0022-538X/85/060720-06$02.00/0
Copyright ©D1985, American Society for Microbiology
Identification of
a
New
Polypeptide Coded by Reovirus Gene
Si
GOBINDA SARKAR,' JERRY PELLETIER,' RHONDA BASSEL-DUBY,"2ANULAJAYASURIYA,2 BERNARD N. FIELDS,23'4AND NAHUM SONENBERGl 5*
Department of
Biochemistryl
andMcGill CancerCenter,5McGill University, Montreal, Quebec, Canada H3G I Y6, and DepartmentofMicrobiology and Molecular Genetics, Harvard Medical School,2DepartmentofMedicine, Division of Infectious Diseases, Brigham and Women's Hospital,3 andShipley Institute of Medicine,4Boston, Massachusetts 02115Received 7 December1984/Accepted 25 February 1985
The reovirus S1genehasrecentlybeenshownpotentiallytoencodetwopolypeptides (fromtwooverlapping reading frames) having predicted molecularweights of 49,071 and 16,143 (Nagata etal., Nucleic AcidsRes. 12:8699-8710, 1984; Bassel-Dubyetal.,Nature [London], in press). The largerpolypeptideisreovirusprotein
or1, butsynthesis of the smaller polypeptide hasnotbeendescribedtodate.Atruncated clone oftheS1gene
inwhichthefirst ATG is deletedwasexpressedinanin vitroproteinsynthesissystem toyielda-13-kilodalton
polypeptide,asdeterminedfrommigrationonsodiumdodecylsulfate-polyacrylamide gels.Apolypeptidewith asimilarmigrationpatternonsodiumdodecylsulfate-polyacrylamidegelswaspresentinreovirus-infectedcells and absent frommock-infected cells. Comparative tryptic peptide analysis of the 13-kilodalton polypeptides producedin vivoandinvitro showed themtobeidentical. Thus, thesl mRNA of reovirustype3isapparently bicistronic, and wesuggestthat the -13-kilodalton polypeptide be calledors(standing forcf small).
The reovirusgenomeconsistsof10double-stranded RNA
molecules which are transcribed into 10 mRNA species. Each mRNA has, until recently, been considered
function-ally monocistronic, thus resulting in the production of 10 functional or structural reovirus proteins. Recently, how-ever,studiesontranslationinitiationsitesofreovirusmRNAs by ribosome protection assays with cell-free systems re-vealed that one of the mRNA
species-si
mRNA-had twoinitiation sites (12), asdefined by the protection of RNA by 80Sribosomesagainstribonucleasedigestion (15). Sequenc-ing of the 5' end of the
Si
gene demonstrated two AUG codons separated by 55 nucleotides (17). Furthermore,Ce-natiempo et al. (4), using a dipeptide synthesis assay, showed that the
Si
gene directed the synthesis of twodifferent formylmethionyl dipeptides initiating at the two
5'-proximal AUGs in a reconstituted translation initiation
system. In addition, they demonstrated that the second AUGfromthe 5' endoftheslmRNAdirectedthesynthesis ofdipeptideswith50%efficiency,ascompared withthefirst AUG.
The process by which ribosomes bind to eucaryotic mRNAs and subsequently find the appropriate initiation codon isthoughttooccurviaa"bind andscan" mechanism
(10). This model states that43Spreinitiationcomplexes bind at or near the 5' end of the mRNA and subsequently slide
along the leaderregionuntil the appropriate (in general the
first)AUG isencountered,whereupona60Sribosome binds toformafunctional80Scomplex. Toexplaintheselection of
the appropriate AUGcodon forinitiation, Kozak(11),upon examination of the flanking sequences of initiator AUGs, found the consensus sequence GXXAUGG. However, a morerecentsurveyshowed less evidence for thepreference
ofguaninein theposition justdownstreamfrom the initiator AUG (14). The importance of the adenine 3 nucleotides upstream from the initiator AUG has now been demon-strated experimentally by mutational analysis (13). These
findingsargue thatthecontextof the initiator AUG codon is
important for its recognition as an initiation codon. An
analysis of theflanking nucleotides of the 5'-proximalAUG
*Correspondingauthor.
