JOURNAL OFVIROLOGY, May 2007, p. 5385–5394 Vol. 81, No. 10 0022-538X/07/$08.00⫹0 doi:10.1128/JVI.02516-06
Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Major Subsets of Human Dendritic Cells Are Efficiently Transduced by
Self-Complementary Adeno-Associated Virus Vectors 1 and 2
䌤
Philippe Veron,
1,2Vale
´rie Allo,
1Christel Rivie
`re,
1Jacky Bernard,
3Anne-Marie Douar,
1and Carole Masurier
1*
Genethon, CNRS UMR 8115, 91002 Evry Cedex, France1; Genosafe S.A., 91002 Evry Cedex, France2; and
Institut Jean Godinot, Unite´ d’Immunothe´rapie Cellulaire, Reims, France3
Received 15 November 2006/Accepted 12 February 2007
Dendritic cells (DC) are antigen-presenting cells pivotal for inducing immunity or tolerance. Gene transfer into DC is an important strategy for developing immunotherapeutic approaches against infectious pathogens and cancers. One of the vectors previously described for the transduction of human monocytes or DC is the recombinant adeno-associated virus (rAAV), with a genome conventionally packaged as a single-stranded (ss) molecule. Nevertheless, its use is limited by the poor and variable transduction efficiency of DC. In this study, AAV type 1 (AAV1) and AAV2 vectors, which expressed the enhanced green fluorescent protein and were packaged as ss or self-complementary (sc) duplex strands, were used to transduce different DC subsets generated ex vivo and the immunophenotypes, states of differentiation, and functions of the subsets were carefully examined. We show here for the first time that a single exposure of monocytes (Mo) or CD34ⴙ
progenitors (CD34) to sc rAAV1 or sc rAAV2 leads to high transduction levels (5 to 59%) of differentiated Mo-DC, Mo-Langerhans cells (LC), CD34-LC, or CD34-plasmacytoid DC (pDC), with no impact on their
phenotypes and functional maturation of these cells, compared to those of exposure to ss rAAV. Moreover, we show that all these DC subpopulations can also be efficiently transduced after commitment to their differen-tiation pathways. Furthermore, these DC subsets transduced with sc rAAV1 expressing a tumor antigen were potent activators of a CD8ⴙ-T-cell clone. Altogether, these results show the high potential of sc AAV1 and sc AAV2 vectors to transduce ex vivo conventional DC, LC, or pDC or to directly target them in vivo for the design of new DC-based immunotherapies.
Dendritic cells (DC) are antigen-presenting cells (APC) piv-otal for regulating immune responses (43). They have a role in the activation and function of both innate and adaptive im-mune responses (2, 3). Furthermore, depending on the DC subset, their maturation stage, and the inflammatory stimuli that they encounter, DC can induce either immunity or toler-ance (28, 44). At least three subsets of mature human DC are known: myeloid conventional DC (cDC), Langerhans cells (LC), and plasmacytoid DC (pDC), with evidence of functional plasticity for a given DC subset (36). Indeed, the subsets ex-hibit some differences in their abilities to regulate T-cell re-sponses, to produce antiviral type I interferon, and to cross-present exogenous antigens to CD8⫹T cells.
Human DC, LC, and pDC can be easily generated from monocytes or CD34⫹progenitors, allowing the procurement, in considerable amounts, of these otherwise scarce cells, which represent approximately 0.2% of total white blood cells (5–8, 19, 21, 22, 32, 37, 39). The genetic modification of DC sub-populations offers great potential for the development of im-mune response-regulatory strategies for purposes ranging from active immunization to tolerance induction. This approach may provide long-lasting expression of the entire array of epitopes directly processed from a native protein and poten-tially allow their presentation by the complete set of human leukocyte antigen (HLA) molecules present on the APC.
Efficient gene delivery to human DC has been achieved mainly by using lentiviral vectors (LV) (40, 42, 48). One of the alternate vectors used to transduce monocytes or DC has been the recombinant adeno-associated virus (rAAV), with a ge-nome conventionally packaged as single-stranded (ss) mole-cules (9, 27, 34), despite the fact that the cloning capacity of this vector is more limited than that of LV (4.7 kb instead of 8 to 9 kb) (13, 50). Indeed, rAAV is unique among viruses that are being developed for gene transfer in that the wild-type virus has never been shown to cause human disease. Like LV, rAAV presents the ability to transduce both dividing and non-dividing cells, which may allow the transduction of DC from a broad range of sources and in various activation and matura-tion states (13, 50). Another advantage of rAAV is the absence of viral coding sequences, which may diminish the elimination of transduced DC by virus-specific cytolytic T cells (26, 55). So far, preclinical studies based on DC immunotherapies have been limited by the poor and variable levels of efficiency of DC transduction with ss rAAV (26, 34, 52).
Recently, AAV vectors have been diversified by alternate serotypes with different cell tropisms (51). It was recently shown that the ss AAV type 5 (ss AAV5) vector presents a higher degree of tropism for human DC than vectors of other serotypes, which had poor levels of efficiency (54). Moreover, some studies have demonstrated that the packaging of rAAV2 as a self-complementary duplex strand (sc) allows earlier and higher levels of transgene expression by bypassing the synthesis of the complementary strand of the AAV viral genome, which constitutes a limiting step for efficient cell transduction (30,
* Corresponding author. Mailing address: Laboratoire d’Immunologie, Genethon, CNRS UMR 8115, 91002 Evry Cedex, France. Phone: (33) 1-69-47-29-84. Fax: (33) 1-60-77-86-98. E-mail: [email protected].
䌤Published ahead of print on 21 February 2007.
5385
on November 8, 2019 by guest
http://jvi.asm.org/
49). A recent study reported the efficient transduction of mouse bone marrow-derived DC with sc rAAV6 (1).
In this study, we compared the levels of efficiency and the time courses of transduction of human DC subsets (cDC, LC, and pDC) with ss rAAV1 and ss rAAV2 or with their self-complementary counterparts. Furthermore, we carefully exam-ined the immunophenotypes, states of differentiation, and functionalities of these transduced DC subsets. We found that in contrast to exposure to ss rAAV, a single exposure of mono-cytes or CD34⫹progenitors to sc rAAV1 or sc rAAV2 enables high levels of transduction of cDC, LC, or pDC, with no impact on their phenotypes and functional maturation. We further showed that all these DC subsets can be efficiently transduced after commitment to their differentiation pathways. Finally, our results indicate that these cDC, LC, and pDC derived from monocytes or CD34⫹ progenitors are potent activators of a specific CD8⫹-T-cell clone after transduction with sc AAV1 vector expressing a tumor antigen. Altogether, these results show the strong potential of sc AAV1 and sc AAV2 vectors for cDC-, LC-, or pDC-based immunotherapy applications.
