Identification of
a
Doubly Spliced
Viral Transcript
Joining the
Separated Domains for Putative
Protease
and
Reverse
Transcriptase
of
Hepatitis B Virus
PEI-JER CHEN, CHIOU-RONG CHEN, JUEI-LOWSUNG, ANDDING-SHINNCHEN*
Graduate Institute of Clinical Medicine, HepatitisResearch Center, and Department of InternalMedicine, National
Taiwan
University
College
of
Medicine and theUniversity
Hospital, Taipei,
Taiwan10016,
Republic of
ChinaReceived24 March1989/Accepted13 June 1989
Hepatitis Bvirus(HBV), like retroviruses, replicates through reversetranscription. However, the identity
and mechanism for the synthesis of HBV reverse transcriptase remain unknown. The open reading frame
(ORF) forHBVputativereverse transcriptase(pol), as a consequenceof overlapping with the whole ORF of
envelope proteins (hepatitis B surface antigens), includes a hypervariable region at the N terminus. Thus,
comparedwithretroviruses, it isunlikely that HBVreversetranscriptase is translated fromcompletepol ORF inthe full-length pregenomicRNA. We havenowdetected ininfected human liversanoveldoublysplicedRNA in whichonesplicingeventremoved the hypervariable region of thepolgenebutretained the conserved region
homologoustoretroviral reversetranscriptase. The other splicingeventdeleted the central region of hepatitis B core antigen and thus brought the protease domain which is important for maturation of reverse transcriptase closetothat ofpol. For thissequenceorganization,the spliced RNAasthepossible template for thesynthesis ofHBVreversetranscriptase isdiscussed.
The hepatitis B virus (HBV) has been showntoreplicate throughreversetranscription (18, 27), and its overall genetic
organization is similar to that of retroviruses in containing
open reading frames (ORFs) for (i) the nucleocapsid protein (hepatitis Bcoreantigen [HBcAg]); (ii) the envelope proteins
(large, middle, and small hepatitis B surface antigens [HB-sAgs]); (iii)theputativereversetranscriptase (pol ORF),and
(iv) a protein of unknown function (hepatitis B X ORF),
respectively. However, the size of HBVgenomeis smaller, about 3.2 kilobasepairs (kbp)inlength, largelyas aresult of
the overlapping of the pol ORF with the whole ORF of envelope proteins (HBsAgs) (30).
The genetic organization of HBV creates a potential
problembecause of thepresenceofahypervariable regionin
HBsAgs. It has been noted that the N-terminal ORF of the envelope protein (pre-Sl and S2, about 500 bases) is the
most variable region in the genome of different HBV
sub-types (20). Thefrequency of nucleotide substitution in this
regionis about15%, comparedwith 5%for the totalgenome
(21). Many of the nucleotide sequence changes in this
hypervariable region affect not only the amino acid
se-quencesofpre-Sl and S2 but alsodramatically affect the N terminus of theputativepol ORF,whichoverlaps thepre-S
sequence in a differentreading frame (21). Incontrast, the
sequencesof the middleregionof thepol ORF,whichshares
some homology with retroviral reverse transcriptase, are
highlyconserved(5, 6). Thus, ithas beensuggestedthat the
part of the pol ORF that overlaps with the hypervariable region of HBsAgs (pre-Si and S2) is too variable to be
functional (21, 25). In a recent report, the putative viral
reverse transcriptase molecule identified by specific
anti-bodyis also noted to lack the N terminus of thepol ORF
(13). Nevertheless, how HBV overcomes this problem to
remove the hypervariable regionremains unknown. In this report, we present evidence for the presence of a novel
spliced mRNA species in HBV-infected human livers. This
* Correspondingauthor.
mRNA is coterminal with pregenomic RNA but lacks the hypervariable region ofpol, and we propose that it is a
candidate mRNA for the synthesis of HBV reverse
tran-scriptase.
MATERIALS ANDMETHODS
Primary hepatocellular carcinomas. Tissueswereobtained
fromtwochronicHBsAg carriers who developed
hepatocel-lularcarcinomas (HCCs)and underwentsurgical resection. Thetumortissuesweredissected fromnontumor partsofthe
liver and both were immediately cut into smallpieces and
thenputinto liquid nitrogenuntil use.
Southern blot analysis. The cellular DNAs were isolated
from tissuesby sodium dodecyl sulfate-protease Kdigestion
and phenol-chloroform extraction as described previously
(12). For blotting, 10 ,ug ofDNAswaseithernondigestedor
digested with EcoRI, HindIII, and BamHI (Bethesda Re-search Laboratories, Inc., Bethesda, Md.) in the
recom-mended buffers. The DNAswerefractionatedby
electropho-resis ina0.7% agarosegel, transferredtonylon membrane
(Nytran;Schleicher &Schuell, Inc., Dassel,Federal
Repub-lic ofGermany), and thenhybridizedwith an HBV-specific
probe (kindly provided by W. S. Robinson, Stanford
Uni-versity, Stanford, Calif.).
