0022-538X/87/113463-07$02.00/0
Copyright C) 1987, American Society for Microbiology
Varicella-Zoster
Virus-Specific
Cytotoxic T Lymphocytes (Tc):
Detection and Frequency Analysis of
HLA
Class
I-Restricted
Tc
in Human
Peripheral
Blood
JULIAN K. HICKLING, LESZEK K. BORYSIEWICZ, AND J. G. PATRICK SISSONS*
MRC Clinical Immunology ResearchGroup, Royal Postgraduate MedicalSchool, London W12 OHS, United Kingdom Received 24June1987/Accepted4August 1987
The cytotoxic T-cell (Tc) response to varicella-zoster virus (VZV) is incompletely characterized. We investigated whether VZV-specificTc restricted by class I products of the majorhistocompatibilitycomplexcan
begeneratedfrom theperipheral blood of VZV-immune donors. Cell lineswere established from peripheral
bloodlymphocytes (PBL)of seropositive donors by secondary in vitro restimulation. If cell-freeVZVwasused
asthestimulating antigen, theresulting lineswerepredominantly CD4+ and didnotshow class I-restricted cytotoxicity; whenautologous infected fibroblasts wereusedfor in vitro stimulation, the resultant lineswere
usually cytotoxic,although inonly4of11subjectstestedwasthiscytotoxicityHLArestricted and virus specific.
PBL were also tested for Tc activity without prior restimulation; VZV-specific Tc activity was only
demonstrable in thePBLofasubjectconvalescentfollowingzosterbutnotfrom subjects withrecentvaricella infectionorfrom normal subjects. VZV-specificTcprecursorfrequencieswerethendetermined in six selected
subjects bylimiting-dilution analysis. Ameasurable frequency was detectable in four of thesix seropositive
subjects, ranging from 11/106T cells inan asymptomatic carrier, to63/106 T cells ina subject with recent zoster.Weconcludethatvirus-specific majorhistocompatibility complex class I-restrictedTcprecursors may
be present intheperipheral blood of normal individuals seropositive forVZV butatafrequencylower than
thatforotherherpesviruses with nonneuronal sites of latency.
Varicella-zoster virus (VZV) is an alpha herpesvirus of humans. Primary infection may be associated with the
clinicalsyndrome ofvaricella(chickenpox), followingwhich thevirus establisheslatentinfection insensory nerveganglia
(7). Reactivation may subsequently occur and is associated
with dermatomallesions (zoster). An increased frequency of virus reactivation and greater likelihood of disseminated
infection are both associated with acquired or inherited
defects ofcellular, rather than humoral, immunity (12). It may thusbe inferredthat cellularimmune mechanisms are
important in controlling both primary infection by and
reactivation ofVZV.
Aknowledge ofthe natureoftheT-cell response to VZV
would aid understanding ofhow viruses establishing classi-callatency withinthenervous system maybecontrolledby the immune response. Theproliferative response of mono-nuclearcellstoVZVantigenshaspreviouslybeen studiedas a measure of T-cell immunity and has been shown to
increase during recovery from varicella (1, 10) and zoster
(15, 19). However, in comparison with the other human
herpesviruses,thecytotoxicTcell(Tc)responsetoVZV has
receivedrelativelylessattention.Tccanbedetected follow-ing infection with other human herpesviruses and are be-lievedtohavearole inlimiting reactivationepisodes during persistent infection. By using
techniques
of secondary in vitrorestimulation, class I-restricted Tclinescan be gener-atedfromperipheral bloodlymphocytes
(PBL)ofasympto-matic subjects
seropositive
for humancytomegalovirus
(HCMV),Epstein-Barrvirus(EBV),orherpes
simplex
virus(HSV) (2, 18, 21); HLAclass I-restrictedTc are
reportedly
directly detectable during HCMVinfection, and their
pres-* Correspondingauthor.
ence may be associated with survival from infection by marrowtransplant recipients (16).
More recently, Hayward et al. (9) have reported the
generation in vitro ofCD4+, class II-restricted Tc clones
specificfor VZV. SimilarCD4+ Tc clones specific for HSV havepreviously beengeneratedand have been showntobe
specific forHSVglycoproteins, in particular glycoproteinD (22).
Previous worksuggests that in general CD8+ HLA class
I-restricted Tc precursors (Tc-P) appear to be the
numeri-callypredominantTcpopulationin humanperipheral blood,
asshownby limiting-dilution analysis againsttargetcells in the presenceof lectin(13). Wethereforeset out todetermine
whetherCD8+, HLA class I-restricted VZV-specific Tc, or Tc-P could be demonstrated in the peripheral blood of
asymptomatic,
VZV-seropositive subjects. We reportmeth-ods by which VZV-specific Tc may be generated in bulk culture fromsome but notall seropositive donors. In
addi-tion, by usinglimiting-dilution analysis,weobtained
prelim-inary estimates ofthe frequency ofVZV-specific Tc-P in
normal subjects and in those recovering from zoster and found that the frequency of
VZV-specific
Tc-P was lowrelativeto thatfor herpesviruses with nonneuronal sites of latency.