MATERIALS AND METHODS
AAV vector construction and production. Pseudotyped AAV vectors were generated by packaging AAV2-based recombinant genomes in AAV1 or AAV2 capsids. All the vectors used in the study were produced using the three-plasmid transfection protocol as described elsewhere (38). Briefly, HEK293 cells were tritransfected with the adenovirus helper plasmid pXX6 (53), a pAAV packaging
plasmid expressing therepandcapgenes (pACG2.1 for AAV2 and pLT-RC02
for AAV1), and the relevant pAAV2 vector plasmid. ss AAV vectors were produced with the conventional pGG2 AAV2 vector plasmid expressing en-hanced green fluorescent protein (E-GFP) under the transcriptional control of the cytomegalovirus (CMV) immediate early promoter associated with a simian virus 40 poly(A) signal. To produce sc AAV vectors, a modified pGG2-scCMV-GFP plasmid was constructed by deleting the D sequence and the terminal
resolution site (trs) from one of the inverted terminal repeats by using MscI
digestion, leaving the other repeat intact (49). To generate self-complementary vectors expressing the MART-1 peptide, the CMV GFP sequence of pGG2-scCMV-GFP was replaced with the MART sequence (ELAGIGLTV) (47) cou-pled with the phosphoglycerate kinase promoter. Recombinant vectors were
purified by double CsCl2ultracentrifugation followed by dialysis against sterile
phosphate-buffered saline. Physical particles were quantified by real-time PCR,
and vector titers are expressed as viral genomes (vg) per milliliter. The propor-tion of sc AAV vector genomes was assessed by alkaline agarose electrophoresis.
Cell line.The OP9 stroma cell line expressing human delta 1 (OP9-Del1) was kindly provided by A. Olivier (Genethon, Evry, France) and maintained as previously described (32).
Culture of peripheral blood monocytes and CD34ⴙcells.Each DC subset was
differentiated from monocytes or CD34⫹progenitors from at least three
differ-ent donors. Monocytes were generated from samples from normal volunteers
after the elutriation of peripheral blood according to the E´ tablissement Franc¸ais
du Sang procedures (J. Bernard, Reims, France). This method yielded purified
(92.2%⫾5.1%) CD14⫹CD45⫹cells as assessed by flow cytometry. Briefly,
cryopreserved monocytes were cultured in 6-well plates at a density of 106cells/ml
in RPMI 1640 (Invitrogen Life Technology, Auckland, New Zealand)
supple-mented with 10% fetal calf serum (FCS; HyClone, Logan, UT) and 1%L
-glutamine (Invitrogen). Monocytes were differentiated either into cDC (Mo-DC) in the presence of 50 ng/ml of recombinant human granulocyte-macrophage
colony-stimulating factor (Novartis, Baˆle, Switzerland) and 15 ng/ml of
recom-binant human interleukin-4 (IL-4; Tebu-bio, Le Perray, France) or into LC (Mo-LC) by the addition of 20 ng/ml of recombinant human transforming growth
factor(Abcys SA, Paris, France) (21). Maturation was induced in some
exper-iments by the addition of lipopolysaccharide (LPS; 7g/ml [Sigma-Aldrich, St.
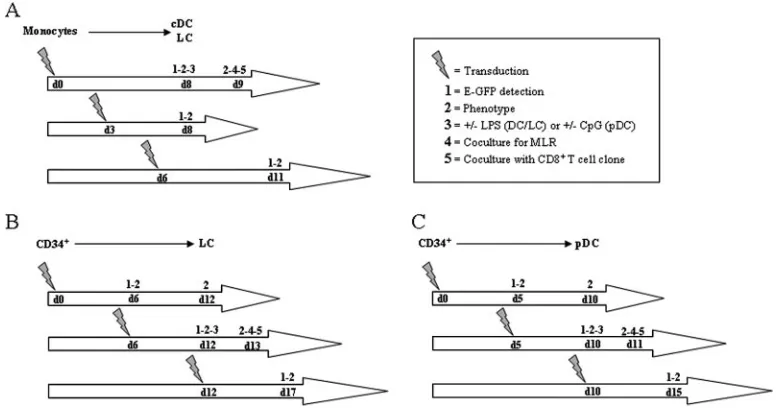
Louis, MO]) at day 8 for 24 h (Fig. 1A).
LC were generated from cord blood CD34⫹cells (CD34-LC) by following
protocols previously described by Caux et al. (6) with some modifications. Briefly,
CD34⫹progenitor cells were separated from cord blood mononuclear cells by
using the CD34 isolation kit (Miltenyi Biotech, GmbH, Bergisch Gladbach,
Germany). Cryopreserved CD34⫹progenitor cells were seeded at a density of
2⫻105
cells/ml in 24-well tissue culture plates in RPMI 1640 supplemented with
10% FCS, 1%L-glutamine (GIBCO/BRL, Rockville, MD), 50 ng/ml of
recom-binant human granulocyte-macrophage colony-stimulating factor, 15 ng/ml of recombinant human IL-4, 15 ng/ml of recombinant human transforming growth
factor, and 20 ng/ml of recombinant human tumor necrosis factor alpha. At day
6, cells were collected and plated in new 12-well plates at a density of 8⫻105
cells/ml. In some experiments, the maturation of CD34-LC was induced by the
addition of LPS (7g/ml) at day 12 for 24 h (Fig. 1B).
pDC were generated from cord blood CD34⫹cells (CD34-pDC) by following
protocols previously described by Olivier et al. (32). CD34⫹progenitors (2⫻
104) were added to OP9-Del1 cells seeded 1 day before in 24-well plates at 3⫻
104
cells/well. Cells were cultured in RPMI 1640 (Invitrogen) supplemented with
10% FCS (HyClone), 1%L-glutamine, and 1% penicillin-streptomycin (GIBCO)
in the presence of recombinant human Fms-like tyrosine kinase 3 ligand (5 ng/ml) and rIL-7 (5 ng/ml; R&D Systems, Minneapolis, MN). The maturation of CD34-pDC was induced in some experiments by the addition of CpG
oligode-oxynucleotide type A (ODN 2216 at 2M) at day 10 for 24 h (Fig. 1C). All cells
[image:2.585.97.488.69.273.2]were cultured in a humidified incubator at 37°C and 5% CO2.
FIG. 1. Graphic depiction of the experimental protocols. d, day; CD34⫹, CD34⫹progenitors;⫹, with;⫺, without.
5386 VERON ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
Flow cytometric analysis.The DC and T-cell phenotypes were assessed using three-color immunostaining with biotinylated phycoerythrin-, Cy-Chrome-, and allophycocyanin-conjugated monoclonal anti-CD86 (FUN-1), anti-CD11c (B-Ly6), anti-HLA-DR (G46.6), anti-Langerin (DCGM4), anti-cutaneous leukocyte antigen (anti-CLA; HECA-452), anti-CD8 (RPA-T8), and anti-CD3 (HIT3a; all purchased from Becton-Dickinson, Mountain View, CA) and anti-CD123 (AC145; Miltenyi Biotech). Data were acquired using a FACSCalibur flow cytometer (Becton Dickinson), and data analyses were performed using the CellQuest program (Becton Dickinson).
Transduction of monocytes and CD34ⴙcells with rAAV.After thawing,
mono-cytes and CD34⫹cells were transduced with 105
vg/cell of ss and sc rAAV types
1 and 2 at a fixed concentration of 1⫻107and 1⫻106cells/ml, respectively, in
RPMI 1640. After 3 h at 37°C, cells were cultured in complete medium as previously described and were analyzed by flow cytometry at different times, as indicated in Fig. 1.
Transduction of differentiated Mo-DC, Mo-LC, CD34-LC, and CD34-pDC with rAAV.Monocytes and CD34⫹progenitors were differentiated as described above into Mo-DC, Mo-LC, CD34-LC, or CD34-pDC. Semiadherent and non-adherent cells in culture were harvested at different times as depicted in Fig. 1 and transduced with ss and sc rAAV types 1 and 2 under the same conditions
described above for monocytes and CD34⫹cells. After 3 h at 37°C, cells were
re-placed into the same complete medium and then cultured for 5 to 6 additional days (Fig. 1).
