0022-538X/06/$08.00⫹0 doi:10.1128/JVI.02094-05
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Sequential Immunization with V3 Peptides from Primary Human
Immunodeficiency Virus Type 1 Produces Cross-Neutralizing
Antibodies against Primary Isolates with a Matching
Narrow-Neutralization Sequence Motif
Yasuyuki Eda,
1Mari Takizawa,
2Toshio Murakami,
1Hiroaki Maeda,
1Kazuhiko Kimachi,
1Hiroshi Yonemura,
1Satoshi Koyanagi,
1Kouichi Shiosaki,
1Hirofumi Higuchi,
1Keiichi Makizumi,
1Toshihiro Nakashima,
1Kiyoshi Osatomi,
1,3Sachio Tokiyoshi,
1Shuzo Matsushita,
4Naoki Yamamoto,
2and Mitsuo Honda
2*
The Chemo-Sero-Therapeutic Research Institute, Kyokushi, Kikuchi, Kumamoto 869-1298, Japan1; AIDS Research Center,
National Institute of Infectious Diseases, Shinjuku-ku, Tokyo 162-8640, Japan2; Nagasaki University,
Nagasaki 852-8521, Japan3; and Center for AIDS Research, Kumamoto University,
Kumamoto 860-0811, Japan4
Received 5 October 2005/Accepted 9 March 2006
An antibody response capable of neutralizing not only homologous but also heterologous forms of the CXCR4-tropic human immunodeficiency virus type 1 (HIV-1) MNp and CCR5-tropic primary isolate HIV-1 JR-CSF was achieved through sequential immunization with a combination of synthetic peptides representing HIV-1 Env V3 sequences from field and laboratory HIV-1 clade B isolates. In contrast, repeated immunization with a single V3 peptide generated antibodies that neutralized only type-specific laboratory-adapted homologous viruses. To determine whether the cross-neutralization response could be attributed to a cross-reactive antibody in the immunized animals, we isolated a monoclonal antibody, C25, which neutralized the heterologous primary viruses of HIV-1 clade B. Furthermore, we generated a humanized monoclonal antibody, KD-247, by transferring the genes of the complementary determining region of C25 into genes of the human V region of the antibody. KD-247 bound with high affinity to the “PGR” motif within the HIV-1 Env V3 tip region, and, among the established reference antibodies, it most effectively neutralized primary HIV-1 field isolates possessing the matching neutralization sequence motif, suggesting its promise for clinical applications involving passive immunizations. These results demon-strate that sequential immunization with B-cell epitope peptides may contribute to a humoral immune-based HIV vaccine strategy. Indeed, they help lay the groundwork for the development of HIV-1 vaccine strategies that use sequential immunization with biologically relevant peptides to overcome difficulties associated with otherwise poorly immunogenic epitopes.
In humans, antibodies, whether actively induced or passively transferred, neutralize viruses and therefore protect against viral diseases like hepatitis and influenza (6, 15). However, the specific antibodies that confer protective immunity against hu-man immunodeficiency virus type 1 (HIV-1) infection are not well known, since most primary strains of HIV-1 are relatively resistant to neutralization (40, 47). Studies with recombinant monomeric gp120 have not been successful at predicting the neutralization of primary isolates (12, 39). However, consider-able progress in understanding HIV pathogenesis, namely, in determining that both antibody- and cell-mediated immune responses are likely responsible for controlling viral load, has recently been made (10, 44, 60). With regard to the role of neutralizing antibody responses in HIV-1 infection, broadly reactive neutralizing antibodies such as 2G12 (54), 2F5 (45), and 4E10 (5) have been proven to suppress immune deficiency virus infection in macaques (14, 38) and humans (62).
How-ever, it remains to be seen whether high-titered and cross-reactive neutralizing antibodies will be produced by active im-munization with a novel viral antigen.
In this study, we attempted to develop a sequential immu-nization strategy that has proven to be effective at eliciting neutralization antibodies to primary HIV-1 by targeting the HIV-1 Env V3 neutralization epitope site as a model antigen. Previously, we demonstrated that the Gly-Pro-Gly-Arg (GPGR) core sequence at the tip of the V3 region of gp120 is relatively conserved in both field and clinical isolates of HIV-1 clade B, while the flanking regions of the V3 tip are more variable (1, 66). While in theory, high-affinity antibodies that recognize the relatively conserved GPGR epitope could potentially neutral-ize many strains of HIV-1 clade B, in practice, such antibodies in sera from HIV-infected individuals show little neutralization activity in vitro (2), suggesting that the immunogenicity of the GPGR sequence is similarly low. To overcome this problem, we sequentially immunized mice with V3 peptides from HIV-1 clade B field isolates, resulting in the induction of cross-reac-tive antisera that strongly bound to V3 peptides from homol-ogous and heterolhomol-ogous primary isolates. Furthermore, a cross-reactive neutralizing monoclonal antibody (MAb), C25,
* Corresponding author. Mailing address: AIDS Research Center, National Institute of Infectious Diseases, Shinjuku-ku, Tokyo 162-8640, Japan. Phone: 81-3-5285-1111, ext. 2737. Fax: 81-3-5285-1183. E-mail: [email protected].
5552
on November 8, 2019 by guest
http://jvi.asm.org/
was established. KD-247, a reshaped MAb derived from a C25 gene that had been reshaped to a humanized antibody, also efficiently neutralized primary isolates of HIV-1.
Although anti-V3 antibodies elicited by active immunization with the V3 peptide and HIV Env have been reported to neutralize HIV-1 both in cell culture and in animal challenge studies, these antibodies have not yet been fully exploited be-cause they are type specific (13, 29, 33, 46). In contrast, anti-V3 MAbs generated by heterohybridoma (22) using peripheral blood mononuclear cells (PBMCs) from HIV-infected individ-uals have recently been shown to contain cross-neutralizing anti-V3 MAbs and neutralize primary isolates (11, 18, 19, 21, 23, 25). Furthermore, it has been suggested that the neutral-ization sensitivity of primary isolates is regulated by the V1/V2 domain of Env gp120 (48). In this study, we demonstrate that cross-neutralizing anti-V3 antibodies against primary isolates can be produced via sequential immunization with the V3-neutralizing epitope peptides of HIV-1. Furthermore, we dis-cuss how sequential immunization with Env peptides including a neutralization epitope could pave the way for the generation of cross-reactive neutralization antibodies.
MATERIALS AND METHODS
Animals. C3H/HeN mice were bred at The Chemo-Sero-Therapeutic Re-search Institute Experimental Animal Center and used when they were between 4 and 8 weeks of age for immunization and generation of MAbs. The study was conducted with the approval of an institutional committee for biosafety and animal welfare. The mice were housed in accordance with the Guidelines for Animal Experimentation of the Japanese Association for Laboratory Animal Science under the Japanese Law Concerning the Protection and Management of Animals (59) and were maintained in accordance with the guidelines set forth by the Institutional Animal Care and Use Committee of Kaketsuken, Japan.
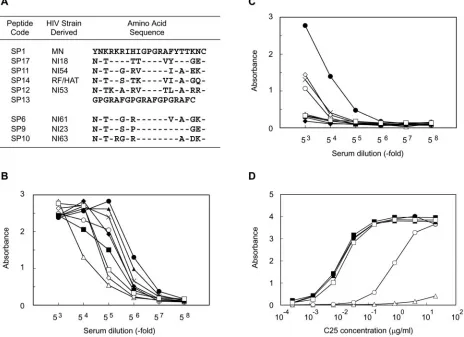
[image:2.585.61.525.75.412.2]Isolation of MAb C25 and sequencing of the V region gene.V3 sequences from HIV-1 clade B isolates were determined as previously described (50) and used to generate peptides SP1, SP17, SP11, SP14, SP12, and SP13 for sequential immu-nization of mice (Fig. 1A). Of the six V3 peptides generated, only the SP1 peptide was likewise repeatedly administered to another group of mice as a reference for the sequential immunization. A cysteine residue was added to each peptide for coupling to the keyhole limpet hemocyanin, and the peptides were then emulsified with complete and incomplete Freund’s adjuvant at a concen-tration of 100g/ml. Peptides were administered at 1-week intervals by intra-peritoneal injection, with the exception of SP13, which was injected intravenously 1 week after the SP12 inoculation. Hybridomas were generated from mice with antisera that are highly cross-reactive to HIV-1, as described previously (32), and clone C25 was selected by a combination of enzyme-linked immunosorbent assays (ELISAs) and neutralization assays (26). RNA was extracted from the C25 hybridoma by conventional methods and used as a template for the synthesis of FIG. 1. Generation of cross-reactive antibodies by sequential immunization with different V3 peptides originating from HIV-1 clade B field isolates. (A) Aligned V3 sequences used for sequential immunization and ELISA screening. (B) Binding antibody titers of antisera from an animal sequentially immunized with the V3 peptides. (C) Binding antibody titers of antisera from animals immunized six times with SP1 alone. (D) Binding antibody titers of an isolated C25 MAb against various V3 peptides. Binding antibody titers were similarly determined by monitoring the antibody reactivity in a peptide-based ELISA. The coated peptides were SP1 (F), SP17 (E), SP11 (Œ), SP14 (‚), SP12 (■), SP13 (䊐), SP6 (}), SP9 (〫), and SP10 (⫻).
on November 8, 2019 by guest
http://jvi.asm.org/
first-strand cDNA. Primers used for PCR amplification of the 5⬘mouse immu-noglobulin (Ig) V regions and the 3⬘J region were designed based on the nucleotide sequence database for mouse Ig as classified previously by Kabat et al. (30). The PCR primers used to obtain the mouse heavy-chain Ig gene were MHL4ver.4 (5⬘-TTCGAAGCTTGCCGCCACCATGGAATGGAGCTGGGTC TTT-3⬘) and MHCver.1 (5⬘-GGATCCCGGGAGTGGATAGACCGATG-3⬘), and the primers used to obtain the mouse light-chain Ig gene were ML7ver.2 (5⬘-ACTAGTCGACATGGGCATCAAGATGGAGT CACAGATTCTGG-3⬘) and MCver.1 (5⬘-GGATCCCGGGTGGATGGTGGGAAGATG-3⬘). Restric-tion enzyme sites for HindIII (V region) and BamHI (J region) were incorpo-rated into the primers to facilitate cloning. PCR amplification was performed using cDNA as a template and the V and J region primers, as described above. The resulting PCR products were then cloned into the HincII site of pUC18, and the sequence of the inserted V region gene was confirmed by the Sanger dideoxy-mediated chain termination method.