Northern (RNA)blotanalysis. RNAswereextracted from
tissues by the guanidium isothiocyanate-cesium chloride
centrifugationmethod(4). Poly(A)+ RNAswere purified by
passagethrough anoligo(dT) columntwice; theywerethen
preservedin 70% ethanol at -20°C. For Northernanalysis, poly(A)+ RNAswere glyoxalated, separatedinagelof 1%
agarose, transferred to nylon membrane, and then
hybrid-ized(3).
Construction and screening of cDNA library. For cDNA
library construction, 2 ,ug of poly(A)+ liver RNAs were
converted to double-stranded DNAs by the method of
Gubler and Hoffman (7) by using a kit supplied by
Boehr-inger Mannheim GmbH, Mannheim, Federal Republic of
Germany. The cDNAs were methylated with EcoRI
meth-4165
0022-538X/89/104165-07$02.00/0
Copyright C) 1989, American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
Kb
94r 61
.4I
A
Kb M i 1 2 3 4
2:3 1
1.i
:1it
M 1 2 3 4
23 .*a
0
0
acids from liver or tumor tissues containing actively replicating
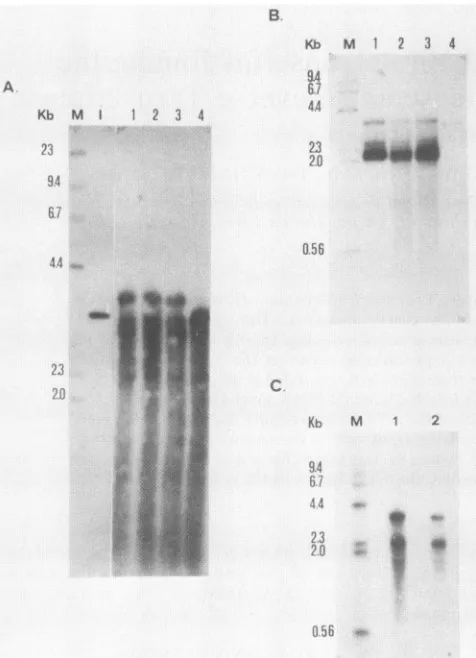
HBV. (A)Southern blotanalysis of totalDNAfromoneliver tissue sample. Lanes: M, HindIll fragments of lambda DNA; I, linear, 3.2-kbp HBV genome. Ten micrograms of tissue DNA was
undi-gested(lane 1)ordigested withHindIII(lane 2), EcoRI (lane 3),or
BamHI (lane 4) and then subjected to electrophoresis and
trans-ferred andhybridized with HBV-specific probe. (B) Northern blot analysis of poly(A)+ HBV RNA. Lane M, Hindlll fragments of lambdaDNA.Poly(A)+RNAsfrom the first HCC (lane 1), second HCC (lane 2), and nontumor liver (lane 3) were glyoxalated, fractionated by gelelectrophoresis, transferred, and hybridized with full-length HBV probe. Lane 4,Denatured 3.2-kbp HBVgenome.
(C) Hybridization of poly(A)+RNAfromasample ofnontumorliver tissue (lane 1) and HCC tissue (lane 2) with an HBcAg-specific probe. Lane M contains the size markers. Results offractionated poly(A)+ RNAs from two independent tissue samples hybridized withtheBglIl-BglII fragmentareshown (lanes1 and 2).
ylase, ligated with EcoRI linkers, and then inserted into a
Agtll vector by using a cloning kit purchased from
Amer-M 2 sham International plc, Amersham, United Kingdom. The library contained about 200,000 clones, and 90% were re-combinants. Forscreening, 10,000 plaqueswere platedin a
98-mm-diameter petri dish, transferredto nitrocellulose
pa-pers, and then hybridized with core-specific probes
(BglII-BgIII
fragment; see Fig. 2A).Subcloning and sequencing. A positive clone (CRC12), which contained a 0.75-kbp insert and was found not to
hybridize with the BglII-EcoRI probe but with the
EcoRI-RsaIprobe, waspickedup for sequencing (see Fig. 2A).The
0.75-kbp
insert was subcloned into apGEM7
vector(PromegaBiotec,Madison, Wis.),andsequencingwasdone
bythechain-terminationmethod with the supplied primers.
A
~3100_ tAn 3.6 kb RNA -CAtv An 2.6 kb RNA _1255 An 2.2 kb RNA
PbI
[INS,
13I
s ]JEceol[cIRZ.AsaI
-Ft:rI:
HBV
genome
1
__1xL
(tandem)FIG. 2. Mapping ofthe novel viraltranscript. (A)Organization ofthe HBV genome. Apartial dimer of theHBV genomeisshown, andthefour ORFsareindicated.C,ORFforHBcAg;Pol, ORF for theputativepolymerase of HBV; PreSl,S2, and S, HBsAgs. TheX
ORF is also shown. The first nucleotideof the initiation codon of HBcAg is designated 1. Three previously identified transcripts of
HBVareshown. The3.6-kbtranscript is thepregenomic RNA;the 2.6-kb species is the pre-Sl RNA; and the 2.2-kb species is the
pre-S2+S mRNA(11, 33). TerminalrepeatsofpregenomicRNAs areshown( )and arelabeledR. The 5'endsofthethree RNAs are shown (11). The probes used for subgenomic mapping are
indicated below the HBVgenome. (B)Mapping ofthe novel viral transcript with subgenomic probes. Lane M,HindIII fragments of lambdaDNA.Thepoly(A)+RNAsweresubjectedtoNorthernblot analysis,and thefilterwashybridized with wholeHBVprobe(lane
1).After stripping asdescribed previously (3), the samefilter was thenhybridizedwiththeBglII-BglIIfragment (lane 2). The results of hybridization with the BglII-EcoRIfragment (lane 3)and with the
EcoRI-RsaIfragment (lane 4)areshown. C
B
I
x .I .I
-1
--LN1
Kb M I 2 3 4
Oa
44
;~~~~4w
i -.*r;1:1...;~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.
