03 3 33.03 3 — — — — 17.27 2 22.73 4 P45 29.72 11 26.36 11 29.46 11 26.15 5 — — — — Ac74 36.23 8 34.34 8 — — — — — — — — Protein tyrosine phosphatase 41.67 5 50.00 8 15.48 3 — — — — — — 35K 11.37 4 13.04 4 15.72 5 11.71 4 — — — — Ac5 44.04 4 67.89 6 45.87 4 45.87 4 — — — — Ac82 8.89 2 4.44 1 — — — — — — — — ssDNA binding protein 9.81 3 14.24 4 — — — — — — — — Ac145 36.36 2 27.27 1 22.10 1 26.32 1 — — — — Ac60 16.09 2 8.05 1 — — — — — — — — GP16 14.15 2 7.55 1 — — — — — — — — GP64 5.86 3 13.87 5 — — — — — — — — LEF-3 7.53 4 10.13 4 — — — — — — — — Ac91 — — — — 10.39 1 10.39 1 — — — — Ac51 — — — — 8.78 3 — — 31.44 5 27.84 4 (Continued on next page)
on November 6, 2019 by guest
http://jvi.asm.org/
TABLE 1 (Continued) Protein AcMNPV BmNPV HearNPV Test 1 Test 2 Test 1 Test 2 Test 1 Test 2 Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides Coverage (%) No. of unique peptides ME53 — — — — 3.10 2 2.12 1 — — — — Ac75 — — — — 7.52 1 7.52 1 — — — — LEF-12 — — — — — — — — 20.18 4 — — Ha100 — — — — — — — — 33.33 16 24.31 13 Ha44 — — — — — — — — 15.08 6 14.29 8 Ha83 — — — — — — — — 21.21 2 23.64 6 PP78/83 — — — — — — — — 14.53 4 2.91 1 a—, the protein was not identified in the sample by LC-MS/MS.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:6.585.140.267.75.746.2]BN-PAGE and Western blot analysis confirmed that the AcMNPV PIF complex was
approximately 500 kDa (Fig. 1A), as reported previously (3, 22, 23). By use of
anti-AcMNPV PIF antibodies, a similar,
⬃
500-kDa complex was detected in the closely
related virus BmNPV (Fig. 1B). Moreover, antiserum raised against HearNPV PIF proteins
confirmed the presence of a similar complex in the virus (Fig. 1C). These data suggest
that an
⬃
500-kDa PIF complex in the envelope of ODVs is likely to be conserved in
baculoviruses.
Consistent with previous reports (3, 22, 23), the AcMNPV PIF complex contained
eight PIFs (PIF0 to -4 and PIF6 to -8) and lacked PIF5 (Fig. 1A). In addition to being in
the PIF complex, PIF8 is also present in the ODV envelope, as an approximately 220-kDa
protein (Fig. 1A). The predicted size of PIF8 is
⬃
96 kDa, but it was detected as a 110-kDa
protein via SDS-PAGE (13); we speculate that the
⬃
220-kDa protein may be a dimer of
PIF8. It is worth noting that in the envelopes of BmNPV and HearNPV, PIF5 was also
absent from the complexes, and a putative dimer of PIF8 was detected (Fig. 1B and C).
To identify other possible components, proteomic analyses were carried out on the
⬃
500-kDa PIF complexes from AcMNPV, BmNPV, and HearNPV. LC-MS/MS analyses
identified 25 viral proteins shared by all three viruses, including the 9 PIFs and 16 other
proteins (Fig. 1D; Table 1). Among these proteins, ubiquitin, P24, and Ac26 were
detected in insignificant amounts (Table 1) and were previously deemed to not be
involved in oral infection (2), and they were therefore excluded from further analysis.
Antibodies against the other 13 proteins were raised, but only anti-Ac108 recognized
the PIF complex (Fig. 1E), and anti-PIF1 was used as a positive control in the experiment.
Ac108, an 11.8-kDa protein encoded by
ac108
, is a homolog of
Spodoptera frugiperda
MNPV (SfMNPV) Sf58 (26) and BmNPV Bm91 (27). By using anti-Ac108 antiserum, we
detected the presence of its homolog in the BmNPV PIF complex (Fig. 1B). An
ac108
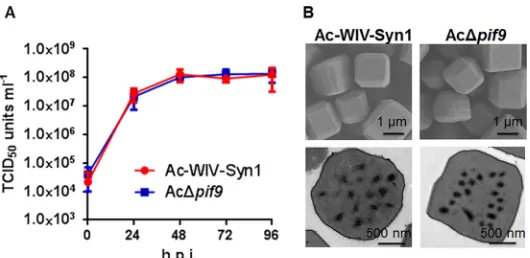
-knockout mutant (Fig. 2K) had no effect on infectious BV production (Fig. 3A) or OB
morphogenesis (Fig. 3B), but the knockout virus lost its oral infectivity to
Spodoptera
exigua
larvae (Table 2). Therefore,
ac108
was proven to encode an authentic PIF, which
was designated PIF9. None of the other 12 remaining viral proteins were detected in
the AcMNPV PIF complex (Fig. 1E), even though they were detected in the ODV
envelope by SDS-PAGE and Western blot analysis (data not shown). This suggests that
there are no other viral proteins in the complex. In summary, the
⬃
500-kDa PIF
complex of AcMNPV consisted of nine PIF proteins: PIF0 to -4 and PIF6 to -9. As the
accumulated size of the nine PIFs (
⬃
360 kDa) is smaller than the observed size of
500 kDa, we further studied the contribution of each individual component to the
formation of the entire complex.
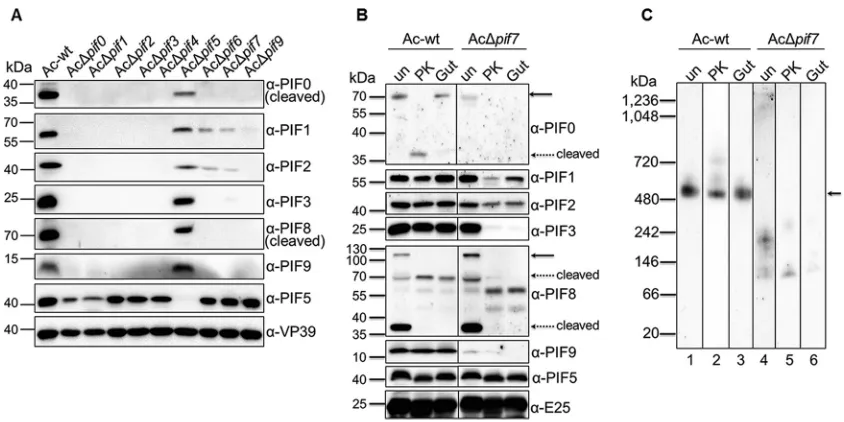
Contributions of individual PIFs to complex formation.
Previous reports showed
that deletion of certain PIFs led to the loss of an intact PIF complex (13, 20, 22, 23, 28).