Preparation of humanized MAb KD-247.Transfer of the complementary de-termining regions (CDRs) and partial framework regions (FRs) of C25 into the human V regions was carried out according to established methods for preparing a humanized MAb. In brief, CDRs and a part of the FRs of the VHregion of C25 were transferred into the VHFR of human subgroup I, while CDRs of the VL region of C25 were transferred into the FR of the human-chain REI. CDR grafting was performed by PCR-based in vitro mutagenesis. Computer-aided molecular modeling was used to design the initial CDR-grafted and resurfaced versions of the constructs, which were then cloned into a glutamine synthetase expression vector (GS system; Lonza Biologics PLC, Berkshire, United King-dom). The vector was transfected into NS0 myeloma cells (Lonza), and glu-tamine-independent transfectants were isolated. Cells producing KD-247 were expanded in large-scale cultures, and the antibody was purified from the culture supernatants by ion-exchange and affinity chromatography.
Surface plasmon resonance.The kinetic constants for the binding of MAbs to the SP1 peptide were determined by surface plasmon resonance measurements (31) using a BIAcore 1000 system (BIAcore AB, Uppsala, Sweden). The SP1 peptide was coupled with biotin-PE-maleimide (Dojindo Laboratories, Kum-amoto, Japan) through a thiol-containing cysteine residue that was deliberately placed at the carboxyl terminus. Biotinylated SP1 was then coupled to strepta-vidin on the surface of a sensor chip (Sensor Chip SA; BIAcore AB, Uppsala, Sweden) by injection of 10l of a 100-ng/ml solution in HBS-EP buffer (BIA-core). Approximately 5 resonance units of SP1, a value recommended by the manufacturer, were immobilized on the chip. MAbs KD-247, R5.5, and C1 (41) were diluted in HBS-EP buffer to concentrations of 40, 60, 80, 100, and 160 nM, and 20l of each MAb was injected over the immobilized antigen at a flow rate of 20l/min. At the end of the association phase, the dissociation of each MAb was measured over an 8-min period at the same flow rate. The surface of the chip was regenerated with 20l of 10 mM HCl. Kinetic rate constants were calculated from data generated at each concentration of MAb using BIAevalu-ation version 3.0 software (BIAcore). The model for simultaneouska/kd deter-mination was selected for bivalent analysis because the samples were regarded as intact antibodies.
Pepscan analysis.The following peptides were synthesized by an Epitope Scanning kit (Chiron Mimotopes Pty., Ltd., Victoria, Australia) according to the Pepscan method (17) and examined for binding activity to KD-247: IHIGPGR AFY, IRVGPGRTLY, IRVGPGRAIY, and LSVGPGRSFY (corresponding to the V3 tip of HIV-1 strains MN, NI53, NI54, and TM2, respectively). We also determined which peptides had consecutive amino acids deleted, which had an overlapping series of peptides of 4 to 10 amino acids in length derived from the HIV-1MNV3 tip sequence IHIGPGRAFY and which had each of their amino acids consecutively replaced by one of the 19 naturally occurring amino acids. The reactivity of KD-247 with bound peptides was determined by a peptide-based ELISA. Polyethylene rods with bound synthetic peptides were precoated with 2% bovine serum albumin–0.1% Tween 20 in phosphate-buffered saline for 1 h at room temperature, followed by a reaction with 2g/ml of KD-247 in the precoated buffer overnight at 4°C. After the reaction of KD-247 with a peroxi-dase-labeled goat anti-human kappa antibody (Southern Biotechnology Associ-ates, Birmingham, AL), the presence of the MAb was detected by reaction with peroxidase substrate, and the resultant color was read at an optical density at 450 nm.
In vitro virus neutralization assay.The international reference antibodies 1006-15D, 694/98D, 1331-D, and 1367-D were kindly provided by Susan Zolla-Pazner, New York University School of Medicine, New York, NY, and 447-52D was obtained from the AIDS Research and Reference Reagent Program, Na-tional Institutes of Health, Rockville, MD (20, 21, 42). R5.5 and C1 (41) were also used as reference antibodies. An MT-4 cell-based virus neutralization assay was performed to screen for hybridomas and to evaluate MAbs as described
here. Briefly, serial dilutions of test samples were plated onto eight wells of a microtiter plate. HIV-1 (100 50% tissue culture infective doses [TCID50s]) was added to each well and incubated at 37°C in a CO2incubator for 1 h. One hundred microliters of MT-4 cell suspension (1⫻105cells/ml) was then added to each well and cultured at 37°C in a CO2incubator for 5 days without agitation. After cultivation of MT-4 cells for 5 days, the cells were dissociated in each well, and the presence or absence of syncytium was examined under a light micro-scope. Neutralizing activity (90% inhibitory concentration [IC90]) was defined as the minimum effective concentration of antibody that inhibited infection in all wells at the same concentration. The neutralization assay using the MT-4 cell line was conducted for the screening of hybridomas and the evaluation of MAbs against laboratory and primary isolates. Laboratory-adapted HIV-1MN (H9/ HTLV-III MN) and HIV-1IIIB(H9/HTLV-IIIB NIH1983) were obtained from the AIDS Research and Reference Reagent Program. Virus stocks of primary HIV-1 clinical isolates were prepared by coculturing phytohemagglutinin (PHA)-activated human PBMCs from both HIV-seropositive and healthy individuals, as described previously (11, 28).
GHOST cell neutralization assays were performed to study the neutralization activity of antibodies against HIV-1 as previously described (7, 58). Briefly, both GHOST-X4 and -R5 cells, which express the CXCR4 and CCR5 coreceptors, respectively, were used as target cells for viral infection. Cells used in neutral-ization assays were analyzed by FACSCalibur flow cytometry (Becton Dickinson, San Jose, CA), and 15,000 events were scored. The cutoff value was considered to be the mean number of fluorescent GHOST cells used for background control plus 2 standard deviations. A primary clinical isolate, HIV-1MNp, was kindly provided by J. Sullivan of the University of Massachusetts Medical School, Worcester, MA. The virus was confirmed as one of the neutralization-resistant isolates by Susan Zolla-Pazner (7). Laboratory-adapted HIV-189.6and HIV-1MN were obtained from the AIDS Research and Reference Reagent Program. Simian/human immunodeficiency virus (SHIV) strain C2/1 was provided by K. Shinohara of the National Institute of Infectious Diseases (NIID), Tokyo, Japan (52, 56). SHIV strain 86.9 PD was kindly provided by Y. Lu of the Virus Research Institute, Cambridge, MA.
PBMC-based virus neutralization assay.Primary isolates were cultured with PHA-activated PBMCs in the presence of 40 units/ml of human interleukin-2 (Shionogi Pharmaceutical Co., Osaka, Japan) for 7 days, virus stocks were ti-trated on PHA-activated normal PBMCs, and the TCID50of each virus was determined (28). The primary isolates of HIV-1JR-CSFand the CS series were provided by Y. Koyanagi, Kyoto University Virus Research Institute, Kyoto, Japan, and JCI series isolates were also used. In vitro virus neutralization assays using diluted serum antibodies were performed as previously described (9, 28). In brief, 50l of various concentrations of serum IgG was preincubated with 50
l of 100 TCID50s of each virus strain in triplicate for 1 h at 37°C in a round-bottomed 96-well plate (Costar, New York, NY).One hundred microliters of 105 PHA-activated PBMCs originating from a single batch was added to that mixture for 1 h. After being washed, the cells were cultured in the presence of human interleukin-2 every day for approximately 7 to 21 days, with the period of culture depending on the property of each virus, and the amount of HIV was then measured by a p24 antigen ELISA (Dainabot, Tokyo, Japan). The neutralization titer is given as the concentration of serum IgG antibody or the reciprocal of serum dilution that reduced p24 antigen production by 90% (IC90) or 50% (IC50) compared to control wells with purified serum IgG from healthy individuals and preimmune mouse sera.