MLRs.Enriched naı¨ve CD2⫹CD45⫹T cells were recovered after the
elutria-tion of monocytes. This method yielded purified (82.4%⫾6.1%) CD2⫹CD45⫹
cells as assessed by flow cytometry. CD2⫹CD45⫹T cells were labeled with
carboxyfluorescein diacetate succinimidyl ester (CFSE) at a final concentration
of 0.5M for 20 min at 37°C before being extensively washed. E-GFP-negative
and -positive Mo-DC and Mo-LC were sorted on a MoFlow flow cytometer (Dako, Glostrup, Denmark). For mixed leukocyte reactions (MLRs), matured allogeneic Mo-DC and Mo-LC (Fig. 1A) were extensively washed and cultured
in 96-well U-bottom plates at different densities with 1⫻105CFSE-labeled
CD2⫹CD45⫹T cells. On day 4, cells were harvested, washed, labeled for T
specificity with anti-CD3 antibody, and analyzed by flow cytometry. The
percent-age of dividing T cells was linearly correlated with the decrease in CFSE fluo-rescence.
Activation of a MART-1 CD8ⴙ-T-cell clone by transduced DC subpopulations.
Matured HLA-A2⫹DC subpopulations were obtained after the transduction of
cells with sc rAAV1 expressing a MART-1 peptide under the control of a CMV
promoter, as depicted in Fig. 1. Nontransduced matured HLA-A2⫹DC
sub-populations were pulsed with the high-affinity MART-1 peptide (ELAGIGLTV)
at 10M for 3 h and then washed extensively before coculture. These
nontrans-duced, transnontrans-duced, or pulsed DC subpopulations were cocultured in 96-well U-bottom plates at different ratios with a specific MART-1 HLA-A2-restricted
CFSE-labeled CD8⫹-T-cell clone (LT12) at 105
cells/well as described above for MLRs. On day 5, cells were harvested, washed, labeled with an CD8 anti-body, and analyzed by flow cytometry. The percentage of dividing T cells was linearly correlated with the loss of CFSE fluorescence.
Statistical analyses.Results are presented as means⫾standard deviations
(SD). Student’sttest for paired data was used to determine significant
differ-ences between two groups. AP value of ⬍0.05 was considered statistically
significant.
RESULTS
[image:3.585.98.480.75.306.2]Transduction of monocytes and CD34ⴙprogenitors with ss and sc rAAV1 and rAAV2.We first compared the gene transfer efficiency with ss and sc rAAV1 and rAAV2 (i) into monocytes that were then differentiated into cDC or LC and (ii) into CD34⫹progenitors that were further differentiated into LC or pDC with the appropriate cytokine cocktails as previously de-scribed (32, 48). These vectors expressing E-GFP driven by the ubiquitous CMV promoter were used under identical condi-tions. E-GFP expression is easily and accurately monitored by fluorescence-activated cell sorter analysis (FACS), allowing
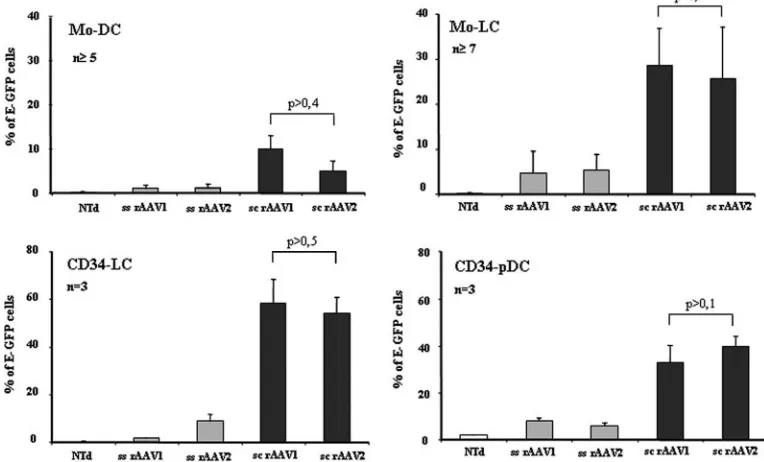
FIG. 2. Transduction efficiencies of monocytes and CD34⫹progenitors with ss and sc rAAV1 and rAAV2 serotypes. Monocytes or CD34⫹ progenitors were transduced with ss and sc rAAV1 and rAAV2 serotypes expressing E-GFP and then differentiated into cDC, LC, or pDC in the presence of the corresponding cytokine cocktail. Mo-DC, Mo-LC, CD34-LC, and CD34-pDC were analyzed 5 to 8 days posttransduction, as indicated in Fig. 1. For each subpopulation, the graphs show the percentages of E-GFP-expressing cells after transduction with the indicated vectors or without transduction (NTd). The percentages of E-GFP-expressing cells were obtained by flow cytometry. Cells were gated in forward scatter and side scatter; cDC, LC, and pDC were analyzed for E-GFP expression and markers characteristic of each DC subset, CD11c, CLA, and CD123, respectively. Results are expressed as the mean percentages of cells⫾SD as determined in the numbers of independent experiments indicated (n). Note that thePvalues indicated are always greater than 0.05, which represents a nonsignificant difference between the considered groups.
VOL. 81, 2007 TRANSDUCTION OF HUMAN DC WITH sc rAAV1 AND sc rAAV2 5387
on November 8, 2019 by guest
http://jvi.asm.org/
the enumeration of DC subpopulations on a single-cell basis. Preliminary experiments performed with different amounts of viral particles (5 ⫻ 103 to 1.5 ⫻ 104 vg/cell) at a fixed cell
density showed that maximum transduction levels were reached with 104 vg/cell, with no cellular toxicity (data not
shown). E-GFP expression and FACS analysis for DC subset identification were monitored 8 days posttransduction for Mo-DC and Mo-LC and 5 to 6 days posttransduction for CD34-LC and CD34-pDC, since the latter are dividing cells and AAV vectors are mainly episomal (Fig. 1). Mo-DC were analyzed for CD11c expression, Mo-LC and CD34-LC were analyzed for CLA expression, and CD34-pDC were analyzed for CD123 expression (32, 48). A single exposure of both monocytes and CD34⫹progenitors to ss rAAV1 or rAAV2 led to low levels of transduced Mo-DC, Mo-LC, CD34-LC, and CD34-pDC, with percentages of E-GFP-positive cells ranging from 1 to 8% (Fig. 2). In contrast, the transduction efficiencies of each DC subpopulation with sc rAAV under similar condi-tions were significantly increased: sc rAAV1 was 10-fold, six-fold, foursix-fold, and fourfold more efficient than its ss counter-part for Mo-DC, Mo-LC, CD34-LC, and CD34-pDC, respectively. Similarly, sc rAAV2 led to transduction levels 4.5-fold, 4.7-fold, 6.7-fold, and 6.6-fold higher than ss rAAV2 for Mo-DC, Mo-LC, CD34-LC, and CD34-pDC, respectively (Fig. 2). Of note, Mo-LC and CD34-LC, which were described as more immature DC (35), were more efficiently transduced
with both serotypes of sc rAAV than Mo-DC and CD34-pDC (Fig. 2). Altogether, these results show that sc rAAV vectors of serotypes 1 and 2 (i) are very efficient for the transduction of monocytes and CD34⫹progenitors compared to their ss coun-terparts and (ii) equivalent in transducing both cell types, fur-ther allowing for the generation of gene-modified cDC, LC, or pDC.
