0022-538X/88/093201-09$02.00/0
Copyright© 1988, American Societyfor Microbiology
Insufficiency
of
Transformation by Simian Virus
40,
Polyomavirus,
EJ-ras,
or v-myc
Oncogenes for Conversion of
Ethanolamine-Responsive Mammary Cells
to
Ethanolamine-Nonresponsive Cells
TAMIKO KANO-SUEOKA* ANDDAVIDM. KING
Department ofMolecular, Cellular, and Developmental Biology, University of Colorado, Boulder, Colorado80309-0347
Received23 February 1988/Accepted 16 May 1988
Normalmammaryepithelial cells (ethanolamineresponsive)requireethanolaminetoenable themtogrowin
defined culture medium because theycannotsynthesize de novo asufficient amountof phosphatidylethanol-amine. Mammarytumorcells whichretainproperties of thenormal tissuearealsolikelytobeethanolamine
responsive, whereasdedifferentiated, highly tumorigenic mammarytumorcellsareethanolamine nonrespon-sive. The nonresponsivetumorcellsareabletosynthesizethenecessaryamountofphosphatidylethanolamine to sustain growth. Therefore, the progression of mnalignancy seems to convert ethanolamine-responsive
mammary cells to ethanolamine-nonresponsive ones. In anattempt to provethe above assumption and to
understand themechanismresponsible forthe conversionduring the progressionofmalignanttransformation,
mammarytumorcellline64-24,which istypically ethanolamine responsive,wastransfected withsimian virus
40, polyomavirus, EJ-ras, or v-myconcogenes,and theresultingtransfectantswereexamined for their growth
responsetoethanolamine. Many of the transfectants exhibited typical transformed phenotypes; however,none
of the transfectants converted to ethanolamine-nonresponsive cells. Some of the SV40 and polyomavirus transformantswereable togrowinthe absence ofethanolamine, although theygrewbetter in thepresenceof ethanolamine, unlike typical ethanolamine-nonresponsive cells. These cells could grow in the absence of ethanolamine,eventhough theirmembranephospholipidwasphosphatidylethanolaminedeficient. The present
study indicates that theexpressionofany oneof the fouroncogenes tested, whichallows the cellstoexhibit transformed phenotypesin 64-24cells, isnotsufficientfor the conversion ofethanolamine-responsive cellsto
-nonresponsivecells.
Mammalian cells can be classified as ethanolamine (Etn) responsive ornonresponsive with regardto their prolifera-tion in definedculture medium (fora review, see reference
19). Without supplemental Etn, Etn-responsivecellscannot
synthesize asufficientamount ofphosphatidylethanolamine (PE); thus, thecomposition of their membrane phospholipid becomes PE deficient and develops a phosphatidylcholine
(PC)excess.This is followedbyaslowingdownorcessation
ofgrowth (17, 20).Thecessation ofgrowthprobablyoccurs because membrane-associated functions do not proceed properlyin cellshavinganabnormal phospholipid
composi-tion (21). TheEtn-responsivetypeincludes epithelial cells of ectodermalandendodermal origins.Known cells of thistype arenormal rat, human, andmousemammaryepithelialcells (8, 13, 36); rat mammary carcinoma cell lines 64-24 and MTW9/PL(4, 12, 14);thehuman breastcarcinoma cell line T47D (14); humanepidermal keratinocytes and lung, bron-chial, and prostate epithelial cells (26, 28, 33, 41); rat
esophageal epithelialcells(1);ratpituitarytumorcells(36a); andmouseplasmacytoma, SP2/O,and all thehybridomasso farexamined(15, 30). TheEtn-nonresponsivetypeincludes several mammary carcinoma cell lines ofrats and humans (22-1, WRK-1, HBL100, MDA-MB231, and HS578T); rat
hepatoma;rat,hamster,or mousefibroblasts; andseveralrat
neuronal andglial tumorcell lines(11, 15).
Themostwidelystudied cellswithrespect toEtn
require-mentare mammary cells. Accordingto the evidence so far
accumulated, normal mammary epithelial cells and their
tumorcell lineswhichtendtoretainproperties ofthe normal tissue (less progressed tumors)areEtnresponsive, whereas
*Corresponding author.
tumor cells which have lost many of the characteristics of normal mammary cells (progressed tumors) are Etn
nonre-sponsive(10). Accordingly,wehypothesized that(i) normal epithelial cells require Etn to grow in culture, and (ii) a
particular process(es) duringthe progressionofmalignancy renders the cells Etn nonresponsive, and therefore the loss of Etn responsiveness may be the one ofpleiotropic mani-festations ofneoplastictransformation. If the above
assump-tion is true, transformation of Etn-responsive mammary
cells by a certain oncogene or oncogenes is expected to convertthe cells from theEtn-responsive tothe Etn-nonre-sponsivetype.
Inaneffortto testtheaboveassumptionandtoidentifyan
oncogene(s) that confers Etn nonresponsiveness, we initi-ated transfection ofratmammarycarcinoma cell line 64-24, which is typically hormone and Etnresponsive (12, 18), by using fouroncogenes,simian virus 40(SV40), polyomavirus, EJ-ras, and v-myc oncogenes. The resulting transfectants were analyzed for their growth responseto Etn, aswell as for other characteristics. The results indicated two
signifi-cantfacts. (i)Some of theSV40 andpolyomavirus
transfec-tants became Etn nonresponsive, whereas allof the ras or myctransfectants remained Etnresponsive. The membrane phospholipid composition of the SV40 transfectants, which became Etnnonresponsive, was,however, still PE deficient when the cells were cultured without Etn, indicating that
SV40T antigen and probablypolyomavirus Tantigens are
capable of circumventing the PE deficiency of the cells, which is usually detrimental to the cells, by some other mechanism to support the growth. (ii) Many ras or myc
transfectants manifested properties of characteristically transformed cells. However,none of them has convertedto 3201
on November 10, 2019 by guest
http://jvi.asm.org/
theEtn-nonresponsivetype, indicatingthat the transformed phenotypeisnotsufficient fortheconversion.
MATERIALS AND METHODS
Cellsand cell culture. The present studieswerecarriedout with a clonal rat mammary carcinomacell line, 64-24 (18). 64-24 cells are oftypical secretory epithelial cell morphol-ogy, and their growth is responsive to Etn and prolactin.
