JOURNALOFVIROLOGY, June1986, p.757-764 Vol. 58, No. 3
0022-538X/86/060757-08$02.00/0
CopyrightC) 1986,AmericanSocietyforMicrobiology
Localization and Fine Structure of
a
Vaccinia Virus Gene Encoding
an
Envelope Antigen
PIERRE HIRT,' GERHARD HILLER,'tANDRICCARDO WITTEKL*
InstitutdeBiologieAnimale, Universitede Lausanne, CH-1015 Lausanne, Switzerland,' and Max-Planck Institutefor
Biophysical Chemistry,
D-3400Gottingen,
FederalRepublic
ofGermany2Received 3 February1986/Accepted 10 March 1986
Themajor antigen on theenvelopeof extracellular vaccinia virusparticlesisapolypeptidewithanapparent molecular weight of 37,000 (p37K; G. Hiller and K. Weber, J. Virol. 55:651-659, 1985). Thegeneencoding
p37K was mapped in the vaccinia virus genome by hybrid selection of RNA followed by in vitro translation. p37K was then identified among the in vitro translation products by immunoprecipitation witha monoclonal antibody. The gene is located closetotheright-hand end of theHindlllFfragment. Thecorrespondingregion
of the DNAwassequenced, andanopenreading frame encodingapolypeptide of 41,748 daltonswasobserved. The 5' end of the mRNA, as defined by nuclease Si analysis, maps within only a few nucleotides of the translation initiation codon. Examination of the DNA sequence around the putative initiation site of transcription revealed a characteristic sequence, TAAATG, which includes the ATG translation initiation codon and whichisconserved in all butonelate genesofaranalyzed. It is therefore likely that thissequence
isanimportantregulatory signal for lategeneexpressionin vaccinia virus.
Vaccinia virus is a member of the poxvirus family. The
virionsarecharacterized bytheir large size and typical brick
shape. The genome of vaccinia virus consists of a linear,
double-stranded DNA molecule ofca. 190 kilobase pairs.
This large amount of genetic information is expressed in a
temporally regulated fashion. Earlygenes aretranscribedby
the virion-associated RNA polymerase before complete
uncoating ofthe viral genome. After DNAreplication, late
genes encoding predominantly structural polypeptides are
expressed (14).
In contrast to most other animalDNAviruses, the
multi-plication cycle of vaccinia virus is confinedtothe cytoplasm
ofthe infected host cell, where replication occurs within
discrete cytoplasmic foci termed "factories" (4). Progeny
virus is assembled within or close to the factory areas in a
complex morphogenesis process(reviewed in reference5).
Uncertainty exists with respect to the mechanism of
release ofprogeny virus, a process which is inefficient in mostcell culture systems used for the propagation of
vac-cinia virus. Most investigators therefore recover progeny
virus after experimentally induced cell lysis, which yields
large amounts of infectious virus particles. However, virus
naturally released from the living cell (extracellular virus)
differs antigenically fromvirusrecovered fromtheinterior of
thecell (1).
Recently, cell cultures and virus strains which produce
sufficient amounts ofextracellular virus to permit detailed
biochemicalanalysis havebeen described(17). The
charac-teristic feature of extracellular virus is the presence of an
envelope (1, 24) containing several virus-specific antigens (16, 17), the majorof which is a polypeptide with a molecular
weight of 37,000 (p37K; 8). The polypeptide has been
classifiedasalateone, since it isonly expressedin cells after
the onsetofviral DNAreplication(9).
*Correspondingauthor.
tPresent address: Boehringer GmbH, D-6800 Mannheim 31, FederalRepublic of Germany.
757
In uIuO
IN UITRO- - aE
TOT IPP IPP TOT
_
U
U,i---S
FIG. 1. Immunoprecipitation ofp37Ksynthesizedin vivo and in vitro.Vaccinia virus-infected cell monolayers were labeledfor3 h with[35S]methionine and [35S]cysteineat 4h after infection. Total cell lysates were analyzed either directly (lane TOT) or after immunoprecipitationwith theanti-p37K antibody (lane IPP). Total RNAfrom vaccinia virus-infected cells was translated invitro,and the products were analyzed either directly (lane TOT) or after immunoprecipitation with the anti-p37K antibody (lane IPP). A
fluorograph ofa14% polyacrylamide gel isshown.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:1.612.370.514.347.636.2]758 HIRT ET AL.
A
H i nd fragments
2 35
C NMK F E 01 G LJ H D
4 5
A
I
111
Ii
[11 1
11
L
10 kb
1 2 3 4 5 m m N N K F
.
.
'
~~~~~200-So"'I"'4
_ 9 -U.*Rrn.
II
U
,- 69
,-46
U
U
up
b
C
FIG. 2. Gel analysis of immunoprecipitated cell-free translation products. Lanes 1 to 5 and N, M, K, and F in panels B and C show the immunoprecipitated polypeptides made by RNA selected on pools of restriction fragments or on individual restriction fragments,
respectively, asindicatedintheHindlIl map of the vacciniavirus DNA shown in panel A. The arrows indicate the positions of twoSallsites within the unique sequence of theHindlll B fragment. Afluorograph of a 14% polyacrylamide gel is shown. The molecular weights (in thousands) of polypeptides used as size markers (lanes m) are indicated. kb, Kilobases.