MATERIALSAND METHODS
Virusstocks. Plaque-purifiedEllen strain VZV(American
TypeCultureCollection, Rockville, Md.)wasgrownin Flow 5000 humanembryo fibroblasts
(Flow
Laboratories,
Irving,
Scotland) by
passaging
infectedmonolayers
showing
75%cytopathic effect onto fresh
monolayers
at a ratioof 1:5 of infected to uninfected cells. VZV stocks were stored inliquidnitrogenascell-associatedvirusandwereshowntobe
mycoplasma free by culture and Hoechst
dye (no.
33258)
3463on November 10, 2019 by guest
http://jvi.asm.org/
staining. Plaque-purified HSV and CMV (ATCC AD169) stockswereharvestedfrom thesupernatant of infectedFlow 5000monolayersandstoredat -70°Cascell-free virusat106 to
107
PFU/ml.Human fibroblast lines. Primary human fibroblast lines
wereestablishedfrom skinbiopsiesfromHLA-typeddonors as described by Borysiewicz et al. (2). Primary fibroblasts
were grown in Eagle minimum essential medium (Flow)
supplemented with10% fetal calfserum(Imperial
Laborato-ries), 1% nonessential amino acids (GIBCO Ltd., Paisley, Scotland), 2 mM L-glutamine, 105 IU ofpenicillin perliter, and 100mgofstreptomycin (Flow)perliter.
PreparationofT-cell lines. Donors werehealthy
seroposi-tive individuals asjudged by a titer of complement-fixing
antibody to VZV ofgreater than 1 in 8 and/or lymphocyte
proliferationinresponsetoVZV.Peripheralblood
mononu-clearcells(PBM)wereobtainedby Ficoll-Paque (Pharmacia Fine Chemicals, Uppsala, Sweden) gradient centrifugation andsuspendedat106/ml in 24-wellplates (BectonDickinson Labware, Oxnard, Calif.) in RPMI 1640 (Flow) with 10% heat-inactivated human AB serum. The stimulating antigen
was either 5 x 104autologous VZV-infected fibroblastsper
well (autologous or partially matched for HLA A and B antigens) or cell-free VZV antigen. Cell-free VZV antigen was prepared by sonication of infected fibroblasts(showing greaterthan 90% cytopathic effect)followedby heat inacti-vation at 56°C. This wasused ata concentration shown to induce proliferative responses as measured by tritiated thymidine uptake in preliminary experiments. Interleukin 2 (IL-2;CellularProducts, Buffalo, N.Y.)wasaddedonday3 (or day7 inearly experiments) andevery subsequent3 or4
days. Fresh antigen was added on day 7, with autologous irradiated(3,500rads) PBMasfeeder cells(106/ml). After 7 to 26 days invitro, cells were harvested, washed, and used
aseffector cells ina6-h51Cr-releaseassay.Thephenotypeof the cells wasdetermined byflowcytometry.
Cytotoxicity assay. Target cellswere autologous (orHLA A- and B antigen-matched) and HLA-mismatched fibro-blasts,used eitheruninfectedorinfectedwithcell-associated VZV(1:5ratio ofinfectedtouninfectedcells)72 hbefore the
assay.Targetcellswerelabeledwith51Cr(200 p.Ci/106cells;
Amersham International, Amersham, England) overnight, trypsinized, washed thoroughly, and suspended to 5 x
104/ml. Target cells(100 pulperwell) wereadded to96-well round-bottom microtiterplates (Becton Dickinson) already containing 100 pl of effector cells, giving the required effector-to-target-cellratiosintriplicate. After 6h, 100 of supernatantwasharvested,andradioactivitywascounted in
a gamma-counter (LKB Wallac Multigamma, Turku, Fin-land); specific chromium release was calculated by the standardformula. Maximumrelease wasobtained by using 1% TritonX-100. Average spontaneous release from VZV-infected target cells was 22.5% of maximum in 6-h assays and 30% of maximum in 18-h assays. At least 70% ofthe VZV-infected target cells were always positive by indirect
immunofluorescence by using zoster-immune globulin (Cen-tral Public Health Laboratory Service, Colindale, England) and fluorescein-conjugated rabbit anti-human immunoglob-ulinG, A, andM, kappaand lambda(Dakopatts, Glostrup, Denmark).