They rarely
formtumors inanimals andhardly
grow insoft agar, and the level of plasminogen activator is that of nontransformed cells(T. Kano-Sueokaand P.Hsieh, unpub-lished results; see below). The cells were normally main-tained in high-glucose Dulbecco modified Eagle medium withpenicillin (50U/ml), streptomycin(50
pug/ml),
5%horse serum (Irvine Scientific, Santa Ana, Calif.), and2.5%fetal calfserum (Irvine Scientific) at37°C in ahumidified atmo-sphere of air containing 10% CO2 and were subcultured every 4days. Anchorage-independent growthwastestedby plating the cells in 0.35% agar containing the standard culture medium over the layer of 0.7% agarcontaining
the medium.After2weeks ofgrowththecolonieswere stained with neutral red and counted.Measurement ofgrowth stimulation. Cells (5 x
104)
were plated in duplicate or triplicateand grown in plastic tissue culture dishes (60 by 15 mm) either in serum-free medium describedpreviously
(16),exceptthat25 ,gof bovineserum albumin per ml was used, or in Dulbecco modified Eagle mediumcontaining 1.5% calfserum (Colorado Serum Co., Denver,Colo.)in thepresence or absenceof5,uM Etn. The cellswerereleased from plates withtrypsin andcounted ina Coulter counter(CoulterElectronics, Inc.,Hialeah,
Fla.).Plasmids. Fiveplasmidswere usedfor transfection. Four of them carried a
neomycin
resistance marker(Neo').
pSV3neo containing
Neor and an entireearly region
ofthe SV40 genome (38) andpSV5neo containing
Neor and an entire earlyregion
ofthepolyomavirus
genome (38) werepurchased
from the AmericanType
Culture Collection,Rockville,
Md. pSV6.6neo containingEJ-ras,
which is an allele of thehuman H-rasl gene, was constructedbyinsert-ing
a6.6-kilobaseBamHI fragmentfrom pEJ6.6(agift from R. A. Weinberg, Massachusetts Institute of Technology,Cambridge
[32]) into theBamHI site ofpSV2neo (38) and confirmedby
restrictionmapping by
C.Sackerson,
Univer-sity
ofColorado,
Boulder. The 6.6-kbfragment containing
the EJ-ras gene includes its endogenous promoter.
pSV7.3neo,
whichcontainsv-myc derivedfrom MC29avianmyelocystomatosis
virus, was similarly constructed frompSVv-myc
(25, 42) (obtained from the American Type Cul-tureCollection)
and pSV2neo by C. Sackerson. The 7.3-kilobase insert frompSVv-myc has theMC29long terminal repeatinthe upstreamregion
whichcontrols theexpression
of v-myc gene.
ptsA58
contains atemperature-sensitive
SV40
T-antigen
gene, tsA58(39),andwasobtained from L.Sompayrac,
UniversityofColorado,Boulder.Transfection and selection oftransfectants. Plasmid DNA wasprepared bythe standard alkali-sodiumdodecyl sulfate method (27). Essentially, the method described byGraham et al. (7),
involving
the calcium phosphate coprecipitationtechnique,
wasused totransfect 64-24 cells. Tologarithmi-cally
growingcells,25 ,ug ofplasmidDNA suspendedin 125 mMCaCl2,
140 mMNaCl, 25 mM N-2-hydroxyethylpipera-zine-N'-2-ethanesulfonic acid(HEPES),
and 0.75 mMNa2HPO4
(pH 7.2) was added, and the culture plates were incubatedfor24h.The cells were thenwashed, trypsinized, andplated
withaseriesof dilutions.Oneday later,when thecells wereattached and started toproliferateagain,500,ugof G418(GIBCOLaboratories,GrandIsland, N.Y.)permlwas added. Fivedays after the addition of
G418,
all untransfec-ted cells were dead and the G418-resistant colonies were clearly observable. Individual G418-resistant clones were then isolated and propagated in the presence of 500jig
of G418perml. Once the clonal cell lineswereestablished,the cells were maintained in the presence of250jig
of G418 per ml. The frequency of transfection with regard to G418 resistanceranged between 3 x 10-6 and 7.5 x i0-4. G418-resistant transformants with pSV2neo, which carries no oncogene, were also prepared as a control. SinceptsA58
doesnotcarrytheNeormarker,the cellswerecotransfected with pSV2neo DNA as well asptsA58.
Phospholipid analysis. The lipids were extracted fromthe cells in late log phase, andphospholipids were separatedby two-dimensionalthin-layerchromatographyandquantitated asdescribed before (20).
Expression of SV40 and polyomavirus T antigens and ras-p21. Expression ofT antigens in the cells transfected with plasmid DNA containing SV40 or polyomavirus T antigens was examined by fluorescence microscopy. The primaryantibodies used were PAB 419 monoclonal antibody against SV40 T antigen (a giftfrom E. Harlow, ColdSpring Harbor Laboratory, Cold Spring Harbor, N.Y.) and poly-clonal polyomavirus anti-T antibody (a gift from T. Benja-min, Harvard University, Boston, Mass.), and the second-ary antibodies were fluorescein-conjugated, affinity-purified goatanti-mouse immunoglobulin G for SV40 T antigen and goatanti-rat immunoglobulinG for polyomavirus Tantigen (Cooper Biomedical, West Chester, Pa.). The cells were grown onglass coverslips and fixed with acetone at -70°C for 10min. The fixed cells were incubated with the primary antibodies for 30 min at 37°C in a humidified atmosphere, and after the cells had been washed twice, 100-fold-diluted secondaryantibodieswere usedtostainthe cells at37°Cfor 30min.
Monoclonal antibody against ras-p21 was obtained from Eastman Kodak Co., Rochester, N.Y. The cells grown on glass cover slips were fixed with acetone-methanol (1:1) at -20°C. The fixed cells were reacted with 50-fold-diluted primary antibody at room temperature for 30 min and then with 80-fold-diluted fluorescein-conjugated goat anti-mouse immunoglobulin Gfor 30 min.
Assayofplasminogen activator. Theactivities of plasmino-genactivator in the crude cell lysates wereassayed byusing a chromogenic substrate by the method developed by D. Abbott-Brown, Synergen, Boulder, Colo. (unpublished results). The cells were lysed with 0.5% Triton X-100 in sodium phosphate (pH 8.1). The reaction mixture (150 ,ul) contained 4 ,g of human Glu-type plasminogen (American Diagnostica, Greenwich, Conn.), 40 ,umol of Spectrozyme PL(AmericanDiagnostica), and various amountsofthe cell lysate in0.05%Triton X-100 and 60 mMTrisbuffer(pH8.5). Microdilution plates were used to incubate the reaction mixtures at37°C for 3 to 6 h, and the amount of the product was measuredby its opticaldensityat 405 nmwith a Titertek Multiscan Spectrophotometer. Urokinase (Calbiochem-Behring, San Diego, Calif.) was used as the enzyme stan-dard. Theactivities were expressed as units permilligramof protein. Each assay included thetumorigenic ratmammary carcinomacellline22-1, and the activities for othersamples were normalized by taking the activity of 22-1 cells as 100.00.
on November 10, 2019 by guest
http://jvi.asm.org/
-D:.
'..I
q r-.