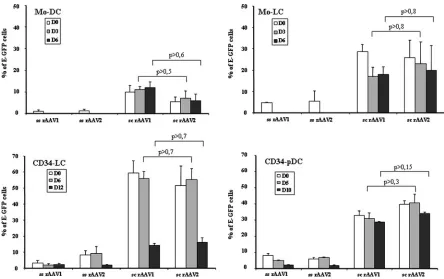
[image:4.585.68.512.66.345.2]Effects of DC differentiation on the efficiency of transduction with ss and sc rAAV1 and rAAV2.We evaluated whether cells already committed toward cDC, LC, or pDC could also be transduced with sc and ss rAAV1 and rAAV2 expressing E-GFP. Committed cells were transduced 3 or 6 days after the induction of differentiation (Mo-DC and Mo-LC), 6 or 12 days after the induction of differentiation (CD34-LC), and 5 or 10 days after the induction of differentiation (CD34-pDC) and then analyzed 5 to 6 days posttransduction by flow cytometry to detect E-GFP expression (Fig. 1). Day-3 and -6 Mo-DC and Mo-LC were efficiently transduced with sc rAAV1 and rAAV2 at levels similar to those at day 0 (6 to 23%). In contrast, no transduction with the ss counterpart vectors was seen (Fig. 3). As observed at day 0, cells committed toward LC were more efficiently transduced with both vectors than cells committed toward DC. Levels LC transduction were 1.5- and 2.5-fold higher than those DC when transduced with sc rAAV1 and sc rAAV2, respectively, 3 days after the induction of differenti-ation. These differences reached 1.9- and 3.5-fold when the
FIG. 3. Transduction efficiencies of DC subpopulations with ss and sc rAAV1 and rAAV2 serotypes. Mo-DC and Mo-LC were transduced at day 0 (D0) or at day 3 or 6 after the induction of differentiation. CD34-LC and CD34-pDC were transduced at day 0, 6, or 12 and at day 0, 5, or 10 after the induction of differentiation, respectively (Fig. 1). Cells were transduced with ss or sc rAAV and rAAV2 and then cultured in the same medium in the presence of the appropriate cytokine differentiation cocktail. Cells in final cultures were analyzed for E-GFP expression by flow cytometry 5 to 6 days posttransduction. The percentage of E-GFP-expressing cells in each DC subpopulation was obtained as described in the legend to Fig. 2 (n⫽3). Note that thePvalues indicated are always greater 0.05, which represents a nonsignificant difference between the considered groups.
5388 VERON ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
transduction occurred 6 days after the induction of differenti-ation (Fig. 3). In contrast to monocyte-derived cells, CD34⫹ progenitor-derived cells could still be transduced with both ss rAAV serotypes after differentiation, although with very low effectiveness (less than 9%). Furthermore, CD34-LC were ef-ficiently transduced 6 days after the induction of differentiation but not at 12 days and CD34-pDC were efficiently transduced at both 5 and 10 days with both sc serotypes (14 to 56%) after commitment to their differentiation pathways, like CD34⫹ pro-genitors (Fig. 3). In addition, day-6 CD34-LC were 1.8- and 1.4-fold more efficiently transduced with sc rAAV1 and rAAV2, respectively, than CD34-pDC (Fig. 3). These results indicate that monocytes and CD34⫹progenitors already com-mitted toward cDC, LC, or pDC can be efficiently transduced by a single infection with sc rAAV1 and sc rAAV2 serotypes.
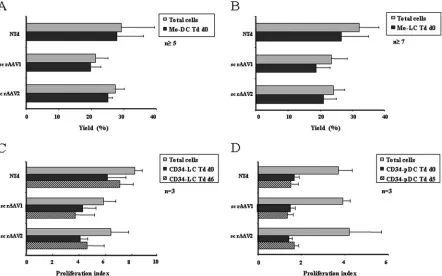
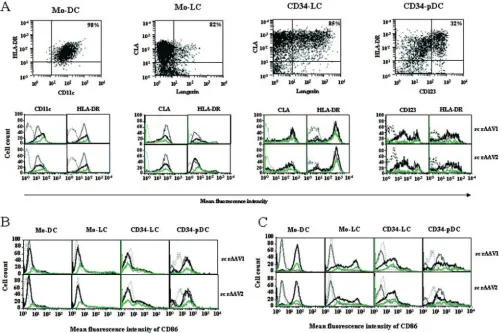
Yields and immunophenotypical analysis of sc rAAV-trans-duced cDC, LC, and pDC.In order to better understand the mechanisms involved in the immunogenicity of sc AAV vec-tors, it is interesting to investigate the effects of these vectors on DC phenotypes and maturation states. We characterized immunophenotypically the different nontransduced and sc rAAV-transduced DC subpopulations generated either from monocytes or from CD34⫹progenitors. Nontransduced mono-cytes were differentiated into either cDC or LC, and final yields of total cells were 29%⫾10% or 32%⫾6% of the original cell populations, respectively (Fig. 4 A and B). The phenotypical analysis showed that more than 95% of the cells were Mo-DC,
with a homogeneous population expressing high levels of HLA-DR and CD11c and low levels of the costimulatory mol-ecule CD86 (Fig. 5A and B). More than 87% of the cells were Mo-LC, with homogeneous expression of CLA, heterogeneous expression of HLA-DR, and low expression of CD86 and Lan-gerin (Fig. 5A and B). The transduction of nondividing mono-cytes with sc AAV vectors at day 0 did not significantly modify the yield of Mo-DC or Mo-LC (Fig. 4A and B) or their phe-notypes or maturation stages (Fig. 5A and B) at the end of the culture period. Furthermore, an upregulation of the CD86 costimulatory marker was observed when sc rAAV-transduced Mo-DC and Mo-LC were activated by LPS for 24 h, showing that their response to LPS was unaltered (Fig. 5C).
[image:5.585.72.514.69.345.2]CD34⫹-derived cells were transduced at day 0, 5, or 6 and analyzed at the end of the culture period (Fig. 1). Results are expressed as the index of proliferation instead of yields, since CD34⫹progenitors are proliferating cells. The proliferation index for total nontransduced cells in LC and CD34-pDC cultures were 8%⫾1% and 3.7%⫾1% (Fig. 4C and D). The phenotypical analysis of CD34-LC transduced at day 6 showed that more than 77% of cells were LC, with the expres-sion of HLA-DR and CLA and a low level of expresexpres-sion of CD86, and the phenotypical analysis of CD34-pDC transduced at day 5 showed that more than 30% of cells were pDC, with the expression of HLA-DR, a high level of expression of CD123, and homogeneous expression of CD86 (Fig. 5A and B). It should be noted that the proliferation index for
FIG. 4. Cell culture characterization of sc rAAV-transduced DC subpopulations. (A and B) Yields of total Mo-DC or Mo-LC that were nontransduced (NTd) or transduced at day 0 (Td d0) with sc rAAV1 or sc rAAV2 were measured at day 8. (C and D) The proliferation index of total CD34-LC that were nontransduced or transduced at day 0 or day 6 (Td d6) with sc rAAV1 or sc rAAV2 were determined at day 12. The proliferation index of total CD34-pDC that were nontransduced (NTd) or transduced at day 0 (Td d0) or day 5 (Td d5) with sc rAAV1 or sc rAAV2 were determined at day 10. Yields and proliferation index were determined by cell counting and flow cytometry analysis. Results are expressed as the mean percentage of cells or the index of cell proliferation⫾SD determined in the numbers of independent experiments indicated (n).