Limiting-dilution analysis. PBL obtained from Ficoll-Paque centrifugation were incubated with AET (Sigma Biochemicals, Poole, England)-coated sheep erythrocytes (Tissue Culture Services, Slough, England) (4), and the AET-E+ population was collected after centrifugation on
Ficoll-Paque and suspended in RPMI 1640 with 10% fetal
calfserum atconcentrationsrangingfrom 5 x 105/mlto 1 x
104/ml. Cellsuspension (100
RI)
wasadded toround-bottom microtiterwells,
with 30 replicate wells at each concentra-tion. Each well alsocontained 1 x105
irradiatedautologous
PBM asfeeder cells and 2 x
103
VZV-infectedautologous
fibroblasts. Inone
experiment, lymphocytes
wereincubatedwith concanavalin A (5 ,ug/ml) for 48 h and then used as
responder cells in the absence of antigen. This
polyclonal
stimulationprotocolresulted infrequencies comparablewith those from antigen stimulation in control experiments. Su-pernatantfromMLA 144 cells wasusedas a sourceof IL-2 and was present from day 0. Cultures were refed every 3
days and maintained for 9 to 12 days. At the end of this
period, each well was split into four aliquots for testing againstfour targets ina12-h51Cr-releaseassay.Targetsused were K562in the presence ofphytohemagglutinin (PHA)(4 pug/ml), VZV-infectedautologous fibroblasts, CMV-infected
autologous fibroblasts, and uninfected autologous fibro-blasts. A positive result against a given target was definedas aresult greater than 3 standard deviations above spontane-ous release obtained in the presence of target and feeder cells only. The number of positive wells mediating lysis of VZV-infected targets and K562 plus PHA but not control targetswasdetermined for each concentration. The propor-tion ofnegativewellsateach concentrationwasdetermined and plotted on alog scale against input cell number per well. The frequency was calculated from the input cell number giving 37% negative wells. Tc-P frequencies were calculated by using least mean squares regression analysis(correlation
coefficient, r > -0.9 in most cases) (11) and maximum likelihood analysis (5). There was no significant difference between the results obtained by each method.
Determinationof cellphenotype. Aliquots of effector cells were incubated for 45 min with anti-Tll, -T8, -T4 (Coulter
Electronics Ltd., Luton, England) or Leu-7 (Becton Dickinson) at a 1/40 dilution, followed by the addition of
fluorescein-conjugated goat anti-mouse immunoglobulin (Coulter). Analysis was performed on an EPICS-C flow cytometer.
RESULTS
Secondary in vitro stimulation with cell-free VZV. Data from other workers (1, 10, 15, 19) and preliminary
experi-ments performed by us show that PBL from subjects
seropositive for VZV proliferate in response to cell-free VZV antigen preparations (data not shown). Experiments wereperformedtodetermine whether therespondingcells in these cultures were capable oflysing VZV-infected fibro-blasts.
PBL from healthy seropositive subjects were cocultured with heat-inactivated cell-free VZV and expanded in the presence of IL-2. Cultures weremaintainedfor 7 to 26 days before being tested in a 6-h 51Cr-release assay. Lines from seven donors were tested, but no cytotoxic activity was observedagainst autologous ormismatched VZV- or HSV-infectedfibroblasts (data not shown). The phenotype of the
respondingcells wasgreater than80%CD2+, and the ratio of
CD4+ toCD8+ was alwaysgreater than 2:1 (Table 1).
Secondary in vitro restimulation with infected autologous fibroblasts. Twomajor possibilities, notmutually exclusive, could explain the failure to generate VZV-specific HLA class I-restricted Tc by cococulture with cell-free antigen.
First,itmightbe thatantigen isnotpresented in the context ofappropriate major histocompatibilitycomplex antigens, or
second, theappropriate VZV antigens required to generate
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Phenotypes ofshort-term T-cell bulk cultures established in response to VZV
Stimulating antigen %Positive (range)
CD4+/CD8+b
(no.of subjects) CD2+ CD4+ CD8+ Leu-7
Cell-free VZV (n = 6) 88.8 (84-96) 60.7(50-81) 18.5 (6-25) 5.6 (2-11) 4.98 + 4.7
VZV-infected fibroblasts (n = 10) 84.8(72-97) 28.9 (15-46) 46.6 (30-60) 5.6(2-8) 0.63 ±0.23 aExpressed as mean percent of positive cells.
bMeanratio of CD4+ to CD8+ + 1 standard deviation.
Tc invitro mightbe absent fromthecell-free virion prepa-ration. In view of ourpreviousexperiencewithCMV (2), the latter possibility was investigated by using VZV-infected
autologous fibroblasts as the stimulating antigen; these wouldcontain nonstructuralVZVproteins, absent from the
cell-free preparation. IL-2 wasaddedonday 7 of culture(in
later experiments, the addition of IL-2 on day 3 made no
difference intheresults).