,.st4
H fiJ(LIL
iLlA S S f;s*Str;#AM A
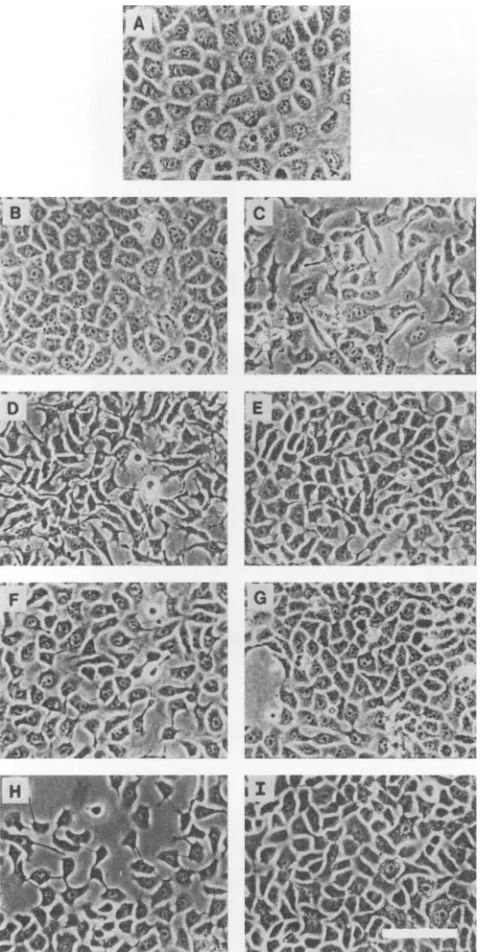
FIG. 1. Phase-contrast micrographs of 64-24 cells transfected withSV40, polyomavirus,v-myc, and H-rasoncogenes. (A) 64-24; (B) Sv-3; (C) Sv-7; (D) Py-5; (E) Py-21; (F) My-5; (G) My-11; (H) Ra-2; (I) Ra-10. Bar, 100 ,um. The cells were grown in regular maintenance mediumasdescribed in Materials andMethods.
RESULTS
Transfections with the SV40 oncogene. pSV3neo DNA,
which contains the SV40 oncogene, T-antigen gene, and
Neormarker,wastransfected into 64-24cellsasdescribed in Materials and Methods. The frequency of transfection against Neor was 4 x 10-4. Nine clonal lines of
G418-resistant cells were established and analyzed for various
properties. About half of these clonal lines looked very similartotheparental 64-24 cells, exhibiting a cobblestone-likesmooth sheetof cells(Fig. 1A and B). Four of the lines,
on theotherhand, had angular and irregular cell shapes and were less securely attached to the substratum (Fig. 1C). These cells tend to form irregular multilayer structures at confluency. Seven lines were tested for the presence of T antigen, and all of them were shown to be positive (repre-sentatives are shown inFig. 2).In mostcells theantigenwas expressed innuclei; however, in some cells only the cyto-plasm contained the stain. There were variations in the extentofstainingamong the clonal lines as well as within the culture of agivenline(Table 1; Fig. 2).Theirabilitytogrow insoft agar was examined to see whether theexpression of Tantigen allowsthecells to exhibittransformedphenotypes. These cloned lineshardlygrew in soft agar under standard conditions, except oneline, whichgrew at alowfrequency (Table 1). Difficulty in growing rat cells transformed with SV40 haspreviouslybeenreported (37).The level of plasmi-nogenactivatoractivity inthese transfectantswastherefore compared with that in untransfected 64-24 cells and its tumorigenicvariantcellline,22-1cells(18).The intracellular andextracellular levels ofplasminogen activatoractivityin 64-24and 22-1 cells have previouslybeen determinedbyD. Rifkin,NewYork University,NewYork, with
'25I-fibrin
as thesubstrate. 22-1cellshavean8- to10-fold-higher activity
than64-24cells do:theactivityof 22-1 cells is that oftypical malignant cells, and that of64-24 cells is typical of nontu-morigenic cells (31; D. Rifkin and T. Kano-Sueoka, unpub-lished results). In the present study, 64-24, 22-1, and four lines ofthe pSV3neo transfectants (all fourarepositive for SV40Tantigen)wereusedforthe assay. 22-1cellshad about 10-fold-higher activity than nontransfected and transfected 64-24 cells
did,
and none of the transfectants contained significantly higher activities than the parental64-24cellsdid (Table 2).The growth responseofthe nineclones toEtn was then examined to see whether the expression ofT antigen con-ferred Etnnonresponsiveness on thecells. The response of individual lines to Etnwasvariable
(Fig.
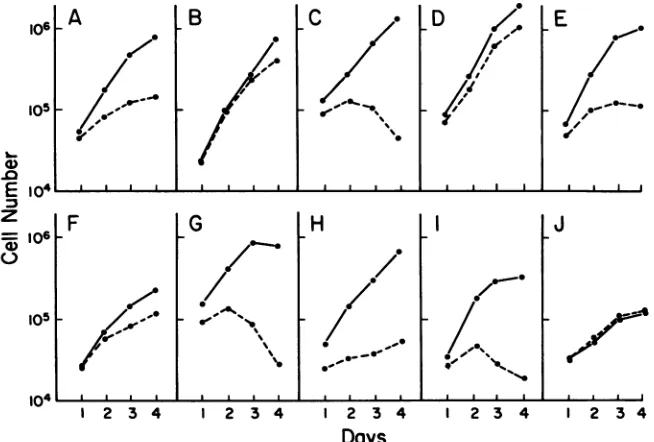
3AtoE;Table 1). The growth property of four lines (Sv-2, Sv-3, Sv-4, and Sv-9)wasvery similartothatof 64-24 cells(Fig. 3A andE). Three (Sv-1, Sv-5, and Sv-8) were more dependenton Etn than 64-24 cells were(Fig. 3C), and, finally,
twolines (Sv6-andSv-7)grew wellwithoutanEtnsupplement(Fig.3B and D). These two lines exhibit the morphology of transformed cells, and the levelsofTantigen
expressed,asexaminedby
immunostaining, seemedtobehigherthanin the other cells. However,thereareother
lines,
suchasSv-1 andSv-9,
which exhibit a transformed morphology or aline,
such as Sv-5, which expresses T antigen at a high level, although their growth behavior is similar to that of nontransfected 64-24 cells. Inaddition,
there is no distinction in the level ofactivity
ofplasminogen
activator whencomparing
Etn-responsiveandEtn-nonresponsive transfectants. Therefore, there seemtobe no notablecharacteristics thatdistinguish transfectants which became Etn nonresponsive from those which remained Etnresponsive.Phospholipid compositionsof theSV40 transfectantswere examined to findwhether theSv-6orSv-7lines, as aresult ofSV40transformation,wereable togrow in the absence of Etn because of their ability to synthesize sufficient PE whether the medium contains Etn or not.Accordingly, Sv-6, Sv7-,Sv-4, anduntransfected 64-24 cellsweregrown in the presence or absenceof5,uMEtninDulbecco modifiedEagle mediumcontaining 2% calf serum, and their
phospholipids
were analyzed as described in Materials and Methods. The results(Table 3)indicate that themannerinwhich the PEor PC content altered in all three transfected cell lines when