.1
z
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.63.301.82.411.2] [image:2.612.118.562.431.723.2]A.
A_
NF7
_
S_I
-WP"a
G
T
cc
GG
T
Tc TT
TCCG
AC
C
G
rAG G
GGA
G
TG
GG
TGr
B.
GCACTCAGACGAAGA--- ---TCTCAPTGGGOAACA cDNA
GCACTCAGACGACGA --TCTCAATGGGGAGCA HBV(adr)
CAAGC- TTGTTATrAGT----TTCTA
C.
1 *
I
I
--"
M
PO'
C f~~~~~~eSIS1S~~~~~~~ Xim~r HBV genome
(tandem)
I6 a4541 1564
An 2.2 kb RNA
[image:3.612.134.480.75.506.2](vspliced) 46480
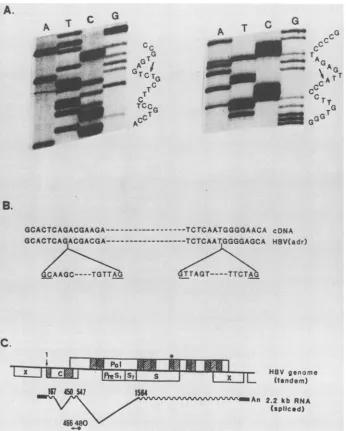
FIG. 3. Structure of thedoubly spliced HBV transcript deduced from cDNA clones. (A) Autoradiograms of sequences oftwosplicing junctions as revealed by the cDNA clone. The junction inside the HBcAg is shownattheleft; that between HBcAg and HBsAg isattheright. (B)Alignment of nucleotide sequences with that of HBV (subtype adr) (5). The intronsareindicated, and theconsensussequencesforsplicing
junctions are underlined. (C) The splicedRNAcontained theconserved andfunctional portions of the pol ORF.The twosplicing sitesare
marked. The conservedregionsof HBV ORFareindicatedbythe hatched boxes. Areas of 30% amino acidhomology( )andof 70to80%o
homology ( ) amongthehepadnaviruses (adapted from reference 25)areshown. Theregion with strong homologytoretroviralreverse
transcriptaseis marked withanasterix. Theoligonucleotide used forprimer extension analysis is also shown.
To complete the sequencing process, an oligonucleotide
complementary to bases 466to480ofthe HBV genome(5'
AGTTCTTCTTCTAGG 3') was synthesized (model 381A;
Applied Biosystem, Foster City, Calif.) and utilized as an
internalprimer.
Primer extension. The oligonucleotide described above
was labeled at its 5' end by using polynucleotide kinase
(Bethesda Research Laboratories) and
[32P]ATP
(5,000 Ci/mmol). Labeled oligonucleotide was isolated by passage
throughaG-50column. About 100,000cpmof
oligonucleo-tide was mixed with 1 ,ug of poly(A)+ liver RNA in a
hybridization solution {40 mM PIPES
[piperazine-N,N'-bis(2-ethanesulfonic acid)] [pH 6.4], 1 mMEDTA, 400mM
NaCl, and 60% formamide}, denatured at
80°C
for 10 min,and annealed at 42°Cfor 1 h (33). Total nucleic acidswere
ethanol precipitated. The pellet was dissolved in 10
,ll
ofreaction buffer (100 mM Tris hydrochloride [pH 8.3], 140
mM KCl, 8 mM MgCl2, 2.5 mM
dithiothreitol,
5 mMdeoxynucleoside triphosphates)
without and with dactino-mycin (1.5 ,ug/ml), and then 10 U ofreverse transcriptase(Seikagaku America, Inc., St.
Petersberg, Fla.)
was addedand themixturewasincubatedat
42°C
for 2 h. Theelongated
products were then separatedin a7 M urea-5%
polyacryl-amidegel.
RESULTS
Identification of a novel viral transcript in liver tissues. Three
specimens,
twofroman HCC and onefromadjacent
on November 10, 2019 by guest
http://jvi.asm.org/
0.561
M 1 2
8Ag'
'r.!..
.-U-",.4
:~~~~~~~~~~~~~~~~~~-;
-28?