FIG 3Deletion ofac108(pif9) had no effects on infectious BV production and OB morphogenesis. (A) One-step growth curves for Ac-WIV-Syn1 and AcΔpif9. Sf9 cells were infected with Ac-WIV-Syn1 and AcΔpif9at an MOI of 5 TCID50units/cell, cell supernatants were harvested at the indicated time points,and BV titers were measured by endpoint dilution assay. Data points are mean values of titers derived from three independent infections. Error bars represent the standard deviations. (B) Electron microscopy of OBs of Ac-WIV-Syn1 and AcΔpif9. Upper and lower panels represent the results of SEM and TEM, respectively.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:7.585.74.339.70.199.2]Because those results were interpreted from a partially denatured complex by
SDS-PAGE, which has a relatively low resolution, we decided to systematically explore the
contributions of all nine PIFs to complex formation by BN-PAGE and Western blot
analyses. We used deletion or point mutation strategies to inactivate
pif
genes from the
AcMNPV genome (Fig. 2B to K); the resulting recombinants are all referred to as
knockout mutants in this report. Individual
pif
-knockout AcMNPV mutants were
gen-erated by synthetic baculovirus technology (29) (see Fig. 4A for an example of a
synthesis flowchart [for AcΔ
pif1
]; details of the knockout strategy are shown in Fig. 2B
to K). The resulting viruses were authenticated by sequencing and the absence of an
immune response to PIF-specific antibodies by SDS-PAGE (Fig. 4B). None of the deletion
mutants affected BV production (data not shown), but as expected, they all lost oral
infectivity (Table 2).
We then analyzed the impacts of these
pif
-knockout mutants on PIF complex
formation (Fig. 5). As expected, knockout of
pif5
(Fig. 5A) had no effect on the integrity
of the
⬃
500-kDa PIF complex. The PIF8 dimer was also detected and was associated
with PIF9. On the other hand,
pif8
knockout (Fig. 5B) led to a smaller complex, of
[image:8.585.41.370.83.227.2]approximately 400 kDa, containing at least PIF1 to -3 and PIF9 (PIF4, -6, and -7 were not
TABLE 2Feeding assay of OBs (3⫻109OBs ml⫺1) on 4th-instarS. exigualarvae
Virus
Mortality (no. of dead larvae/total no. of larvae) Test 1 Test 2
Mock 0/24 0/24
Ac-wt 24/24 24/24
AcΔpif0 0/24 1/24
AcΔpif1 0/24 0/23
AcΔpif2 1/24 0/23
AcΔpif3 0/24 0/24
AcΔpif4 0/23 0/21
AcΔpif5 0/23 0/21
AcΔpif6 0/23 0/24
AcΔpif7 2/24 1/24
AcΔpif8 0/23 0/24
AcΔpif9 1/20 0/22
FIG 4Generation and verification ofpif-knockout AcMNPVs. (A) Schematic illustrating the generation of
pif1-knockout AcMNPV. Fragment A36, which containspif1, was modified by overlapping PCR to knock outpif1. After three rounds of transformation-associated recombination (TAR) in yeast cells, the synthetic genome of AcΔpif1was obtained. The detailed strategies for knocking out thepifgenes are shown in Fig. 2B to K. (B) Western blots of ODVs for identification of synthesizedpif-knockout AcMNPVs. The envelope protein ODV-E25 and the nucleocapsid protein VP39 were used as protein loading controls. In order to inactivate endogenous protease in larva-derived OBs, they were heated at 80°C for 30 min prior to ODV purification.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:8.585.67.347.483.660.2]tested due to a lack of antibodies). When either
pif1
,
pif2
, or
pif3
was deleted, no
complex was detected, except for the PIF8 dimer and its associated PIF9 (Fig. 5C to E).
Knockout of
pif0
,
pif4
,
pif6
,
pif7
, or
pif9
(Fig. 5F to J) resulted in an
⬃
230-kDa complex
consisting of PIF1, -2, and -3. The PIF8 dimer was detected in these mutants, and PIF9
was found to be associated with the PIF8 dimer in the case of
pif0
,
pif4
, and
pif6
deletions (Fig. 5F to H). The PIF8 dimer was slightly smaller than the
⬃
230-kDa
complex, and this was more obvious when phosphate-buffered saline (PBS) was used,
as represented by Fig. 5K. We assume that under PBS conditions the conformation of
the complexes may be more natural and therefore results in better resolution by
BN-PAGE. The picture that emerges from these experiments indicates a core complex
of
⬃
230 kDa, consisting of PIF1 to -3, and a subcomplex of about 400 kDa when
pif8
is
deleted.
Assembly of the PIF complex in the cytoplasm.
When the PIF complex was
com-promised by deletion of one of the PIFs, the other PIFs were still found in the ODV
envelope (Fig. 4B). The question is whether the PIF complex is assembled in the nucleus
or initially in the cytoplasm. To address this issue, cells were infected with AcMNPV and
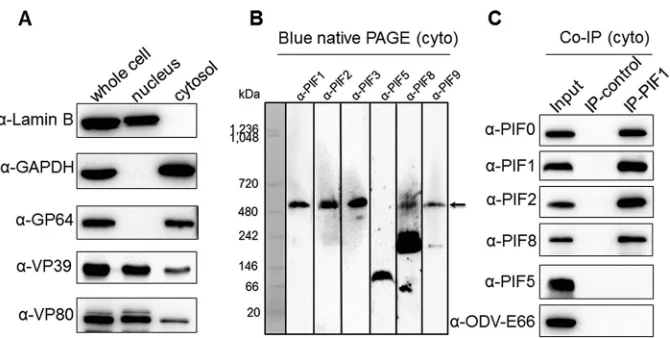
subjected to nuclear and cytoplasmic fractionation. Detection of the cytoplasmic
protein marker glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (absent in the
nucleus) and the nuclear marker lamin B (absent in the cytoplasm) showed that the
fractionation was successful (Fig. 6A). The major BV envelope fusion protein GP64
(cytoplasmic) and two viral nucleocapsid proteins, VP39 and VP80 (nucleocytoplasmic),
were also detected, further confirming the successful virus infection and cell
fraction-ation (Fig. 6A). The cytoplasmic fractions were subjected to BN-PAGE and Western
analysis (Fig. 6B). Anti-PIF1, -PIF2, -PIF3, -PIF8, and -PIF9 antibodies indicated that the
⬃
500-kDa complex was formed in the cytoplasm (Fig. 6B). It is interesting that the PIF8
dimer was already associated with PIF9 in the cytoplasm. Next, a
coimmunoprecipita-tion (Co-IP) experiment was further conducted with anti-PIF1 to confirm the existence
FIG 5Contributions of individual PIFs to PIF complex formation. (A to J) ODVs of AcΔpif0to AcΔpif9were purified in the presence of SBTI, and the ODV pellets were resuspended with ddH2O and then subjected to envelope protein extraction. BN-PAGE and Western blotting were performed on the AcΔpif0to AcΔpif9
ODV envelope fractions, using the indicated antibodies for probing. (K) BN-PAGE and Western blotting were conducted under the same conditions as those for panels A to J, except that the purified ODVs were resuspended in PBS (instead of ddH2O). The PIF8 dimer was found to be smaller than the core complex.