Amonospecific antibodyhasrecentlybeen usedtofurther
characterize p37K and also to study the mechanism by
whichvaccinia virusacquiresits envelope (9). After
synthe-sis, p37K, which is unglycosylated but contains palmitic
acid, accumulates in Golgi membranes. These membranes
with viral protein(s) incorporated are then used to enwrap intracellular vaccinia virus with a double-walled coat (9).
These virus-carryingvesicles then appeartobetransported to the cell surface, wheretheiroutermembrane fuses with
the cytoplasmic membrane, thus releasing a virus particle
surroundedby asinglemembrane.
Inthiscommunicationwereportthemapping ofthe gene
encoding p37K on the vaccinia virus genome. We also
presentthe fine structure of the gene and itsflanking regions.
MATERIALS ANDMETHODS
Virus and cells. The WR strain of vaccinia viruswasused inthisstudy. Rabbitkidneycells(RK-13) and African green
monkey kidney cells (CV-1) were grown in Eagle minimal
essentialmediumsupplementedwith5%fetal calfserumand inDulbecco modifiedEaglemediumsupplementedwith10% fetal calf serum, respectively. Cells were infected with 10 PFUof vaccinia virus per cell.
Monoclonalantibodyeevl. The material usedfor immuni-zationconsisted oftheBrij-solubleandtrichloroacetic
acid-precipitable components of crude extracellular vaccinia
vi-rus preparations. Spleencells from successfully immunized
Lourats werefused with theratmyelomaline YB2/0(11)to J. VIROL.
301.11
1-1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.146.465.71.507.2]VOL.58,1986~~~~~~~VACCINIA VIRUS ENVELOPE ANTIGEN 759
A
Hind III F
9~~~~~~~~~~O r9C F
1 2 3 4 5 6 7 6
1 kb
1 2 34'5 67 6 m 5 6 7 8 m
" '200"W
S.
92
Sa
U,',W
l..-iI"
46
350
iMP
[image:3.612.133.488.71.460.2]B
C
FIG. 3. Immunoprecipitationof invitro-translatedpolypeptides.TotalRNAwasselectedby hybridizationtoelectrophoretically separated DNArestrictionfragments(B)ortosubcloned DNAfragments (C).Lanes 1to8 show theimmunoprecipitated polypeptides obtained from RNAselectedonthe DNAsegmentsindicated inpanelA. Afluorographofa14%polyacrylamide gelis shown. The molecularweights(in thousands)ofsizemarkers (lanes m)areindicated. Symbols: IHindIII; F' Sall; F. EcoRI;~,BamHl.
createhybridomasinaccordance with theinformationgiven in a recent review (7). A selected clone produced eev1 antibodies,whichspecificallyandselectively immunoprecip-itated the previously recognized acylated p37K of extracel-lular virus (manuscriptinpreparation).
For immunoprecipitation experiments purified eev1 anti-bodies were covalently coupled to Sepharose (Pharmacia,
Uppsala, Sweden) by standardprocedures.
Preparationof labeled cell extracts. Monolayer cultures of CV-1 cellsin 3-cmdisheswereinfected with vacciniavirus.
At 4 hafter
infection,
thegrowth
mediumwasreplaced' by
1ml of medium
containing
1/10 of theregular
methionineconcentration and 20 p~Ci each of
[35S]methionine
and[35S]cysteine.
After an additional 3 h ofincubation, cells were washed withphosphate-buffered
saline and thenlysedby
theaddition of 0.5mlof0.0625 M Trishydrochloride (pH6.8)-3% sodium dodecyl sulfate-15% glycerol-1%
2-mercaptoethanol-0.015% bromophenol
bluefor subsequent analysisofpolypeptides
bySDS-polyacrylamide
gelelectro-phoresis. For
immunoprecipitation
cells were lysed by theaddition of1 ml of10 mM Tris hydrochloride (pH 7.4)-l%
Triton X-100-0.5% sodium
deoxycholate-0.1%
SDS-150mMNaCl-1 mM EDTA. After3 hat
40C,
insoluble materialwas removed
by
centrifugation
at 100,000 X gfor 90 minat40C.
Imimunoprecipitation. Polypeptides
wereimmunoprecipi-tated from tota cell extracts or from in vitro translation
mixtures asdescribed previously(28).
RNA extraction. Earlyand late RNAs were purifiedfrom vaccinia virus-infected cellsasdescribed in detail elsewhere
(28).
Hybrid selection ofRNA. RNAspecific todefinedregions
of the vaccinia virusgenomewas
purified by hybridization
tocloned DNA restriction fragments as described previously
(28).
In vitro translation. Total and
hybrid-selected
RNAswere translated inamessage-dependent
rabbitreticulocyte lysate assay as describedpreviously
(28).Nuclease Si analysis. The procedure for
mapping
RNA transcripts by nuclease Si analysis (2) with 5'-end-labeled DNA fragments as hybridization probes (25) has beende-scribedpreviously (28).
DNAsequencing.DNAfragmentswerecloned into
single-stranded
phage
vectors (13) and sequencedby
the chainVOL.58, 1986
.1'..-.4,: 4..
on November 10, 2019 by guest
http://jvi.asm.org/
760 HIRT ET AL.
A mRNA p37 k
L E L
F~ F¶ - f
91g--250 bp
3. a
41.