VOL. 81, 2007 TRANSDUCTION OF HUMAN DC WITH sc rAAV1 AND sc rAAV2 5389
on November 8, 2019 by guest
http://jvi.asm.org/
CD34-LC decreased slightly when cells were transduced with both sc rAAV serotypes at day 0 or at day 6 (Fig. 4C). The phenotypes and maturation states of LC and CD34-pDC were not significantly changed when transduced with ei-ther sc rAAV vector (Fig. 5A and B). As previously, we ob-served an upregulation of the CD86 costimulatory marker when transduced CD34-LC and CD34-pDC were activated by LPS and CpG, respectively (Fig. 5C). Transduction of DC at later time points in the differentiation pathways did not affect the yields or phenotypes of these cells (data not shown). Lastly, infection with ss rAAV vectors was poorly efficient and did not affect the yields and the phenotypes of the DC subpopulations (data not shown).
Altogether, our results indicate that sc rAAV1 and sc rAAV2 can efficiently transduce cells already committed to the
different DC differentiation pathways without inducing pheno-typic and maturation changes.
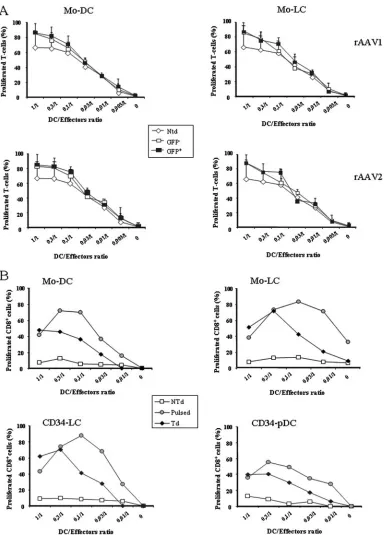
Functional properties of DC subpopulations transduced with sc rAAV1 and rAAV2.We evaluated the ability of trans-duced monocyte-derived cells to stimulate allogeneic T cells in MLRs. Monocytes transduced with sc rAAV1 or sc rAAV2 expressing E-GFP were differentiated into cDC or LC as pre-viously described, matured in LPS for 24 h, and then sorted by flow cytometry on the basis of E-GFP expression. Nontrans-duced, E-GFP-negative and -positive sorted cDC and LC were used to stimulate CFSE-labeled allogeneic T cells. As shown in Fig. 6A, the E-GFP-negative and -positive Mo-DC and Mo-LC displayed allostimulatory capacities comparable with those of nontransduced DC. The capacities of the transduced DC sub-populations to activate a CD8⫹-T-cell clone were then
evalu-FIG. 5. Immunophenotypes of sc rAAV-transduced DC subpopulations. Phenotypes of Mo-DC and Mo-LC at day 8, after transduction with sc rAAV at day 0, and phenotypes of CD34-LC and CD34-pDC at days 12 and 10, after transduction with sc rAAV at days 6 and 5, respectively. (A) Flow cytometry analysis was performed to monitor the expression of markers characteristic of cDC, LC, and pDC. Cells gated in forward scatter-side scatter parameters were analyzed by double staining for the expression of HLA-DR and CD11c for Mo-DC, CLA and Langerin for Mo-LC and CD34-LC, and HLA-DR and CD123 for CD34-pDC. Values indicate the percentages of cells in the corresponding quadrant. Overlay histograms show the expression of relevant antigens for nontransduced cells (thin black lines), total transduced cells (thick black lines), and E-GFP-positive gated cells (green lines) versus those for isotype-matched controls (dotted lines). (B) Comparative phenotypes of nontransduced and transduced DC subpopulations. Overlay histograms show the expression of the CD86 costimulatory molecule for nontransduced (thin black lines), total transduced (thick black lines), and E-GFP-positive gated (green lines) DC subpopulations versus those for isotype-matched controls (dotted lines). (C) Comparative phenotypes of transduced DC in the absence or presence of LPS or CpG for 24 h. Overlay histograms show the levels of expression of CD86 by immature (thin black lines) and total mature (thick black lines) and E-GFP-positive gated mature (green lines) transduced DC subpopulations versus those of isotype-matched controls (dotted lines). The results are representative of those from at least three experiments.
5390 VERON ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.43.546.69.402.2]FIG. 6. Functions of sc rAAV-transduced DC subpopulations. (A) The alloantigen presentation capacities of Mo-DC and Mo-LC were assessed 8 days posttransduction after maturation in LPS for 24 h and cell sorting on an E-GFP expression basis. Total nontransduced cells (Ntd) and E-GFP-negative and E-GFP-positive sorted Mo-DC (left graphs) and Mo-LC (right graphs), transduced with either sc rAAV1 (upper graphs) or sc rAAV2 (lower graphs), were incubated with allogeneic T cells stained with CFSE. After 5 days of coculture, percentages of CD8⫹dividing T cells measured by flow cytometry were linearly correlated with the loss of CFSE fluorescence. The data are shown as the means⫾SD of triplicate results and represent one out of three independent experiments. (B) In vitro antigen presentation capacities of sc rAAV1-transduced HLA-A2 DC subpopulations. Cells were transduced with sc rAAV1 expressing the MART-1 peptide, as depicted in Fig. 1, or pulsed for 3 h with the peptide the day of the coculture. Then mature nontransduced (NTd), transduced (Td), or peptide-pulsed (Pulsed) DC were cocultured with the HLA-A2-restricted CD8⫹-T-cell clone specific for the MART-1 peptide (LT12) stained with CFSE. After 5 days of coculture, percentages of CD8⫹ dividing T cells measured by flow cytometry were linearly correlated with the loss of CFSE fluorescence. The data are representative the results of one of two independent experiments.
VOL. 81, 2007 TRANSDUCTION OF HUMAN DC WITH sc rAAV1 AND sc rAAV2 5391
on November 8, 2019 by guest
http://jvi.asm.org/
ated. Nonproliferating HLA-A2⫹ monocytes transduced at day 0 were differentiated into DC or LC, whereas proliferating HLA-A2⫹CD34-LC or CD34-pDC were transduced at day 6 with sc rAAV1 expressing the MART-1 peptide. The different DC subpopulations obtained were efficient in activating a spe-cific CD8⫹-T-cell clone (Fig. 6B). The capacities of the differ-ent transduced DC subpopulations to secrete characteristic cytokines, such as IL-12, IL-10, and tumor necrosis factor al-pha for cDC and LC and alal-pha interferon for pDC, were then assessed. For the four subpopulations analyzed, no change in the secretion profiles compared to those of the nontransduced cells was seen (data not shown).
Altogether, these results indicate that the functional prop-erties of Mo-DC, Mo-LC, CD34-LC, and CD34-pDC are not altered by transduction with sc AAV1 and AAV2 vectors. Furthermore, all these transduced DC were able to activate a CD8⫹-T-cell clone.
DISCUSSION
DC are the professional APC of the immune system, fully equipped to initiate primary immune responses, including tolerogenic responses (3, 43, 44). For these reasons, DC are an attractive target for genetic manipulation of the immune sys-tem to increase otherwise insufficient immune responses in cases of infectious disease and cancer. Immunization with ex vivo-generated DC has proven feasible and permits the en-hancement as well as the dampening of antigen-specific im-mune responses in humans. Nevertheless, DC-based vaccines have yet to be improved since clinical responses are rarely complete and long lasting (33). Therefore, a future vaccine generated ex vivo will probably be heterogeneous and com-posed of several DC subsets, each presenting specific tar-geting to a given immune effector.