on November 10, 2019 by guest
http://jvi.asm.org/
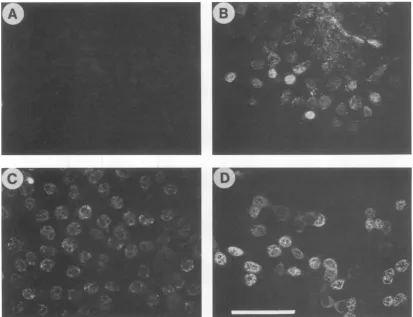
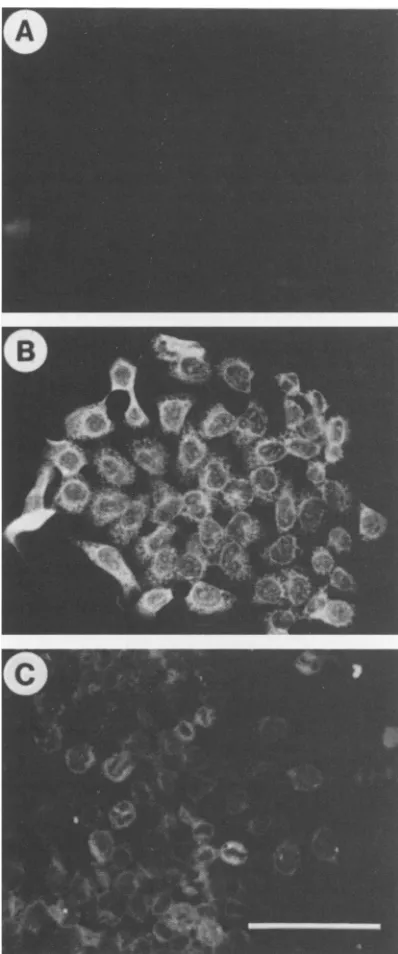
[image:3.612.56.296.69.545.2]FIG. 2. Fluorescencestainingof64-24cells transfectedwithSV40oncogenewithantibody against SV40 T antigen. (A) 64-24; (B)Sv-6; (C) Sv-1;(D) Sv-7. Bar, 100 ,m.
they were grown in the presence or absenceofEtnwasvery similar to thatofnontransfected 64-24cells. Namely, these transfected cellswere notable to maintain thenormal level ofPE orPC inthe absence ofsupplemental Etnas typical Etn-nonresponsive cells are able to do. The results thus suggestthatthe Sv-6and Sv-7 lines acquired theability to grow in the absence of Etn, although their phospholipid compositionisabnormal.
[image:4.612.103.516.72.389.2]To ensure that the Etn nonresponsiveness acquired by Sv-6orSv-7 cellswasduetothefunction of SV40Tantigen,
TABLE 1. Propertiesof64-24cells transfectedwithpSV3neoa
Cell Morhloy Etn Soft-agar T
anignd
line orphology responseb growth(%) Tanigen
Sv-1 Transformed + + <0.1 +
Sv-2 Parental + <0.1 NTe
Sv-3 Parental + <0.1 +
Sv-4 Parental + <0.1 +
Sv-5 Parental + + <0.1 + +
Sv-6 Transformed - 1.4 + +
Sv-7 Transformed - <0.1 + +
Sv-8 Parental + + <0.1 +
Sv-9 Transformed + <0.1 NT
aAveragefrequencyoftransfectionwith respect to
Neor
was3.5 x1O-4.
bSymbols:++, moreresponsivethan 64-24cells;+, asresponsiveas64-24
cells;-,nonresponsive.
c Thevalueswereobtainedby dividingthe number ofcolonies formedin soft agarbythe numberof colonies obtainedon aplastic surface.
dSymbols: +,clearly positive; ++,greaterthan twice the +value.
eNT,Nottested.
plasmidDNA containing atemperature-sensitive T-antigen gene,ptsA58,wastransfectedinto 64-24 cells withpSV2neo, and the resulting G418-resistant transfectants (not cloned, twoindependently isolated but mixedpopulations of trans-fectants) were examined fortheirgrowth
properties
at33°C
(permissivetemperature) and39°C (nonpermissive tempera-ture). Nontransfected64-24 cellswere clearlydependenton Etn for growth at both temperatures, and the dependence was more pronounced at 39°C (Table 4). Also, in the transfectants thecells behaved in a similar fashionas64-24 cells at39°C, whereasat 33°C both isolates ofthe transfec-tantsgrewrather wellwithoutEtn,althoughtheaddition of Etn shortened the generation time significantly (Table 4). Namely, at 33°C the generation times of transfected and nontransfected cells were very similar when they were
TABLE 2. Plasminogenactivator activities of
pSV3neotransfectants
Cellline %Activityto 22-1 cells"relative
22-1... 100.00
64-24... 9.31 + 1.76
Sv-1... ... 10.29± 2.23
Sv-5... ... 4.88 + 0.32
Sv-6... 5.94 ± 1.49
Sv-7... 10.69 + 1.70
a22-1cells hadanaverageactivityof 372
U/4ig
ofprotein.Activitiesareexpressedas meanststandard errorsof seven separate assays. Theactivities
werenormalizedbytakingtheactivityof 22-1 cells as 100.00.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.60.298.559.667.2] [image:4.612.319.557.608.697.2]..0
E
z
0 106
F
tO
G J
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
[image:5.612.148.475.71.292.2]Days
FIG. 3. GrowthresponsetoEtnof 64-24 cells transfectedwithSV40,polyomavirus,v-myc,and EJ-rasoncogenes. The cellswereplated
atadensity ofS x 104cellsperplate(60by10mm)inserum-free mediumorDulbecco modifiedEaglemedium containing 1.5% calfserum
with (-)orwithout(---)5,uMEtn.The cellswerereleased fromtheplateswith trypsin andcounted inaCoultercounter.Eachdatum
pointrepresentstheaverageofduplicateortriplicate samples. (A)64-24;(B) Sv-7; (C) Sv-1; (D) Sv-6; (E) Sv-2; (F) Py-6; (G) Py-2;(H) Ra-29;
(I) My-11; (J)22-1.
growninthepresenceof Etn,whereas in the absence of Etn
the growth of 64-24 cells literally stopped, but transfected cellskeptgrowing, withgeneration times less than twice the time ofthosegrownwith Etn. The above results strengthen
the idea that acquisition of Etn independence by these transformants is duetothe action of T antigen.
Transfection with the polyomavirusoncogene.Theeffect of
DNAvirusoncogeneon64-24cellswasfurtherexamined by
transfecting the cells with plasmid DNA containing poly-omavirusT-antigengene,pSV5neo. A total of 22 Neor clonal
lineswereisolated, and 6 of themwereexamined for various
characteristicsasdescribedabove.SimilarlytothepSV3neo transfectants, theirmorphologies ranged from those indistin-guishable from that of parental 64-24 cells to those of typically transformed cells with angular and spiky shapes and less adherence to the plate (Table 5; Fig. 1D and E).