In contrast to theresultsobtained from lines generated in
response to cell-free antigen, cultures stimulated with in-fected fibroblastswerepredominantly CD8+ (meanratioof CD4+ to CD8+, 0.63) (Table 1). The cytotoxic activity observed in 11
asymptomatic
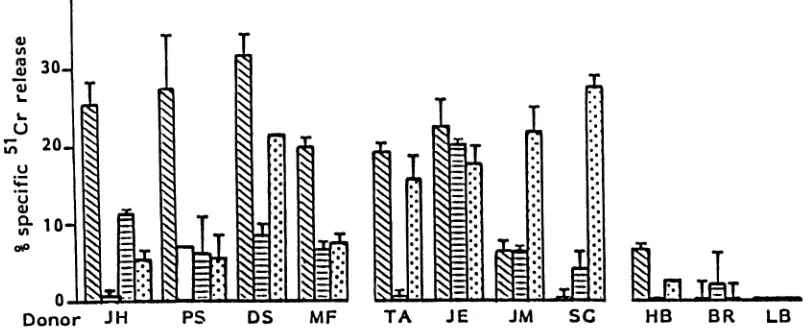
seropositive donors could bedividedinto threegroups(Fig. 1).Lines from4of11donors
showed HLA class I-restricted cytoxicity against
VZV-infected targets (JH, PS, DS, and MF). Linesfrom donors
TA, JE, JM, and SG displayedcytotoxic activity, although
this was not HLA restricted. Finally, lines from a further three donors (HB, BR, and LB) displayed little or no
cytotoxicity. Inrepeatexperiments, T-cell lines established fromanindividualsubjectgenerallygave the same patternof
response.VZV-specific, class I-restrictedTc lines have been
consistently generated from donor JH (7 of10lines
gener-atedshowed class I-restricted cytotoxicity).
To confirm the virus specificity ofthe responding cells, lines from two donors (JH and PS) which showed
HLA-restricted cytotoxicity against VZV-infected targets were
also tested against autologous fibroblasts infected with HSV (Fig. 1). HSV-infected autologous fibroblastswerenotlysed above background levels.
Studies on subjects with recent varicella or zoster. The
failure to generate VZV-specific Tc lines from some
seropositive donors mightbe due to alow frequency of Tc-P cells in these individuals. To determine whether the fre-quencyof VZV-specificTc wastemporallyrelated torecent
primary infection orreactivation, donors who hadrecently experienced
varicella
or herpes zoster were tested. Donor MF had varicella 12 months before testing, and donorsTA and HB had varicella 2 months before testing. When their PBL were stimulated with infected fibroblasts, the resultsdid not differ significantly from those obtained in other seropositive subjects (Fig. 1). A Tc response was obtained from MF,although notfrom the subjectswith more recent
infection (TA and HB).
Inaddition,PBLfromthesesubjectsweretesteddirectly in
51Cr-release
assays without prior secondary in vitro restimulation (Fig. 2). There was no significant lysis at 6h,and therefore the results from the 18-h assays are shown. Four donors with recent varicella (TA, MH, HB, and EA)
showednopreferential lysis of VZV-infectedtargetssharing
one or two HLA class I A and B antigens (autologous fibroblasts were not available) compared with
HLA-mismatched fibroblasts. Similar results were obtained with
asymptomatic seropositive subjects with no recentepisodes of varicella or zoster. The
nonspecific lysis
observed with donorEA(whohadvaricella2monthspriortoassay)could be reduced by rosetting the effector population with oxerythrocytes coated with rabbit ox erythrocyte anti-body. The remainingcellpopulation mediated virus-specific HLA-restricted lysis(Fig. 3).
PBL from two subjects with recent episodes of zoster werealsotestedwithoutpriorrestimulationinvitro, i.e., PC
and SB, 3 months and 4 weeks, respectively, after their initialpresentation. Bothhadincreasedlevelsof CD8+ cells
40.
30-20)
0J
Donor JH PS DS MF TA JE
.JM
SG HB BR LBFIG. 1. Cytotoxicityexpressed byT-cell linesestablishedinresponsetoinfected fibroblasts. Culturesweretestedonday14. Resultsare expressedasspecific 51Cr release obtainedat aneffector-to-T-cell ratio of 20:1(+ 1 standarddeviation)in a6-hassayfrom thefollowing targets. F, Autologous (or partially HLA class I-matched) VZV-infected fibroblasts; i, Autologous uninfected fibroblasts; g,
HLA-mismatched,VZV-infectedfibroblasts; [1, autologousHSV-infectedfibroblasts.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.102.507.515.680.2]30 w
u 2
O10
OlS
AT-
SIV
Donor CC TA MH EA HB PC SB
FIG. 2. Cytotoxicity
produced directly by
fresh PBL from donors withrecentvaricellaorzoster. Barsrepresent51Cr
release inan 18-hassayat aneffector-to-T-cell ratio of 20:1 (+1 standard deviation). Subjects TA, MH, EA, andHB weretested2months afterpresentation with varicella. Subject PC was tested 3 months after presentation with zoster. Subject SB was tested 3 weeks afterpresentation with zoster. Subject CG is anasymptomaticseropositive control. For explanation of bars,seelegendtoFig. 1.