B
1
2 33
4
-m p
t
e I p I pt
Ie
m p I p pI
Es~~~~~~~~~~~~~~~~~__
622
d
622 45
527
4S
527 *
403 am
_ __ 403
309
3093
242 238 f
217 to 201t
242
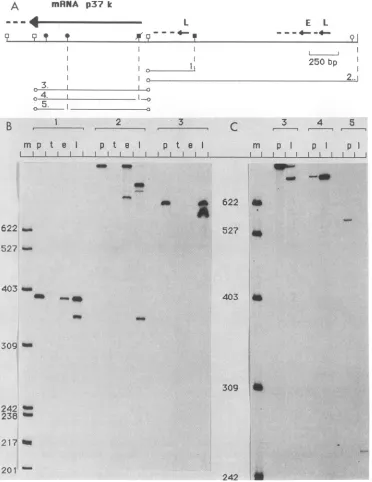
FIG. 4. Nuclease Si mappingof RNAtranscripts. (A)ADNAsegmentfrom theright-handHindIIIsite of theHindIIIFfragmenttothe
extremeleft-handSallsitewasused fortranscriptional mapping.Variousfragmentsderivedfrom thisregionwerelabeled at the 5' end(open circles) and usedashybridization probes. Eachprobe(1to5)wasanalyzedeitherdirectly (BandC,lanesp)orafterhybridizationtotRNA
(B, lanes t) or to early (B, lanes e) or late (B and C, lanes 1) RNA from infected cells. Si-resistant material was analyzed on a 6%
polyacrylamide sequencing gel.The sizes(in bases)oflengthmarkers(lanes m)areindicated.Symbols:
I,
HindIII; y,Sall; ?,BglII;-A,HincII; I, ClaI; y,EcoRI. (NotethatonlytheBgII, HincII, andClaIsitesofparticularinterestareindicated;this regionof the DNA
contains additional restriction sites for theseenzymes.) Themapposition and direction oftranscriptionof thep37K mRNA,aswellasof
additionalearly (E) and late (L) RNAs,is shownbythearrows.The broken lineatthetipof eacharrowindicatesthatthe3' end of theRNA hasnotbeenmapped;vertical broken lines indicate the relative positionsofparticularrestriction sites in theappropriate Si probe. termination method (21). In some cases, the chemical
deg-radationmethodwas used(12).
RESULTS
Immunoprecipitation of in vitro- and in vivo-synthesized p37K. To identify p37K, a characteristiccomponent of the
envelopemembrane of extracellular vacciniavirus,wemade
use ofa newly establishedrat monoclonal antibody (eevl) whichselectivelyreactswith thispolypeptidein
immunopre-cipitationaswellasin Westernblotexperiments (G. Hiller, manuscriptinpreparation).
When synthesized in vivoin the presence ofradioactive palmitic acid, p37Kwaslabeled(9). Acylationofproteinsis
aposttranslationaleventwhich does notoccurincelllysates
J. VIROL.
am
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.127.499.72.554.2]VACCINIA VIRUS ENVELOPE ANTIGEN 761
used for in vitro protein synthesis. We therefore examined
the ability of monoclonal antibody
eev1
to react with in vitrotranslation products directed by total RNA from vaccinia
virus-infected
cells.
As a control, the in vivo-synthesizedpolypeptide
was
immunoprecipitated from infected cellex-tracts in parallel. In both cases, a single polypeptide of the expected size was detected (Fig. 1). The in vitro translation product showed a slightly increased electrophoretic mobility as compared with the in vivo-synthesized polypeptide; this increase can best be explained by the lack of posttransla-tional modification of the in vitro-synthesized product.
Mapping of the p37K gene within
HinduI
restrictionfrag-ments. RNA from vaccinia virus-infected
cells
was selectedby hybridization to cloned
HindIII
restrictionfragments
ofthe vaccinia virus genome. Five nitrocellulose membranes containing the indicated fragments (Fig. 2A) were used in a first experiment. After hybridization, the filters were
washed, and
bound
RNA was then eluted and translated.After immunoprecipitation, the products were analyzed by
polyacrylamide
gelelectrophoresis
(Fig. 2B). The mRNAencoding p37Kwasclearly selected byfilter1containing the
HindIII
fragments N, M, K, and F. These fragments were therefore used separately to select RNA, and the polypep-tide was obtained from RNA selected on the HindIII Ffragment (Fig.
2C).
Localization of the p37K gene within the
HindIll
Ffrag-ment. To further map the gene, we prepared a detailed restriction map of the HindIII F fragment (ca. 13.5 kilobase pairs) (Fig. 3A). The recombinant plasmid was cleaved with
EcoRI,
BamHI, HindIII, andSalI,
and the resultingfrag-ments were then separated by gel electrophoresis and trans-ferred to nitrocellulose membranes. The membranes were cut such that each piece contained a given restriction frag-ment. The DNA was used for hybrid selection of RNA. Selected RNA was translated in vitro, and p37K was
iden-tified by immunoprecipitation (Fig. 3B). A
SalI
fragment ofca. 840 base pairs (bp) (Fig. 3B, lane 7) efficiently selected the mRNA. Smaller amounts of the mRNA were also
selected on a
SalI
fragment of 230 bp (Fig. 3B, lane 6), andtrace amounts were selected on all other fragments. Since about 1,000 nucleotides are sufficient to encode a
polypep-tide with a
molecular
weight 37,000, it is somewhatsurpris-ing that the mRNA was selected on such a long stretch of
DNA. This result may be
explained
at least in part by thefact that
late-infection
vaccinia virus transcripts are veryheterogeneous in length, do not contain discrete 3' ends (27), and may exceed in length the minimal coding requirement severalfold. However, some cross-contamination of restric-tion fragments or inaccuracy in cutting the membranes or both cannot be excluded. A small polypeptide was also immunoprecipitated from the in vitro translation products of RNA selected on fragment 2. This polypeptide presumably
has an antigenic
site
in common with p37K and is apparentlydetectable only when its mRNA is sufficiently enriched. The experiment nevertheless clearly indicated that the
coding sequence of
p37K
is located at the right-hand end ofthe
HindIII
F fragment. Four DNA fragments from this region were therefore subcloned, and equimolar amounts ofeach recombinant
plasmid
wereimmobilized
onnitrocellu-lose filters and used to select RNA. After in vitro translation,
p37K was
immunoprecipitated
(Fig.3C).