However, polyomavirus transfectants exhibited typical transformed morphology more frequently, and their
trans-formedphenotype wasmorepronouncedthan thatofSV40 transfectants (Fig. 1D). Our results agree with previous findings with rat fibroblasts that polyomavirus gives fully transformedphenotypes, whereas SV40 givesan
intermedi-ateleveloftransformation (34, 37). All of themsofartested
werepositive for the expression of polyomavirus T antigens
(Fig. 4), and, unlike pSV3neo transfectants, all of the pSV5neo transfectantswereabletogrowinsoftagar(Table 5).Withregardtothegrowthresponseofthese transfectants to Etn, one (Py-6) of six lines tested grew without supple-mentalEtn, butnotaswellaswith Etnaddedasin thecase
of Sv-6 or Sv-7 lines (Fig. 3F). The majority ofpSV5neo
transfectants was more dependent on Etn than the 64-24
cellswere(Fig. 3G). Notable characteristics associatedwith
TABLE 3. Phospholipidcompositions ofpSV3neotransfectants
Cell Mean amt(%)+SEof:
line SMa
plb
p5c PC PE64-24 - 2.5 ± 1.1 9.0 + 2.5 10.7 ± 3.7 62.0± 8.2 15.8 ± 2.1
+ 2.7 ± 1.0 9.4 ± 1.2 13.0± 1.5 52.0± 2.3 22.7 ± 1.0
Sv-4 - 2.5 ± 1.0 7.7 ± 0.9 12.6± 1.4 63.9 ± 3.1 13.4 ± 0.8
+ 2.4 ± 0.8 9.1 ± 1.5 11.6± 1.2 52.4 ± 1.5 24.6 + 4.2
Sv-6 - 1.6 ± 0.2 10.0 ±0.5 10.5 ± 1.2 62.8 + 2.6 15.3 ± 0.9
+ 1.7 ± 0.3 9.7 ± 1.5 11.2± 0.5 56.3 + 1.2 21.1 ± 0.3
Sv-7 - 2.6 ± 1.1 9.1 ±0.7 11.1 ± 0.8 60.2 ± 2.4 17.0 ± 1.0
+ 3.1 ± 0.9 6.6 ± 0.6 9.6± 0.8 52.0+ 2.3 28.6 ± 1.0
aSM,Sphingomyelin. b
Pl,
Phosphatidylinositol.cPS, Phosphatidylserine.
I I a I
--so
r 0
0
4".., 0--el
I I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.60.562.564.699.2]TABLE 4. Growth responseofptsA58 transfectants to Etn at permissive and nonpermissive temperatures
ICiell Temp (°C) Generation time(hr)a
line
Tm(C)+
Etn -Etn64-24 33 12.0 ± 1.6 59.3 ± 13.2
39 13.3 ±0.9 NGb
ts-3A 33 13.3 ± 2.4 19.3 ± 1.3
39 11.7 ± 1.3 NG
ts-3C 33 10.8± 1.3 19.3 ± 1.5
39 11.5 ± 2.5 NG
aMeans ±standarderrorsofthree orfourseparateexperiments. bNG,Nogrowth.
their Etn-responsive growth was again notfound as in the case of SV40 transfectants. The phospholipid composition was not analyzedfor the Py-6line,which was able to grow without Etn.
Transfections with EJ-ras and v-myc oncogenes. A totalof 33 and 11 lines were isolated for pSV6.6neo (EJ-ras) and pSV7.3neo (v-myc) transfectants, respectively, and 6 repre-sentative lines ofeach werefurtherexamined.The morphol-ogy ofmost ofthe v-myc transfectants was verysimilar to thatofthe parental cells. Cellsof one line,My-5, exhibited some transformed phenotypes when they were semicon-fluent.However, theconfluentcultureofMy-Swasvirtually indistinguishable from that of 64-24 cells (Table 6; Fig. 1F andG). Thegrowthin softagarofthese cells wasgenerally negative except for one line, My-11, which grew at a low efficiency, and, finally, all lines needed Etn to grow in a similar fashion to 64-24 cells. Interestingly, My-11, which exhibitssignificantgrowth insoftagar, was moredependent onEtnthan theparental linewas (Fig. 31).
Thefrequency oftransfection for pSV7.3neo, whichwas selected for Neor, was unusually low (1.2 x
10-6
to 3 x10-6)
compared with other plasmids.The reason maybethat the majority oftransfected cells containmultiple copies of mycDNA, andelevatedlevelsofmycproductarecytotoxic (43); therefore, only a few which do not contain excess plasmid DNA have survived. This may be the reasonwhy literally all of the myc transfectants are similar to the parental cells.In contrast to v-myc transfectants, transformed cells of literally all EJ-ras transfectants had distinct characteristic morphologies (Table 7). The cells had angular edges and
TABLE 5. Properties of64-24cells transfected with
pSV5neoa
CellMorphology
Etn Soft-agar Tantigen'line
~~~~response
b growth(%)C64-24 Parental + 0.2
22-1 Transformed - 21.1
Py-2 Parental ++ 0.7 +
Py-3 Parental ++ 4.7 +
Py-4 Transformed ++ 2.8 +
Py-6 Transformed - 11.9 ++
Py-18 Transformed ++ 10.1 + +
Py-21 Transformed + 8.2 + +
a Averagefrequency of transfectionwas 7.5x10-4.
bSymbols:++,moreresponsivethan64-24cells; +,asresponsive as 64-24 cells; -, nonresponsive.
c See Table 1, footnotec.
d See Table 1, footnoted.
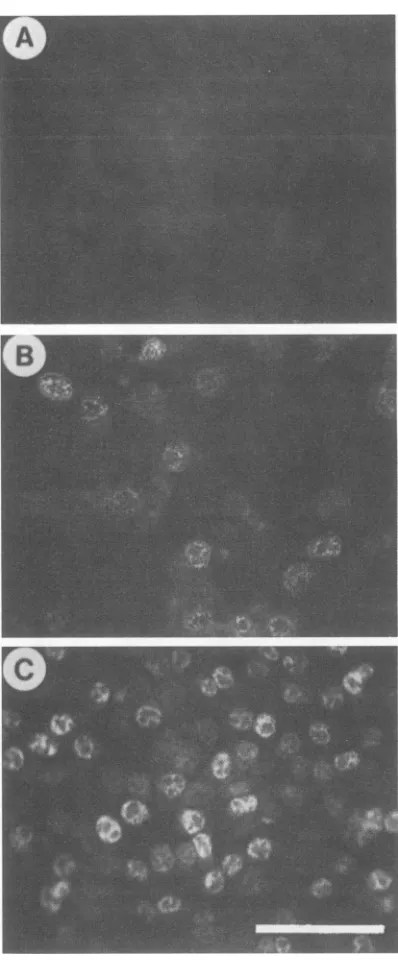
FIG. 4. Fluorescence staining of 64-24 cells transfected with polyomavirus oncogene. (A) 64-24; (B) Py-6; (C) Py-21. Bar, 100
,um.
werelessattached to the substratum(Fig. 1HandI),andtwo of six lines tested grewpositively in soft agar(Table 7). All lines tested have clearly overproduced
ras-p21 protein
as tested by fluorescence microscopy (representatives are shown inFig.5).