intheirPBL(CD4+ toCD8+ ratio of 0.5:1 in both cases as determined byflowcytometry). Donor SB showed no
pref-erentiallysis of VZV-infectedtargetssharingtwo HLAclass Iantigens compared with completely HLA-mismatched con-trols. In contrast, PBL from PC exhibited class I-restricted
lysis of VZV-infected fibroblasts. HSV-infected autologous
fibroblasts matched at three class I loci and
HLA-mis-matched fibroblasts were not lysed, suggesting that
VZV-specificTc were present in PBL at a sufficientfrequencyto
be detected without invitrorestimulation.
Limiting dilution analysis ofVZV-specific Tc-P frequency. To measurethefrequency ofVZV-specific Tc-Pin different individuals and compare it with those for other human
herpesviruses, we used the technique of limiting-dilution analysis as described in Materials and Methods (and by
L. K. Borysiewicz, S. Graham, J. Hickling, C. Green, and J. G.P. Sissons [submitted for publication]).
40
-as
In -) 30
-L.
i
20-u
m10_
uI
0-O
-IIt
J .
-1 I
-- I
A
Theability of cells from each well in the assay to mediate
lectin-dependent cytotoxicitywas used to measure the fre-quency of all cytotoxic cells regardless oftheir specificity
and was between 67 and 500 per 106Tcells. Results from a
typical experiment are given in Fig. 4, and a summary of
results from sixsubjects isgivenin Table 2. In all cases, the
frequency of Tc-P mediating specific lysis of autologous
uninfected and CMV-infected fibroblast target cells was low orundetectable. In twoasymptomatic seropositivesubjects
(SH and AR), the VZV-specific-TcP frequency was no greater than that for uninfected target cells. However,
de-tectable frequencies of 11 and 29 per 106 T cells were
obtained from asymptomatic subjects JH and DS,
respec-tively; PBL from both these donors had previously been
observed to give rise to VZV-specific, class I-restricted
cytotoxiclines in the bulk culture system(Fig. 1).
The highest VZV-specific Tc-P frequency (63
VZV-407
30_
50 25 12.5
E:T
6.25
B
I I I
-I
50 25 12.5 6.25
E:T
FIG. 3. Cytotoxicity expressed by fresh PBM from subject EA 2 months after varicella. (A) Unfractionated PBM. (B) After depletion of FcR+ cells. Cells were tested in an 18-h51Cr-releaseassayagainst VZV-infected autologous fibroblasts(0), uninfected autologous fibroblasts
(0),VZV-infected HLA-mismatchedfibroblasts (A), and K562 cells (A). Standard deviations were<10%of meanspecific-release values.
20-
10-oi
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.110.517.66.264.2] [image:4.612.154.473.511.686.2]Input
cells/well,
104
0 1.0
.0
t, .6
ci
.
.0 .37-,
C
o
0
L.
IL
.2
2 4 6 8
,e. ,eeseeeS 59
10
1/9x
104
1/4 x
103
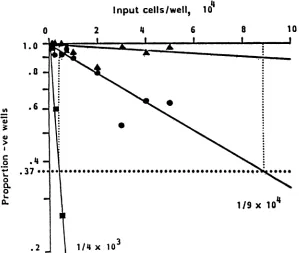
FIG. 4. Frequency analysis of VZV-specific Tc-P in
donor
JH. The results fromlimiting-dilution analysis are shown as asemilogplot of results from a representative experiment. After 13 days in vitro, cells from each well were split and tested against K562 plus PHA(a), autologous VZV-infectedfibroblasts (O), and uninfected fibroblasts (A). There was no lysis of CMV-infected fibroblasts. It can be seen that thecalculated precursor frequency cells mediating lectin-dependent cytotoxicity was 1 in 4,000, and for VZV-specific Tc, the frequency was 1in90,000 (250 and 11per 106Tcells, respectively). Lines were plotted by using linear regression analysis.specific Tc-P per
106
T cells) was detected in PBL fromdonor DB, who was tested 3 months after an episode of zosterbutwasotherwisehealthy.Incontrast, donorSBwas
tested 4 weeks afterzoster butwas on immunosuppressive therapyfollowingarenalallograft. Inthisexperiment, PBL
were polyclonally activated with concanavalin A for 48 h
prior to plating out in limiting dilution, as opposed to
stimulation
withVZV-infectedfibroblasts.Thefrequency ofVZV-specific
Tc-Pobtained(29 VZV-specific Tc-P per 106 Tcells)
is
comparable withthehighest frequency observed in anyasymptomatic seropositive subject.DISCUSSION
Inthisreport, wehavedescribedamethodofsecondaryin vitro restimulation which is capable of generating major
histocompatibility
complex class I-restricted VZV-specific Tc lines from a proportion of asymptomatic seropositiveTABLE 2. Frequencies ofVZV-specificTc-Pderivedfrom limiting-dilution analysis
No.ofspecificTc-Pper106Tcells withtarget:
Donor K562 cells VZV-infected CMV-infected Uninfected
+ PHA fibroblasts fibroblasts fibroblasts
JH 250 11 <2.0 2.5
DBW 100 63 <2.0 <2.0
SH 500 <2.0 <2.0 3.3
AR 115 <2.0 <2.0 <2.0
DS 122 29 <2.0 <2.0
SBb 323 29 <2.0 <2.0
aTested3monthsafterpresentationyvithzoster.