[image:4.612.112.260.75.359.2]0.125,
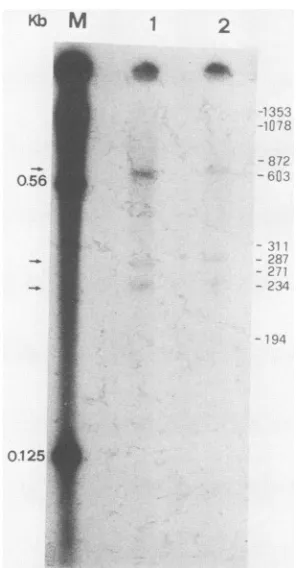
FIG. 4. Mappingof the 5' end ofsplicedRNAby primer exten-sion. An oligonucleotide complementary toexon2 waslabeled at the 5' end, annealed with poly(A)+ RNA from the liver, and
elongated byreverse transcriptase (33)withorwithout dactinomy-cin. Productswere analyzed by electrophoresisina7 M urea-5%
polyacrylamide gel. Lanes 1 and 2 show the products withoutor
with the presence ofdactinomycin, respectively.Lane M,HindlIl
fragmentsoflambda DNA. The positionsofHaeIII-digested
frag-ments of the PX174 replicative form are shown at the right.
Reactionproductsareindicatedbythearrowsat the left of thegel.
livertissues,were surgicallyremoved fromHBsAgcarriers. All three tissue samples were found to have only actively replicating HBV DNA (Fig. 1A), without integrated viral DNA. Northern blot analyses of poly(A)+ RNAs from the three samplesareshowninFig. 1. Usingwhole HBV DNA
as a probe, three discrete RNA species 3.6, 2.6, and 2.2 kilobases (kb) in length, respectively, were detected (Fig. 1B), corresponding to the pregenomic RNA,
pre-Sl
RNA,and pre-S2+S RNA (Fig. 2A) previously characterized in
human liver (33) and HBV DNA-transfected hepatoma cell lines (11, 28).
The threespecimenswerefurtherexamined todetermine whetherthereisasplicedRNAwhich deletes the
hypervari-able region ofpre-S sequences, thus potentially servingas
themRNAforpolprotein. Since thisputativeRNAspecies
shouldtheoreticallycontainsequencesofnucleocapsid
pro-tein(34), anHBcAg gene-specific BglII-BglII fragment (Fig.
2A)wasused as aprobe for Northernanalysis.As shown in
Fig. 1C,two RNA speciesweredetected in twoindependent
tissue specimens, a 3.6-kb pregenomic RNA and an
addi-tional RNA approximately 2.2 kb in length. Although the
latterRNAhasapproximatelythesamesizeasthepre-S2+ S RNA, itisanovel RNAspecies sincepre-S2+S RNAdoes
notcontainHBcAgsequences(Fig. 2A).Thesamefilterwas
also hybridized with a glyceraldehyde phosphate
dehydro-genase cDNAprobe (32) and showed no RNAdegradation (data notshown).
determine the structure of this novel RNAspecies, hybrid-ization experiments were done with different subgenomic fragments of HBV (Fig. 2A). In Fig. 2B, lane 1, all three RNAs were detected with thefull-length HBVprobe. When the HBcAggene-specific BglII-BglII probe was used, only 3.6- and 2.2-kb species were detected (lane 2). The probe from the next region (BglII-EcoRI) detected two speciesas
expected, the 3.6-kb pregenomic and the 2.6-kb
pre-Sl
RNAs(lane 3). Another probe further downstream (EcoRI-RsaI), containing sequences of the HBsAg gene, hybridized with all three bands (lane 4). These results suggest that the 2.2-kb RNA contains sequences of the C and S regions but has an internal deletion in the pre-S region.
cDNAcloning and sequencing indicated that thetranscript is adoubly spliced RNA. Because the amounts of available RNAswere toolimitedtodoS1 nuclease mapping to further define the structure of this RNA, we performed cDNA cloning of poly(A)+ RNAs from these liver tissues and screened the clones withBgII-BgIII probes. Positive clones werefurtherhybridizedwithaBglII-EcoRI probe, and those which werenegative at thesecond hybridizationwere con-sidered candidate cDNA clones derived from this novel 2.2-kb RNA. By this approach, several cDNA clones were
picked up. The sequence of one representative clone
(CRC12) was obtained. After comparison with HBV
se-quences, this clone was found to start from the 5' end of pregenomic RNA and to skip the next 282 nucleotides
correspondingtothe middle ofHBcAg (Fig. 3A, left). There
isaseconddeletion of 1016nucleotidescorrespondingtothe
regionbetween the endofHBcAgand thebeginningof the S
region (Fig. 3A, right). The deleted sequences are bracketed
atthe 5' endby GCorGT andatthe 3' endby AG (Fig. 3B), theconsensusboundarysequencesof the introns(19, 24). In
aprevioustransfectionexperimentdone withCOS cells, the
second acceptor site definedbythiscDNAwasaccidentally
discovered (1). Thus, we concluded that the cDNA clone
represented adoubly spliced HBV transcript (Fig. 3C).