RESULTS
Sequential immunization with synthetic V3 peptides from HIV-1 clade B field isolates generates a cross-reactive anti-body response.Our previous study, which described anti-V3 antibody production, used single HIV-1 Env V3 peptides and produced homologous neutralizing antibodies. To find a more appropriate procedure to elicit a cross-reactive anti-body, we opted for sequential immunization using six syn-thetic peptides (SP1, SP17, SP11, SP14, SP12, and SP13) overlapping the tip of the V3 region of gp120 from both field and laboratory HIV-1 strains (Fig. 1A). Antibodies present in sera from the immunized mice exhibited high levels of binding to all the six homologous peptides and were highly cross-reactive with peptides (SP6, SP9, and SP10) derived
on November 8, 2019 by guest
http://jvi.asm.org/
from heterologous HIV-1 clade B field isolates containing the V3 core GPGR sequence (Fig. 1B). In contrast,
re-peated immunization with HIV-1MN Env V3 SP1 peptide
alone generated antibodies with specificity only for SP1 (Fig. 1C). Thus, sequential immunization using synthetic peptides derived from the V3 tip of HIV-1 gp120 generated site-specific and cross-reactive binding antibodies.
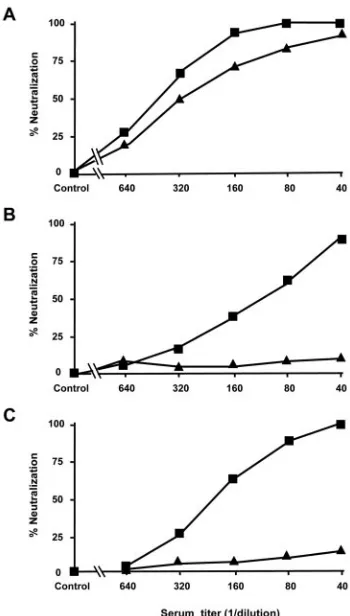
In order to analyze the neutralizing ability of the mouse anti-sera, a PBMC-based virus neutralization assay was performed
using T-cell-line-adapted (TCLA) HIV-1MN, CXCR4-tropic
pri-mary HIV-1MNp, and CCR5-tropic primary HIV-1JR-CSF. Both
types of immunizations successfully induced high
neutraliza-tion titers with IC50values of 450 in immune sera from
se-quentially immunized animals and with IC50values of 300 in
immune sera from mice receiving repeated immunizations of
single SP1 peptides against HIV-1MN(Fig. 2A). Furthermore,
the immune sera from the mice sequentially immunized with
the peptides efficiently neutralized HIV-1MNp with an IC50
titer of 110 and an IC90titer of 40 and neutralized HIV-1JR-CSF
with an IC50of 200 and IC90of 60 (closed squares in Fig. 2B
and C, respectively). In contrast, the immune sera from mice
immunized with the single SP1 peptide had no ability to
neu-tralize the HIV-1MNpand HIV-1JR-CSFisolates (closed
trian-gles in Fig. 2B and C, respectively). Preimmune and healthy control sera had no ability to neutralize the above-described three viruses. In summary, sequential immunization with rep-resentatives of Env V3 peptides produced high levels of cross-reactive neutralization responses in the immunized animals.
Production and neutralizing capacity of mouse MAb C25 and its humanized MAb, KD-247.It is important to determine whether the cross-reactive neutralization responses could be due to the effect of cross-reactive neutralization antibodies and to compare the data with those of established reference neu-tralization antibodies (7, 21). We first used the immunized mice with highly cross-reactive antibody responses and se-lected a hybridoma, named C25, which was reactive with het-erologous V3 peptide (Fig. 1D). MAb C25 reacted efficiently with the synthetic V3 peptides from multiple heterologous
strains of HIV-1 at concentrations of less than 0.01 g/ml;
however, SP17 (Fig. 1D, open circles) was less reactive, and SP14 (Fig. 1D, open triangles) showed no response at all. Furthermore, MAb C25 was highly reactive with all three clin-ically isolated heterologous V3 peptides (SP6, SP9, and SP10) (Fig. 1D). In short, sequential immunization proved to be effective at eliciting cross-reactive binding antibodies against the V3 tip epitopes of HIV-1. Furthermore, since C25 pos-sessed cross-reactive neutralization abilities as described Table 1, we produced a humanized antibody using the gene of the C25 mouse hybridoma. The CDRs and the partial FRs in the C25 gene were transferred into the human V region gene (data not shown) to produce a stable reshaped antibody clone known as KD-247. To reshape KD247, we aligned the amino acid se-quences deduced from the genes for C25 and KD-247 with the
FR of the REI VLand the FR of the subgroup I VHregions
(Fig. 3).
To assess the neutralizing abilities of C25 and KD-247, we evaluated their neutralization abilities against reference labo-ratory and clinical isolates by using MT-4 cell-based and PBMC-based neutralization assays. As shown in Table 1, we selected those target viruses that possessed a GPGR sequence in the Env V3 tip region. C25 and KD-247 similarly neutralized laboratory and primary isolates of CXCR4 and CCR5, with the
exception of HIV-1IIIB, which has an insertion of QR in front
of the glycine residue of the V3 tip sequence. As expected based on previous research, MT-4 cell- and PBMC-based neu-tralization assays showed that various concentrations (0.1 to
34.0g/ml of antibodies for IC90) of both the C25 and KD-247
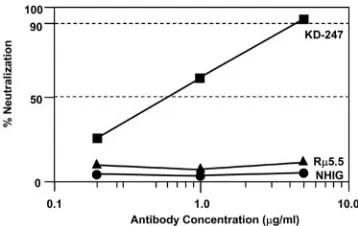
antibodies were capable of neutralizing target viruses that pos-sessed a GPGR sequence in the V3 tip sequence. To evaluate the neutralization abilities of KD-247, we compared its neu-tralization activity with those of established reference antibod-ies by GHOST cell neutralization assays using GHOST-X4 and GHOST-R5 cells (7) expressing the chemokine receptors CXCR4 and CCR5, respectively. As shown in Fig. 4, KD-247
efficiently neutralized HIV-1MNpon GHOST-X4 cells with an
IC90of approximately 5g/ml, while neither normal human
IgG nor a similar reshaping anti-V3 antibody, R5.5, which
recognizes the V3 tip sequence IHIGPGRAFYT, neutralized
HIV-1MNp. Thus, although the V3 tip sequence of HIV-1MNp
(RIHIGPGRAFYTTKN) (Table 2) was identical to the V3
[image:4.585.75.250.71.379.2]sequence of R5.5 (underlined), R5.5 did not neutralize
FIG. 2. Neutralization of HIV-1MNand clinical isolates HIV-1MNp
and HIV-1JR-CSFby sera from sequentially immunized mice with
di-verse V3 tip peptides but not by sera from mice repeatedly immunized with a single SP1 peptide. Shown are data from neutralization of HIV-1 strains MN (A), MNp (B), and JR-CSF (C) by immune sera from sequentially immunized mice with diverse V3 tip peptides (■) or by immune sera from mice with repeated immunization with a single SP1 peptide (Œ) determined by a PBMC-based virus neutralization assay.
on November 8, 2019 by guest
http://jvi.asm.org/
HIV-1MNp, but KD-247 did. These results suggest that the
neutralization epitope of HIV-1MNpmay be narrower than that
of the V3 sequence of R5.5 and that KD-247 may recognize
the narrow-neutralization epitope of the V3 tip sequence.
Testing of the laboratory isolate HIV-1MNby the GHOST cell
assay confirmed that KD-247 neutralized HIV-1MNefficiently,
with an IC90and IC50of 1.0 and 0.1g/ml, respectively.
HIV-1AD8, SHIV 89.6, and SHIV C2/1, whose V3 tip sequences
were confirmed to be KSIHIGPGRAFYT, RLSIGPGRA FYA, and RLSIGPGRAFYA, respectively, were less sensitive
to neutralization by KD-247, with IC90values of 10, 5, and 5
[image:5.585.42.545.82.262.2]g/ml, respectively. In contrast, those three viruses were not
TABLE 1. Neutralizing activities of C25 and KD-247 against HIV-1 clade Ba
Target HIV-1 isolate Env V3 sequenceb IC90(g/ml)
C25 KD-247c
MT-4 cells HIV-1MN CTRPNYNKRKRIHI GPGRAFYTTKNIIGTIRQAHC 2.0 2.0
HIV-1IIIB –––––N–T–––KS–QR––––––V–IGK– –NM––––– ⬎500 ⬎500
HIV-1N154 –––––N–IK––––– –––––––––GE–––N–––––– 1.0 1.0
HIV-1N161 –––––N–T––G–R– –––––V–A–GK–––D–––––– 3.9 2.0
HIV-1DBA3 –––––N–T––G–R– –––––V–A–EK–––D–––––– 3.9 2.0
HIV-1N163 –––––N–T–RG–R– –––––––A–GK–––D–––––– 0.5 1.0
HIV-1N153 –––––N–T––A–RV ––––TL–A–RR–––D–––––– 31.2 31.2
HIV-1TM2 –––––NKA–G–LSV ––––S––––RQ–T–D–––––– 3.9 3.9
PBMCs HIV-1MN CTRPNYNKRKRIHI GPGRAFYTTKNIIGTIRQAHC 0.1 0.1
HIV-1JR-CSF ––––SN–K––S––– –––––––––GE–––D–––––– 5.0 5.0
HIV-1CS3-5 –––I–N–T––S––– –––––––A–GE–––N–K–––– 10.0 10.0
HIV-1CS6-8 –I–––N–T––G––– –––––––A–D––––N–––––– 8.0 5.0
HIV-1JCI-3 –––––N–I––H––– –––––––––RG –RD––K––– 10.0 4.0
HIV-1JCI-9 –––––––T––G––– ––––––V–––G –RD––K––– 21.0 ND
HIV-1JCI-11 –––––––TS–G–R– –––––––ASER –RD––K––– 34.0 ND
HIV-1JCI-22 –––––N–I––H––– –––––––––RG –RD––K––– 12.0 ND
a
The HIV-1 sequences were confirmed by proviral DNA sequencing of virus-infected cells.
b
Dashes indicate sequence homology to HIV-1MN, and spaces represent the presence of a deletion.
c
ND, not done.