Further, theextent ofras-p21 production seemsto becorrelated with theefficiencyofsoft-agar colony formation (Fig. SB and C). However, none of thembecame Etnnonresponsive or more dependenton Etn as aresultof EJ-rastransfection.DISCUSSION
Ourprevious observations regardingthe Etn
responsive-nessof various mammary cells in culture ledusto
postulate
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.54.294.93.202.2] [image:6.612.57.295.583.684.2]TABLE 6. Properties of 64-24 cells transfected with pSV7.3neoa
Cell Etn Soft-agar
line Morphology responseb growth(%)
My-4 Parental + <0.1
My-5 Transformed + <0.1
My-8 Parental + <0.1
My-9 Parental + <0.1
My-10 Parental + <0.1
My-11 Parental + + 1.1
a Average frequency of transfection was 1.2 x 10-6 to 3 x 10-6.
bSymbols:++,moreresponsive than 64-24 cells; +,asresponsiveas64-24 cells.
that some step or stepsduringtheprogression of malignancy should alter the mammarycellsfrom the Etn-responsive to the Etn-nonresponsive type. To testthe above hypothesis, wehaveexamined theeffect ofoncogenes onEtn-responsive mammary cells. Ifthe results arepositive, Etn nonrespon-siveness could be used as a new parameter for tumor progression. Four oncogenes (twoDNA virusand mycand rasoncogenes) were chosenforthe presentstudy.
Transfection with these oncogenes brought about some noticeable changes characteristic oftransformed cells in the recipient mammary cells. In addition, 3 clonal isolates of SV40 and polyomavirusoncogenetransfectantsoutof15so
far testedwereable to growwithout supplementalEtn. The analysis ofthe growth response ofthe three transfectants seemingly converted to the Etn-nonresponsive type indi-cated that these cells were not completely independent of Etnasinthe caseoftypically Etn-nonresponsive 22-1 cells (Fig. 3J), suggesting that the nature ofEtn-nonresponsive growth ofthesetransfectants may be different from that of typically Etn-nonresponsivetumorcells. Indeed, the analy-sisofphospholipidsinthese cellsindicatedthattheirability togrowwithoutEtndid not occur becausetheywereableto synthesize sufficientPEde novo and hencemaintain normal phospholipid composition.Fibroblasts suchasmouse3T3or babyhamsterkidney cells transformedwithSV40 havebeen knownto produce aprotein growth factor(s) which stimu-latesthe growth ofthe untransformedfibroblast cells (2, 3, 35). Accordingly, the ability of Sv-6, Sv-7, orPy-6 cells to growin theabsenceofEtnin serum-free mediummay occur because these transformants produce an autocrine growth factorwhichcan stimulate thegrowth ofthe cells irrespec-tive oftheir abnormal phospholipid composition. Prelimi-nary results indicate that these cells indeed produce some kind of factororfactors whichcan stimulate thegrowth of parental64-24cellsin the absence ofEtn.The natureofthe factor(s) isnotknown.
The hypothesis that acquisition of Etn-nonresponsive growth aftertransfection with SV40 or polyomavirus
onco-TABLE 7. Properties of64-24cells transfected withpSV6.6neoa
Cell Etn Soft-agar
line Morphology response6 growth(%)
Ra-2 Transformed + <0.1
Ra-3 Transformed + <0.1
Ra-10 Transformed + 0.1
Ra-11 Parental + 0.1
Ra-15 Transformed + 5.0
Ra-29 Transformed + 0.6
a Average frequencyof transfection was 6.5x
1O-4.
b+,Asresponsiveas64-24 cells.
FIG. 5. Fluorescence staining of 64-24 cells transfected with H-ras oncogene. (A)64-24; (B) Ra-15;(C)Ra-29.Bar, 100 ,um.
genes is due to the expression of the T antigens was confirmed by using a mutant with a temperature-sensitive mutation of the SV40 T-antigen gene, tsA58. Growth pat-terns ofthe tsA58 transfectants behaved according to our predictions;
i.e.,
atthenonpermissive
temperaturethe trans-fected cellswere dependenton Etn forgrowth, whereas atthepermissivetemperature theycould grow in the absence of Etn. In this case, however,thecells were notcompletely independent or nearly independent of Etn as were Sv-6, Sv-7,and Py-6. This may be because mixedpopulations of transfectants selected for Neor were used for the growth analysis,andthereforetheculturemustconsist ofamixture ofEtn-responsiveandEtn-nonresponsivetypes if theptsA58 transfection gave riseto asimilarproportion of
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.54.296.84.167.2] [image:7.612.56.295.624.706.2]sive and Etn-nonresponsive types as the pSV3neo transfec-tion did. An interesting observation regarding the behavior of 64-24 cells at different temperatures is that the cells are more sensitive to the Etn requirement at higher tempera-tures.Although the cells remain alive at 37°C in the absence of Etn, even if their growth stops, they cannot survive at 39°C without Etn. It may be that at 39°C ahigh PC content makes the cellular membrane too fluid to function properly or tomaintain an intact cellular structure.
SV40 does not fully transform normal rodent cells (37). The present study indicates that the SV40 oncogene is not able toconfer transformed phenotypes such as the ability to grow in soft agar orhigher levels ofplasminogen activator activityon 64-24 cells. In contrast to SV40, transfection with thepolyomavirus T-antigen gene caused most of the cells to havetypicallytransformed characteristics, as in the case of rat fibroblasts (34, 37). The effect of those DNA viral oncogenes on Etn responsiveness seems, however, very similar.
For the present study we have chosen two oncogenes, myc and ras, from a number ofavailable oncogenes whose normal counterparts exist in the host cells, since these oncogenes are often activated in mammary tumor cells. The myc gene has often been found tobe amplified in primary humanbreast carcinomas (6) or human breastcarcinomacell lines(23, 29). The ras genes are also often overexpressed in breast carcinoma tissues (9, 40). Particularly interesting is that the c-Ha-ras oncogene confers on the estrogen-respon-sive human breast carcinoma cell line, MCF-7, estrogen autonomyforgrowth both in cultureandin animals (5, 22). Our results indicated that some myc-transfected cells and literally all of the ras-transfected cells manifested some characteristics of transformed cells. However, neither ras nor myc alone converted the Etn-responsive cells to Etn-nonresponsive cells. Introduction of the EJ-ras gene into 64-24cells altered not only themorphology ofthecells,but also their ability to grow in soft agar, suggestingthat these cells haveprogressed atleastastep further in the develop-ment of malignancy. Furthermore, the amount of ras-p21 present in ras-transfectedcells seems to be correlated with the ability of the cells to grow in soft agar.