Spliced RNA is coterminal with pregenomic RNA. The
approximate 5' and 3' ends of this transcript were
subse-quentlydetermined. Since there isonlyonepolyadenylation
signal in the HBV genome (33), all poly(A)+ transcripts of
HBV were 3'-end coterminal. Subtraction of two introns from the pregenomicRNA wouldgive a size equivalent to that of the spliced2.2-kb RNA (Fig. 3C). Thus, it is likely that the spliced transcript is coterminal with pregenomic RNA. To test this possibility, an oligonucleotide comple-mentaryto nucleotides466 to480ofthe HBV genome was used forprimer extension. Usingpoly(A)+ RNA from these humantissuesasthe template, three primer-extended
prod-uctswereobtained(Fig.4).The product, about 600 bases in
length, should represent the extended primer on unspliced
pregenomicRNA. The other two products, about 300 bases
inlength, probablyrepresented the extended primer on the
spliced2.2-kb RNA. The presenceof two bands may be due
tomicroheterogeneityatthe 5' ends of the HBV transcripts.
This result suggests that the 2.2-kb spliced RNA is cotermi-nal with the 3.6-kb pregenomic RNA.
Analyses oftheORFs of the spliced RNA and the
conserva-tion of sequences around splicing junctions. The coding
ca-pacity of this splicedRNAwas analyzed. The first splicing
eventtookplace in the ORF of the HBcAg gene. Since 282 bases were removed, the reading frame of HBcAg was
preserved.The secondsplicing event removed the
termina-tion codon ofHBcAg and the whole hypervariable regions of
I
on November 10, 2019 by guest
http://jvi.asm.org/
A
10 20
A T
AACTTTTCACCTCTOCCTAA
30 40 50 60
-.
TCATCTCATAGCATGTCCTACTOTTCAAGCCTCCAAGC
LeuPheHisLeuCysLeuI 1e1leSerCymS.rCyuProThrVal0lnAlaSerLysLeu
70 80 90 100 110 120
TGTGCCTTGGCTGGCTTTOGOGCATGGACATTGACCCOTATMAGAATTTOGAGCTTCTO
CysLeuGlyTrpLeuTrp0ly AspIleAspProTyrLysGluPheGlyAlaS@rVal IlUcAg-.
130 140 150
*--:
160 170
:
190 200 210 220 230 240
CCGCCTCTGCGCTGTATCGGAGOCCTTAGAVTCTCCGAACATTG1TCACCTCACCATA
AlaSerAlaLeuTyrArgOluAlaLeuGluSerProGluHlaCysSerProUisUiNlTr
250 260 270 280 290 300
S1 : :
^-or-'o-*-^->^^--*-*-*
> ^n-LyuLysA.nSerLouAla.rGlanThrLys
AlaLeuArgAraArgalvrArS8rProArfArgArgThrProS@rProArIAEAtrArAg
310 320 330 340 3350 360
GGTCTCAAkTCGCCGCGTCGCA0AAGATCTCAATCTC0GGGATCTCZAAT0A0CTACAC
ValSerIleAlaAlaSerOlnLysl*l-SrIleS*rGlyl0 8e*
Iy;rSrTyrThr
Se rG
lnSerProAreArgArgArgS-rGInS-rArgOly8orA
p abrPro380 390 400 410
CCGTGTOTCTTOOCCAAAATTCOCAOTCCCAAATCTCCAGTCACTCACCAACCTOTOTC
ArgVa1S.rTrpProLyaPbeAasValProAsa1eu08rL.Th3raLeuL@uSr
ValCysL.uGlyGlnAsAaSrOlSn.rOlnlle8erSerKiSSIrProP1rCyCCySPZo
B
1st S.D. 1st S.A. 2nd S.D. 2nd S.A.
adr ac
tag/GCAAGC--TGTTAG/a09-9--otoaat/aTTAGT--TTCTAG/W99ag
adr4
&Ctcag/GCAAGC--TGTTAG/a-ca-a--ctoaat/GTTAGT--TTCTAG/g
M agadw actc-g/GCAAGC--TGTTA:/acg
ga--ctcaat/GTTAGT--TTCTAG/9gga
tayw actcag/GCAAGC--TGTTAG/acgag--ctcaat/GTTAGT--TTCTAG/mgaa
[image:5.612.150.467.62.610.2]adyw actcag/GCAAGC--TGTTAG/acgaga.--ctoaat/GTTAGT--TTCTAG/ggg9aa
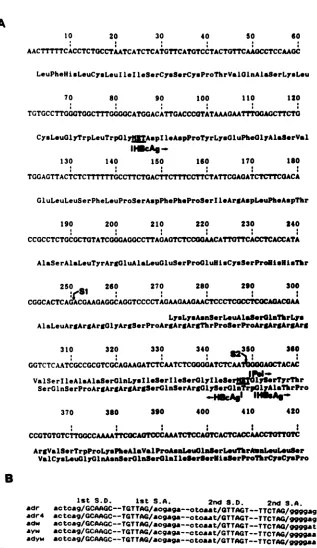
FIG. 5. Analyses of coding capacities of the spliced RNAs andsequenceconservation around the splicing sites. (A) The mapped5'end ofpregenomic RNAwasusedasthe first nucleotide (11). S1, The first splicing junction;S2,the second splicingjunction. The overlapping ORFs between shortened HBcAg andpol are shown. (B) Conservation of nucleotide sequences around the splicing sites among HBV
subtypes. Sequences from HBV subtypes adr (5), adr4 (20), adw (20), ayw(6),and adyw (21)arealigned with respecttothetwosplicing junctions. S.D., Splice donor site; S.A., spliceacceptorsites.
pre-Sl and pre-S2.Asaresult, the shortened ORF ofHBcAg
is followed by a Trp and then is contiguous with that of
HBsAg. The N terminus of the putative polymerase gene
was removed, and a new initiation codon was generated,
which is in phase with the truncated ORFs ofthepolymerase
gene (Fig. 5A). The truncated pol includes the most
con-servedregionand theregionhomologoustothat ofretroviral
reversetranscriptase (Fig. 3C).