FIG. 3. Design to synthesize an amino acid sequence that encoded the reshaping antibody KD-247. Amino acid sequences of VHand VLregions
of human IgG, KD-247, and C25 are shown. Boxes indicate amino acids in the CDRs and a portion of the FR segments in the reshaped human V regions, which were transferred from the mouse MAb. Dashes indicate a deletion. The sequence of a control reshaping MAb, R5.5, derived from a murine MAb obtained by immunization with SP1 peptide alone is also shown.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.585.57.533.417.685.2]neutralized by the other previously reported anti-V3
neutral-izing MAbs, R5.5 and C1.
To further substantiate that KD-247 neutralizes primary iso-lates, we also tested seven primary isolates by comparing them to the following international reference antibodies: anti-V3 MAb 447-52D, anti-V3 MAb 1006-15D, anti-V3 MAb 694/ 98D, anti-C5 MAb 1331-160, and anti-gp41 MAb 1367-D (Table 3). KD-247 most efficiently neutralized HIV-1 clade B primary isolates from the AIDS Research and Reference Re-agent Program, NIAID, and from the HIV Isolation Project,
NIID, with IC50s at various concentrations, from 0.78 to 34.26
g/ml, although it did not prove effective against the N-NIID
isolate, which possessed a GPGR sequence in the Env V3 tip region. These results demonstrate that KD-247 more effec-tively neutralizes primary isolates with both CXCR4 and CCR5 tropism than do any of the reference antibodies (Tables 2 and 3).
Properties of the anti-HIV envelope V3 MAb KD-247 that neutralize viral infection.To more precisely address the neu-tralization mechanisms of KD-247, we investigated its bio-chemical properties.
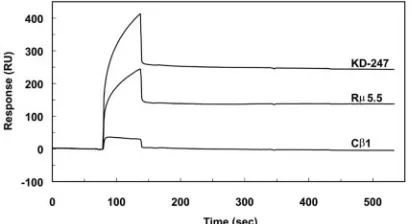
(i) Peptide affinity.The affinity of KD-247 for the SP1 pep-tide was evaluated by surface plasmon resonance-based mea-surements using a BIAcore instrument and then compared to
the affinity of R5.5 and C1 for HIV-1MN and HIV-1IIIB,
respectively (41). Sensorgrams showed that C1 bound slightly
to SP1, while KD-247 bound more than did R5.5 (Fig. 5). The
equilibrium dissociation constants (KDs) of KD-247 and R5.5
were 1.3⫻10⫺9M (ka, 1.3⫻105M⫺1s⫺1;kd, 1.7⫻10⫺4s⫺1)
and 2.9⫻10⫺10M (ka, 1.0⫻105M⫺1s⫺1;kd, 2.9⫻10⫺5s⫺1),
respectively.
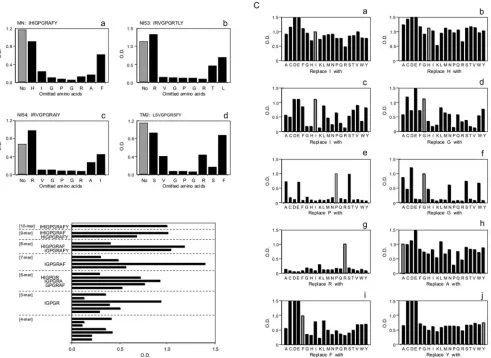
(ii) Epitope mapping.We tested KD-247 for epitope map-ping by reacting the antibody with decamer peptides derived from the V3 tip sequences of three different clinical HIV-1
isolates (NI53, NI54, and TM2) and HIV-1MN. Deletion of
amino acids lying in the central region of each peptide resulted in a reduction in KD-247 binding, while deletion of amino acids near either terminal had little effect (Fig. 6A). To deter-mine the shortest reactive peptide, KD-247 was tested for binding activity against a series of overlapping peptides that varied in length from 4 to 10 amino acids. For optimal binding, KD-247 required peptides of five or more amino acids con-taining an IGPGR sequence (Fig. 6B). In addition, sets of peptide analogues that differed from the original decamer pep-tide by only one amino acid were synthesized. Each amino acid within a given peptide was replaced in turn by one of 19 naturally occurring amino acids, and the resulting peptides were tested for reactivity against KD-247 (Fig. 6C). The amino acids I, H, A, F, and Y in the flanking region of the core sequence could easily be replaced by many other residues without the loss of KD-247 binding (Fig. 6C, panels a, b, h, i, and j). Although a few amino acids in the core PG sequence could be replaced (Fig. 6C, panels e and f), the arginine resi-due was found to be critical for maintaining KD-247 binding at levels equivalent to those of the original peptide (Fig. 6C, panel g). In summary, KD-247, a humanized antibody with
high affinity (KDof 1.3⫻10⫺
9M), recognized the PGR of the
narrow V3 tip sequence and should therefore act as an effec-tive neutralizing antibody.
DISCUSSION
Sequential immunization with multiple V3 peptides of HIV-1 Env is desirable in that it can evoke cross-reactive HIV-1-neutralizing antibodies, while repeated immunizations with a single peptide elicit only a non-cross-reactive and virus-specific neutralizing antibody response. Of these two types of antibody responses, only the cross-reactive one produced by sequential immunization neutralized CCR5-tropic primary and other heterologous isolates. Because it appears to raise an effective antibody specific for the HIV-1 Env neutralization epitope, this sequential immunization regimen may show con-siderable promise for use in vaccine development by an active immunization procedure.
A single C4-V3 peptide immunogen from HIV-189.6 was
shown to partially protect immunized animals from
homolo-FIG. 4. Neutralization of HIV-1MNp by KD-247 in GHOST-X4
[image:6.585.73.252.69.182.2]cells. Each dilution of MAbs KD-247 and R5.5 was tested in tripli-cate, and results are expressed as the means of three different exper-iments. Normal human IgG (NHIG) was used as the control IgG for the neutralization assay; results are expressed as the means of data from four different assays of triplicate samples.
TABLE 2. Neutralizing activity of KD-247 against R5 and R5/X4 dual-tropic viruses by GHOST cell assaya
Isolate Env V3 sequence
GHOST cell assay
KD-247 R5.5 (V3) C1 (V3)
IC90 IC50 IC90 IC50 IC90 IC50
HIV-1MN CTRPNYNKRKRIHI GPGRAFYTTKNIIGTIRQAHC X4 1 0.1 1 0.2 ⬎50 ⬎50
HIV-1MNp –––––N–R–T–––– ––––––––––––––––––––– X4 5 0.6 ⬎50 ⬎50 ⬎50 ⬎50
HIV-1IIIB –––––N–T–––KS–QR––––––V–IGK– –NM––––– X4 ⬎50 ⬎50 ⬎50 ⬎50 5 0.5
HIV-1AD8 –––––N–T––S––– –––––––––GD–––D–––––– R5 10 5 ⬎50 ⬎50 ⬎50 ⬎50
SHIV 89.6PD –––––N–T–R–LS– –––––––ARR––––D–––––– R5/X4 5 0.5 ⬎50 ⬎50 ND ND
SHIV C2/1 –––––N–T–E–LS– –––––––ARR––––D–––––– R5/X4 5 0.5 ⬎50 ⬎50 ND ND
aThe amino acid sequences were confirmed by proviral DNA sequencing of virus-infected cells. Dashes indicate sequence homology to HIV-1
MN; spaces represent the presence of a deletion. ND, not done.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.44.541.616.708.2]gous SHIV 89.6P challenge (35). Furthermore, recent progress in the development of an HIV-1 vaccine expressing multiple Env genes has been made, suggesting the potential production of cross-clade neutralizing antibodies (1, 8, 37, 55). These findings also suggest that the use of sequential immunization with biologically relevant peptides or proteins for the develop-ment of HIV-1 vaccines could overcome difficulties associated with otherwise poorly immunogenic epitopes of virus neutral-ization.
The ability of KD-247 to neutralize HIV-1 may be depen-dent on site-specific binding to epitopes on the viral envelope glycoprotein. The CDRs of KD-247 were transferred from mouse MAb C25, which was designed to have cross-reactive neutralization activity against HIV-1 clade B isolates. MAb C25 was elicited by sequential immunization using six different peptides containing the V3 GPGR sequence. Antigen recog-nition by KD-247 was little affected by the genetic reshaping of the C25 gene. Indeed, the results of the Pepscan analysis sug-gest that KD-247 can react with core V3 sequences from many HIV-1 clade B isolates. The recognition site of KD-247 was mapped to five or six amino acids around the PGR core se-quence at the tip of the V3 region of gp120. The shortest peptide that was reactive with KD-247 was regarded as IGPGR, but the epitope is stabilized by the addition of one or more supplemental amino acids. The GPGR sequence of the V3 tip is highly con-served among HIV-1 strains (27, 66). Furthermore, IGPGRA and GPGRAF sequences were detected in the majority of HIV-1 isolates from donors in the United States (34). Using the previ-ously published sequences found in the Los Alamos HIV-1
se-quence database, we confirmed that these sese-quences are present in the majority of HIV-1 clade B isolates (36, 66).