bTested 4weeks after presentation with zoster and receiving
immuno-suppressive therapyattime oftesting.
donors. VZV-specific Tc were only generated if infected
fibroblasts, rather than cell-free VZV, were used as the
stimulating antigen. Thefrequency of such cells, estimated
bylimiting-dilution analysis,wasbetween29 and <2 per 106 cells in thoseindividuals we studied. Furthermore, prelimi-nary results suggest that there is little difference in the response produced by donors 2monthsafteranepisode of
varicella compared with that of asymptomatic seropositive
individuals. However, a recent episode ofzoster may be
associated with an increased frequency of VZV-specific
Tc-Pasmeasured bylimiting-dilution analysis.
The importance ofthevirus preparationused to restimul-atePBLis shownbythegeneration ofpredominantlyCD4+ T-cell lines to cell-free virus and CD8+ T-cell lines to
autologous virus-infected fibroblasts, as previously shown for HCMV(2).Oneexplanation forthis is that nonstructural
virus antigens expressed only in the infected cells are
dominanttargetantigens for Tc,asinthe caseof HCMV(2). Thispossibility is difficulttoinvestigate because the strongly cell-associatednatureof VZVmitigates against synchronous infectionand thesubsequentuseofmetabolic inhibitors such
as phosphonoformate which only permit
expression
ofthe virus immediate-early and early proteins. An alternativeexplanation isthat naturalvirusinfectionof stimulatorcells may beimportantifendogenoussynthesisof viralproteinsis
required for their presentation in the context of major histocompatibility class I antigens (6), as shown for the
generationofinfluenza
hemagglutinin-specific
Tc(14). However, evenbyusing autologous
infectedfibroblasts,
classI-restricted
VZV-specific
Tclineswereonly
generatedin one-third ofthedonors tested. In the other
subjects,
thenonspecific cytotoxic
activity
waspresumably
attributableto natural killer (NK) or
lymphokine-activated
killer cells.on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.154.453.64.317.2] [image:5.612.53.295.596.692.2]This variation among seropositive subjects could reflect a
lower frequency of VZV-specific Tc-P in some donors. In
view
of thispossibility,
weused
limiting-dilution analysis
toquantitatively
assess thefrequency of VZV-specificTc-P in a small group of seropositive subjects. A measurable fre-quency ofVZV-specific
Tc-P was detected in two of fourhealthy
asymptomatic subjects. Both thesesubjects hadalsogiven
good responses in thebulk culture system, suggestingthat there maybeacorrelation betweenTc-Pfrequencyand the
ability
to generate Tcin culture. However, theseVZV-specificTc-Pfrequenciesare considerably lower than those
described forotherherpesviruses. Forinstance, ananalysis of HCMV-specific Tc-P frequencies in asymptomatic
sub-jects seropositive
for both HCMVand VZV suggests that theHCMV-specific
Tc-Pfrequencies
are in the range of50 to200 per 106 T cells, at least fivefold higher than those for
VZV
(Borysiewicz
et al., submitted). In addition, the fre-quency of EBV-specific Tc-P (although estimated by adifferent
method,
theregression
assay) is reportedly in the range of 100 to 1,000 per106
T cells (17). The highestVZV-specific
Tc-P frequencies were detected in subjects withrecentepisodes ofzoster(DB and SB). These increasedlevels of Tc-P are consistent with recent restimulation in
vivoand suggest that such cells mayplayarole incontrolling reactivation
episodes.
It is possible that the VZV-specific Tc-Pfrequency
observed in SB was reduced owing to theimmunosuppressive
therapy the subject was receiving, ashas been
previously
reported for EBV-specific Tc-P(20).