codons of reovirus mRNAs showed that the majority of
themcontainthe
GXXAUGG
consensus sequence(14).The5'-proximal AUGcodonofthe
Si
gene,however,residesin the unfavorable context CXXAUGG, whereas the second AUG resides in the context AXXAUGG. Consequently, it wasproposed by Kozak (12) that a certainproportion of the 40S ribosomal subunits might pass through the 5'-proximal AUG and starttranslationat thesecond AUG.Recently, the entire
Si
geneof reovirus type 3 (Dearing strain) was cloned and sequenced (19; R. Bassel-Duby, A.Jayasuriya, D. Chatterjee, N. Sonenberg, J. Maizel, and B.
Fields, Nature [London], in press). The
Si
geneconsists of 1,416 nucleotides and has two open reading frames. The 5'-proximal AUG is followed by an open reading frameencodingaprotein of455 aminoacids whichcorrespondsto theu1protein.ThesecondAUG from the 5' endis followed
by a short open reading frame potentially encoding a
120-amino-acidpolypeptide thatwould have amolecularweight of 16,143 (according to the sequence ofBassel-Duby et al.
[in press]). The two AUGs of the
Si
gene are in differentreading frames, thus potentially making the S1 gene
func-tionally bicistronic. In this communication, we provide
direct evidence that the
Si
gene,inadditiontocoding forthea 1 protein product, codes for a second
polypeptide
of-13-kilodaltons (kDa) in vitro and in vivo. Thus, we have identifieda new reoviruspolypeptidewhichmightbe
signif-icant in the reovirusreplicative cycle.
MATERIALSAND METHODS
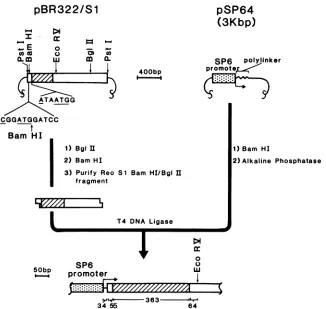
Molecularcloning.Aportionof thereovirus(type3,
Dear-ing strain)
Si
gene was subcloned into expression vectorpSP64(Fig. 1).PlasmidpSP64,obtainedfromD.Melton and M.Green(Harvard University, Cambridge, Mass.),contains the SP6 promoter, a polylinker region, and a selectable
ampicillin gene. A clone containing the full-length reovirus
Si
gene in thePstI siteofpBR322wasdigestedwith BamHI and BglII (Boehringer Mannheim Biochemicals,Indianapo-lis, Ind.). The
Si
BamHIIBglll
fragmentwasisolated froman agarosegelbytheglass powdertechnique
ofVogelstein
andGillespie(25).pSP64waslinearized withBamHI and treated with calf alkaline phosphatase. The two fragments were
720
on November 10, 2019 by guest
http://jvi.asm.org/
NEW REOVIRUS GENE PRODUCT 721
pBR322/S1
pSP64
(3Kbp)
400bp
I_
1) Bgl E 2) Bam HI
3) Purify Reo SI Bam HI/Bgl II f ragment
T4 DNA Ligase
4
SP6 polylinker promot
..Y
1)Bam HI
2) Alkaline Phosphatase
I
04 o
I
!,AL' 363 l..
34 55 64
FIG. 1. Schematicdiagramillustratingthesubcloning procedureforinsertion of theBamHI-BglII fragmentof thereovirus(Reo) S1gene
intotheexpressionvectorpSP64(see thetextfor details of the construction). RestrictionenzymeBamHIcleavesjustdownstream from the firstAUG. The hatchedboxdenotesthecoding regioninitiatingfromthe secondAUG;theflankingnucleotidesequencesof this AUGare
indicated. Thenumber of nucleotides derived frompSP64andfrom thecodingandnoncoding regionsof theS1geneisindicatedbelow the pSP64/S1construct. Kbp,Kilobasepairs;bp,basepairs.
ligated with T4 DNA ligase, and recombinant molecules containingtheS1geneintheorientationshown inFig.1were
chosen for further studies. All recombinant techniqueswere
carriedoutbasically asdescribedbyManiatis et al.(18)and in references therein.