AcΔpif0is shown as an example here, but similar results were obtained with AcΔpif4, AcΔpif6, AcΔpif7, and AcΔpif9.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:9.585.45.539.71.341.2]of a PIF complex in the cytoplasm. The results showed that the components of the PIF
complex (PIF0, PIF1, PIF2, and PIF8), but not unrelated ODV envelope proteins (PIF5 and
ODV-E66), were immunoprecipitated by anti-PIF1 antibody as confirmed by Western
blot analysis (Fig. 6C). These results showed that the PIF complex is formed in the
cytoplasm, possibly prior to transport into the nucleus.
The full complex, but not the subcomplex, is resistant to proteolytic
degrada-tion.
The alkaline environment in the insect midgut induces the dissolution of OBs to
release occluded ODVs. Since proteolytic enzymes are ubiquitous in the insect midgut
(30), ODVs need to resist proteolytic digestion prior to entry into epithelial columnar
cells. It was reported previously that the PIF complex is important in protecting PIF
proteins from degradation by OB endogenous proteinase (23). We further investigated
the resistance of PIF proteins to proteolysis in the different forms of the complex.
Individual
pif
deletions (except for
pif5
deletion) led to the rapid degradation of
other PIF proteins during the process of ODV purification (Fig. 7A). In fact, we had to
add proteinase inhibitors to obtain sufficient protein amounts for experiments with PIF
deletion mutants (Fig. 5). The results suggest that only the full complex provides
sufficient protection from proteolytic degradation for PIF proteins in the larval midgut
during oral infection.
To further verify this hypothesis, ODV envelope proteins of wild-type AcMNPV
(Ac-wt) and AcΔ
pif7
(as an example of
pif
knockouts) were purified in the presence of
soybean trypsin inhibitor (SBTI) to avoid degradation by OB endogenous proteases. The
envelope fractions of ODVs were extracted and exposed to proteinase K or
S. exigua
gut
fluid. In the presence of proteinase K (40 ng/ml) or gut fluid (0.2% [vol/vol]), PIF proteins
from Ac-wt were more resistant to proteolysis than those from AcΔ
pif7
(Fig. 7B).
BN-PAGE and Western blot analyses showed that the intact,
⬃
500-kDa PIF complex of
Ac-wt could still be detected with anti-PIF1 antibody under different treatment
condi-tions; however, in the case of AcΔ
pif7
, the subcomplex of
⬃
230 kDa was barely
detectable after digestion (Fig. 7C). These results suggest that the full PIF complex is
resistant to proteolytic digestion and remains intact prior to attachment to and entry
into columnar epithelial cells.
DISCUSSION
Knowledge of the composition and structure of the PIF complex is fundamental to
understanding its mode of action. Since PIFs were previously identified individually, we
FIG 6The PIF complex is assembled in the cytoplasm of infected cells. (A) Cytoplasmic and nuclear fractionation. Sf9 cells were infected with wild-type AcMNPV at an MOI of 5 TCID50units/cell, and cells wereharvested at 72 h p.i. and separated into nuclear and cytoplasmic fractions. The purification of the fractions was analyzed by SDS-PAGE followed by Western blotting with the indicated antibodies. Lamin B and GAPDH were used as a cellular nucleus marker and a cytoplasmic marker, respectively. (B and C) PIF complexes could be detected in the cytoplasmic fraction. The cytoplasmic fractions from panel A were subjected to blue native PAGE (B) or used for Co-IP assay with mouse PIF1 antibody (C) and probed with the indicated antibodies. The complex in the BN-PAGE gel is indicated with an arrow.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:10.585.43.378.68.237.2]decided to take an integrated investigation to disclose all the possible components of
the PIF complex. To this end, three different baculoviruses were chosen. AcMNPV and
BmNPV both belong to the group I alphabaculoviruses, while HearNPV is a member of
the group II alphabaculoviruses. Our results showed that PIF complexes of all three
baculovirus are
⬃
500 kDa (Fig. 1), indicating that the size and composition of the PIF
complex are conserved in members of the genus
Alphabaculovirus
, and possibly in all
baculoviruses. LC-MS/MS analysis of these three PIF complexes revealed 25 shared
proteins as potential candidates for complex components (Table 1). Western blot
analysis confirmed the previous finding that PIF0 to -4 and PIF6 to -8 are constituents
of the complex, and Ac108 was revealed as a novel component.
Ac108 is a homolog of SfMNPV Sf58, whose deletion abolished oral infectivity of
SfMNPV in
S. frugiperda
(26). Previously, Ac108 was found to be associated with PIF1 by
LC-MS/MS (22). Our results showed that it exists in the PIF complexes of both AcMNPV
and BmNPV (Fig. 1B and E). Its deletion had no effects on BV production and OB
morphogenesis (Fig. 3) but resulted in disassembly of the full PIF complex (Fig. 5J) and
the loss of oral infectivity of AcMNPV in
S. exigua
(Table 2). Therefore, it is a genuine PIF
and is designated PIF9. It is important to note that the previously identified nine PIFs
(PIF0 to -8) are conserved in all sequenced baculoviruses, to date. PIF9, however, is
conserved only in alpha-, beta-, and gammabaculoviruses and may thus be considered
an auxiliary PIF in the complex and essential in some cases. The PIF9 homolog of
BmNPV, Bm91, is dispensable for oral infectivity (27), which also alludes to an auxiliary
function of this PIF in BmNPV ODV entry.
Because other viral proteins identified by LC-MS/MS were not found to be associated
with the
⬃
500-kDa complex (Fig. 1E), it is convincing that the nine PIFs (PIF0 to -4 and
PIF6 to -9) constitute all of the PIF components of the PIF complex. The documentation
of the full members of the PIF complex is important for further investigations of its
structure, stoichiometry, and assembly.
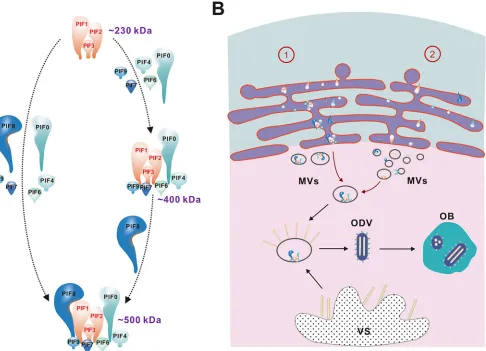
We next analyzed the impacts of individual PIFs on complex formation to gain
insight into the structure of the complex. Notably, deletion of any
pif
gene other than
pif5
eliminated the production of the full 500-kDa complex (Fig. 5). In detail, deletion
of
pif8
yielded an
⬃
400-kDa complex, suggesting that PIF8 is present as a monomer
FIG 7The full complex, but not the subcomplex, confers PIF stability. (A) OBs of Ac-wt orpif-deletion AcMNPVs were left untreated prior to ODV purification. Western blots show the remaining PIFs in ODVs. AcΔpif8was not included in the figure due to the presence of fewer ODVs, but it showed degradation patterns similar to those of the otherpifdeletion mutants. (B) Western blots of Ac-wt and AcΔpif7ODVs purified in the presence of SBTI and exposed to proteinase K (PK; 40 ng/ml) or gut lumen fluid (0.2% [vol/vol]). Full-sized PIF0 and PIF8 are indicated with solid arrows, while the cleaved proteins are indicated with dashed arrows. (C) The intact,⬃500-kDa PIF complex (indicated by an arrow) of Ac-wt was still detected by use of PIF1 antibodies during BN-PAGE analysis after digestion as described for panel B, while the subcomplex of AcΔpif7was barely detectable after proteinase K or gut lumen fluid treatment.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:11.585.40.462.69.280.2](
⬃
110 kDa) in the complex and that its absence does not affect the assembly of other
components. Deletion of
pif0
, -
4
, -
6
, -
7
, or -
9
reduced the complex to
⬃
230 kDa.