Owing
to thedifficulty
infinding patients
with acutezosterwith haplotypes matching those available in our panel of
primary
fibroblast lines, relatively few donors have beentested to
date,
and noneduring
the acute phase of thedisease.
Previousattempts to compare the levels of T-cell
immu-nity
to VZV insubjects
with varicella or zoster havefocussedon
lymphoproliferative
responses toVZVantigens(1, 10, 15, 19).
These have shown that the mean in vitroblastogenic
response declines to low levels within 30 to 50days
oferuption
of the varicella rash yet still remainsextremely
high
for thesamelength
of time after the appear-anceofzoster. This would beconsistentwith
ourresultsoncytotoxic
cells obtainedfromlimiting-dilution
analysis. There are alsoprevious
reports onthedetection ofVZV-specific
Tc.Hayward
et al. (9)recently
reportedthegener-ation of
VZV-specific CD4+
Tcclones whicharecapableoflysing
HLA-DR-matched EBV-transformed B-cell linessuperinfected
withVZV. Itshould be noted thatitispossiblethatthe
CD4+
linesgeneratedin ourexperiments didpossesscytotoxic potential
but that such activitywas notdetectableby using primary
fibroblasts astargetcells, as they do not express HLA class IIantigens,
which are the restricting elementfor theexpression
of effector function by CD4+ T cells.Inanotherstudy, Bowden etal. (3) used a direct
cytotox-icity
assaysystem
(without restimulation in vitro) toinves-tigate lysis
ofVZV-infected fibroblastsby PBL fromasymp-tomatic
healthy subjects.
They found that cytotoxicitywasenhanced
by
matchingthetarget and effector cells for HLA class Iantigens (in
contrast tothe results we obtained) and was reduced by panningthe effector populations with anti-T8monoclonalantibodies, suggestinga role for CD8+Tc. Inaddition,
monoclonal antibody blockingand depletionstud-ies also indicated involvement of CD4+ and NK cells. Furtherworkby Haywardetal. (8) also suggested that NK cells contribute to the lysis of VZV-infected fibroblasts in sucha systemand that it is difficult to distinguish between
lysis mediateddirectly by Tc andthatmediated byNK cells amplified by lymphokine release from T cells. We
assayed
PBLfrom both normalseropositive donorsand donors with
recent varicella indirect cytotoxicity assays(in addition to assays after secondary restimulation in vitro). PBL from
both groups exhibited non-HLA-restricted
cytotoxicity
when assayed directly. Although we have not performed
detailed studies to determine the phenotype of the cell
population(s) mediating this nonspecific lysis, it was noted that the population responsible couldbe removed by
deple-tion of FcR+ cells, consistent with there being an NK component. Two subjects with recent zoster were also tested in direct cytotoxicity assays without preculture of
PBL, and cells from one of these subjects (PC) mediated VZV-specific, HLA class I-restricted cytotoxicity.
In summary, we demonstrated the presence of major histocompatibility complex class I-restricted Tc-P
specific
for VZV in the peripheral blood ofsome seropositive sub-jects. Our initial results suggest that these Tc-P are present at a relatively low frequency, which may be higher after
reactivation of VZV. It might be supposed that
herpes-viruses exhibiting classical neuronal latency, such as VZV, would produce less restimulation in vivo than those which
persist in lymphoid and epithelial cells, with intermittent
virus production, such as EBV and HCMV, and this could be one explanation for the relatively low frequency of
VZV-specific Tc-P. Further work is needed to define the precise specificity of VZV-specific Tc-P. The use of expres-sion systems containing isolated VZV genes would provide a means of sodefining specificity, and we are currently pur-suing this end.
ACKNOWLEDGMENTS
We are grateful to J. R. Batchelor, Department of Immunology, Royal Postgraduate Medical School, for the HLA typing, and we thank K. Bateson for preparation of themanuscript.
This work is supported by the Medical ResearchCouncil. J.K.H. is in receiptofa Medical Research Council researchstudentship, and L.K.B. isa ListerResearchFellow.
LITERATURECITED
1. Arvin, A. M., C. M. Koropchak, B. R. G. Williams, F. C. Grumet, and S. K. H.Foung. 1986. Early immuneresponse in healthy and immunocompromised subjects with primary varicella-zoster virus infection. J. Infect. Dis. 154:422-429. 2. Borysiewicz, L. K., S.Morris, J. D. Page, and J. G.P. Sissons.
1983. Humancytomegalvirus-specificcytotoxic Tlymphocytes: requirements for in vitro generation and specificity. Eur. J. Immunol.13:804-809.
3. Bowden, R. A., M. S.Levin, R. H. Giller, D. G. Tubergen,and A. R. Hayward. 1985. Lysis of varicella zoster virus infected cells by lymphocytes from normal humans and immunosup-pressed pediatric leukaemic patients. Clin. Exp. Immunol. 60:387-395.