Invitrotranscriptions andtranslations. Plasmid pSP64/S1
was linearized with EcoRV. In vitro transcriptions were
performed essentiallyasdescribedbyGreenetal.(8)except that the cap analog, m7GpppG (P-L Biochemicals Inc., Milwaukee, Wis.),wasincluded in the reaction mixture ata
concentration of 500 ,uM and the GTP concentration was
reduced to 20 ,uM. [3H]CTP (20 ,uCi, 25.4 Ci/mmol; New EnglandNuclearCorp., Boston, Mass.)wasincludedin the
reaction mixture to allow for quantitation of the mRNA. Transcription reaction mixtures were incubated for 2 h at
37°C, after which 2 ,ug ofRNase-free DNase I (Bethesda ResearchLaboratories, Inc., Gaithersburg, Md.) and1,ug of calf livertRNA(BoehringerMannheim)wereadded and the
mixturewasfurtherincubatedat37°C for 10min. The RNA
was extracted with phenol-chloroform, passed over aG50
spun-dialysis column, precipitatedtwice with 2.5 volumesof ethanol containing 2 M ammonium acetate, precipitated
oncewithethanol containing0.15 Mpotassiumacetate,and washed with 80% ethanol. In vitro translations were
per-formed inwheat-germ extractswith 8 ,ugofmRNApermlat 25C for 2 haspreviously described (22).
Infection of L cells with reovirustype3(Dearing strain). L cells insuspension weregrowntoaconcentrationof5 x 105
cells per ml in modified Eagle medium (Flow Laboratories Inc., McLean, Va.). Infection of
107
cells per ml with reovirus at 30 PFU per cell was performed as describedby Banerjee and Shatkin (1). Cells were labeled with [35S]methionine (>1,000Ci/mmol;NewEnglandNuclear) 20 h postinfection as follows. Cells were pelleted and sus-pended in methionine-free medium at 5 x 106 cells per ml. Cold methionine (50 ,uM)and 80 P,Ciof[35S]methionineper mlwereadded,and incubationwascarriedout at37°Cfor4 h. Afterbeinglabeled, cellswerepelleted,washed withcold phosphate-buffered saline, andlysedbyincubation(10 min)
in hypotonic buffer containing 10 mM N-2-hydroxyethyl-piperazine-N'-2-ethanesulfonic acid (HEPES) (pH 7.6), 10 mMpotassiumacetate, 0.75mM magnesiumacetate, and 2 mM dithiothreitol. Total extracts were mixed with sodium dodecyl sulfate(SDS) gel electrophoresis samplebuffer(16) andresolved by SDS-polyacrylamide gel electrophoresis.
Processing of [35S]methionine-labeled polypeptides for tryptic peptide mapping. After electrophoresis on a 10 to 18%SDS-polyacrylamide gel, carried outby the method of Laemmli (16), gelswere treated essentially as described by
Beemon and Hunter (2)but with some modifications. Gels were shaken in waterwith mixed-bed resin [AG 501-X8(D) (Bio-Rad Laboratories, Richmond, Calif.)] for 1 h, after which thegelsweredried andautoradiographed. Desiredgel pieces were cut out and swelled in 50 mM NH4HCO3, and the backing paper and front plastic were scraped off. Gel
pieceswerehomogenizedwithaPolytron homogenizer,and
I
L
5Obp- SP6 promoter
m
VOL.54,1985
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.146.472.64.373.2]the mixture was made 5% with ,-mercaptoethanol, boiled for 5min, and shaken overnight at 37°C to elute the protein. Gelfragments were pelleted at 10,000 xgfor10min,and the extracted protein in the supernatant was precipitated with 20% trichloroacetic acid with 100
jig
of bovine serumalbuminperml ascarrierat4°C for4h. Theprecipitatewas
pelletedat 12,000 x g for 15minat4°C, washedsuccessively
withcold ethanol and ethanol-ether (1:1), andincubatedwith 10 ,ul of chilled performic acid (prepared by mixing 50% H202and 98% formic acid [1:9] for 2 h at room temperature)
for2.5 h at4°C. The sample wasdilutedwith 1 mlofwater,
lyophilized,andredissolvedin 100 mMNH4HCO3. Tosylsul-fonyl phenylalanyl chloromethyl ketone-treated trypsin (50
,ug/ml) was added, and the sample was incubatedovernight
at37°C and lyophilized. The materialwasdissolved in10,ul
ofelectrophoresis buffer (butanol-pyridine-acetic acid-water [2:1:1:18, vol/vol], pH 4.7),spotted onCEL300(Polygram)
plates (20by 20 cm), andelectrophoresedat 800 Vfor50 min
essentiallyasdescribed by Gibson (6).Platesweredriedand chromatographed for 3 hinthe second dimension
(butanol-pyridine-acetic acid-water [97:75:15:60, vol/vol], pH 5.3). After chromatography, plates were dried, soaked in 7% 2,5-diphenyloxazole (PPO)in ether, dried again, and
autor-adiographed.