However,
this was notsufficient to convert the cells to Etn nonresponsive-ness. myc transfection resulted in the least strong manifes-tation of transformed phenotypes inrecipient cells.The myc gene product was not examined in these transfectants. However, since all of the Neorisolates forthetransfectants from three other oncogene-containing plasmids expressed significantamountsof oncogene products, it can be reason-ablyassumed that the selected Neor cells forpSV7.3neoalso integrated myc DNA. However, since it is highly possible that the reason for the lowfrequency of transfection with pSV7.3neo is the cytotoxicity ofthe myc gene product at elevated levels (43),survivingNeorisolates may notproduce excess myc gene product. If theabove is the case, the fact that cells converted to Etn nonresponsiveness were not foundamong myctransfectantsmay not besignificantunless only a small amountof myc gene product is required for the conversion.The present study suggests that oncogenes of DNA vi-ruses are not capable of converting Etn-responsive mam-marycellstoEtn-nonresponsive cells and that conversion of thecells from Etnresponsiveness to nonresponsiveness may require the activation of a cellular oncogene other than myc or rasor the activation of more than one oncogene. Rear-rangement ofchromosomes or the loss of the allele of one of theoncogenes in question may be necessary in addition to
the expression of oncogenes for converting cells from a responsive to a nonresponsive state. Forexample, the loss of a normal allele of the ras oncogeneseemstobeimportant for theprogression ofmalignancy in breast cancercells(24, 40). The examination of oncogenes other than the ones used in the present study and double transfections withvarious combinations ofoncogenes are under way. The long-term culture of ras and myc transformants under conditions without Etn to select forEtn-nonresponsive cells is also in progress inorder to examine thepossibility that a secondary eventafter the introduction of myc or ras DNAis necessary forobtaining the Etn nonresponsiveness. This line of study should contribute to theunderstanding of the phenomenon ofmalignancy progression.
ACKNOWLEDGMENTS
We aregrateful toRichard Frederickfor able technical assistance. Thisworkwassupported by Public Health ServicegrantCA30545 from the National Institutes ofHealth andgrantPCM8313161from
the National ScienceFoundation.
LITERATURE CITED
1. Babcock, M. S., M. R. Marino, W. T. CunningIII, and G. D. Stoner. 1983. Clonalgrowth and serialpropagation ofrat
esoph-ageal epithelial cells. In Vitro(Rockville) 19:403-415.
2. Bourne, H. R., and E. Rozengurt. 1976. An 18,000 molecular
weight polypeptide inducesearly events and stimulates DNA
synthesisincultured cells.Proc. Natl. Acad. Sci. USA
73:4555-4559.
3. Burk, R. R. 1976. Induction of cell proliferation byamigration factor released fromatransformed cell line.Exp. CellRes. 101: 293-298.
4. Danielpour,D., T. L.Riss,M.Ogasawara, and D. A. Sirbasku. 1988. Growth ofMTW9/PL2estrogenresponsiveratmammary tumorcellsinhormonally defined serum-free medium. InVitro (Rockville)24:42-52.
5. Dickson, R. B., A.Kasid,K. K.Huff,S. E.Bates, C.Knabbe,D. Bronzert, E. P.Gelmann,and M. E.Lippman. 1987.Activation ofgrowth factor secretionintumorigenicstatesof breastcancer
induced by 17p-estradiol or v-Ha-ras oncogene. Proc. Natl. Acad. Sci. USA84:837-841.
6. Escot, C., C. Theillet, R. Lidereau, F. Spyratos, M.-H.
Cham-peme, J.Gest, and R. Callahan. 1986.Genetic alteration ofthe c-myc protooncogene (MYC) in human primary breast
carci-noma. Proc.Natl. Acad. Sci. USA83:4834-4838.
7. Graham,F.L.,S.Bacchetti, and R.McKinnon. 1980.
Transfor-mation of mammalian cells withDNAusing calciumtechnique,
p. 3-25. In R. Baserga, C. Croce,andG. Rovera(ed.), Intro-duction of macromolecules into viable mammaliancells. Alan R. Liss, Inc., NewYork.
8. Ham, R. G. 1982. Importance ofthe basalnutrient medium in the design ofhormonally defined media. Cold SpringHarbor
Conf. CellProliferation 9:39-60.
9. Hand, P. H., A. Thor, D. Wunderlich, R. Muraro,A. Caruso,
and J. Schlom. 1984. Monoclonal antibodies of predefined specificitydetect activatedrasgeneexpressioninhuman
mam-mary and colon carcinomas. Proc. Natl. Acad. Sci. USA 81: 5227-5231.
10. Kano-Sueoka,T. 1983. Growth ofratmammarytumor cells in
serum-free hormone-supplemented medium, p. 89-104. In D. W. Barnes, 1).A. Sirbasku, andG. H. Sato(ed.), Methods
forserum-free cultureofcellsoftheendocrinesystem. Alan R.
Liss, Inc. New York.
11. Kano-Sueoka,T., G. R.Campbell,and M.Gerber. 1977.Growth
stimulatingactivity inbovine pituitary extractspecificfora rat
mammary carcinomacell line. J. Cell. Physiol.93:417-424. 12. Kano-Sueoka, T., D. M. Cohen, Z. Yamaizumi, S. Nishimura,
M.Mori,and H.Fujiki.1979.Phosphoethanolamineas agrowth factor ofa mammary carcinoma cell line of rat. Proc. Natl.
Acad. Sci. USA 76:5741-5744.
13. Kano-Sueoka, T., and J. E. Errick. 1980. A pituitary-derived
on November 10, 2019 by guest
http://jvi.asm.org/
growth factor for rat mammary tumor cells:
phosphoethanol-amine,p.299-303. In L.Jimenezde Asua, R. Levi-Montalcini, R.Shields,andS.lacobelli (ed.), Control mechanismsin animal
cells: specificgrowth factors. Raven Ptess, New York. 14. Kano-Sueoka, T., and J. E. Errick. 1981. Effects of
phospho-ethanolamine andethanolamine on growth of mammary carci-nomacellsinculture. Exp. Cell Res. 136:137-145.
15. Kano-Sueoka, T., and J. E.Errick. 1982. Roles of
phosphoetha-nolamine, ethaphosphoetha-nolamine,andprolactin in mammary cell growth.
ColdSpring Harbor Conf. Cell Proliferation9:729-740. 16. Kano-Sueoka, T., J. E. Errick, and D. M. Cohen. 1979. Effects
of hormones and a novel mammary growth factor on a rat mammarycarcinoma inculture.ColdSpring Harbor Conf. Cell Proliferation 6:499-512.