Again, a largeamount of p37K was obtained from RNA selected on the
SalI
fragment of ca. 840 bp (Fig. 3C, lane 7), andsucces-sively decreasing
amounts
were obtained from RNAse-lected on the adjacent left-hand fragments. Thus, hybrid selection of RNA clearly showed that a large portion of the
HIND III FRAGMENTS
F
L...j 10 kb
4-Ikb
2 840 Sal
Sal
I It ~ ~~~~~~IXba
Alu
I Dra
Bgl
---I A Sau
IOO bp
A 40 4-3 u WA do
B. 4 °0
FIG. 5. Sequencing strategy. A detailed restriction map of the DNA that was sequenced is shown. Various restriction fragments
were either cloned into single-stranded phage vectors and se-quenced by the chain termination method (A) or labeled at the 5' end (circles at end of arrows) and sequenced by the chemical degrada-tion method (B). The direcdegrada-tion and extent of sequencedetermination
are indicated by arrows. kb, Kilobases.
coding sequence of p37K maps within aSalI fragment of840
bp located near the right-hand end of theHindIlI F fragment.
Mapping of the p37K mRNA. The p37K mRNA was
mapped by nucleaseS1 analysis. Five 5'-end-labeled
frag-ments derived from the region to which the p37Kgene had
been mapped by hybrid selection of RNA were used as
hybridization probes (Fig. 4A). Each probe was analyzed
directly (Fig. 4B and C, lanes p) or hybridized totRNA as a
control (Fig. 4B, lanes t) or to early (Fig. 4B, lanese) or late
(Fig. 4B and C, lanes 1) RNA isolated from vaccinia
virus-infected cells. After hybridization, single-strandedDNA was
digested with nucleaseS1, and resistant material was
ana-lyzed by gel electrophoresis.
With probe 1, a SalI-ClaI fragment, the entire probe was
protected with both early and late RNAs. In addition, late
RNA also protected a fragment of 360 nucleotideswhichwas
also obtained with the longer probe 2 butwhich was labeled
at the same end. The map position of this late RNAisshown
in Fig. 4A. It is unlikely that this transcript represents the
p37K mRNA, since a cloned fragment from this region was
negative in the hybridization selection procedure (Fig. 3C,
fragment 8). The map positions of additional major earlyand
late RNAs detected with probe 2 are also indicated in Fig.
4A. With probe 3, the fragment which very efficiently
selected the p37K mRNA, late RNA protected the entire
probe and, in addition, a smaller fragment of ca. 750
nucle-otides. Since probe 3 was labeled at both ends, it was not
possible to determine the direction of transcription of this
late RNA, which most likely represents the p37K mRNA.
Probe 3 was therefore cleaved asymetrically with either
HincII (probe 4) orBglII(probe 5; note thatonly one BglII
site had been cleaved). The digested fragments (without
isolating the individual fragments) werethenanalyzed either
VOL.58, 1986
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.316.554.70.323.2]762 HIRT ET AL.