RESULTS
The 5'-proximal AUG ofreovirus sl mRNA directs the
synthesis of v 1, and the next AUG from the 5' end is followed byanopenreadingframe with acodingcapacity of
ca. 16 kDa (Bassel-Duby et al., in press). However, no
evidence has been presentedtodatetodemonstrate thatthe putative -16-kDa polypeptide is indeed synthesized during reovirus infection. We probed for the presence of the
-16-kDapolypeptide in reovirus-infected cellsdirectly and
compared it withthe polypeptide synthesized in vitro from
an mRNAderivedfrom a subclone of theSigene. Aportion
of the
Si
gene was subcloned next to theSP6promoter(Fig.1). A full-length clone of the
Si
gene was digested with BamHI and BglII restriction enzymes. Excision of the1.0-kilobase-pair
Si
fragment resultedinthelossofthefirstAUGcodon and 414 base pairs corresponding to the 3' end
ofthe mRNA. The5'-proximal AUGwas now55basepairs from the 5' end of the transcript, followed by the open
reading frame encoding the putative -16-kDa polypeptide. This fragment was then inserted into pSP64 to place
tran-scription under the control ofthe highly efficient SP6 pro-moter(8). The plasmid was thenlinearized withEcoRVand
utilizedas atemplatefor SP6 polymerasetoyieldmicrogram quantitiesof thetruncatedreovirussl mRNAforsubsequent functional assays. The transcripts produced in this system areuncapped and would not serve asefficient templatesfor in vitro translation in eucaryotic systems. However, by adding
m7GpppG
as a cap analogto prime transcription, ithasbeenshown that >90% of thetranscripts synthesized are
capped (20) and are efficientlytranslated in in vitro transla-tion systems.
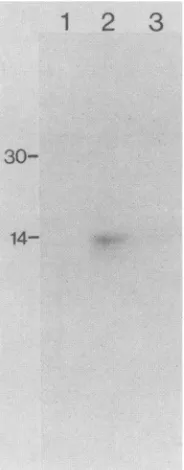
When the truncated
si
transcript was translated in a wheat-germ extract, a small polypeptide with an apparent massof-13 kDa was synthesized (Fig. 2). The control lane(lackingexogenous RNA, lane 1) shows that thisproteinwas not produced from endogenous mRNA in the extract, whereas the addition of the
si
transcript yielded the -13-kDapolypeptide (lane 2). Translation of the -13-kDapoly-peptide was dependent on the presence of a cap structure,
sincetranslation of the uncapped mRNA was -10-fold less efficient than thatof the capped mRNA, as determined by
12
3
30-
14-FIG. 2. Polyacrylamide gel electrophoresis of proteins synthe-sized ina wheat-germ extract programmed with invitro-transcribed truncated reovirus sl mRNA. Translations were done in 12.5-,ul reaction mixtures containing 0.1 ,ug of [3H]CTP-labeled mRNA (cappedoruncapped) made frompSP64/S1 asdescribed in thetext. Incubations were carried out for2h at25°C,at which time1
pR1
of the incubation mixture was withdrawn,mixed with electrophoresis buffer, andrunon a 10 to18%SDS-polyacrylamide gel; the gel was then treated with En3Hance (New England Nuclear), dried, and autoradiographed. Gelswereexposed to XAR-5 X-ray filmfor1h. Radioactive bands were quantified by soft-laser densitometry (LKB Instruments, Inc., Rockville, Md.). Lanes: 1,no mRNAadded; 2, cappedmRNAfrompSP64/S1;3,uncappedmRNAfrompSP64/S1. The numbers at the leftindicate kDa of markerproteins.laserdensitometry ofthe labeled bands (compare lane 3 to lane 2). Similarresults wereobtained when the mRNA was translatedin rabbitreticulocyte lysate (datanotshown).The apparent molecular weight of the synthesized polypeptide
(-13,000), as determinedfrom its migrationon a10 to18%
SDS-polyacrylamide gradient gel relativetothatofamarker
protein (lysozyme; molecular weight, 14,300), was lower
than the molecular weight predicted from the cDNA se-quence (molecular weight, 16,143). This somewhat anom-alous behavior could be attributed to clustering of hydro-phobic or charged amino acids in the polypeptide or to
post-translationalmodification. Henceforth, wewillreferto the -13-kDapolypeptide asus(standing forusmall).