Direct DC transduction with antigen-expressing vectors of-fers potential advantages over regular peptide- or protein-loading protocols: (i) it ensures long-lasting expression of the antigen and production of an entire array of epitopes pre-sented by the autologous HLA molecules, and (ii) antigens are delivered to both endogenous major histocompatibility com-plex class I and class II antigen presentation pathways (2, 23). In this study, we investigated the potential of AAV1 and AAV2 vectors, with the conventional ss genome or sc genome, to transduce human Mo-DC, Mo-LC, LC, and CD34-pDC. We showed that a single exposure of monocytes or CD34⫹progenitors to sc rAAV1 or sc rAAV2 results in high levels of transduced Mo-DC, Mo-LC, LC, or CD34-pDC (5 to 59%) without impacting the phenotypes, functions, and abilities of these cells to respond to inflammatory signals. We previously showed that cells committed toward LC, but not DC, can be efficiently transduced with LV without any facili-tating agents such as Polybrene (48). Here, we showed that not only LC but also cDC and pDC can be efficiently transduced with sc AAV1 and sc AAV2 vectors after commitment to their differentiation pathways, offering a great advantage over trans-duction with LV. Furthermore, it appears clear that even though most rAAV genomes remain episomal (11, 31, 41), they permit long-term persistence and stable transgene expres-sion (12, 25, 56) in nondividing cells. Gene delivery to human DC has also been achieved with other viral vectors such as
adenovirus- and vaccinia virus-derived vectors, but the appli-cability of these vectors in vivo is rather limited since most humans have preexisting immunity to these viruses (24). The preexistence of humoral immunity to some wild-type AAV in humans has been described previously (10, 16). Nevertheless, rAAV may overcome these limitations due to the development of alternative serotypes which are derived from a variety of human and nonhuman wild-type AAVs (18, 38).
We further showed that 8-day posttransduction Mo-DC and Mo-LC transduced at day 0 of the induction of differentiation and 12-day posttransduction CD34-LC and CD34-pDC trans-duced at day 5 or 6 with sc rAAV1 were potent activators of a CD8⫹-T-cell clone specific for the tumor antigen MART-1. Our results showed that sc rAAV1 and sc rAAV2 are efficient tools for ex vivo DC-based immunotherapy. Furthermore, these ex vivo evaluations suggest that these vectors may also be good candidates for the in vivo targeting of specific DC subsets. This finding would be an important step toward the develop-ment of a DC-based immunotherapy where the induction of a given type of immune response is required. For this purpose, the identification of promoters driving the expression of spe-cific DC subset cell surface molecules, such as DC-spespe-cific intercellular adhesion molecule-3-grabbing nonintegrin ex-pressed by myeloid cDC (20), Langerin exex-pressed by LC (45, 46), and blood DC antigen 2 expressed by pDC (14, 15), will allow specific targeting of each of these DC subsets. One of the limitations of sc rAAV is the limited packaging capacity, which cannot exceed 2.3 kb. However, given the fact that for immu-nization purposes, only a fraction of the coding sequence can be used to generate an immunoreactive epitope, this restricted of the expression cassette should not be a limitation.
Other reports have described the efficient transduction of human Mo-DC with ss rAAV2, but large numbers of vectors or repeated exposures to the vectors were necessary (9, 27, 34). We found here that monocytes, CD34⫹progenitors, and cells committed to differentiation into one of the DC subsets were very poorly transduced by using a single exposure to ss rAAV1 and ss rAAV2 (less than 9%), suggesting, among other expla-nations such as tropism, that the second-strand synthesis may be a limiting step for transgene expression in human DC sub-sets. Several studies have reported that the potential benefit of sc rAAV is the achievement of earlier and higher-level trans-gene expression in various cell types, such as muscle, brain, and liver cells, related to a bypass of the complementary strand synthesis (1, 17, 30, 49). In our experiments, we did not observe significant transduction levels with ss rAAV1 and ss rAAV2 even when a longer time for expression was allowed (data not shown). Our results clearly demonstrate that the major step limiting DC transduction with AAV vectors is the conversion of the single-stranded viral genome.
To date, gene therapy clinical trials performed to correct monogenic diseases such as cystic fibrosis and hemophilia B have been based on conventional ss AAV vectors (4). Results have shown the safety of these vectors but also have been somewhat disappointing in terms of efficacy. Furthermore, the immune response against the AAV capsid imposes some lim-itations (29). The use of sc AAV vectors represents an oppor-tunity to increase gene transfer efficiency by providing tran-scriptionally active recombinant viral genomes to the cell. However, using sc rAAV for in vivo gene therapy applications
5392 VERON ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
may increase the risk of immune responses against the trans-gene product through the transduction of DC. To counteract this highly unwanted consequence, the use of promoters spe-cific to the targeted cells will be an absolute requirement.
Altogether, we show here for the first time that sc AAV type 1 and type 2 vectors allow for the efficient transduction of monocytes, CD34 progenitors, and their derivative DC subsets, before and after commitment to their differentiation pathways, without alteration of their phenotypes and functional matura-tion capabilities. Furthermore, sc rAAV-transduced DC sub-sets are able to present a specific peptide and to activate a CD8⫹-T-cell clone, demonstrating the potential use of this approach for a DC based-immunotherapy aiming at inducing active immunization or peripheral tolerance. Finally, these re-sults also suggest that sc AAV vectors can be of high value for in vivo strategies in which a specific DC subset must be tar-geted.
ACKNOWLEDGMENTS
P.V. is supported by a CIFRE convention from Association Nationale de la Recherche Technique. This work was supported by the Association Franc¸aise contre les Myopathies (AFM).
We thank Vincent Zuliani and Isabelle Lambert for providing viral vectors. We thank Florence Faure for giving us the HLA-A2-restricted MART-1-specific CD8⫹-T-cell clone. We thank Susan Cure for the critical reading of the manuscript.
REFERENCES
1.Aldrich, W. A., C. Ren, A. F. White, S. Z. Zhou, S. Kumar, C. B. Jenkins, D. R. Shaw, T. V. Strong, P. L. Triozzi, and S. Ponnazhagan.2006. Enhanced transduction of mouse bone marrow-derived dendritic cells by repetitive infection with self-complementary adeno-associated virus 6 combined with
immunostimulatory ligands. Gene Ther.13:29–39.
2.Banchereau, J., F. Briere, C. Caux, J. Davoust, S. Lebecque, Y. J. Liu, B. Pulendran, and K. Palucka.2000. Immunobiology of dendritic cells. Annu.
Rev. Immunol.18:767–811.
3.Banchereau, J., and R. M. Steinman.1998. Dendritic cells and the control of
immunity. Nature392:245–252.
4.Carter, B. J.2005. Adeno-associated virus vectors in clinical trials. Hum.
Gene Ther.16:541–550.
5.Caux, C., C. Dezutter-Dambuyant, D. Schmitt, and J. Banchereau.1992. GM-CSF and TNF-alpha cooperate in the generation of dendritic
Langer-hans cells. Nature360:258–261.
6.Caux, C., C. Massacrier, B. Dubois, J. Valladeau, C. Dezutter-Dambuyant, I. Durand, D. Schmitt, and S. Saeland.1999. Respective involvement of TGF-beta and IL-4 in the development of Langerhans cells and
non-Langer-hans dendritic cells from CD34⫹progenitors. J. Leukoc. Biol.66:781–791.