I 10 20 30 40 50 60 70 80 5'
OTCOACTTTORT0RRRRTTTTROCORTRTROCCORTOATRTTCTRORTCOTTORTRGRRCRAORTOTATRROTTTTTRTOTTRRCTRA
89 99 109 119 129 139 149 159 169 179
ATO TOO CCR TTT OCR TCO GTR CCT 0C0 GAP OCR RAR TOT AGO CTO OTA GAR RCA CTA CCA GAR ART ATO GAT TTT AGA TCC GAT CRT TTA [let Trp Pro Phe Ala Ser
UVl
Pro Ala Glg Rio Lys Cys Arg Leu Val Olu Thr Leu Pro Olu Asn[let
Asp Phe Rrg Ser Rsp His Leu180 189 199 209 219 229 239 249 259 269
RCA RCA TTT GAR TOT TTT ARC GAR ATT RTC ACT CTR OCT ARO PAR TAT ATA TRC ATA OCR TCT TTT TOT TOT RAT CCT CTO ROT ACO ACT Thr Thr Phe Glu Cys Phe Asn Olu lie lie Thr Leu Rio Lys Lys Tyr lie Tyr lie Ala Ser Phe Cys Cys Rsn Pro Leu Ser Thr Thr
270 279 289 299 309 319 329 339 349 359
AGO GOR OCO CTT RTT TTT OAT RAR CTA RAR GAO OCR TCT GAR RAR 000 ATT RAR RTR ATA OTT TTO CTR OAT GAR COR 000 RAR ABA RAT
Rrg Oly Rla Leu lie Phe Asp Lys Leu Lys Olu Rla Ser Olu Lys Gig lie Lys lie lie Val Leu Leu Asp Olu Arg Gly Lys Rrg Rsn
360 369 379 389 399 409 419 429 439 449
CTO GOR GAO CTA CAR ROT CRC TOC CCO ORT RTR RAT TTT ATA RCC OTT ART ATA OAT RR RAR RAT RAT OTO GOR CTA CTR CTC GOT TOT Leu GIy Olu Leu Gin Ser HIs Cys Pro Asp lIe Rsn Phe lie Thr Val Asn lIe Asp Lys Lys Rsn Asn Val Olg Leu Leu Leu Glg Cys
450 459 469 479 489 499 509 519 529 539
TTT TOO OTO TCA OAT OAT OAR ROR TOT TAT OTA OGR ARC OCO TCR TTT ACT 0OG GOA TCT PTR CRT ACO RTT AAR ACO TTR GOT OTR TAT Phe Trp Val Ser Asp Asp Olu Arg Cys Tyr Val Gly Asn Ala Ser Phe Thr Gly Gly Ser Ile Hls Thr Ile Lys Thr Leu Gly Val Tgr
540 549 559 569 579 589 599 609 619 629
TCT OAT TAT CCC CCO CTO OCC RCA OAT CTT COT ROR ROR TTT OAT ACT TTT ARA 0CC TTT RAT ROC OCR ARA RPT TCR TOO TTG RAT TTA
Ser Asp Tyr Pro Pro Leu Rla Thr Asp Leu Arg Arg Arg Phe Asp Thr Phe Lys Rla Phe Rsn Ser Ala Lys Rsn Ser Trp LeuAsn Leu
630 639 649 659 669 679 689 699 709 719
TOC TCT OCO OCT TOT TOT TTO CCA OTT ROC RCT OCO TAT CRT ATT PRAG RT CCT ATA GOT GOR OTO TTC TTT ACT OAT TCT CCG OAR CRC
Cys Ser Ala Ala Cys Cys Leu Pro Ual Ser Thr Rio Tyr His Ile Lys Rsn Pro Ile Oly Oly Val Phe Phe Thr Asp Ser Pro Glu Hils
720 729 739 749 759 769 779 789 799 809
CTA TTO GOR TAT TCT ROR ORT CTA OAT ACC OAT OTA OTT RTT OAT RAR CTC RAG TCO OCT RAO ACT ROT RTA OAT ATT ORA CRT TTO 0CC
Leu Leu Glg Tyr Ser Arg Rsp Leu Asp Thr Asp Vol Val Ile Asp Lgs Leu Lgs Ser Rio Lys Thr Ser Ile Asp lIe Glu His Leu Rio
810 819 829 839 849 859 869 879 889 899
ATA OTT CCC ACT RCA COT OTC GRC GOT RAT ROC TAC TRT TOO CCC GRC ATT TRC ARC TCC ATT ATA ORR OCR OCC RTT RAT ROR GOR OTT
lIe Val Pro Thr Thr Rrg Val Asp Glg Rsn Ser Tyr Tyr Trp Pro Asp lIe Tyr Asn Ser lie lie Olu Rla Rla Ile Rsn Arg Glg Val
900 909 919 929 939 949 959 969 979 989
RAG RTC ROR CTT CTA OTT GOT RAT TOO ORT RAG ARC GRC OTA TAT TCT ATO OCR ACC OCC RAR ROT CTR GRC OCO TTO TOT OTT CAR RART Lys lIe Rrg Leu Leu Val Oly Rsn Trp Asp Lys Rsn Asp Val Tyr Ser let Rla Thr Rla Arg Ser Leu Asp Rla Leu Cys Vol GIn Rsn
990 999 1009 1019 1029 1039 1049 1059 1069 1079
GAT CTA TCT OTO RAG OTT TTC ACT ATT CR0 RAT RAT RCA ARA TTO TTG ATA OTC GRC GRC ORA TAT OTT CRT RTC ACT TCO OCR RAT TTC Asp Leu Ser Vol Lgs Vol Phe Thr Ile Gin Rsn Rsn Thr Lys Leu Leu lie Vol Asp Asp Olu Tyr UVl His lie Thr Ser Rio Rsn Phe
1080 1089 1099 1109 ll9 1129 1139 1149 1159 1169
ORC GOR RCC CRT TRC CAR RAT CRC GOR TTC OTC ROT TTT ART ROT RTA OAT RAR CR0 CTT OTA ROC ORG OCT RAR ARA ATA TTT ORG ROR Asp GIg Thr His Tyr OIn Asn His Gly Phe Val Ser Phe Rsn Ser lIe Rsp Lys Gin Leu Val Ser Olu Rio Lys Lys lie Phe Olu Rrg
1170 1179 1189 1i99 1208 1220 1230
OAT TOO GTA TCT ROC CRC ROT PAR TCO TTR PAR ATT TAR RRRRRGARRRTAORGACOTRTRGR Asp Trp Val Ser Ser His Ser Lys Ser Leu Lys Ile ***
FIG. 6. Nucleotide sequence of the p37K gene and flanking regions. The sequence starts at the extreme right-handSalIsite of the HindIII Ffragment. Thederived amino acid sequence of the p37K polypeptide is also shown. The mappositionsofthe 5' ends of the mRNA are indicated by the bar.