Animportant question raisedby thesefindings is whether
theaspolypeptide is produced during reovirus infection.To try to answerthisquestion, weanalyzed a
[35S]methionine-labeled extract from reovirus-infected cells sideby side on anSDS-polyacrylamidegelwiththeaspolypeptide
synthe-sized in vitro in thewheat-germ system. Infection ofLcells in suspension with reovirus type 3 (Dearingstrain) led to a
drastic reduction in cellular
protein
synthesis(Fig.
3,com-parelanes 1 and3),aspreviously describedbyZweerink and
Joklik (26). Concomitantly, efficient synthesis ofthe three reoviruspolypeptide size classes
(X, pu,
anda)tookplacein the infected cellextract(lane 3).These results show that the cells were productively infected with reovirus and that authentic reovirus proteinswere synthesized. Inaddition, iton November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.390.482.70.305.2]NEW REOVIRUS GENE PRODUCT 723
2
3
.O" .1
or
14-FIG. 3. Comparative polyacrylamide gelanalysis of35S-labeled products from an in vitro translation reaction programmed with
mRNA transcribed from pSP64/S1 and ofextracts prepared from reovirus-infected L cells. Lanes: 1, extractfrom noninfected cells (106 cpm); 2, translation products from mRNA synthesized from pSP64/S1 (4 x 104 cpm); 3, extract from reovirus-infected L cells (106cpm). SDS-polyacrylamide gelanalysis wascarriedoutina 10
to 18% gradient gel, which was then dried (no En3Hance) and
exposedtoXAR-5X-rayfilmfor 15 h. Cell extractions and invitro translationswereperformedasdescribedin thetext. Thenumberat
theleft indicates kDaofmarkerprotein lysozyme.
is striking that the in vitro-synthesized a s protein (lane 2)
comigrated witha polypeptide synthesized in extracts from reovirus-infected cells (compare lanes 2 and 3). The latter polypeptide was not synthesized in extracts from mock-in-fected cells (lane 1). These results are consistent with the
possibility that a s is indeed synthesized from reovirus si
mRNA duringreovirus infection.
To determine whether the polypeptides comigrating in Fig. 3, lanes 2 and 3, are the same, wecarried out a tryptic
peptideanalysis of theextracted polypeptides. Aftertrypsin digestion,the resultantpeptideswereresolved by
electropho-resis in the first dimension and chromatography in the second dimension on CEL 300 plates. Figure 4 shows the
autoradiograms of the [35S]methionine tryptic mapsof the cr spolypeptide synthesizedin vitro (Fig. 4A), theputativev s
polypeptide synthesized invivo (Fig. 4C), and amixture of
the trypsin digestion products of the two polypeptides (Fig. 4B). The amino acid sequence ofa s, as deduced from the
nucleotide sequence of the Si gene (Bassel-Duby et al., in press),predicted that with[35S]methionine asthe label, four
peptides shouldbegeneratedbytrypsindigestion. In both in vitro and in vivo preparations, three peptides were of a
relatively strongintensity (indicated by arrowheads; one of
the three peptides was labeledmore strongly than the other
two, most probably because it contained two methionines,
as predicted from the DNA sequence), whereas a fourth
peptide was less intense (indicated by arrows; we do not
have an explanation for the low relative intensity of this
peptide). In addition, severalweaklylabeledpeptides, which
were identicalin bothpreparations, were present (probably incomplete digestion products). Thus, it is clear from the
peptide
map analysis that the -13-kDa polypeptidepro-duced in reovirus-infected L cells is very similar, if not identical to, crs synthesized in vitro.