[image:7.585.45.542.81.196.2]With regard to the properties of the anti-V3 antibody in-duced by active immunization protocols in animals for HIV vaccine development, the difficulty with exploiting the effica-cious antiviral neutralizing antibody responses of anti-V3 an-tibodies lies in the extraordinary diversity of the V3 sequence and in the resulting strict type specificity of anti-V3 neutraliz-ing antibodies. That strict type specificity is thought to be a function of the inaccessibility of the neutralization epitope on the envelope of the virus (3, 51, 64). This is certainly the case with the polyclonal anti-V3 antibody, which was produced by repeated immunizations with a single HIV Env V3 peptide and which neutralized the only homologous virus in this study. However, sequential immunization of mice with a selection of V3 peptides elicited cross-reactive neutralization antibody re-sponses, and we were eventually able to construct a humanized monoclonal antibody, KD-247, which showed a relatively high level of affinity to the narrow “PGR” motif within the V3 determinant. The humanized antibody was capable of neutral-izing a broader range of clade B primary isolates than did the previously reported authentic anti-V3 neutralizing antibody. Thus, the strict type specificity of the anti-V3 antibody elicited by active immunization can be overcome by driving the anti-body responses to a more conserved motif within V3. This can be done through sequential immunization with peptides rep-resentative of V3 sequences. The KD-247 antibody would be expected to have binding activity against a wide range of HIV-1 field isolates, because the IGPGRA and GPGRAF sequences predominate in the majority of the clade B isolates recognized by the MAb. The results of Pepscan analysis with replacement peptides also suggest that KD-247 has broad binding activity for HIV-1. Although few amino acid substitutions were toler-ated in the central PGR sequence of the V3 tip peptide, a number of amino acids substitutions were permitted in the flanking region. It would therefore be expected that KD-247 would bind to HIV-1 quasispecies that have a similar recogni-tion sequence. Interestingly, although the N-NIID isolate showed the same IGPGR V3 tip sequence, it was not neutral-ized by KD-247 or by any of the other antibodies tested (Table 3). In contrast, the Pepscan analysis showed that the INIG PGRA V3 tip peptide of the Env region in the N-NIID isolate bound KD-247 using short synthetic peptides (Fig. 6C, graph b). These results raise the possibility that some amino acids
[image:7.585.61.267.590.702.2]FIG. 5. Sensorgram overlays for the binding of KD-247, R5.5, and C1 to the SP1 peptide.
TABLE 3. Fifty-percent neutralization activity on GHOST/R5 or GHOST/X4a
Isolate Env V3 sequence GHOST
cell
IC50(g/ml)
KD-247 (V3)
447-52D (V3)
1006-15D (V3)
694/98D (V3)
1331-D (C5)
1367-D (gp41)
HIV-1MN CTRPNYNKRKRIHI GPGRAFYTTKNIIGTIRQAHC X4 0.01 0.01 0.02 8.64 ⬎50 ⬎50
HIV-1JR-CSF ––––SN–T––S––– –––––––––GE–––D–––––– R5 8.34 ⬎50 ⬎50 ⬎50 ⬎50 ⬎50
HIV-192US712 –T–––N–T––S––– –––––––––GD–––D––K––– R5 9.03 ⬎50 ⬎50 ⬎50 ⬎50 ⬎50
HIV-1N-NIID –––––N–T––S–N– –––––––A–GD–––D–––––– R5 ⬎50 ⬎50 ⬎50 ⬎50 ⬎50 ⬎50
HIV-192TH022 ––––SN–T–TS–T– –––QV––R–GD–––D––K–Y– R5 ⬎50 ⬎50 ⬎50 ⬎50 ⬎50 ⬎50
HIV-192TH014 –––––N–T––S–PL –––––W–––GQ–––D–––––– R5 0.78 1.50 1.31 1.9 ⬎50 ⬎50
HIV-189.6 –––––N–T–R–LS– –––––––ARR––––D–––––– R5/X4 13.46/11.6 ⬎50 ⬎50 ⬎50 ⬎50 ⬎50
HIV-1NIH1054 ––––SN–T–R–––– –––––––––RQ–R–D–––––– R5/X4 34.26/33.31 ⬎50 ⬎50 ⬎50 ⬎50 ⬎50
a
The amino acid sequences were confirmed by proviral DNA sequencing of virus-infected cells. Results show antibody concentration of IC50(micrograms/milliliter). Dashes indicate sequence homology to HIV-1MN, and spaces represent the presence of a deletion.
on November 8, 2019 by guest
http://jvi.asm.org/
neighboring the neutralization epitope in the viral particle can influence steric hindrance of the binding site, thereby protect-ing the virus from antibody neutralization.
In studies centered on anti-V3 MAb produced by heterohy-bridomas using PBMCs from HIV-infected individuals, Gorny et al. recently showed that the V3 loop is accessible on the surface of most primary HIV-1 isolates and serves as a neu-tralization epitope (23). They also recently identified a quater-nary neutralizing epitope on HIV particles (24). Since the fusion of the heteromyeloma and the PBMCs from HIV-in-fected individuals results in the production of these unique cross-reactive MAbs, a similar sequential immunization with diverse V3 antigens of HIV quasispecies in HIV-infected in-dividuals could lead to the generation of responsible PBMCs, which in turn could produce a cross-neutralizing V3 anti-body (21, 23). Actually, human MAb 447-52D shows functional and epitope-mapping characteristics (4, 21) similar to those of
the 247 humanized antibody described in this study; KD-247 and 447-52D also both possess a similarly narrow binding epitope on the V3 region with high affinity to the “PGR” motif and “GPxR” motif at the center of the V3 region, respectively, and they show cross-reactive neutralization against primary isolates and TCLA isolates. Although KD-247 does not
neu-tralize HIV-1IIIB, which has an insertion of the QR sequence
just before the “GPGR” motif in the center of the V3 region, we cannot as of yet compare the breadth of the neutralization activity of the two antibodies, since we used only six primary isolates for this comparison.
[image:8.585.52.543.70.428.2]The gp120 V1/V2-domain structure and not the sequence variations at the target sites is thought to mediate the neutral-ization sensitivity of HIV-1 pseudotyped with Env proteins derived from HIV-1 strains SF162 and JR-FL as well as the inherent neutralization resistance of JR-FL and presumably of related primary isolates (48). Despite similar binding affinities
FIG. 6. Pepscan with deletion (A), overlapping (B), and replacement (C) peptides. KD-247 binding activity for the peptides is proportional to the value of the optical density (O.D.) at 450 nm. Hatched bars indicate the reactivity of the original sequence without any deletion (A) or replacement (C). The amino acid deleted from the original sequence is indicated on the horizontal axis (A). The original sequences for the set of deletion analogues were IHIGPGRAFY (A, panel a), RVGPGRTL (A, panel b), RVGPGRAI (A, panel c), and SVGPGRSF (A, panel d), derived from HIV-1 strains MN, NI53, NI54, and TM2, respectively. The decamer peptide derived from the V3 region of the MN strain was also synthesized; overlapping peptides of multiple lengths (B) and replacement peptides (C) are shown. The length of each set of peptides and the sequences, which exhibited relatively strong binding activity, are indicated on the vertical axis (B). Every group of 20 lines corresponds to the complete replacement set for 1 of the 10 amino acid positions in the decamer peptide (C, panels a to j). The amino acid replaced with the original sequence is indicated on the horizontal axis (c).
on November 8, 2019 by guest
http://jvi.asm.org/
for MAbs against V3-, V2- and CD4-binding domains, three MAbs of immunoglobulin G, b12, 2G12, and 2F5, neutralized the JR-FL virus. Using single-cell viral transduction assays mediated by both JR-FL Env pseudotypes and SF162 Env pseudotypes, we plan to compare the neutralization sensitivity of the humanized anti-V3 MAb KD-247 and of immune sera from animals sequentially immunized with a selection of V3 peptides.
The current findings suggest that high affinity for antibody binding is required for neutralization as well as site-specific localization of epitopes to the V3 tip. The kinetic parameters of KD-247 were identified to be fast on rates and slow off rates,
similar to those of R5.5, although theKDvalue of KD-247 for
binding to the SP1 peptide was higher than that of R5.5. This
higher value results from the faster rate of association for
KD-247 (1.3⫻105M⫺1s⫺1) than that for R5.5 (1.0⫻105
M⫺1 s⫺1). This finding is reasonable, since the epitope of
KD-247 (IGPGR) is shorter than that of R5.5 (IHIGPGRA
FYT). The higher rate of association of KD-247 might be responsible for the virus neutralization activity of the antibody. These results are consistent with hypotheses that propose to use kinetic parameters of antibody binding to explain virus neutralization.