directly (Fig. 4C, lanes p) or after hybridization to late RNA 230 bp as well as some adjacent left-hand DNA were
(Fig. 4C, lanes 1). The uncleaved fragment (probe 3) was also sequencedbythe strategyoutlinedinFig. 5.The nucleotide includedin the analysis. With probe 3, a majorband ofca. sequence and the derivedamino acidsequenceofthep37K
750 nucleotides was again observed. An intense band gene as wellasthe 5'-and 3'-flanking regionsare shownin
comigratingwiththelargerfragment ofprobe4 wasobtained Fig. 6.
after hybridizationtolate RNA, indicatingthattranscription Location ofthe5'endofthe mRNA. To mapthe 5' endof
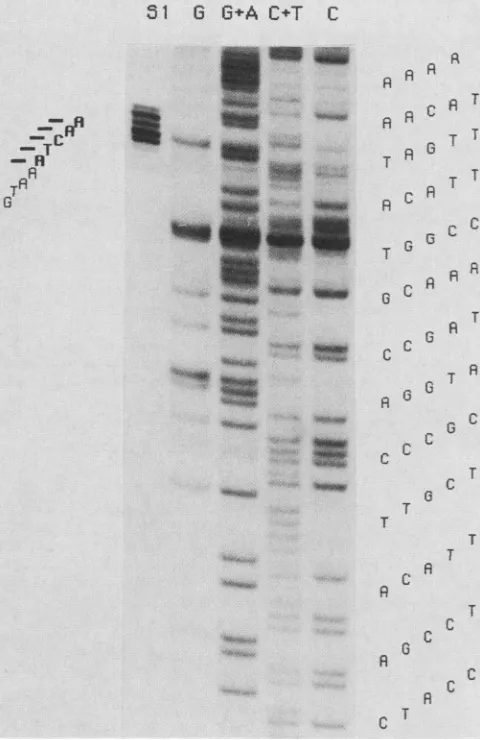
oftheputative p37K mRNAis fromrighttoleft. This result thep37KmRNAprecisely,welabeledanAluI-SalIfragment wasconfirmed with probe 5, withwhich the entirefragment (Fig. 5) spanning theputative initiation site oftranscription from the labeled left-hand end to the second BglII site but at the Alul site and used it for nuclease Si analysis. After
nottheright-hand BgIII-SalI fragmentwas protected. hybridization to late RNA, Si-resistant material was
ana-Sequence of the p37K gene. TheSailfragments of 840 and lyzed
alongside
a "sequence ladder"prepared
from theJ.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.68.558.68.577.2]VACCINIA VIRUS ENVELOPE ANTIGEN 763
same DNA fragment (Fig. 7). Five major Si-protected
fragments were observed. Since the entire region of the
DNA had already been sequenced, the positions of the protectedfragmentscould be located despitesome problems
in the sequencing reactions shown in Fig. 7. The results
indicated that the 5' ends ofthe mRNA map within about
two to six nucleotides upstream of the ATG translation
initiation codon.
Amino acid sequence of p37K. Following the first ATG
downstreamof the 5' end of theRNA, an openreading frame
which ended with a TAA termination codon at nucleotide
1208 wasobserved.The molecular weight of thepolypeptide
calculated fromtheamino acidcomposition was41,748 and
wasthussomewhat larger than thesizeof p37Kestimated by
polyacrylamide gel electrophoresis.
Hydrophilicity-hydrophobicity analysis of p37K. Since
p37K is tightly associated with Golgi membranes, we
searched for a putative signal peptide and possible
mem-brane anchors by determining the hydropathic index (10)
Sl G
6G+A
C+T C40
> 20 1
10
0--20
.Z
1 51 101 151 201 251 301 351
FIG. 8. Average hydropathicity of amino acids of the p37K polypeptide.ThemethodofHopp andWoods(10) withawindowof seven amino acids was used to generate the graph. The x axis indicates the amino acid number, and the y axis indicates the hydropathic index.
alongthepolypeptide chain (Fig. 8).No stronghydrophobic
AR
region
characteristic ofasignal peptide
wasfound atthe N A terminus ofp37K.
Twohydrophobic regions (amino
acids C A T 130 to 157 and 175 to 192) close to the center of the A A T polypeptide could represent putative membrane anchors. -RlT G
qg.p
FIG. 7. Mappingof the 5' end of the mRNAencoi
Alul-Sallfragment(seealsoFig. 5)waslabeledatth,
hybridized tolate RNA from infected cells. Nuclea:
material(lane Si)and theproductsof the sequenceri
G,G+A,C+T,andC)of thesamefragmentwere res
sequencing gel. The map positions of the 5' endE
(indicated by the horizontal bars) are shown at t
sequenceof thenoncodingstrandin the 5' to3' dire
tobottom.