The results presented here are consistent with the notion thatreovirus sl mRNA is functionally bicistronic and that during infection in L cells it is translated to yield two
polypeptides, or 1 and cr s. However, it is also plausible,
althoughunlikely, thatininfected cells the reovirus Si gene canin some way generate two distinct mRNAs, one
direct-ingthe synthesisofcr1and the other directing the synthesis
ofgs.Thispossibility could be excluded only by performing
Si mapping or primer extension experiments with reovirus mRNAspurified from infected cells. However, to determine
ifthe full-length sl mRNA could be translated in vitro to yield both a 1 and C s, we subcloned the entire Si gene into plasmid pSP64 and synthesized a full-length sl mRNA
containingthe AUG codons for both thea1 and a s proteins.
Translation of this mRNA in rabbit reticulocyte lysate yielded both a 1 and a s (data not shown), indicating that translation from sl mRNA can initiate from the two 5'-proximal AUG codons. Thus, in vitro slmRNAbehaves as abicistronic mRNA.
DISCUSSION
The existence of a newly synthesized low-molecular-weight polypeptide after reovirus infection of L cells has been previously demonstrated (27). In addition, the transla-tion of reovirus mRNAs in in vitro systems often yields polypeptides that migrate fasterthan the a3 protein(see, for example,reference 24). Thequestion as to whethertheyare bonafide translationproducts orsimplyanartifactof in vitro translation systems (i.e., early termination products) has been difficult to answer.
In light of our results, reovirus sl mRNA appears to be bicistronic, as predicted by the determination of mRNA translation initiation sitesprotected by80S ribosomes against
RNase digestion (12) and by formylmethionyl dipeptide
synthesis from sl mRNA (4). The ability of a eucaryotic mRNA to code for more than onepolypeptide is not unique
to reovirus sl mRNA. However, the majority of mRNAs analyzed to date that do so use a second and sometimes a third AUGdownstreamfrom but in phase with thefirst AUG
(herpes simplex virus type 1 thymidine kinase mRNA, for example; see reference 21). Thus, in these cases, the result-antpolypeptide is a truncated versionof the primary protein
product. However, in several cases,viraleucaryotic mRNAs have been shown to encode polypeptides which are
synthe-sized from two different reading frames. These examples
include the following: (i) an mRNA of adenovirus early
region 1B that codes for two tumorantigens (3); (ii)the 16S late mRNA ofsimian virus 40encoding the agnoprotein and VP-1 (9); (iii) RNA segment 6 of influenze B viruscodingfor neuraminidase and aglycoprotein (23);and (iv)anmRNA of
Sendai virus that contains two overlapping open reading
frames that encode the P and C proteins (7).
The presence of the cr s protein in reovirus-infected cells
raises many interesting questions concerning its possible
function in the viral replication cycle and in the mechanism andregulation ofas expression. Itwill also beof interestto determine if reovirus types 1 and 2 express a
us-like
polype-ptide. We have found that the truncated reovirus sl mRNA is dependent on the cap structure for translation in vitro. This is consistent with a normal initiation mechanism in which a 40S ribosome binds near the mRNA 5' end and subsequently relocates to the 5'-proximal AUG. However,
we cannot conclude that this is how the a s polypeptide is VOL. 54, 1985
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.110.248.69.293.2]724 SARKAR ET AL.
A.:
. .
:bl1i
.:
..
.. ... ...
*:
*: ... : ::
*: *:; .:
.. :. *.
.:
.:
.. ..
.. .. ...
.4...
*..:.::
.....s.
... ... .<II
a.