The sequences around the two splicing junctions were
found to be remarkably conserved amongthe various
sub-types of HBV (Fig. 5B) (except for one nucleotide). The
homology could be extended somewhat to that of
wood-1o8
370 420
on November 10, 2019 by guest
http://jvi.asm.org/
second
splicing
acceptor site(data
notshown).
It will beinteresting
todeterminewhetheranimalhepadnaviruses
also express similarspliced transcripts.
DISCUSSION
The
transcription
ofHBVboth invivo and in transfected cell lines has been studiedextensively,
and mRNAs formajor
viralprotein
products
havebeenidentified(11,
28, 33).
A2.2-kbviralRNA is showntobe thetranslation
template
for
hepatitis
B surfaceproteins,
and a 3.6-kb RNA is thetemplate
for coreprotein (11, 33). However,
whether the3.6-kb RNA also serves as the mRNA for viral reverse
transcriptase
is not yet settled.Considering
thespecial
genetic organization
ofHBV,
welooked forthe presence ofspliced
RNAas analternative RNAtemplate.
IntwoHBV-infected human liver
specimens,
evidence ofa2.2-kbdoubly
spliced
viraltranscript
was obtained. Infact,
similar RNAspecies
wererecently
also detected in infected humantis-sues
(26).
The resultsclearly
indicate theexistence ofviraltranscripts
of HBV not yet discovered and the need toexplore
thecomplex
viral geneexpression
during persistent
infections.
The
biological
significance
ofthisspliced
RNAcurrently
isunknown.Itshould be
pointed
outthat,
exceptin the reportofSu et al.
(26),
the RNA has not been demonstrated inprevious
studies. This factmayreflectthe lack ofanessen-tial role for this RNA in the viral life
cycle.
However,
thefailure could be due to
(i)
anobscuring
effectby
thepre-S2+S
mRNA thatcomigrates
with thespliced
mRNAspecies
or(ii)
adifferent levelofsplicing efficiency,
since ithas been well documented that retroviruses can
regulate
viralgene
products by
controlling
the extent ofsplicing (8,
14).
Inaddition,
the conservation ofsequences around thesplicing
donor andacceptorsitesamongHBVsubtypes
(Fig.
5B)
suggeststhat RNAsplicing
may havefunctional role. For all the retrovirusescharacterized,
theunspliced
ge-nomicRNA is considered tobethe
template
for translationofreverse
transcriptase
(10, 29). However,
inretroviruses,
the ORF for reverse
transcriptase
isseparated
from that ofthe
envelope
proteins
which containhighly
variableregions
(29).
In contrast, the two ORFs in HBV arecompletely
overlapping.
Thedoubly spliced
form ofpregenomic
RNAidentified inour
study
mayprovide
amechanismtoremovethis
hypervariable region
so that thepolymerase
remainsfunctional.In
addition,
theregion
within the HBVpolORFwhich is
homologous
to theretroviral reverse transcriptaseis retained in this
spliced
RNA(Fig.
3C).Therefore,
it ispossible
thatthisspliced
RNAisthetranslationtemplate
forreverse
transcriptase
ofHBV. Inaplant
virus,
cauliflowermosaic
virus,
thatalsoreplicates through
reversetranscrip-tion
(9),
thetemplate
for translation ofreversetranscriptase has been shown to beasubgenomic
RNAof about 2.5 kb,whichcouldbea
spliced
RNA(22). This result is consistentwith
findings
presented
inourreport. Tofurthersupportit,a
putative
reversetranscriptase
identified in virions byspecific
antibody
has been shown not to contain the Nterminus ofpol
ORF,
thehypervariable
region (13). Theprimary
translation product of retroviral reversetranscriptase
isagag-pol fusionprotein
which isfunctionallyinactive
(10, 29).
Afterbeing
packaged into virionparticles,the fusion
protein
is cleaved byaproteaselocated eitheratthe C terminusof gagor atthe N terminusofpoltorelease
theactivereverse
transcriptase
(16, 29). Asimilarprotease-cleavage
mechanismmayalso beimportant
for activation ofprotease has beenproposedtobe locatedatthe Nterminus of thenucleocapsid protein (HBcAg)
(17).
The removalofaportionof
HBcAg by
RNA splicingcouldbring
the domainof theputativeprotease closerto thehypothetical
cleavage
site ofreversetranscriptase. Thus,thismaybeacrucialstep
inactivatingthe distantly spaced protease.