T A u DISCUSSION
R T Incontrast toother viruses whichacquire theirenvelope
R C A
by
abudding
processthrough
the cellmembrane,
vacciniacC virus
particles
areenwrapped
in the interior ofthe cellby
T
Golgi-derived
membranes.T In this
study
wemapped
andsequenced
thegeneencoding
C A themajorviralantigenpresent on theenvelopeof
extracel-G lularvacciniavirusparticles. This antigenrepresents 5 to7% T of the totalproteinmassof thevirus(8).Hybrid selection of
c RNA,in vitrotranslation, andimmunoprecipitation allowed
C A us to mapthe gene close to the right-hand end ofthe HindlIl A T F fragment. This location corresponds to about 38kilobases A G G from the left-hand end of the 190-kilobase vaccinia virus
C genome. More precise map information was obtained by
C nuclease
S1
analysis,
which also showedthatthe mRNAisC transcribed fromrighttoleftonthe vaccinia virusDNA.
c T The nucleotide sequence of the
p37K
genereveals several T Ginteresting
features. First, the 5' ends of the mRNA as T definedby S1 analysismap withinno more than six nucleo-T tides from the translation initiation codon. Such extremely T short untranslated leaders have been observed in all three C ^ late genes thathave so farbeen sequenced (3, 20, 26) and thus appear to be ageneral
feature of vaccinia virus late C genes.A furtherhighly
conserved feature of lategenesis the G C TAAsequence whichimmediatelypreceeds the ATGtrans-R lation initiation codon. The sequence TAAATG has been
C observedin mostlategenes sequencedelsewhere(3, 19, 26)
A and in several additional late genes sequenced in our
labo-c T ratory
(E.
VanMeir,
unpublished
data).
Sofar,
theonly
exception is alategene analyzed byRosel andMoss(20)in
ding p37K.An which thesequence preceding the ATG translation initiation eAluIsite and codon isTAAATA. Further conserved elements of putative
se Si-resistant late promoters alsoinclude stretches of fivetoeight
thymi-oeadcins
(lane8% dine (3; p37K gene, this paper) or adenine (20, 26) residues sof the RNA starting at 16 to 20 bp upstream of the ATG codon. he left on the The open reading frame reveals a polypeptide composedctionfromtop of 372 amino acids and having a total molecular weight of
41,748. The difference between the calculated molecular
VOL.58, 1986
TT
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.576.303.539.63.228.2] [image:7.576.41.281.266.636.2]764 HIRT ET AL.
weight and thatestimated fromtheelectrophoretic mobility (8)could beexplained bya signalpeptidewhich isremoved
posttranslationally to produce the mature form of p37K.
However, examination ofthe hydropathic index shown in
Fig. 7 does not supportthis idea. Furthermore,the in vitro
translation product migrates slightly ahead of the in vivo
product(Fig. 1),aresultwhich would alsoargueagainstthe
removal of a signal peptide in vivo. It should be borne in
mind, however, that p37K is modifiedin vivobytheaddition
ofpalmitic acid, which presumably also influences its
elec-trophoretic mobility. The fact thatp37Kacquiresitsaffinity
for membranesby thismodification,similartowhathasbeen
proposed, for instance, for the transforming proteins of
retroviruses(22), also has to be considered.
Examination of the hydropathicity graph reveals that
p37K is relatively hydrophobic, consistent with previous
phase partion experiments and the recognized membrane
association (9). Two hydrophobic regions, each comprising
27 amino acids, are located between amino acids 130to 157
and 175 to 192. These regions could possibly serve as
membrane anchors.
An interesting question is whether p37K is essential for
viral replication or not. The fact that nonenveloped virus
particles harvested from experimentally lysed cells are
in-fectioussuggeststhatitmightnotbe. Furthermore, left-hand
DNA sequences are frequently deleted in orthopoxvirus genomes (reviewed in reference 27), and in one rabbit
poxvirus mutant described by Moyer and Rothe (15) the
deletion extends into the HindlIl F fragment. Although
viable inmostcelllinestested,suchdeletionmutantsmay be
hostrangerestricted(6). Clearly, large portionsof left-hand
DNA sequences are not required for viralreplicationunder
mostexperimental conditionsbut mayencode functions that
modify the biological behavior of the virus. Interestingly,
although both intracellular and extracellular viruses are
infectious, they differmarkedlyin their mode ofadsorption
and penetration (18) in cell cultures. This observation sug-gests that at least the functions which influence certain
biological propertiesarecarried ontheenvelopeof the virus
particle.Nowthat thep37Kgene hasbeenmapped,itshould
be possible to genetically engineervaccinia virusby
insert-ing foreign DNA into thep37K locustoinactivate the gene.
Such experiments have become feasible through the
possi-bility of constructing vacciniavirusrecombinantsby
homol-ogous in vivo recombination (reviewed in reference 23).
Provided that suchrecombinantsareviable, theywould offer
anidealtool forstudyingvirusmultiplicationinthe absence
ofp37Kexpression.
ACKNOWLEDGMENTS
We thank Anne Seiler-Tuyns and Walter Wahli for critically
reading the manuscript, Beatrice ten Heggeler for artwork, and HannelorePagelfor editorialhelp.
This workwassupported bytheSwissNational Science Founda-tion.
LITERATURE CITED
1. Appleyard,G.,A.Hapel,and E.A.Boulter. 1971. Anantigenic
difference between intracellular and extracellular rabbit-pox
virus.J. Gen. Virol. 13:9-17.
2. Berk,A.J.,and P. A.Sharp.1977.Sizingandmappingofearly
adenovirusmRNA'sbygelelectrophoresisofS1
endonuclease-digested hybrids. Cell12:721-732.
3. Betholet, C.,R.Drillien,and R.Wittek.1985. Onehundred base
pairs of5' flanking sequence ofavacciniavirus late geneare sufficient totemporally regulate latetranscription. Proc. Natl. Acad. Sci. USA82:2096-2100.