C
FIG. 4. Comparativetryptic peptide map analysis ofthe cr spolypeptide synthesized in vitro and the -13-kDa polypeptidesynthesized in reovirus-infectedLcells. Bothpolypeptides were resolvedon a 10 to18%SDS-polyacrylamide gradient gelasdescribed in the legendtoFig. 3, and therespective bandswereeluted from the dried gel slice and processedasdescribedinthe text;-9,000 cpmwerespottedontoCEL 300 plates. For mixing experiments, -5,000 cpm each from in vivo and in vitro digestion products were spotted together. After electrophoresis and chromatography, plates wereexposedtoXAR-5X-rayfilm for2days. (A)crspolypeptide; (B) rspolypeptideplus the -13-kDa polypeptide fromreovirus-infected L cells; (C) -13-kDa polypeptide from reovirus-infected Lcells.
produced fromthebicistronicsl mRNA in vivo.For
exam-ple, it ispossiblethatribosomesinitiate internallyand hence
by a novel mechanism. It will be of great importance to
examine relativeexpression from thetwo slcistronsinvivo
andinvitro withextractsfrom eithernoninfected or infected
cellstogain insight intothese possibilities.
The construction of reovirusmutantsin orsby manipula-tion ofthe cloned Si gene may allow elucidation of the
putative role ofcrs inreovirusreplication.
We haverecently learnedthat Ernstand Shatkin(5)found that translationof reovirussl mRNAin vitroyielded theu1
proteinand alow-molecular-weight polypeptide (-14 kDa) which most probably corresponds to the cr s protein.
Fur-thermore, they foundthat alow-molecular-weight polypep-tide of-14kDa ispresentinreovirus-infected extracts but not in mock-infected extracts and that it shares common
peptideswith the invitro-synthesized-14kDapolypeptide. Thus, ourresults are inagreement with these findings.
ACKNOWLEDGMENTS
Weareindebted to Sheelin Howard and Christiane Babin for their excellenttechnical assistance. WethankD. Melton andM.Green for providing us with plasmid pSP64 and Kevin Lee for helpful discussions and comments onthemanuscript. Wethank H. Ernst and A. J. Shatkin forcommunicating their datato usbefore publi-cation.
This workwassupported byagrantfrom the MedicalResearch Council of CanadatoAngus Graham and N.S. N.S. isarecipient of aTerry Fox Cancer Research ScientistAward from the National Cancer Institute of Canada. J.P. is a recipient ofa predoctoral studentship from the Medical Research Council of Canada, and R.B.-D. is arecipient ofanFCAC Fellowship (Gouvernement du
Quebec).
LITERATURECITED
1. Banerjee,A.K.,and A.J.Shatkin.1970.Transcriptionin vitro by reovirus-associated ribonucleic acid-dependent polymerase. J. Virol.6:1-11.
2. Beemon, K., and T. Hunter. 1978. Characterization of Rous sarcomavirussrcgeneproducts synthesized in vitro. J. Virol. 28:551-556.
3. Bos, J. L., L.Polder,R.Bernards, P.Schrier, P. van den Elson, A. van der Eb, and H. van Ormondt. 1981. The 2.2 kb Elb mRNAofhumanAdl2 and Ad5 codes for two tumor antigens startingatdifferent AUGtriplets. Cell 27:121-131.
4. Cenatiempo, Y.,T.Twardowski,R.Shoeman,H.Ernst,N.Brot, H. Weissbach, and A.J. Shatkin. 1984. Two initiation sites detected in the smallslspecies of reovirusmRNAby dipeptide synthesisin vitro. Proc. Natl. Acad.Sci. U.S.A. 81:1084-1088. 5. Ernst, H., and A. J. Shatkin. 1985. Reovirus hemagglutinin mRNA codes for two polypeptides in overlapping reading frames. Proc.Natl. Acad. Sci. U.S.A. 82:48-52.
6. Gibson,W. 1974. Polyoma virusproteins: adescription of the structural proteins of the virion based on polyacrylamide gel electrophoresis and peptide analysis. Virology 62:319-336. 7. Giorgi, C.,B. M.Blumberg,and D. Kolakofsky. 1983. Sendai
virus containsoverlapping genes expressed fromasinglemRNA. Cell 35:829-836.