Inthisspliced RNA,thetwo shortened ORFs of
HBcAg
and pol are contiguous but out of phase (Fig. 5A). This
organization is reminiscent of that of theretroviral gene for
the
gag-pol
fusion protein (10, 29). The genetic structureallows the translation of this spliced RNA viaa ribosomal frame-shifting mechanism (34) to generate a
protease-pol
fusionprotein. However,itis noted that the second
splicing
eventcreatesa new initiation codon in shortenedpol
ORF. Therefore, the synthesis of viral reverse transcriptaseby
internal initiationfrom thesplicing site isequally possible,as
isdocumentedbythemechanism for translation of the duck
HBVpolgene (2, 23). Further studiesare needed to show
which modelactually works intranslating thisdoublyspliced transcript of HBV.
Although arguments that thissplicedRNA is the
template
for HBV reverse transcriptase have beenpresented,
previ-ous genetic studies on the duck HBVpol ORF identified severalfeatures whicharerelevanttopolexpression butarenot present in the spliced RNA. For example, the spliced RNAdoesnotcontain the first AUG in thepolORF that is shown to be important for polymerase expression (2,
23).
Certainly it is possible to evoke a mechanismdefending such
adefect(e.g., that the sequence is required for removal of the intron), but until further experimental data become available, the functional assignment of this spliced RNA shouldbe considered hypothetical.
Another consequence of this RNA splicing event is the possible generation of an HBcAg-HBsAg fusion protein, since the two ORFs are fused in this spliced RNA. Such a
protein remains tobe identified.
Finally, since the spliced RNA is coterminal with the pregenome, all thecis-acting sequences required for replica-tion are probably present. However, this RNA species is unlikely to be packaged into virus particles; otherwise, its reverse transcription might interfere with viral replication. Thus, the sequences missing in this spliced mRNA, analo-gous tothose ofretroviruses, may be important for
packag-ing (15).
ACKNOWLEDGMENTS
Wethank Michael M. C. Lai for critical reading of the manuscript and helpful comments, W. S. Robinson for providing the HBV
plasmid, S. J. Tu for invaluable technical assistance, and M. S. Chuang for secretarial assistance.
The workwassupported bygrantsfrom theInstitute of Biomed-icalSciences,AcademiaSinica,the National Science Council, and
theDepartment ofHealth, Taiwan, Republic ofChina.
LITERATURE CITED
1. Cattaneo, R., H. Will, G. Darai, E. Pfaff, and H. Schaller. 1983.
Detectionof an elementofthe SV40late promotor in vectors
usedforexpression studies.EMBO J. 2:511-515.
2. Chang, L.-J., P. Pryciak, D. Ganem, and H. E. Varmus. 1988.
Biosynthesisofthe reversetranscriptaseof hepatitis B viruses
involves de novo translational initiationnot ribosomal
frame-shifting. Nature(London)337:364-367.
3. Chen,P.J.,G.Kalpana, J. Goldberg, W. S. Mason, B. Werner,
J.L.Gerin,andJ. M. Taylor. 1986. Structureand replication of
hepatitis delta virus.Proc. Natl.Acad.Sci. USA83:8774-8778.
4. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W. J.
on November 10, 2019 by guest
http://jvi.asm.org/
Rutter. 1979. Isolation of biologically active ribonucleic acid
from sourcesenrichedin ribonuclease. Biochemistry
18:5294-5300.
5. Fujiyama, A., A. Miyanohara, C. Nozaki, T. Yoneyama, N. Ohtomo, and K. Matsubara. 1983. Cloningandstructrual
anal-ysis of hepatitisBvirusDNAs, subtype adr. Nucleic Acids Res. 11:4601-4616.
6. Galibert, F., E. Mandart, F. Fitoussi, P. Tiollais, and P. Char-nay. 1979.Nucleotidesequence of the hepatitis B virus genome
(subtypeayw)clonedin E. coli. Nature (London) 280:646-650. 7. Gubler, U., and B. J. Hoffman. 1983. Asimpleand veryefficient
method forgeneratingcDNAlibrary. Gene 25:263-270.
8. Gutman, D., and C. J. Goldenberg. 1988.Virus-specificsplicing
inhibitors in extracts from cells infected with HIV-1. Science
241:1492-1495.
9. Hull, R., and S. N. Covey. 1986. Genome organization and
expression of reverse transcribing elements: variation and a
theme. J. Gen. Virol. 67:1751-1758.
10. Jacks, T., and H. E. Varmus. 1985. Expression of the Rous sarcoma virus pol gene by ribosomal frameshifting. Science 85:1237-1242.
11. Katsuyuki, Y., Y. Shirakata, K. Kobayashi, and K. Koike. 1987.
Stable expression and replication of hepatitisBvirusgenome in an integrated state in ahuman hepatoma cell line transfected with the cloned viral DNA. Proc. Natl. Acad. Sci. USA
84:2678-2682.
12. Lai, M. Y., D. S. Chen, P. J. Chen, S. C. Lee, J. C.Sheu, G. T. Huang, T. C. Wei, C. S. Lee, S. C. Yu, H. C. Hsu, and J. L. Sung. 1988. Status ofhepatitis B virusDNAin hepatocellular carcinoma:astudy baseduponpairedtumorandnontumorliver tissues. J. Med.Virol. 25:249-258.