The V3 region of HIV-1 gp120 has been identified as the major determinant of cellular tropism (57) based on corecep-tor specificity (63, 65). It is assumed that anti-V3 region neu-tralizing antibodies inhibit the interaction between the V3 re-gion and its coreceptors. The critical domains, which are concerned with the utilization of the chemokine receptors CCR5 and CXCR4, are located outside of the central IGP GRAF sequence in the V3 region (61). The replacement of amino acids flanking the IGPGRAF sequence had little effect on the binding of KD-247 to peptides, showing that KD-247 neutralized CCR5 and CXCR4 as well as dual-tropic HIV-1 isolates (Fig. 6A and C). In this study, we have shown how neutralizing and nonneutralizing antibodies against primary viruses differ qualitatively. The type-specific anti-V3 MAb
R5.5, which binds a linear epitope at the tip of V3 (IHIGP
GRAFYT), has the ability to neutralize TCLA CXCR4-tropic
virus HIV-1MNbut neutralizes neither the CCR5-tropic strain
HIV-1AD8nor the primary virus HIV-1MNp(Fig. 4). In contrast,
the humanized anti-V3 antibody KD-247 efficiently neutralized these viruses in neutralization assays using both PBMCs and GHOST cells (Tables 1 and 2).
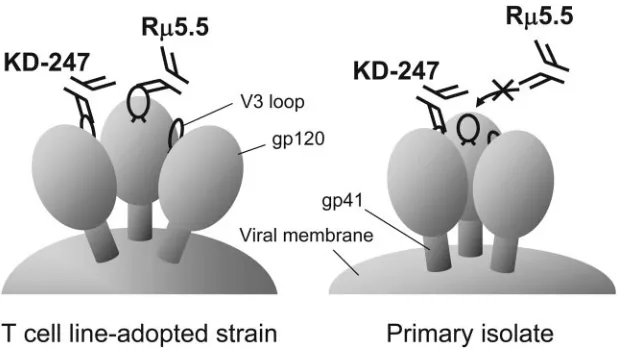
As previously reported, the envelope spikes of HIV-1 consist of a transmembrane gp41 molecule interacting noncovalently with a gp120 molecule to form an oligomeric structure, most likely a trimer. It has been proposed that the gp120 trimer of TCLA strains forms a relatively open conformation and that the primary isolate trimeric complex has a more closed con-formation (4, 49). For these reasons, we propose that a part of
the epitope recognized by R5.5 is hidden by the
oligomeriza-tion of gp120 on primary HIV-1 isolates, reducing its accessi-bility to the antibody. The aaccessi-bility of KD-247 to neutralize both primary and CCR5-tropic isolates suggests that the V3 tip site protrudes from the gp120 trimer or is located within an area accessible to KD-247 on primary CCR5-tropic isolates but not
to R5.5 (Fig. 7). However, R5.5, produced by repeated
immunization with SP1, did effectively neutralize homologous TCLA strains of HIV-1 (Fig. 7).
Our speculation that the V3 tip epitope on the virion is one of the neutralization targets of primary isolates accords well with recent reports suggesting that the V3 epitope is more exposed on intact virions than are the CD4-binding domains and the V2 and gp41 regions (67). Conceivably, nonconforma-tional epitopes may also be responsible for neutralization, since the neutralizing antibody 2F5 recognizes a linear epitope, ELDKWA, in the membrane-proximal region of HIV-1 gp41 (43). Furthermore, HIV-1 envelope proteins have at least three conformational states: an unbound state on the surface of virions, a CD4-bound state, and an end-product state in which the gp120 proteins have dissociated from gp41 trimers (16, 53). Since KD-247 suppresses the ex vivo generation of primary HIV-1 quasispecies in PBMC cultures from
HIV-in-FIG. 7. Schematic neutralization model of the cross-reactive anti-V3 MAb KD-247 compared with the type-specific anti-V3 MAb R5.5. Neutralization of primary CCR5-tropic isolates by KD-247 suggests that the narrow V3 tip epitope for KD-247 binding and neutralization sticks out from the gp120 trimer of primary CCR5-tropic isolates, or it is located in an accessible site of the gp120 trimer during neutralization by KD-247. However, R5.5 may not be able to bind to the V3 tip epitope, since the neutralization epitope for R5.5 is broader than KD-247’s epitope (Fig. 6), which may not be fully expressed and are more efficiently masked in the primary R5-tropic viruses compared to TCLA strains.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:9.585.138.450.68.243.2]fected individuals (13a), the binding of KD-247 to an appro-priate binding epitope on the V3 region of primary HIV-1 can be presumed to occur and to express itself at some point during the three conformational states. KD-247 could then neutralize primary HIV-1.
ACKNOWLEDGMENTS
We thank Richard M. Krause and Malcolm Martin, National Insti-tute of Allergy and Infectious Diseases, National InstiInsti-tutes of Health, Bethesda, Md., for their helpful comments.
Valuable reagents and assistance were contributed by Susan Zolla-Pazner with the support of an NIH grant funding the Viral Immunol-ogy Core of the NYU Center for AIDS Research (AI 27742). The Panel on AIDS of the US-Japan Cooperative Medical Science Pro-gram, the Human Science Foundation, Japan, and the Japanese Min-istry of Health, Labor, and Welfare also supported this study.
REFERENCES
1.Akahata, W., Z. Y. Yang, and G. J. Nabel.2005. Comparative immunoge-nicity of human immunodeficiency virus particles and corresponding poly-peptides in a DNA vaccine. J. Virol.79:626–631.
2.Boudet, F., M. Girard, J. Theze, and M. Zouali.1992. Antibodies of HIV-1 positive subjects and experimentally immunized primates and rodents bind to sequence divergent regions of the third variable domain (V3) of gp120. Int. Immunol.4:283–294.
3.Bou-Habib, D. C., G. Roderiquez, T. Oravecz, P. W. Berman, P. Lusso, and M. A. Norcross.1994. Cryptic nature of envelope V3 region epitopes pro-tects primary monocytotropic human immunodeficiency virus type 1 from antibody neutralization. J. Virol.68:6006–6013.
4.Burton, D. R.1997. A vaccine for HIV type 1: the antibody perspective. Proc. Natl. Acad. Sci. USA94:10018–10023.
5.Cardoso, R. M., M. B. Zwick, R. L. Stanfield, R. Kunert, J. M. Binley, H. Katinger, D. R. Burton, and I. A. Wilson.2005. Broadly neutralizing anti-HIV antibody 4E10 recognizes a helical conformation of a highly conserved fusion-associated motif in gp41. Immunity22:163–173.
6.Casadevall, A., and M. D. Scharff.1995. Return to the past: the case for antibody-based therapies in infectious diseases. J. Infect. Dis.21:150–161. 7.Cecilia, D., V. N. Kewalramani, J. O’Leary, B. Volsky, P. Nyambi, S. Burda,
S. Xu, D. R. Littman, and S. Zolla-Pazner.1998. Neutralization profiles of primary human immunodeficiency virus type 1 isolates in the context of coreceptor usage. J. Virol.72:6988–6996.
8.Chakrabarti, B. K., W. P. Kong, B. Y. Wu, Z. Y. Yang, J. Friborg, X. Ling, S. R. King, D. C. Montefiori, and G. J. Nabel.2002. Modifications of the human immunodeficiency virus envelope glycoprotein enhance immunoge-nicity for genetic immunization. J. Virol.76:5357–5368.
9.Chujoh, Y., K. Matsuo, H. Yoshizaki, T. Nakasatomi, K. Someya, Y. Oka-moto, S. Naganawa, S. Haga, H. Yoshikura, S. Yamazaki, and M. Honda.
2001. Cross-clade neutralizing antibody production against human immuno-deficiency virus type 1 clade E and B⬘strains by recombinant mycobacterium bovis BCG-based candidate vaccine. Vaccine20:797–804.
10.Cohen, O. J., A. Kinter, and A. S. Fauci.1997. Host factors in the patho-genesis of HIV disease. Immunol. Rev.159:31–48.
11.Conley, A. J., M. K. Gorny, J. A. Kessler II, L. J. Boots, M. Ossorio-Castro, S. Koenig, D. W. Lineberger, E. A. Emini, C. Williams, and S. Zolla-Pazner.
1994. Neutralization of primary human immunodeficiency virus type 1 iso-lates by the broadly reactive anti-V3 monoclonal antibody, 447-52D. J. Virol.
68:6994–7000.
12.Dolin, R.1995. Human studies in the development of human immunodefi-ciency virus vaccines. J. Infect. Dis.172:1175–1183.
13.Durda, P. J., L. Bacheler, P. Clapham, A. M. Jenoski, B. Leece, T. J. Matthews, A. McKnight, R. Pomerantz, M. Rayner, and K. J. Weinhold.
1990. HIV-1 neutralizing monoclonal antibodies induced by a synthetic pep-tide. AIDS Res. Hum. Retrovir.6:1115–1123.
13a.Eda, Y., T. Murakami, Y. Ami, T. Nakasone, M. Takizawa, K. Someya, M. Kaizu, Y. Izumi, N. Yoshino, S. Matsushita, H. Higuchi, H. Matsui, K. Shinohara, H. Takeuchi, Y. Koyanagi, N. Yamamoto, and M. Honda.2006. Anti-V3 humanized antibody KD-247 effectively suppresses ex vivo genera-tion of human immunodeficiency virus type 1 and affords sterile protecgenera-tion of monkeys against a heterologous simian/human immunodeficiency virus infection. J. Virol.80:5563–5570.
14.Emini, E. A., W. A. Schleif, J. H. Nunberg, A. J. Conley, Y. Eda, S. Tokiyoshi, S. D. Putney, S. Matsushita, K. E. Cobb, C. M. Jett, J. W. Eichberg, and K. K. Murthy.1992. Prevention of HIV-1 infection in chimpanzees by gp120 V3 domain-specific monoclonal antibody. Nature355:728–730.
15.Fearon, D. T., and R. M. Locksley. 1996. The instructive role of innate immunity in the acquired immune response. Science272:50–53.