8. Green, M. R., T. Maniatis, and D. A. Melton. 1983. Human
P-globinpre-mRNAsynthesized invitroisaccuratelyspliced in Xenopus oocyte nuclei.Cell 32:681-684.
9. Jay, G., S. Nomura, C. W. Anderson, and G. Khoury. 1981. Identification of the SV40 agnogene product: a DNAbinding protein.Nature(London)291:346-349.
10. Kozak,M.1981.Mechanism ofmRNArecognition by eukaryo-tic ribosomesduring initiation ofprotein synthesis. Curr. Top. Microbiol. Immunol. 93:81-123.
11. Kozak,M. 1981.Possible role offlanking nucleotides in recog-nition of the AUG initiator codon by eukaryotic ribosomes. Nucleic AcidsRes. 9:5233-5252.
12. Kozak,M. 1982.Analysis of ribosome binding sites from thesl message of reovirus: initiation at the first and second AUG codons.J. Mol. Biol. 156:807-820.
13. Kozak, M. 1984. Pointmutations close to the AUG initiation codon affect theefficiency of translationof ratpreproinsulinin vivo.Nature(London)308:241-246.
14. Kozak, M. 1984. Compilation and analysis of sequences up-streamfromthe translational start site ineukaryoticmRNAs. Nucleic Acids Res. 12:857-872.
15. Kozak, M., and A. J. Shatkin. 1976. Characterization of ribo-some-protected fragments from reovirus messenger RNA. J. Biol. Chem. 251:4259-4266.
16. Laemmli,U.K.1970. Cleavage of structuralproteinsduringthe assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
17. Li, J. K.-K., J.D.Keene,P. P.Scheible,and W. K.Joklik.1980. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
NEW REOVIRUS GENE PRODUCT
Nature of the 3' terminal sequences of the plus and minus
strands of theslgeneof reovirusserotypes1, 2 and 3.Virology 105:41-51.
18. Maniatis, T., E.F. Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual.Cold SpringHarbor Laboratory,
ColdSpring Harbor, N.Y.
19. Nagata, L.,S. A. Masri, D. C. W. Mah, and P. W. K. Lee. 1984. Molecular cloning and sequencing of the reovirus(serotype 3) Si gene which encodes the viral cell attachment proteina 1.
Nucleic Acids Res. 12:8699-8710.
20. Pelletier, J., andN.Sonenberg. 1985. Insertion mutagenesisto increase secondarystructurewithin the 5'noncoding regionofa
eukaryotic mRNA reduces translational efficiency. Cell 40: 515-526.
21. Preston, C. M., and D. J. McGeoch. 1981. Identification and mapping oftwopolypeptides encodedwithin theherpes simplex virus type 1 thymidine kinase gene sequences. J. Virol.
38:593-605.
22. Roberts, B. E., and B. M. Paterson.1973.Efficient translation of
tobacco mosaic virus RNA and rabbit globin 9SRNAinacell
free system from commercial wheat-germ. Proc. Natl. Acad. Sci. U.S.A.70:2330-2334.
23. Shaw, M., P. Choppin, and R. Lamb. 1983. A previously unrecognized B virus glycoprotein from a bicistronic mRNA
that also encodes the viralneuraminidase. Proc. Natl. Acad. Sci. U.S.A. 80:4879-4883.
24.Sonenberg, N., D. Skup, H. Trachsel, and S. Millward.1981.In vitro translation in reovirus- and poliovirus-infected cell
ex-tracts.J. Biol. Chem. 256:4138-4141.
25. Vogelstein,B.,andD.Gillespie.1979.Preparativeandanalytical purification of DNA from agarose. Proc. Natl. Acad. Sci.
U.S.A. 76:615-619.
26. Zweerink, H. J., and W. K. Joklik. 1970. Studies on the intracellularsynthesis ofreovirus-specified proteins. Virology 41:501-518.
27. Zweerink, H. J., M. J. McDowell, and W. K. Joklik. 1971. Essential and nonessentialnoncapsidreovirusproteins.
Virol-ogy45:716-723.
VOL.54, 1985 725