Deletion of
pif1
, -
2
, or -
3
abrogated the assembly of the complex. Taken together, the
data suggest a stable core complex of
⬃
230 kDa containing PIF1, -2, and -3, which
appears to be a prerequisite for full complex assembly. We postulate that during
assembly the core complex may form initially and serve as a docking station for other
constituents to either form a full complex directly (Fig. 8A, left panel) or assemble the
⬃
400-kDa intermediate complex at first and subsequently add PIF8 to form the
⬃
500-kDa complete complex (Fig. 8A, right panel).
Our result is different from previous postulates of the AcMNPV core complex, which
was thought to be
⬃
170 kDa and to contain PIF1 to -4 (3, 22). It is now clear that PIF4
is not a component of the core complex, as deletion of
pif4
still yields an
⬃
230-kDa
complex (Fig. 5G). The difference between current results and previous data can be
explained as follows. (i) The relatively low resolution of partially denaturing SDS-PAGE
made it difficult to distinguish large proteins; the
⬃
150-kDa band, for example, may
represent the
⬃
230-kDa core complex. (ii) As mentioned previously, the partial
dena-turation method could not entirely inhibit proteolytic degradation by OB endogenous
proteinase (23). In our experience, different batches of larvae may have different
FIG 8Proposed model of PIF complex formation and intracellular transport. (A) Process of PIF complex formation. First, PIF1, -2, and -3 form a core complex which serves as a docking station, and then other constituents are recruited to the core, either directly to form a full complex (left) or via an about 400-kDa intermediate complex to which PIF8 is added to form the full PIF complex (right), which consists of nine PIFs. (B) Intracellular formation and transportation of the PIF complex. There are two possibilities for PIF complex formation and transportation, as follows. (1) The PIF complex is formed in the ER, and the INM-SM of the PIFs directs the PIF complex into the inner nuclear membrane; or (2) individual PIFs are transported into the nucleus via INM-SM and assembled in the nucleus. Either way, the entire PIF complex ends up at microvesicles (MVs) and is finally incorporated into the envelopes of ODVs. The ODVs are then embedded into OBs. VS, virogenic stroma.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:12.585.58.544.72.423.2]activities of endogenous proteinases. These proteinases would make the subcomplex
unstable if OBs were not pretreated with heat or proteinase inhibitors. (iii) The
differ-ence in pretreatments of the samples (SBTI inhibition or heating) may also have
different impacts on the forms of the complex.
The nine PIF proteins in the complex have an accumulated size of
⬃
360 kDa, less
than the observed
⬃
500 kDa. We cannot unequivocally exclude the possibility that
there are one or more host proteins associated with the complex. However, it is more
likely that some PIF proteins are present as multimers. The
⬃
230-kDa core complex is
larger than the estimated size (
⬃
127 kDa) of individual PIF1, -2, and -3 subunits, and we
therefore postulate that some may be oligomers. Assuming that the
⬃
230-kDa core
complex consists of PIF1, -2, and -3, the presence of PIF8 (
⬃
110 kDa) and other
monomer-form PIF components (total,
⬃
130 kDa) adds up to about 470 kDa, which is
close to the observed
⬃
500-kDa size of the PIF complex. Due to the lack of specific
antibodies against PIF4, -6, and 7, we are as of yet unable to propose a convincing
composition model of the complex based on the current data. Future experiments
using techniques such as crystal structure analysis and/or cryo-electron microscopy will
be needed to gain a more detailed structure of the PIF complex.
ODV nucleocapsids are assembled and enveloped in the nuclei of infected cells, and
given the size of the PIF complex, we were a bit surprised to find that the full complex
already exists in the cytoplasm (Fig. 7). Taking the stoichiometry of the complex and the
amount produced in the cytoplasm, the PIF complex found in the cytoplasm likely
reflects the genuine process of PIF complex assembly, although we cannot exclude the
possibility that it might be formed incidentally during the experimental process. It is
now conceivable that the PIF proteins are transported as a complex from the cytoplasm
to the nucleus. Many PIFs are rich in conserved cysteines (4), implying the requirement
of protein folding machinery for correct assembly. Formation of the PIF complex in the
cytoplasm may take advantage of such cellular machinery in the endoplasmic reticulum
(ER). The INM-SM of the PIFs may direct the PIF complex into the inner nuclear
membrane, intranuclear microvesicles (MVs), and eventually the envelopes of ODVs as
INM-SM-containing proteins (31). Our results, however, do not exclude the possibility
that individual PIFs can be transported into the nucleus and are assembled into
complexes there. It is notable that the monomeric PIFs, when not in a complex due to
pif
knockout, apparently also move to the nucleus, as evidenced on the envelopes of
ODVs (Fig. 4B). This is likely due to the presence of INM-SM in the PIFs (32). In the future,
it will be interesting to reveal the mechanism for how exactly the PIF complex is
assembled and transported to the nucleus. Based on current knowledge, a model of PIF
complex formation and intracellular transport is proposed and illustrated in Fig. 8.
Our results confirmed the previous finding that the PIF complex protects PIF
proteins from proteolytic degradation in the larval midgut (23) (Fig. 7). In addition, we
showed that deletion of any component of the PIF complex resulted in disruption of
the full complex structure (Fig. 5) and in protein degradation (Fig. 7A). This
demon-strates that each individual component is required for complex formation to allow
successful oral infection. Apart from protecting proteins from degradation, the essential
role of the PIF complex is expected to include binding and fusion activities. So far, it is
unknown which PIF is responsible for binding and/or fusion activities. For future
investigations, we need to distinguish the role of each individual PIF from its impact on
the integrity of the PIF complex. Since our results highlighted the important role of the
core complex in the assembly of the full complex, our next goal is to understand how
the core complex is formed and how it interacts with other PIFs to form the full
complex.
MATERIALS AND METHODS
Amplification and purification of viruses.Ac-wt and AcMNPV-WIV1-Syn1 (29) were propagated by orally infecting fourth-instarS. exigualarvae. AcMNPVs withpifdeletions were amplified by intrahemo-coelic injection of BV intoS. exigualarvae. HearNPV (33) was amplified in fourth-instarH. armigeralarvae, and BmNPV (34) was amplified in fourth-instarB. morilarvae. OBs were collected from diseased larvae and purified as previously described (35). ODVs were purified from OBs as previously reported (24), with
on November 6, 2019 by guest
http://jvi.asm.org/
minor modifications. To prevent protein degradation, SBTI (Sigma-Aldrich, St. Louis, MO) at a final concentration of 1 mg/109OBs was added prior to treatment with alkaline DAS buffer (0.1 M Na
2CO3,
0.15 M NaCl, and 10 mM EDTA; pH 11.0). After DAS treatment at room temperature for 5 min, the solution was neutralized with a 1/10 volume of 500 mM Tris-HCl (pH 7.5). The ODV suspensions were subjected to sucrose gradient (30 to 60% [wt/vol]) ultracentrifugation at 72,000⫻gfor 1 h (SW28 rotor; Beckman Coulter, Brea, CA), after which ODV-containing bands were collected and diluted with double-distilled water (ddH2O), followed by centrifugation at 72,000⫻gfor 1 h. The ODV pellets were then resuspended
in ddH2O for subsequent experiments. ODV purification in this study was performed as described above
unless otherwise indicated.