16.Furuta, R. A., C. T. Wild, Y. Weng, and C. D. Weiss.1998. Capture of an early fusion-active conformation of HIV-1 gp41. Nat. Struct. Biol.5:276–279.
17.Geysen, H. M., R. H. Meloen, and S. J. Barteling.1984. Use of peptide synthesis to probe viral antigens for epitopes to a resolution of a single amino acid. Proc. Natl. Acad. Sci. USA81:3998–4002.
18.Gorny, M. K., A. J. Conley, S. Karwowska, A. Buchbinder, J. Y. Xu, E. A. Emini, S. Koenig, and S. Zolla-Pazner.1992. Neutralization of diverse hu-man immunodeficiency virus type 1 variants by an anti-V3 huhu-man monoclo-nal antibody. J. Virol.66:7538–7542.
19.Gorny, M. K., C. Williams, B. Volsky, K. Revesz, S. Cohen, V. R. Polonis, W. J. Honnen, S. C. Kayman, C. Krachmarov, A. Pinter, and S. Zolla-Pazner.2002. Human monoclonal antibodies specific for conformation-sen-sitive epitopes of V3 neutralize human immunodeficiency virus type 1 pri-mary isolates from various clades. J. Virol.76:9035–9045.
20.Gorny, M. K., J. P. Moore, A. J. Conley, S. Karwowska, J. Sodroski, C. Williams, S. Burda, L. J. Boots, and S. Zolla-Pazner.1994. Human anti-V2 monoclonal antibody that neutralizes primary but not laboratory isolates of human immunodeficiency virus type 1. J. Virol.68:8312–8320.
21.Gorny, M. K., J. Y. Xu, S. Karwowska, A. Buchbinder, and S. Zolla-Pazner.
1993. Repertoire of neutralizing human monoclonal antibodies specific for the V3 domain of HIV-1 gp120. J. Immunol.150:635–643.
22.Gorny, M. K., J. Y. Xu, V. Gianakakos, S. Karwowska, C. Williams, H. W. Sheppard, C. V. Hanson, and S. Zolla-Pazner.1991. Production of site-selected neutralizing human monoclonal antibodies against the third variable domain of the human immunodeficiency virus type 1 envelope glycoprotein. Proc. Natl. Acad. Sci. USA88:3238–3242.
23.Gorny, M. K., K. Revesz, C. Williams, B. Volsky, M. K. Louder, C. A. Anyangwe, C. Krachmarov, S. C. Kayman, A. Pinter, A. Nadas, P. N. Nyambi, J. R. Mascola, and S. Zolla-Pazner.2004. The V3 loop is accessible on the surface of most human immunodeficiency virus type 1 primary isolates and serves as a neutralization epitope. J. Virol.78:2394–2404.
24.Gorny, M. K., L. Stamatatos, B. Volsky, K. Revesz, C. Williams, X. H. Wang, S. Cohen, R. Staudinger, and S. Zolla-Pazner.2005. Identification of a new quaternary neutralizing epitope on human immunodeficiency virus type 1 virus particles. J. Virol.79:5232–5237.
25.Gorny, M. K., T. C. VanCott, C. Hioe, Z. R. Israel, N. L. Michael, A. J. Conley, C. Williams, J. A. Kessler II, P. Chigurupati, S. Burda, and S. Zolla-Pazner.1997. Human monoclonal antibodies to the V3 loop of HIV-1 with intra- and interclade cross-reactivity. J. Immunol.159:5114–5122. 26.Gorny, M. K., V. Gianakakos, S. Sharpe, and S. Zolla-Pazner.1989.
Gen-eration of human monoclonal antibodies to human immunodeficiency virus. Proc. Natl. Acad. Sci. USA86:1624–1628.
27.Hattori, T., K. Shiozaki, Y. Eda, S. Tokiyoshi, S. Matsushita, H. Inaba, M. Fujimaki, T. Meguro, K. Yamada, M. Honda, K. Nishikawa, and K. Takatsuki.
1991. Characteristics of the principal neutralizing determinant of HIV-1 prev-alent in Japan. AIDS Res. Hum. Retrovir.7:825–830.
28.Honda, M., K. Matsuo, T. Nakasone, Y. Okamoto, H. Yoshizaki, K. Kita-mura, W. Sugiura, K. Watanabe, Y. Fukushima, S. Haga, Y. Katsura, H. Tasaka, K. Komuro, T. Yamada, T. Asano, A. Yamazaki, and S. Yamazaki.
1995. Protective immune responses induced by secretion of a chimeric sol-uble protein from a recombinantMycobacterium bovisbacillus Calmette-Gue´rin vector candidate vaccine for human immunodeficiency virus type 1 in small animals. Proc. Natl. Acad. Sci. USA92:10693–10697.
29.Javaherian, K., A. J. Langlois, C. McDanal, K. L. Ross, L. I. Eckler, C. L. Jellis, A. T. Profy, J. R. Rusche, D. P. Bolognesi, S. D. Putney, and T. J. Matthews.1989. Principal neutralizing domain of the human immunodefi-ciency virus type 1 envelope protein. Proc. Natl. Acad. Sci. USA86:6768– 6772.
30.Kabat, E. A., T. T. Wu, M. Reid-Miller, H. M. Perry, and K. S. Gottesman.1987. Sequence of proteins of immunological interest, 4th ed. U.S. Department of Health and Human Services, National Institutes of Health, Bethesda, Md. 31.Karlsson, R., A. Michaelsson, and L. Mattsson.1991. Kinetic analysis of
monoclonal anti-antigen interaction with a new biosensor based analytical system. J. Immunol. Methods145:229–240.
32.Kohler, G., and C. Milstein.1975. Continuous cultures of fused cells secret-ing antibody of predefined specificity. Nature256:495–497.
33.Laman, J. D., M. M. Schellekens, Y. H. Abacioglu, G. K. Lewis, M. Tersmette, R. A. Fouchier, J. P. Langedijk, E. Claasen, and W. J. Boersma.1992. Variant-specific monoclonal and group-specific polyclonal human immuno-deficiency virus type 1 neutralizing antibodies raised with synthetic peptides from the gp120 third variable domain. J. Virol.66:5175.
34.LaRosa, G. J., J. P. Davide, K. Weinhold, J. A. Waterbury, A. T. Profy, J. A. Lewis, A. J. Langlois, G. R. Dreesman, N. Boswell, P. Shadduck, L. H. Holley, M. Karplus, D. P. Bolognesi, T. J. Matthews, E. A. Emini, and S. D. Putney.1990. Conserved sequence and structural elements in the HIV-1 principal neutralizing determinant. Science249:932–935.
35.Letvin, N. L., S. Robinson, D. Rohne, M. K. Axthelm, J. W. Fanton, M. Bilska, T. J. Palker, H. X. Liao, B. F. Haynes, and D. C. Montefiori.2001. Vaccine-elicited V3 loop-specific antibodies in rhesus monkeys and control of a simian-human immunodeficiency virus expressing a primary patient human immunodeficiency virus type 1 isolate envelope. J. Virol.75:4165– 4175.
36.Los Alamos Database and Analysis Staff.2003. Part II. HIV-1/SIVepz com-plete genome alignments, p. 123–317.InT. Leitner, B. Foley, B. Hahn, P. A.
on November 8, 2019 by guest
http://jvi.asm.org/
Marx, F. McCutchan, J. W. Mellors, S. Wolinksy, and B. Korber (ed.), HIV Sequence Compendium 2003. Publication LA-UR 04-7420. Los Alamos National Laboratory, Los Alamos, N.Mex.
37.Mascola, J. R., A. Sambor, K. Beaudry, S. Santra, B. Welcher, M. K. Louder, T. C. VanCott, Y. Huang, B. K. Chakrabarti, W.-P. Kong, Z.-Y. Yang, L. Xu, D. C. Montefiori, G. J. Nabel, and N. L. Letvin.2005. Neutralizing antibodies elicited by immunization of monkeys with DNA plasmids and recombinant adenoviral vectors expressing human immunodeficiency virus type 1 pro-teins. J. Virol.79:771–779.
38.Mascola, J. R., M. G. Lewis, G. Stiegler, D. Harris, T. C. VanCott, D. Hayes, M. K. Louder, C. R. Brown, C. V. Sapan, S. S. Frankel, Y. Lu, M. L. Robb, H. Katinger, and D. L. Birx.1999. Protection of macaques against patho-genic simian/human immunodeficiency virus 89.6PD by passive transfer of neutralizing antibodies. J. Virol.73:4009–4018.
39.Mascola, J. R., S. W. Snyder, O. S. Weislow, S. M. Belay, R. B. Belshe, D. H. Schwartz, M. L. Clements, R. Dolin, B. S. Graham, G. J. Gorse, M. C. Keefer, M. J. McElrath, M. C. Walker, K. F. Wagner, J. G. McNeil, F. E. McCutchan, D. S. Burke, et al.1996. Immunization with envelope subunit vaccine products elicits neutralizing antibodies against laboratory-adapted but not primary isolates of human immunodeficiency virus type 1. J. Infect. Dis.173:340–348.
40.Mathewa, T. J.1994. Dilemma of neutralizing resistance of HIV-1 field isolates and vaccine development. AIDS Res. Hum. Retrovir.10:633–636. 41.Matsushita, S., H. Maeda, K. Kimachi, Y. Eda, Y. Maeda, T. Murakami, S.