17. Kano-Sueoka,T., J. E. Errick, D.King, and L. A. Walsh. 1983.
Phosphatidylethanolamine synthesis in ethanolamine-respon-sive and -nonresponethanolamine-respon-sive cells in culture. J. Cell. Physiol. 117: 109-115.
18. Kano-Sueoka, T., and P. Hsieh. 1973. A rat mammarycarcinoma in vivo and in vitro:establishment ofclonallines ofthe tumor. Proc. Natl. Acad. Sci. USA70:1922-1926.
19. Kano-Sueoka, T., and D. King.1987. Role of ethanolamine and
phosphatidylethanolamine in proliferation ofmammary
epithe-lialcells, p. 39-58. InJ. Enami andR.G. Ham (ed.),Growth anddifferentiation ofmammaryepithelial cells in culture.Japan
Scientific Societies Press,Tokyo.
20. Kano-Sueoka, T., and D. M. King. 1987. Phosphatidylethanol-aminebiosynthesisin ratmammarycarcinoma cells thatrequire
and do not require ethanolamine for proliferation. J. Biol. Chem. 262:6074-6081.
21. Kano-Sueoka, T., and D. M. King. 1988. Effects of phosphati-dylethanolamine and phosphatidylcholine in membrane phos-pholipidonbinding of phorbolesterinratmammarycarcinoma cells. Cancer Res. 48:1528-1532.
22. Kasid, A., M. E. Lippman, A.G. Papageorge, D. R. Lowy, and E.P.Gelmann. 1985. Transfectionofv-rusHDNAintoMCF-7 human breastcancercellsbypasses dependenceonestrogenfor tumorigenicity. Science 228:725-728.
23. Kozbor, D., and C. M. Croce. 1984.Amplification of thec-myc oncogene in one of five human breast carcinoma cell lines. Cancer Res. 44:438441.
24. Kraus, M. H., Y. Yuasa, and S. A. Aaronson. 1984. Aposition 12-activated H-ras oncogene in all HS578T mammary
carci-nomacells butnotnormalmammarycells ofthe samepatient. Proc. Natl. Acad.Sci. USA81:5384-5388.
25. Land, H., L. F. Parada, and R. A. Weinberg. 1983.Tumorigenic conversion of primary embryo fibroblastsrequires atleasttwo
cooperatingoncogenes. Nature(London)304:596-602. 26. Lechner, J. F., A. Haugen,I.A. McClendon, and E. W. Pettis.
1982.Clonal growth of normaladult humanbronchialepithelial cellsinaserum-free medium. InVitro(Rockville)18:633-642. 27. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular
cloning: alaboratory manual. ColdSpring Harbor Laboratory, ColdSpring Harbor,N.Y.
28. Minna, J. D.,D. N.Carney, H.Oie,P. A.Bunn, Jr., and A. F.
Gazdar. 1982. Growth of human small-cell lung cancer in defined medium. ColdSpring Harbor Conf. Cell Proliferation 9:
627-639.
29. Modjtahedi, N., C. Lavialle, M. F. Poupon, R.-M. Landin, R. Cassingena, R. Monier, and 0. Brison. 1986. Increased level of amplification of the c-myc oncogene in tumors induced in nude mice by a human breast carcinoma cell line. Cancer Res. 45: 4372-4379.
30. Murakami, H., H. Masui, G. H. Sato, N. Sueoka, T. P. Chow, and T. Kano-Sueoka. 1982. Growth of hybridoma cells in serum-free medium: ethanolamine is an essential component. Proc. Natl. Acad. Sci. USA 79:1158-1162.
31. Ossowski, L., J. C. Unkeless, A. Tobia, J. P. Quigley, D. B. Rifkin, and E. Reich. 1973. An enzymatic function associated
with transformation of fibroblasts by oncogenic viruses. II.
Mammalianfibroblast cultures transformed by DNA andRNA tumorviruses.J. Exp. Med. 137:112-126.
32. Parada, L. F., C. J. Tabin, C. Shih, and R. A. Weinberg.1982.
Human EJ bladder carcinomaoncogene ishomologue of Har-vey sarcomavirus ras gene. Nature (London)297:474-478.
33. Peehl, D. M., and T. A. Stamey. 1986. Serum-free growth of adult humanprostaticepithelial cells. In Vitro (Rockville)22: 82-90.
34. Perbal, B., and M. Rassoulzadegan. 1980. Distinct transforma-tion phenotypesinducedby polyomavirus andsimian virus40in
ratfibroblastsandtheir controlbyanearlyviral genefunction. J. Virol. 33:697-707.
35. Powers,S.,P. B. Fisher, and R. Pollack. 1984. Analysis of the reduced growth factor dependency of simian virus
40-trans-formed3T3cells. Mol. Cell. Biol. 4:1572-1576.
36. Riss, T. L., and D. A. Sirbasku. 1987. Growth and continuous
passage of COMMA-D mouse mammary epithelial cells in
hormonally defined serum-free medium. CancerRes.
47:3776-3782.
36a.Riss,T.L., B. H.Stewart,and D. A. Sirbasku. 1986.Growth of
GH4C1rat pituitary tumorcells in hormonally defined
serum-free medium.J.Cell Biol. 103:15.
37. Seif,R. 1980. Polyomavirus middletantigen: a tumor
progres-sionfactor. J. Virol. 35:479-487.
38. Southern, P. J., and P. Berg. 1982. Transformation of
mamma-lian cells to antibiotic resistance with a bacterialgene under control of the SV40early regionpromoter.J. Mol.Appl.Genet. 1:327-341.
39. Tegtmeyer,P., M.Schwatz,J. K.Collins,and K. Rundell.1975.
Regulation oftumorantigen synthesis by simian virus 40gene A.J. Virol. 16:168-178.
40. Theillet, C., R. Lidereau, C. Escot, P. Hutzell, M. Brunet, J.
Gest, J. Schlom, and R. Callahan. 1986. Loss ofa c-H-ras-1
alleleandaggressivehumanprimary breast carcinomas.Cancer Res.46:4776-4781.
41. Tsao, M.C., B.J. Walthall,and R.G. Ham.1982.Clonalgrowth ofnormal humanepidermal keratinocytes inadefined medium.
J.Cell.Physiol. 110:219-229.
42. Vennstrom, B., C. Moscovici, H. M. Goodman, and J. M. Bishop.1981. Molecularcloning of the avian myelocytomatosis virusgenome and recoveryof infectious virus by transfection of chickencells. J. Virol. 39:625-631.
43. Wurm, F.M.,K. A.Gwinn,and R. E.Kingston.1986.Inducible overproduction of themousec-mycproteininmammalian cells.
Proc. Natl. Acad. Sci. USA83:5414-5418.