4. Cairns, J. 1960. The initiation of vaccinia infection. Virology
11:603-623.
5. Dales,S.,and B.G.T.Pogo.1981.Biologyofpoxviruses.Virol. Monogr. 18:1:109.
6. Drillien, R.,F.Koehren,and A.Kirn. 1981.Hostrangedeletion
mutant of vaccinia virus defective in human cells. Virology
111:488-499.
7. Galfre, G., andC. Milstein. 1981. Preparation ofmonoclonal antibodies:strategiesandprocedures.MethodsEnzymol.73B: 1-46.
8. Hiller, G., H. Eibl, and K. Weber. 1981. Characterization of intracellular and extracellular vaccinia virus variants:
N1-isonicotinoyl-N2-3-methyl-4-chlorobenzoylhydrazine interferes
with cytoplasmic virus dissemination and release. J. Virol.
39:903-913.
9. Hiller, G.,and K. Weber. 1985.Golgi-derivedmembranes that containanacylatedviralpolypeptideareused for vaccinia virus
envelopment.J.Virol. 55:651-659.
10. Hopp, T. P., and K. R. Woods. 1981. Prediction ofantigenic determinants. Proc.Natl. Acad. Sci. USA78:3824-3828. 11. Kilmartin, J. V.,B.Wright, and C. Milstein. 1982. Rat
mono-clonal antitubulin antibodies derivedby usinga new
nonsecret-ingcell line.J. Cell Biol. 93:576-582.
12. Maxam, A. M., and W. Gilbert. 1977. A new method for
sequencingDNA. Proc. Natl. Acad. Sci. USA 74:560-564.
13. Messing, J.,andJ.Viera. 1982. AnewpairofM13 vectorsfor
selecting either strand of double digest restriction fragments.
Gene 19:269-276.
14. Moss, B. 1985. Replicationofpoxviruses,p. 685-703. InB. N.
Fields,R. M.Chanock,andB.Roizman(ed.),Virology.Raven
Press,Publishers, NewYork.
15. Moyer,R. W.,andC. T. Rothe. 1980.The whitepockmutants of rabbitpoxvirus. I. Spontaneoushost range mutantscontain deletions. Virology102:119-132.
16. Payne, L. G. 1978. Polypeptide composition ofextracellular
envelopedvaccinia virus. J. Virol. 27:28-37.
17. Payne,L. G. 1979. Identification of the vacciniahemagglutinin
polypeptidefromacellsystemyielding largeamountsof
extra-cellularenvelopedvirus. J. Virol. 31:147-155.
18. Payne,L.G.,and E.Norrby.1978.Adsorptionandpenetration
of enveloped and naked vaccinia virus particles. J. Virol.
27:19-27.
19. Plucienniczak, A., E. Schroeder, G. Zettelmeissl, and R. E. Streek. 1985. Nucleotide sequence ofaclusterofearlyand late genes in a conserved segment of the vaccinia virus genome. Nucleic Acids Res. 13:985-998.
20. Rosel, J.,and B. Moss. 1985.Transcriptional and translational
mappingand nucleotide sequence analysis ofavaccinia virus
geneencodingtheprecursorof themajorcorepolypeptide4b. J.
Virol.56:830-838.
21. Sanger, F.,S.Nicklen,and A. R.Coulson.1977.DNA
sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.
USA 74:5463-5467.
22. Sefton, B. M., I. S. Trowbridge, J. A. Cooper, and E. M. Scolnick. 1982. The transforming proteins of Rous sarcoma
virus, Harvey sarcoma virus and Abelson viruscontaintightly
boundlipid.Cell 31:465-474.
23. Smith, G. L., and B. Moss. 1984. Uses ofvaccinia virus as a
vector for the production of live recombinant vaccines. Bio
Essays 1:120-124.
24. Turner, G. S., and E. J. Squires. 1971. Inactivated smallpox
vaccine: immunogenicityof inactivatedintracellularand extra-cellular vaccinia virus.J. Gen. Virol. 13:19-25.
25. Weaver,R. F.,andC. Weissman. 1979. MappingofRNAbya
modification of the Berk-Sharp procedure: the 5' termini of
P-globinmRNAhaveidenticalmap coordinates. Nucleic Acids Res. 7:1175-1193.
26. Weir,J. P., and B. Moss. 1984. Regulationofexpression and nucleotide sequence ofa late vaccinia virus gene. J. Virol. 51:662-669.
27. Wittek, R. 1982. Organization andexpressionofthepoxvirus genome. Experientia32:285-297.
28. Wittek,R.,M. Hanggi,andG. Hiller.1984. Mappingofagene
coding fora majorlate structural polypeptideonthe vaccinia
virus genome.J. Virol.49:371-378.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG.1.RNAvitro.immunoprecipitationwithcelltheimmunoprecipitationfluorograph Immunoprecipitation of p37K synthesized in vivo and in Vaccinia virus-infected cell monolayers were labeled for 3 h [35S]methionine and [35S]cysteine at 4 h after infection](https://thumb-us.123doks.com/thumbv2/123dok_us/1380006.91210/1.612.370.514.347.636/rnavitro-immunoprecipitationwithcelltheimmunoprecipitationfluorograph-immunoprecipitation-synthesized-vaccinia-monolayers-methionine-infection.webp)