Tokiyoshi, and K. Takatsuki.1992. Characterization of a mouse/human chimeric monoclonal antibody (C1) to a principal neutralizing domain of the human immunodeficiency virus type 1 envelope protein. AIDS Res. Hum. Retrovir.8:1107–1115.
42.Moore, J. P., Y. Cao, L. Qing, Q. J. Sattentau, J. Pyati, R. Koduri, J. Robinson, C. F. Barbas III, D. R. Burton, and D. D. Ho.1995. Primary isolates of human immunodeficiency virus type 1 are relatively resistant to neutralization by monoclonal antibodies to gp120, and their neutralization is not predicted by studies with monomeric gp120. J. Virol.69:101–109. 43.Muster, T., F. Steindl, M. Purtscher, A. Trkola, A. Klima, G. Himmler, F.
Ruker, and H. Katinger.1993. Conserved neutralizing epitope on gp41 of human immunodeficiency virus type 1. J. Virol.67:6642–6647.
44.Nabel, G. J.2001. Challenges and opportunities for development of an AIDS vaccine. Nature410:1002–1007.
45.Ofek, G., M. Tang, A. Sambor, H. Katinger, J. R. Mascola, R. Wyatt, and P. D. Kwong.2004. Structure and mechanistic analysis of the anti-human immunodeficiency virus type 1 antibody 2F5 in complex with its gp41 epitope. J. Virol.78:10724–10737.
46.Palker, T. J., M. E. Clark, A. J. Langlois, T. J. Matthews, K. J. Weinhold, R. R. Randall, D. P. Bolognesi, and B. F. Haynes.1988. Type-specific neu-tralization of the human immunodeficiency virus with antibodies to env-encoded synthetic peptides. Proc. Natl. Acad. Sci. USA85:1932–1936. 47.Park, E. J., L. K. Vujcic, R. Anand, T. S. Theodore, and G. V. Quinnan, Jr.
1998. Mutations in both gp120 and gp41 are responsible for the broad neutralization resistance of variant human immunodeficiency virus type 1 MN to antibodies directed at V3 and non-V3 epitopes. J. Virol.72:7099– 7107.
48.Pinter, A., W. J. Honnen, Y. He, M. K. Gorny, S. Zolla-Pazner, and S. C. Kayman.2004. The V1/V2 domain of gp120 is a global regulator of the sensitivity of primary human immunodeficiency virus type 1 isolates to neu-tralization by antibodies commonly induced upon infection. J. Virol. 78:
5205–5215.
49.Poignard, P., E. O. Saphire, P. W. Parren, and D. R. Burton.2001. GP120: biologic aspects of structural features. Annu. Rev. Immunol.19:253–274. 50.Robey, W. G., B. Safai, S. Oroszlan, L. O. Arthur, M. A. Gonda, R. C. Gallo,
and P. J. Fischinger.1985. Characterization of envelope and core structural gene products of HTLV-III with sera from AIDS patients. Science228:593– 595.
51.Roderiquez, G., T. Oravecz, M. Yanagishita, D. C. Bou-Habib, H. Mostowski, and M. A. Norcross.1995. Mediation of human immunodefi-ciency virus type 1 binding by interaction of cell surface heparan sulfate proteoglycans with the V3 region of envelope gp120-gp41. J. Virol.69:2233– 2239.
52.Sasaki, Y., Y. Ami, T. Nakasone, K. Shinohara, E. Takahashi, S. Ando, K. Someya, Y. Suzaki, and M. Honda.2000. Induction of CD95 ligand expres-sion on CD8⫹T-lymphocyte correlates with HLA-DR expression and
con-tributes to apoptosis of CD95-upregulated CD4⫹T-cells in macaques by infection with a pathogenic simian/human immunodeficiency virus. Clin. Exp. Immunol.121:1–10.
53.Sattentau, Q., and J. P. Moore.1991. Conformational changes induced in the human immunodeficiency virus envelope glycoprotein by soluble CD4 bind-ing. J. Exp. Med.174:407–415.
54.Scanlan, C. N., R. Pantophlet, M. R. Wormald, E. O. Saphire, D. Calarese, R. Stanfield, I. A. Wilson, H. Katinger, R. A. Dwek, D. R. Burton, and P. M. Rudd.2003. The carbohydrate epitope of the neutralizing HIV-1 anti-body 2G12. Adv. Exp. Med. Biol.535:205–218.
55.Seaman, M. S., L. Xu, K. Beaudry, K. L. Martin, M. H. Beddall, A. Miura, A. Sambor, B. K. Chakrabarti, Y. Huang, R. Bailer, R. A. Koup, J. R. Mascola, G. J. Nabel, and N. L. Letvin.2005. Multiclade human immuno-deficiency virus type 1 envelope immunogens elicit broad cellular and hu-moral immunity in rhesus monkeys. J. Virol.79:2956–2963.
56.Shinohara, K., K. Sakai, S. Ando, Y. Ami, N. Yoshino, E. Takahashi, K. Someya, Y. Suzaki, T. Nakasone, Y. Sasaki, M. Kaizu, Y. Lu, and M. Honda.
1999. A highly pathogenic simian/human immunodeficiency virus with ge-netic changes in cynomolgus monkey. J. Gen. Virol.8:1231–1240. 57.Shioda, T., J. A. Levy, and C. Cheng-Mayer. 1991. Macrophage and T
cell-line tropisms of HIV-1 are determined by specific regions of the enve-lope gp120 gene. Nature349:167–169.
58.Someya, K., D. Cecilia, Y. Ami, T. Nakasone, K. Matsuo, S. Burda, H. Yamamoto, N. Yoshino, M. Kaizu, S. Ando, K. Okuda, S. Zolla-Pazner, S. Yamazaki, N. Yamamoto, and M. Honda.2005. Vaccination of rhesus ma-caques with recombinantMycobacterium bovisbacillus Calmette-Gue´rin Env V3 elicits neutralizing antibody-mediated protection against simian-human immunodeficiency virus with a homologous but not a heterologous V3 motif. J. Virol.79:1452–1462.
59.Someya, K., K. Q. Xin, K. Matsuo, K. Okuda, N. Yamamoto, and M. Honda.
2004. A consecutive priming-boosting vaccination of mice with simian im-munodeficiency virus (SIV)gag/polDNA and recombinant vaccinia virus strain DIs elicits effective anti-SIV immunity. J. Virol.78:9842–9853. 60.Someya, K., Y. Ami, T. Nakasone, Y. Izumi, K. Matsuo, S. Horibata, K.-Q.
Xin, H. Yamamoto, K. Okuda, N. Yamamoto, and M. Honda.2006. Induc-tion of positive cellular and humoral immune responses by a prime-boost vaccine encoded with simian immunodeficiency virus gag/pol. J. Immunol.
176:1784–1795.
61.Speck, R. F., K. Wehrly, E. J. Platt, R. E. Atchison, I. F. Charo, D. Kabat, B. Chesebro, and M. A. Goldsmith.1997. Selective employment of chemokine receptors as human immunodeficiency virus type 1 coreceptors determined by individual amino acids within the envelope V3 loop. J. Virol.71:7136– 7139.
62.Trkola, A., H. Kuster, P. Rusert, B. Joos, M. Fischer, C. Leemann, A. Manrique, M. Huber, M. Rehr, A. Oxenius, R. Weber, G. Stiegler, B. Vcelar, H. Katinger, L. Aceto, and H. F. Gunthard.2005. Delay of HIV-1 rebound after cessation of antiretroviral therapy through passive transfer of human neutralizing antibodies. Nat. Med.11:615–622.
63.Trkola, A., T. Dragic, J. Arthos, J. M. Binley, W. C. Olson, G. P. Allaway, C. Cheng-Mayer, J. Robinson, P. J. Maddon, and J. P. Moore.1995. Cross-clade neutralization of primary isolate of human immunodeficiency virus type 1 by human monoclonal antibodies and tetrameric CD4-IgG. J. Virol.
69:6609–6617.
64.Trkola, A., T. Dragic, J. Arthos, J. M. Binley, W. C. Olson, G. P. Allaway, C. Cheng-Mayer, J. Robinson, P. J. Maddon, and J. P. Moore.1996. CD4-dependent, antibody-sensitive interactions between HIV-1 and its co-recep-tor CCR-5. Nature384:184–187.
65.Wu, L., N. P. Gerard, R. Wyatt, H. Choe, C. Parolin, N. Ruffing, A. Borsetti, A. A. Cardoso, E. Desjardin, W. Newman, C. Gerard, and J. Sodroski.1996. CD4-induced interaction of primary HIV-1 gp120 glycoproteins with the chemokine receptor CCR-5. Nature384:179–183.
66.Yamanaka, T., Y. Fujimura, S. Ishimoto, A. Yoshioka, M. Konishi, N. Narita, J. Mimaya, M. T. Meguro, T. Nakasone, Y. Okamoto, H. Yoshizaki, K. Yamada, and M. Honda.1997. Correlation of titer of antibody to principal neutralizing domain of HIV MN strain with disease progression in Japanese hemophiliacs seropositive for HIV type 1. AIDS Res. Hum. Retrovir.13:317–326. 67.York, J., K. E. Follis, M. Trahey, P. N. Nyambi, S. Zolla-Pazner, and J. H.
Nunberg.2001. Antibody binding and neutralization of primary and T-cell line-adapted isolates of human immunodeficiency virus type 1. J. Virol.
75:2741–2752.