Antibodies.The rabbit polyclonal antibodies (pAbs) anti-AcPIF0, anti-AcPIF1, anti-AcPIF2, anti-AcPIF5, AcODV-E66, AcODV-E25, AcGP41, GAPDH, lamin B, AcGP64, AcVP80, anti-AcP33, anti-Ac-polyhedrin, anti-AcVP39, anti-HaPIF0, anti-HaPIF1, anti-HaPIF2, and anti-HaPIF3 were produced in our lab as reported previously (36–39). Other antibodies are reported here for the first time; these include the rabbit pAbs anti-AcPIF8 (amino acids [aa] 466 to 847), anti-AcEC43, anti-Ac49K, anti-AcVP80, anti-Ac66, anti-Ac81 (aa 15 to 164), anti-HaPIF5 (aa 1 to 318), and anti-HaPIF8 (aa 180 to 816) as well as the mouse pAbs anti-AcPIF1 (aa 24 to 530) and anti-AcODV-E18 (aa 20 to 62). These pAbs were generated by injecting rabbits or mice with purified prokaryotically expressed full-length or truncated proteins (as indicated above). The monoclonal antibodies (MAbs) anti-AcPIF3 (33A6), anti-AcPIF9 (26A9), and anti-HaPIF6 (38C10) were isolated after 3 rounds of screening of mouse hybridoma cells. An HA tag MAb and horseradish peroxidase-conjugated secondary antibodies against rabbit or mouse IgG were purchased from Sigma-Aldrich.
BN-PAGE and LC-MS/MS analysis.ODV suspensions were incubated with an equal volume of TN buffer (100 mM Tris, pH 7.4, and 2% NP-40) in the presence of a proteinase inhibitor cocktail (Roche, Basel, Switzerland) at 4°C for 30 min. After centrifugation at 20,000⫻gfor 30 min, the supernatant was collected in a new tube and mixed with 4⫻NativePAGE sample buffer (200 mM Bis-Tris, 64 mM HCl, 200 mM NaCl, 40% [wt/vol] glycerol, and 0.004% Ponceau S; pH 7.2), after which 5% Coomassie brilliant blue G-250 was added to the sample at a final concentration of 1/4 the concentration of NP-40. The samples were analyzed by BN-PAGE or stored at⫺80°C until further use. Electrophoresis and staining were performed with a NativePAGE Novex Bis-Tris gel system (Life Technologies, Carlsbad, CA) according to the manufacturer’s instructions. For Western blot analysis, proteins were semidry blotted (Bio-Rad Laboratories, Hercules, CA) with Tris-glycine buffer and developed by MicroChemi (DNR Bio-Imaging Systems, Neve Yamin, Israel).
Visible bands for the PIF complex were excised from the gel and treated with 25 mM ammonium bicarbonate in 50% (vol/vol) acetonitrile, followed by reduction, alkylation, and incubation with 0.01 mg/ml trypsin overnight. The digested peptides were extracted with 5% (vol/vol) formic acid in 50% (vol/vol) acetonitrile and vacuum dried. The digested samples were analyzed using Q Exactive spec-trometers (Thermo Fisher Scientific, Waltham, MA) coupled with an Easy-nLC 1200 Nanoflow liquid chromatography system (Thermo Fisher Scientific). Tryptic digests were loaded onto a C18 column
(75-m internal diameter and 25-cm length; Thermo Fisher Scientific) and eluted by use of a gradient of 5 to 35% (vol/vol) acetonitrile in 0.1% (vol/vol) formic acid at a flow rate of 300 nl/min for 90 min. The MS full-scan range wasm/z350 to 1,300, with resolutions of 700,000 at 200m/z. The top 20 peaks were subjected to MS/MS analysis with resolutions of 17,500 at 200m/z. The dynamic exclusion time was set to 18 s. Data were acquired using Xcalibur software (version 4.0; Thermo Fisher Scientific).
Proteins were identified by searching against NCBI databases by use of Mascot, version 2.3 (Matrix Science, London, United Kingdom), implemented in Proteome Discoverer 2.1 (Thermo Fisher Scientific). Accession numbers in the databases were NC_001623.1 (AcMNPV), NC_001962.1 (BmNPV), and NC_002654.2 (HearNPV). Proteins with at least two unique peptides with a 1% false-discovery rate at the peptide level were considered positive identifications.
Construction of recombinant AcMNPVs.As no specific antibodies to AcMNPV PIF4, -6, or -7 were obtained, recombinant AcMNPVs with HA-tagged PIFs were generated by use of the Bac-to-Bac system as previously reported (9, 11, 12), with minor modifications (Fig. 2A). Briefly, thepif4-knockout bacmid was generated by inserting the chloramphenicol resistance gene (Cmr) and anegfpcassette between nucleotides (nt) 386 and 389 ofpif4by homologous recombination inEscherichia coli.polyhedrin(polh) andpif4fused with HA were used to repair thepif4-knockout bacmid by Tn7-mediated transposition at thepolhlocus. The bacmid was used to transfect Sf9 cells, and progeny viruses were harvested. After confirmation by PCR and sequencing, the correct virus was named Ac-pif4HA. As there is overlap between the sequences oflef3andpif6, a double-knockout bacmid was first generated by replacing the sequence region covering nt 1 to 171 ofpif6and 1 to 792 oflef3with theCmrandegfpcassettes to generate Ac-pif6HA. HA-taggedpif6andlef3together withpolhwere then used to repair the bacmid by transposition. For Ac-pif7HA, nt 29 to 145 ofpif7were replaced with theCmrandegfpcassettes.