7.Caux, C., B. Vanbervliet, C. Massacrier, C. Dezutter-Dambuyant, B. de Saint-Vis, C. Jacquet, K. Yoneda, S. Imamura, D. Schmitt, and J. Banchereau.
1996. CD34⫹hematopoietic progenitors from human cord blood differentiate
along two independent dendritic cell pathways in response to GM-CSF⫹TNF
alpha. J. Exp. Med.184:695–706.
8.Chapuis, F., M. Rosenzwajg, M. Yagello, M. Ekman, P. Biberfeld, and J. C. Gluckman.1997. Differentiation of human dendritic cells from monocytes in
vitro. Eur. J. Immunol.27:431–441.
9.Chiriva-Internati, M., Y. Liu, E. Salati, W. Zhou, Z. Wang, F. Grizzi, J. J. Roman, S. H. Lim, and P. L. Hermonat.2002. Efficient generation of cyto-toxic T lymphocytes against cervical cancer cells by adeno-associated virus/ human papillomavirus type 16 E7 antigen gene transduction into dendritic
cells. Eur. J. Immunol.32:30–38.
10.Chirmule, N., K. Propert, S. Magosin, Y. Qian, R. Qian, and J. Wilson.1999. Immune responses to adenovirus and adeno-associated virus in humans.
Gene Ther.6:1574–1583.
11.Duan, D., P. Sharma, J. Yang, Y. Yue, L. Dudus, Y. Zhang, K. J. Fisher, and J. F. Engelhardt.1998. Circular intermediates of recombinant adeno-asso-ciated virus have defined structural characteristics responsible for long-term
episomal persistence in muscle tissue. J. Virol.72:8568–8577.
12.Duan, D., Z. Yan, Y. Yue, and J. F. Engelhardt.1999. Structural analysis of
adeno-associated virus transduction circular intermediates. Virology261:8–14.
13.Dullaers, M., and K. Thielemans.2006. From pathogen to medicine: HIV-1-derived lentiviral vectors as vehicles for dendritic cell based cancer
immu-notherapy. J. Gene Med.8:3–17.
14.Dzionek, A., A. Fuchs, P. Schmidt, S. Cremer, M. Zysk, S. Miltenyi, D. W.
Buck, and J. Schmitz.2000. BDCA-2, BDCA-3, and BDCA-4: three markers for distinct subsets of dendritic cells in human peripheral blood. J. Immunol.
165:6037–6046.
15.Dzionek, A., Y. Sohma, J. Nagafune, M. Cella, M. Colonna, F. Facchetti, G. Gunther, I. Johnston, A. Lanzavecchia, T. Nagasaka, T. Okada, W. Vermi, G. Winkels, T. Yamamoto, M. Zysk, Y. Yamaguchi, and J. Schmitz.2001. BDCA-2, a novel plasmacytoid dendritic cell-specific type II C-type lectin, mediates antigen capture and is a potent inhibitor of interferon alpha/beta
induction. J. Exp. Med.194:1823–1834.
16.Erles, K., P. Sebokova, and J. R. Schlehofer.1999. Update on the prevalence of serum antibodies (IgG and IgM) to adeno-associated virus (AAV).
J. Med. Virol.59:406–411.
17.Fu, H., J. Muenzer, R. J. Samulski, G. Breese, J. Sifford, X. Zeng, and D. M. McCarty.2003. Self-complementary adeno-associated virus serotype 2 vec-tor: global distribution and broad dispersion of AAV-mediated transgene
expression in mouse brain. Mol. Ther.8:911–917.
18.Gao, G., L. H. Vandenberghe, and J. M. Wilson.2005. New recombinant
serotypes of AAV vectors. Curr. Gene Ther.5:285–297.
19.Gatti, E., M. A. Velleca, B. C. Biedermann, W. Ma, J. Unternaehrer, M. W. Ebersold, R. Medzhitov, J. S. Pober, and I. Mellman.2000. Large-scale culture and selective maturation of human Langerhans cells from
granulo-cyte colony-stimulating factor-mobilized CD34⫹progenitors. J. Immunol.
164:3600–3607.
20.Geijtenbeek, T. B., R. Torensma, S. J. van Vliet, G. C. van Duijnhoven, G. J. Adema, Y. van Kooyk, and C. G. Figdor.2000. Identification of DC-SIGN, a novel dendritic cell-specific ICAM-3 receptor that supports primary immune
responses. Cell100:575–585.
21.Geissmann, F., C. Prost, J. P. Monnet, M. Dy, N. Brousse, and O. Hermine.
1998. Transforming growth factor beta1, in the presence of granulocyte/ macrophage colony-stimulating factor and interleukin 4, induces differenti-ation of human peripheral blood monocytes into dendritic Langerhans cells.
J. Exp. Med.187:961–966.
22.Herbst, B., G. Kohler, A. Mackensen, H. Veelken, P. Kulmburg, F. M. Rosenthal, H. E. Schaefer, R. Mertelsmann, P. Fisch, and A. Lindemann.
1996. In vitro differentiation of CD34⫹hematopoietic progenitor cells
to-ward distinct dendritic cell subsets of the Birbeck granule and MIIC-positive
Langerhans cell and the interdigitating dendritic cell type. Blood88:2541–
2548.
23.Jaraquemada, D., M. Marti, and E. O. Long.1990. An endogenous process-ing pathway in vaccinia virus-infected cells for presentation of cytoplasmic
antigens to class II-restricted T cells. J. Exp. Med.172:947–954.
24.Jenne, L., G. Schuler, and A. Steinkasserer.2001. Viral vectors for dendritic
cell-based immunotherapy. Trends Immunol.22:102–107.
25.Jiang, H., G. F. Pierce, M. C. Ozelo, E. V. de Paula, J. A. Vargas, P. Smith, J. Sommer, A. Luk, C. S. Manno, K. A. High, and V. R. Arruda.2006. Evidence of multiyear factor IX expression by AAV-mediated gene transfer to skeletal muscle in an individual with severe hemophilia B. Mol. Ther.
14:452–455.
26.Jooss, K., Y. Yang, K. J. Fisher, and J. M. Wilson.1998. Transduction of dendritic cells by DNA viral vectors directs the immune response to
trans-gene products in muscle fibers. J. Virol.72:4212–4223.
27.Liu, Y., M. Chiriva-Internati, F. Grizzi, E. Salati, J. J. Roman, S. Lim, and P. L. Hermonat.2001. Rapid induction of cytotoxic T-cell response against cervical cancer cells by human papillomavirus type 16 E6 antigen gene delivery into human dendritic cells by an adeno-associated virus vector.
Cancer Gene Ther.8:948–957.
28.Mahnke, K., E. Schmitt, L. Bonifaz, A. H. Enk, and H. Jonuleit.2002. Immature, but not inactive: the tolerogenic function of immature dendritic
cells. Immunol. Cell Biol.80:477–483.
29.Manno, C. S., G. F. Pierce, V. R. Arruda, B. Glader, M. Ragni, J. J. Rasko, M. C. Ozelo, K. Hoots, P. Blatt, B. Konkle, M. Dake, R. Kaye, M. Razavi, A. Zajko, J. Zehnder, P. K. Rustagi, H. Nakai, A. Chew, D. Leonard, J. F. Wright, R. R. Lessard, J. M. Sommer, M. Tigges, D. Sabatino, A. Luk, H. Jiang, F. Mingozzi, L. Couto, H. C. Ertl, K. A. High, and M. A. Kay.2006. Successful transduction of liver in hemophilia by AAV-factor IX and
limi-tations imposed by the host immune response. Nat. Med.12:342–347.