4. Falkoff, R. J. M., M. Peters, and A. S. Fauci. 1982. T cell enrichmentanddepletion of human peripheral blood mononu-clear cell populations: unexpected findings in the study of functional activities of the separated populations. J.Immunol. Methods50:39-49.
5. Fazekas DeSt. Groth,S.1982. The evaluation of limiting dilution assays. J. Immunol. Methods 49:1211-1223.
6. Germain, R. N. 1986. The ins and outs ofantigenprocessingand presentation. Nature (London)322:687-689.
7. Gilden, D. H., A. Vafai, Y. Shtram, Y. Becker, M. Devlin, and M. Wellish. 1983. Varicella zoster virus DNA inhuman sensory ganglia.Nature (London) 306:478-480.
8. Hayward, A. R., M.Herberger, and M. Lazslo. 1986. Cellular interactionsin thelysisof varicellazoster virus infected human
on November 10, 2019 by guest
http://jvi.asm.org/
fibroblasts. Clin. Exp. Immunol. 63:141-146.
9. Hayward, A. R., 0. Pontesilli, M. Herberger, M. Laszlo, and M. Levin. 1986. Specific lysis of varicella zoster virus-infected B lymphoblasts by human T cells. J. Virol. 58:179-184,
10. Kumagai, T.,Y.Chiba,Y.Wataya, H. Hanazono, S. Chiba, and T. Nakao. 1980. Development andcharacteristics of the cellular immune response to infection with varicella zoster virus. J. Infect. Dis. 147:7-13.
11. Lefkovits, I., and H.Waldmann. 1979. Limiting dilution analysis of cells in the immune system. Cambridge University Press, Cambridge.
12. Merigan, T. C., and D. A. Stevens. 1971. Virus infections in man associated with acquired immunologicaldeficiency states. Fed. Proc. 30:1858-1864.
13. Moretta, A., G. Pantaleo, L. Moretta, M. C. Mingari, and J. C. Cerrotini. 1983. Quantitative assessment of the pool size and subset distribution ofcytolytic Tlymphocytes within resting or alloactivated peripheral bloodTcellpopulations. J. Exp. Med. 158:571-585.
14. Morrison, L. A., A. E. Lukacher, V. L. Braciale, D. P. Fan, and T.J.Braciale. 1986.Differences in antigen presentation to MHC class I andclass II restricted influenza virus-specific cytolyticT lymphocyte clones. J. Exp. Med. 163:903-921.
15. Patel, P.A., S. Yoonessi, J. O'Mally,A.Freeman,A.Gershon, and P.L.Ogra.1979.Cell mediatedimmunity to varicella zoster virus infection in subjects with lymphoma or leukaemia. J. Pediatr. 94:223-230.
16. Quinnan, G. V., N. Kirmani, E. Esber, R. Saral, J. F. Manischewitz, J. L. Rogers, A. H. Rook, G. W. Santos, and W.H.Burns. 1981.HLArestrictedcytolyticTlymphocyte and nonthymic cytotoxic lymphocyte responses to cytomegalovirus infection of bone marrow transplant recipients. J. Immunol. 126:2036-2041.
17. Rickinson, A. B., D. J. Moss, L. E. Wallace, M. Rowe, I. S. Misko, M.A.Epstein, and J. H. Pope. 1981. Long-term T-cell-mediated immunity to Epstein-Barr virus. Cancer Res. 41:4216-4221.
18. Rickinson, A. B., L. E. Wallace, and M. A. Epstein. 1980. HLA-restricted T cell recognition of Epstein-Barr virus-infected B cells. Nature(London) 283:865-867.
19. S0rensen, 0. S.,S.Haahr,A.M0ller-Larsen,and K.Wildenhoff. 1980. Cell-mediated and humoral immunity to herpesviruses dur-ing and after herpes zoster infections. Infect. Immun. 29:369-375. 20. Yao, Q. Y., A. B. Rickinson, J. S. H.Gaston, and M. A. Epstein. 1985.Invitroanalysis of the Epstein-Barr virus: hostbalance in longtermrenal allograftrecipients. Int. J. Cancer 35:43-49. 21. Yasukawa, M., T. Shiroguchi, and Y. Kobayashi. 1983.
HLA-restricted T lymphocyte-mediated cytotoxicity against herpes simplex virus-infected cells in humans. Infect. Immun. 40:190-197.
22. Zarling, J. M., P. A. Moran, L. A. Lasky, and B. Moss. 1986. Herpes simplex virus (HSV)-specific humanT-cell clones rec-ognize HSV glycoprotein Dexpressed byarecombinant vac-cinia virus.J. Virol. 59:506-509.