To generate 10 individual pif-knockout viruses, a synthetic biology method similar to that for generating AcMNPV-WIV1-Syn1 (29) was employed (Fig. 2B to K and Fig. 4A). This method is based on three rounds of transformation-associated recombination (TAR) in yeast cells. Briefly, 45 overlapping fragments (A1 to A45;⬃3 kb each) covering the entire genome of AcMNPV were produced by PCR and recombined into nine fragments (B1 to B9;⬃15 kb each) by TAR in yeast. The B fragments were further recombined into three larger intermediates (C1 to C3;⬃45 kb each), and the complete AcMNPV-WIV1-Syn1 genome was finally obtained by TAR with three C fragments in yeast. The synthetic virus was rescued by transfecting Sf9 cells with genomic DNA. To generatepif0-knockout bacmids, the region of nt 119428 to 121229, located on fragments A41 and A42 of the AcMNPV genome (accession number
KM667940), was deleted by overlapping PCR (Fig. 2B). The AcΔpif0bacmid was obtained by three rounds of TAR as described above, but with modified A41 and A42. Similarly, forpif1knockout, fragment A36 was
on November 6, 2019 by guest
http://jvi.asm.org/
modified by deleting nt 100709 to 102384 of AcMNPV to obtain the AcΔpif1bacmid by three rounds of TAR in yeast (Fig. 2C). To generate AcΔpif2, AcMNPV nt 17388 to 18453, which contained the majority of thepif2open reading frame (ORF), were deleted; however, part of the 5=end of the gene, encoding the first 24 aa ofpif2, was retained because it contains the transcriptional initiation motif of the neighboring genearif1(40) (Fig. 2D). To ensure that the remaining amino acids were not synthesized, the transcrip-tional initiation motif TAAG ofpif2was mutated to TGAG, and a G was inserted right after nt⫹4 of the translational start codon (ATG) to generate a frameshift mutation (Fig. 2D). To generate thepif3knockout, nt 99281 to 99874 of AcMNPV were deleted in fragment A35 (Fig. 2E). To knock outpif4without affecting neighboring genes, the transcriptional initiation motif TAAG ofpif4was mutated to GAGG. To further inhibit the synthesis of PIF4, a stop codon (TGA) was generated by mutating TTG just a few nucleotides downstream of the start codon, and an insertion of TA generated another stop codon (TAA) (Fig. 2F). To knock outpif5, AcMNPV nt 129091 to 130235 were removed from fragments A44 and A45 (Fig. 2G). In the case ofpif6, the transcriptional initiation motif TAAG ofpif6was converted to TGAG, and a TGG just downstream of the ATG was mutated to the stop codon TGA. A second stop codon (TGA) was introduced when a C was converted to T. In addition, an inserted C generated a frameshift mutation (Fig. 2H). To knock outpif7, the transcriptional initiation motif TAAG was mutated to TCAG, and a T was inserted right after the translational start codon (ATG) to generate a stop codon (Fig. 2I). Forpif8, the majority of the sequence was deleted, except for a fragment of 325 nt at the 5=end and an essential region for nucleocapsid assembly in the middle of the gene (41). To ensure that the remaining sequence does not encode amino acids, the transcriptional initiation motif TAAG ofpif8was mutated to TAAA, and a T at nt 58 of the ORF was deleted to generate a stop codon and a frameshift mutation (Fig. 2J). Similar to that ofpif7,pif9knockouts were generated by mutating the transcriptional initiation motif TAAG to TCAG and adding a T right after the translational start codon (ATG) to generate a stop codon (Fig. 2K). After confirmation by sequencing, the synthesized bacmids were used to transfect Sf9 cells, and then progeny viruses were collected for further amplification and infectivity testsin vitroandin vivo. Viruses with individually inactivatedpifgenes were designated AcΔpif0to AcΔpif9.
One-step growth curve assay.Sf9 cells (3⫻106) were infected with the viruses AcMNPV-WIV-Syn1
and AcΔpif9at a multiplicity of infection (MOI) of 5 50% tissue culture infective dose (TCID50) units/cell.
At 0, 24, 48, 72, and 96 h p.i., the cell supernatants were collected and the virus titers were measured by endpoint dilution assay. Virus infections were performed in triplicate. BV titers at specified time points were analyzed statistically by one-way analysis of variance (ANOVA), and the results were presented using GraphPad Prism 6 software.
Electron microscopy.Purified OBs of AcMNPV-WIV-Syn1 and AcΔpif9were analyzed by scanning electron microscopy (SEM) and transmission electron microscopy (TEM) as previously reported (24). Briefly, OBs were dried on tinfoil paper and sputter coated with gold for SEM analysis (Hitachi SU-8010 instrument). For TEM analysis, OBs were fixed with 2.5% (vol/vol) glutaraldehyde at 4°C overnight and ultrathinly sectioned for examination by TEM (Hitachi H-7000FA instrument).
Feeding assays.OBs of Ac-wt and AcΔpif0to AcΔpif9were purified from diseased larvae infected through BV injections and then used in feeding assays by the droplet method (42). Briefly, OBs were diluted with feeding buffer (1% [wt/vol] brilliant blue and 10% [wt/vol] sucrose) to a final concentration of 3⫻109OBs/ml and fed to 16-h-starved early-fourth-instarS. exigualarvae. Mortality was recorded at
3 days postinfection (p.i.). The feeding assay was performed twice independently.
Cytoplasmic and nuclear fractionation.Fractionation of the cytoplasm and nucleus was performed according to the method in a previous report (11), with modifications. Sf9 cells (1.2⫻108) were infected
with wild-type AcMNPV budded virus at an MOI of 5 TCID50units/cell. At 72 h p.i., infected cells were
harvested and washed with cold PBS three times. Cells were resuspended in 2 ml of NP-40 lysis buffer (PBS, 0.5% NP-40, proteinase inhibitor cocktail) and incubated on ice for 30 min. Lysis of the plasma membrane but not the nuclear membrane was confirmed by microscopy. Cell lysates were spun at 1,000⫻gfor 3 min. The supernatant (1.8 ml was taken for subsequent experiments, and the rest [0.2 ml] was discarded) and the pellet (washed with 1 ml lysis buffer and then resuspended with 2 ml lysis buffer) were collected as the cytoplasmic and nuclear fractions, respectively. GAPDH and lamin B were used as marker proteins to show the efficiency of fractionation. For BN-PAGE analysis of the cytoplasmic samples, addition of 4⫻NativePAGE sample buffer and digitonin (final concentration, 1%) plus G-250 and 10 s of sonication were required before sample loading.
Co-IP analysis.Protein A beads (NewZongKe Viral Disease Control Bio-Tech LTD, China) were precoated with PIF1 mouse polyclonal antibody or preimmune mouse serum as a control and incubated with the AcMNPV-infected cytosolic fraction overnight at 4°C. After washing 4 times with lysis buffer, the precipitated proteins were analyzed by Western blotting using the different PIF antibodies.
Proteinase protection assays.Gut lumen fluid was taken from the mouths of fifth-instarS. exigua
larvae by squeezing their abdomens. Collected gut fluid was centrifuged at 2,400⫻gfor 5 min, and then the supernatant was harvested, divided into 10-l aliquots, and stored at⫺80°C until further use. The proteinase activity of the collected gut fluid was determined by dose-dependent digestion of bovine serum albumin (BSA).
Envelope proteins were extracted from AcMNPV ODVs by incubating purified virions with 1% NP-40 in ddH2O at 4°C for 30 min, followed by harvesting of supernatants after centrifugation at 20,000⫻gfor
30 min. The envelope fractions (ODV envelope proteins from 1⫻109OBs) were digested with proteinase
K (40 ng/ml) or gut lumen fluid (0.2% [vol/vol]) at 37°C for 5 min. Proteins were loaded at 1⫻108
OB-derived proteins per lane for Western blot analysis.
on November 6, 2019 by guest
http://jvi.asm.org/
ACKNOWLEDGMENTS
We thank Youling Zhu and Fan Zhang from the Experimental Animal Center, Wuhan
Institute of Virology, for help with antibody preparation; Gengfu Xiao from the Wuhan
Institute of Virology for providing the TAR clone vector pGF; Yongping Huang from
the Institute of Plant Physiology and Ecology, Chinese Academy of Sciences, for
providing
B. mori
; and Basil M. Arif and Mark Goettel for manuscript editing.