30.McCarty, D. M., H. Fu, P. E. Monahan, C. E. Toulson, P. Naik, and R. J. Samulski.2003. Adeno-associated virus terminal repeat (TR) mutant gen-erates self-complementary vectors to overcome the rate-limiting step to
transduction in vivo. Gene Ther.10:2112–2118.
31.Nakai, H., S. R. Yant, T. A. Storm, S. Fuess, L. Meuse, and M. A. Kay.2001. Extrachromosomal recombinant adeno-associated virus vector genomes are
primarily responsible for stable liver transduction in vivo. J. Virol.75:6969–
6976.
32.Olivier, A., E. Lauret, P. Gonin, and A. Galy.2006. The Notch ligand delta-1 is a hematopoietic development cofactor for plasmacytoid
den-dritic cells. Blood107:2694–2701.
33.O’Neill, D. W., S. Adams, and N. Bhardwaj.2004. Manipulating dendritic
cell biology for the active immunotherapy of cancer. Blood104:2235–2246.
34.Ponnazhagan, S., G. Mahendra, D. T. Curiel, and D. R. Shaw.2001. Adeno-associated virus type 2-mediated transduction of human monocyte-derived
VOL. 81, 2007 TRANSDUCTION OF HUMAN DC WITH sc rAAV1 AND sc rAAV2 5393
on November 8, 2019 by guest
http://jvi.asm.org/
dendritic cells: implications for ex vivo immunotherapy. J. Virol.75:9493– 9501.
35.Pulendran, B.2004. Modulating vaccine responses with dendritic cells and
Toll-like receptors. Immunol. Rev.199:227–250.
36.Pulendran, B.2005. Variegation of the immune response with dendritic cells
and pathogen recognition receptors. J. Immunol.174:2457–2465.
37.Reid, C. D., A. Stackpoole, A. Meager, and J. Tikerpae.1992. Interactions of tumor necrosis factor with granulocyte-macrophage colony-stimulating fac-tor and other cytokines in the regulation of dendritic cell growth in vitro
from early bipotent CD34⫹progenitors in human bone marrow. J. Immunol.
149:2681–2688.
38.Riviere, C., O. Danos, and A. M. Douar.2006. Long-term expression and repeated administration of AAV type 1, 2 and 5 vectors in skeletal muscle of
immunocompetent adult mice. Gene Ther.13:1300–1308.
39.Romani, N., S. Gruner, D. Brang, E. Kampgen, A. Lenz, B. Trockenbacher, G. Konwalinka, P. O. Fritsch, R. M. Steinman, and G. Schuler.1994.
Pro-liferating dendritic cell progenitors in human blood. J. Exp. Med.180:83–93.
40.Rouas, R., R. Uch, Y. Cleuter, F. Jordier, C. Bagnis, P. Mannoni, P. Lewalle, P. Martiat, and A. Van den Broeke.2002. Lentiviral-mediated gene delivery in human monocyte-derived dendritic cells: optimized design and procedures for highly efficient transduction compatible with clinical constraints. Cancer
Gene Ther.9:715–724.
41.Schnepp, B. C., K. R. Clark, D. L. Klemanski, C. A. Pacak, and P. R. Johnson.2003. Genetic fate of recombinant adeno-associated virus vector
genomes in muscle. J. Virol.77:3495–3504.
42.Schroers, R., I. Sinha, H. Segall, I. G. Schmidt-Wolf, C. M. Rooney, M. K. Brenner, R. E. Sutton, and S. Y. Chen.2000. Transduction of human PBMC-derived dendritic cells and macrophages by an HIV-1-based lentiviral vector
system. Mol. Ther.1:171–179.
43.Steinman, R. M.1991. The dendritic cell system and its role in
immunoge-nicity. Annu. Rev. Immunol.9:271–296.
44.Steinman, R. M., D. Hawiger, and M. C. Nussenzweig.2003. Tolerogenic
dendritic cells. Annu. Rev. Immunol.21:685–711.
45.Valladeau, J., V. Duvert-Frances, J. J. Pin, C. Dezutter-Dambuyant, C. Vincent, C. Massacrier, J. Vincent, K. Yoneda, J. Banchereau, C. Caux, J. Davoust, and S. Saeland.1999. The monoclonal antibody DCGM4 recog-nizes Langerin, a protein specific of Langerhans cells, and is rapidly
inter-nalized from the cell surface. Eur. J. Immunol.29:2695–2704.
46.Valladeau, J., O. Ravel, C. Dezutter-Dambuyant, K. Moore, M. Kleijmeer, Y.
Liu, V. Duvert-Frances, C. Vincent, D. Schmitt, J. Davoust, C. Caux, S. Lebecque, and S. Saeland.2000. Langerin, a novel C-type lectin specific to Langerhans cells, is an endocytic receptor that induces the formation of
Birbeck granules. Immunity12:71–81.
47.Valmori, D., J. F. Fonteneau, C. M. Lizana, N. Gervois, D. Lienard, D. Rimoldi, V. Jongeneel, F. Jotereau, J. C. Cerottini, and P. Romero.1998. Enhanced generation of specific tumor-reactive CTL in vitro by selected
Melan-A/MART-1 immunodominant peptide analogues. J. Immunol.160:
1750–1758.
48.Veron, P., S. Boutin, J. Bernard, O. Danos, J. Davoust, and C. Masurier.
2006. Efficient transduction of monocyte- and CD34(⫹)-derived Langerhans
cells with lentiviral vectors in the absence of phenotypic and functional
maturation. J. Gene Med.8:951–961.
49.Wang, Z., H. I. Ma, J. Li, L. Sun, J. Zhang, and X. Xiao.2003. Rapid and highly efficient transduction by double-stranded adeno-associated virus
vec-tors in vitro and in vivo. Gene Ther.10:2105–2111.
50.Warrington, K. H., Jr., and R. W. Herzog.2006. Treatment of human disease
by adeno-associated viral gene transfer. Hum. Genet.119:571–603.
51.Wu, Z., A. Asokan, and R. J. Samulski.2006. Adeno-associated virus
sero-types: vector toolkit for human gene therapy. Mol. Ther.14:316–327.
52.Xiao, W., N. Chirmule, M. A. Schnell, J. Tazelaar, J. V. Hughes, and J. M. Wilson.2000. Route of administration determines induction of T-cell-inde-pendent humoral responses to adeno-associated virus vectors. Mol. Ther.
1:323–329.
53.Xiao, X., J. Li, and R. J. Samulski.1998. Production of high-titer recombi-nant adeno-associated virus vectors in the absence of helper adenovirus.
J. Virol.72:2224–2232.
54.Xin, K. Q., H. Mizukami, M. Urabe, Y. Toda, K. Shinoda, A. Yoshida, K. Oomura, Y. Kojima, M. Ichino, D. Klinman, K. Ozawa, and K. Okuda.2006. Induction of robust immune responses against human immunodeficiency virus is supported by the inherent tropism of adeno-associated virus type 5
for dendritic cells. J. Virol.80:11899–11910.
55.Yang, Y., Q. Su, and J. M. Wilson.1996. Role of viral antigens in destructive cellular immune responses to adenovirus vector-transduced cells in mouse
lungs. J. Virol.70:7209–7212.
56.Yue, Y., and D. Duan.2003. Double strand interaction is the predominant pathway for intermolecular recombination of adeno-associated viral
ge-nomes. Virology313:1–7.
5394 VERON ET AL. J. VIROL.