13. Mack, D. H., W. Bloch, N. Nath, and J. J. Sninsky. 1988.
HepatitisBvirusparticlescontainapolypeptide encoded by the largestopenreading frame: aputativereversetranscriptase. J.
Virol. 62:4786-4790.
14. Malim, M. H., J. Hauber, R. Fenrick, and B. R. Cullen. 1988.
Immunodeficiency virus rev trans-activator modulates the
expression of the viral regulatory genes. Nature (London) 335:181-183.
15. Mann, R. S., R. C. Mulligan, and D. Baltimore.1983. Construc-tion ofa retroviral packaging mutantand its use to generate
helper-free defective retrovirus. Cell33:153-159.
16. McClure, M. A., M. S. Johnson, and R. F. Doolittle. 1987.
Relocation ofaprotease-likegene segmentbetweentwo
retro-viruses. Proc. Natl. Acad. Sci. USA84:2673-2677.
17. Miller, R. H. 1987.Proteolytic self-cleavage ofhepatitisBvirus
core protein may generate serume antigen. Science 236:722-725.
18. Miller,R.H.,P. L.Marion,and W.S. Robinson. 1984.Hepatitis
Bviral DNA-RNAhybrid molecules inparticles from infected liverareconvertedtoviralDNA moleculesduringan
endoge-nousDNApolymerase reaction. Virology139:64-72.
19. Myerowitz, R. 1988. Splice junction mutation in some
Ash-kenaziJewswithTay-Sachsdisease: evidenceagainst asingle
defect with this ethnic group. Proc. Natl. Acad. Sci. USA
85:3955-3959.
20. Ono, Y., H.Onda, R. Sasada, K. Igarashi, Y. Sugino, and K. Nishioka. 1983. The complete nucleotide sequences of the
cloned hepatitis B virus DNAs subtype adrand adw. Nucleic AcidsRes. 11:1747-1757.
21. Pasek, M., T. Goto, W. Gilbert, B. Zink, H.Schaller, P. McKay, G. Leadbetter, and K. Murray. 1979. Hepatitis B virus genes
andtheir expressionin E.coli. Nature(London) 282:575-580.
22. Plant, A. L., S. N.Covey, and D. Grierson.1985. Detectionofa subgenomic mRNA for gene V, the putative reverse
tran-scriptasegeneof cauliflower mosaic virus. Nucleic AcidsRes. 23:8305-8321.
23. Schlicht, H.-J., G. Radziwill, and H. Schaller. 1989. Synthesis and encapsidation of duck hepatitis B virus reverse
tran-scriptase do not require formation of core-polymerase fusion proteins. Cell 56:85-92.
24. Sharp, P. A. 1981. Speculation on RNA splicing. Cell 23:
643-645.
25. Sprengel, R., C. Kuhn, H. Will, and H. Schaller.1985. Compar-isonsequenceanalysis of duckand humanhepatitisBvirus.J. Med. Virol. 15:323-328.
26. Su, T.-S., W.-Y. Lui, L.-H. Lin, S.-H. Han, and F.-K. Peng. 1989.Analysisof hepatitisBvirustranscripts in infectedhuman
livers. Hepatology9:180-185.
27. Summers, J., and W. S. Mason. 1982.Replicationofthegenome
ofahepatitis B-like virus by reversetranscription ofanRNA
intermediate. Cell29:403-415.
28. Sureau,C., J. L. Romet-Lemonne, J. I. Mullins, and M. Essex. 1986.Production ofhepatitisB virusbyadifferentiated human
hepatoma cellline after transfection withcloned circularHBV DNA.Cell 47:37-47.
29. Swanstrom, R., and H. E. Varmus. 1985. Replication of
retro-viruses, p.369-513.InR. Weiss,N. Teich,H. Varmus,andJ.
Coffin (ed.), RNA tumorviruses, vol. 1. Cold SpringHarbor Laboratory,ColdSpring Harbor,N.Y.
30. Tiollais, P., C. Pourcel, and A. Dejean. 1985. The hepatitisB
virus. Nature(London) 317:489-495.
31. Toh, H.,H. Hayashida,and H.Miyata. 1983. Sequence
homol-ogybetweenretroviralreversetranscriptase andputative
poly-merase of hepatitis B and cauliflower mosaic virus. Nature
(London) 305:827-829.
32. Tso,J.Y., X. H.Su,T. H.Kao, K. S.Reece, and R. Wu.1985.
Isolation and characterization ofratandhuman glyceraldehyde-3-phosphate dehydrogenase cDNAs: genomic complexity and molecular evolution of the gene. Nucleic Acids Res. 13:2485-2502.
33. Will, H., W. Reiser, T. Weimer, E. Pfaff, M. Buscher, R.
Sprengel,R.Cattaneo,and H. Schaller. 1987.Replication strat-egyofhumanhepatitisB virus.J. Virol. 61:904-911.
34. Will, H., J. Salfield, E.Pfaff,C. Manso,L.Theilmann, and H. Schaller. 1986. Putative reverse transcriptase intermediates of human hepatitis B virus inprimary liver carcinomas. Science 231:594-596.