This work was supported by the Key Research Program of Frontier Sciences of
the Chinese Academy of Sciences (grant QYZDJ-SSW-SMC021), the Strategic Priority
Research Program of the Chinese Academy of Sciences (grant XDB11030400), the
National Natural Science Foundation of China (grants 31621061 and 31130058), and
the Virology Key Frontier Science Program of the State Key Laboratory of Virology
(grant klv-2016-03).
REFERENCES
1. Harrison RL, Herniou EA, Jehle JA, Theilmann DA, Burand JP, Becnel JJ, Krell PJ, van Oers MM, Mowery JD, Bauchan GR, ICTV Report Consortium. 2018. ICTV virus taxonomy profile: Baculoviridae. J Gen Virol 99: 1185–1186.https://doi.org/10.1099/jgv.0.001107.
2. Rohrmann G. 2013. The AcMNPV genome: gene content, conservation, and function, p 1– 48.InBaculovirus molecular biology, 3rd ed. National Center for Biotechnology Information. Bethesda, MD.
3. Boogaard B, van Oers MM, van Lent JWM. 2018. An advanced view on baculovirus per os infectivity factors. Insects 9:84. https://doi.org/10 .3390/insects9030084.
4. Wang X, Liu X, Makalliwa GA, Li J, Wang H, Hu Z, Wang M. 2017. Per os infectivity factors: a complicated and evolutionarily conserved entry machinery of baculovirus. Sci China Life Sci 60:806 – 815.https://doi.org/ 10.1007/s11427-017-9127-1.
5. Faulkner P, Kuzio J, Williams GV, Wilson JA. 1997. Analysis of p74, a PDV envelope protein of Autographa californica nucleopolyhedrovirus re-quired for occlusion body infectivity in vivo. J Gen Virol 78:3091–3100.
https://doi.org/10.1099/0022-1317-78-12-3091.
6. Kikhno I, Gutierrez S, Croizier L, Croizier G, Ferber ML. 2002. Character-ization of pif, a gene required for the per os infectivity of Spodoptera littoralis nucleopolyhedrovirus. J Gen Virol 83:3013–3022. https://doi .org/10.1099/0022-1317-83-12-3013.
7. Pijlman GP, Pruijssers AJ, Vlak JM. 2003. Identification of pif-2, a third conserved baculovirus gene required for per os infection of insects. J Gen Virol 84:2041–2049.https://doi.org/10.1099/vir.0.19133-0. 8. Ohkawa T, Washburn JO, Sitapara R, Sid E, Volkman LE. 2005. Specific
binding of Autographa californica M nucleopolyhedrovirus occlusion-derived virus to midgut cells of Heliothis virescens larvae is mediated by products of pif genes Ac119 and Ac022 but not by Ac115. J Virol 79:15258 –15264.https://doi.org/10.1128/JVI.79.24.15258-15264.2005. 9. Fang M, Nie Y, Harris S, Erlandson MA, Theilmann DA. 2009. Autographa
californica multiple nucleopolyhedrovirus core gene ac96 encodes a per os infectivity factor (PIF-4). J Virol 83:12569 –12578.https://doi.org/10 .1128/JVI.01141-09.
10. Sparks WO, Harrison RL, Bonning BC. 2011. Autographa californica mul-tiple nucleopolyhedrovirus ODV-E56 is a per os infectivity factor, but is not essential for binding and fusion of occlusion-derived virus to the host midgut. Virology 409:69 –76.https://doi.org/10.1016/j.virol.2010.09 .027.
11. Nie Y, Fang M, Erlandson MA, Theilmann DA. 2012. Analysis of the Autographa californica multiple nucleopolyhedrovirus overlapping gene pair lef3 and ac68 reveals that AC68 is a per os infectivity factor and that LEF3 is critical, but not essential, for virus replication. J Virol 86: 3985–3994.https://doi.org/10.1128/JVI.06849-11.
12. Liu J, Zhu L, Zhang S, Deng Z, Huang Z, Yuan M, Wu W, Yang K. 2016. The Autographa californica multiple nucleopolyhedrovirus ac110 gene en-codes a new per os infectivity factor. Virus Res 221:30 –37.
13. Javed MA, Biswas S, Willis LG, Harris S, Pritchard C, van Oers MM, Donly BC, Erlandson MA, Hegedus DD, Theilmann DA. 2017. Autog-rapha californica multiple nucleopolyhedrovirus AC83 is a per os
infectivity factor (PIF) protein required for occlusion-derived virus (ODV) and budded virus nucleocapsid assembly as well as assembly of the PIF complex in ODV envelopes. J Virol 91:e02115-16.https:// doi.org/10.1128/JVI.02115-16.
14. Braunagel SC, Summers MD. 2007. Molecular biology of the baculovirus occlusion-derived virus envelope. Curr Drug Targets 8:1084 –1095. 15. Wang Y, Bininda-Emonds OR, van Oers MM, Vlak JM, Jehle JA. 2011. The
genome of Oryctes rhinoceros nudivirus provides novel insight into the evolution of nuclear arthropod-specific large circular double-stranded DNA viruses. Virus Genes 42:444 – 456.https://doi.org/10.1007/s11262 -011-0589-5.
16. Garcia-Maruniak A, Abd-Alla AM, Salem TZ, Parker AG, Lietze VU, van Oers MM, Maruniak JE, Kim W, Burand JP, Cousserans F, Robinson AS, Vlak JM, Bergoin M, Boucias DG. 2009. Two viruses that cause salivary gland hypertrophy in Glossina pallidipes and Musca domestica are related and form a distinct phylogenetic clade. J Gen Virol 90:334 –346.
https://doi.org/10.1099/vir.0.006783-0.
17. Gauthier L, Cornman S, Hartmann U, Cousserans F, Evans JD, de Miranda JR, Neumann P. 2015. The Apis mellifera filamentous virus genome. Viruses 7:3798 –3815.https://doi.org/10.3390/v7072798.
18. Bézier A, Theze J, Gavory F, Gaillard J, Poulain J, Drezen JM, Herniou EA. 2015. The genome of the nucleopolyhedrosis-causing virus from Tipula oleracea sheds new light on the Nudiviridae family. J Virol 89: 3008 –3025.https://doi.org/10.1128/JVI.02884-14.
19. Bézier A, Annaheim M, Herbiniere J, Wetterwald C, Gyapay G, Bernard-Samain S, Wincker P, Roditi I, Heller M, Belghazi M, Pfister-Wilhem R, Periquet G, Dupuy C, Huguet E, Volkoff AN, Lanzrein B, Drezen JM. 2009. Polydnaviruses of braconid wasps derive from an ancestral nudivirus. Science 323:926 –930.https://doi.org/10.1126/science.1166788. 20. Peng K, van Oers MM, Hu Z, van Lent JW, Vlak JM. 2010. Baculovirus per
os infectivity factors form a complex on the surface of occlusion-derived virus. J Virol 84:9497–9504.https://doi.org/10.1128/JVI.00812-10. 21. Wittig I, Braun HP, Schagger H. 2006. Blue native PAGE. Nat Protoc
1:418 – 428.https://doi.org/10.1038/nprot.2006.62.
22. Peng K, van Lent JW, Boeren S, Fang M, Theilmann DA, Erlandson MA, Vlak JM, van Oers MM. 2012. Characterization of novel components of the baculovirus per os infectivity factor complex. J Virol 86:4981– 4988.
https://doi.org/10.1128/JVI.06801-11.
23. Boogaard B, van Lent JWM, Theilmann DA, Erlandson MA, van Oers MM. 2017. Baculoviruses require an intact ODV entry-complex to resist teolytic degradation of per os infectivity factors by co-occluded pro-teases from the larval host. J Gen Virol 98:3101–3110.https://doi.org/10 .1099/jgv.0.000974.
24. Wang X, Chen C, Zhang N, Li J, Deng F, Wang H, Vlak JM, Hu Z, Wang M. 2018. The group I alphabaculovirus-specific protein, AC5, is a novel component of the occlusion body but is not associated with ODVs or the PIF complex. J Gen Virol 99:585–595. https://doi.org/10.1099/jgv.0 .001031.
25. Blissard GW, Theilmann DA. 2018. Baculovirus entry and egress from insect cells. Annu Rev Virol 5:113–139.https://doi.org/10.1146/annurev -virology-092917-043356.
26. Simon O, Palma L, Williams T, Lopez-Ferber M, Caballero P. 2012. Anal-ysis of a naturally-occurring deletion mutant of Spodoptera frugiperda multiple nucleopolyhedrovirus reveals sf58 as a new per os infectivity factor of lepidopteran-infecting baculoviruses. J Invertebr Pathol 109: 117–126.https://doi.org/10.1016/j.jip.2011.10.010.
27. Tang Q, Li G, Yao Q, Chen L, Lv P, Lian C, Chen K. 2013. Bm91 is an envelope component of ODV but is dispensable for the propagation of
on November 6, 2019 by guest
http://jvi.asm.org/
Bombyx mori nucleopolyhedrovirus. J Invertebr Pathol 113:70 –77.
https://doi.org/10.1016/j.jip.2013.01.006.
28. Huang H, Wang M, Deng F, Wang H, Hu Z. 2012. ORF85 of HearNPV encodes the per os infectivity factor 4 (PIF4) and is essential for the formation of the PIF complex. Virology 427:217–223.https://doi.org/10 .1016/j.virol.2012.01.022.
29. Shang Y, Wang M, Xiao G, Wang X, Hou D, Pan K, Liu S, Li J, Wang J, Arif BM, Vlak JM, Chen X, Wang H, Deng F, Hu Z. 2017. Construction and rescue of a functional synthetic baculovirus. ACS Synth Biol 6:1393–1402.https://doi.org/10.1021/acssynbio.7b00028.
30. Hu X, Zhu M, Wang S, Zhu L, Xue R, Cao G, Gong C. 2015. Proteomics analysis of digestive juice from silkworm during Bombyx mori nucleopo-lyhedrovirus infection. Proteomics 15:2691–2700. https://doi.org/10 .1002/pmic.201400475.
31. Hong T, Summers MD, Braunagel SC. 1997. N-terminal sequences from Autographa californica nuclear polyhedrosis virus envelope proteins ODV-E66 and ODV-E25 are sufficient to direct reporter proteins to the nuclear envelope, intranuclear microvesicles and the envelope of occlu-sion derived virus. Proc Natl Acad Sci U S A 94:4050 – 4055.
32. Braunagel SC, Williamson ST, Saksena S, Zhong Z, Russell WK, Russell DH, Summers MD. 2004. Trafficking of ODV-E66 is mediated via a sorting motif and other viral proteins: facilitated trafficking to the inner nuclear membrane. Proc Natl Acad Sci U S A 101:8372– 8377.https://doi.org/10 .1073/pnas.0402727101.
33. Hu Z, Peters S, Vlak JM, Wang H, Chen X, Tarchini R, Lankhorst RK, IJkel WFJ, Zuidema D, Sandbrink H, Sun X. 2001. The sequence of the Heli-coverpa armigera single nucleocapsid nucleopolyhedrovirus genome. J Gen Virol 82:241–257.https://doi.org/10.1099/0022-1317-82-1-241. 34. Huang J, Hao B, Deng F, Sun X, Wang H, Hu Z. 2008. Open reading frame
Bm21 of Bombyx mori nucleopolyhedrovirus is not essential for virus replication in vitro, but its deletion extends the median survival time of infected larvae. J Gen Virol 89:922–930. https://doi.org/10.1099/vir.0 .83504-0.
35. Hou D, Zhang L, Deng F, Fang W, Wang R, Liu X, Guo L, Rayner S, Chen X, Wang H, Hu Z. 2013. Comparative proteomics reveal fundamental structural and functional differences between the two progeny pheno-types of a baculovirus. J Virol 87:829 – 839.https://doi.org/10.1128/JVI .02329-12.
36. Wang R, Deng F, Hou D, Zhao Y, Guo L, Wang H, Hu Z. 2010. Proteomics of the Autographa californica nucleopolyhedrovirus budded virions. J Virol 84:7233–7242.https://doi.org/10.1128/JVI.00040-10.
37. Li Y, Shen S, Hu L, Deng F, Vlak JM, Hu Z, Wang H, Wang M. 2018. The functional oligomeric state of tegument protein GP41 is essential for baculovirus budded virion and occlusion-derived virion assembly. J Virol 92:e02083-17.https://doi.org/10.1128/JVI.02083-17.
38. Kuang W, Zhang H, Wang M, Zhou NY, Deng F, Wang H, Gong P, Hu Z. 2017. Three conserved regions in baculovirus sulfhydryl oxidase P33 are critical for enzymatic activity and function. J Virol 91:e01158-17.https:// doi.org/10.1128/JVI.01158-17.
39. Song J, Wang X, Hou D, Huang H, Liu X, Deng F, Wang H, Arif BM, Hu Z, Wang M. 2016. The host specificities of baculovirus per os infectivity factors. PLoS One 11:e0159862. https://doi.org/10.1371/journal.pone .0159862.
40. Chen YR, Zhong S, Fei Z, Hashimoto Y, Xiang JZ, Zhang S, Blissard GW. 2013. The transcriptome of the baculovirus Autographa californica mul-tiple nucleopolyhedrovirus in Trichoplusia ni cells. J Virol 87:6391– 6405.
https://doi.org/10.1128/JVI.00194-13.
41. Huang Z, Pan M, Zhu S, Zhang H, Wu W, Yuan M, Yang K. 2017. The Autographa californica multiple nucleopolyhedrovirus ac83 gene con-tains acis-acting element that is essential for nucleocapsid assembly. J Virol 91:e02110-16.https://doi.org/10.1128/JVI.02110-16.
42. Hughes PR, van Beek NAM, Wood HA. 1986. A modified droplet feeding method for rapid assay of Bacillus thuringiensis and baculoviruses in noctuid larva. J Invertebr Pathol 48:187–192.https://doi.org/10.1016/ 0022-2011(86)90122-9.
on November 6, 2019 by guest
http://jvi.asm.org/