0022-538X/89/051897-10$02.00/0
Copyright © 1989, American Societyfor Microbiology
Control of mRNA Stability
by the Virion Host Shutoff Function
of
Herpes
Simplex
Virus
ASHA A. OROSKAR ANDG. SULLIVAN READ*
Department of Microbiology and Program in Molecular Biology, Loyola University of Chicago, Stritch School
of
Medicine,
2160
Soluth
FirstAvenue,Maywood,
Illinois 60153Received 3 October 1988/Accepted 25 January 1989
vhslisamutantofherpessimplex virustype1thatis defective in the virion hostshutoff function responsible forthedegradation ofcellular mRNAs and the concomitant shutoffofhostprotein synthesis.In thisstudy,the effect ofthe vhsl mutationonthe metabolismof viralmRNAswasexamined by measuringthe half-lives and
patternsof accumulation of10 different viralmRNAsrepresenting all kinetic classes. The vhsl mutationhad theeffect ofdramatically lengtheningthecytoplasmic half-lives ofall10mRNAs. Inwild-typevirusinfections, the 10 mRNAshad similar half-lives,suggesting that little,ifany,targetmRNAselectivitywasexhibitedby the
vhs function. The vhsl mutation caused overaccumulation of a number of mRNAs. The effect was most
dramatic for thealpha (immediate-early) mRNA for ICP27and the beta(early) mRNAsencoding thymidine kinase, ICP8, andDNApolymerase. Whereasinwild-typeinfections these mRNAs increasedtopeak levels and subsequently declined in abundance, in vhsl infections they continued to accumulate until late times. A significant but less dramatic overaccumulation was observed for several beta-gamma (delayed-early) and
gamma (late) mRNAs. The results suggest that the vhs protein plays an important role indetermining the half-lives of viral mRNAs belonging to all kinetic classes and in so doing is important in the normal
downregulation atlate times ofalphaand betageneexpression.
Incells infected with herpes simplex virustype1(HSV-1), viral gene expression is regulated in a complex cascade
fashion (22). For most viral gene products, the rate of synthesisofaproteinisdirectly proportionaltotheamount of thecorresponding mRNAin thecytoplasm (14, 27). Since mRNAlevelsarenecessarily governed bytherateofmRNA degradation as well as the rate of synthesis, factors that control either process will affect gene expression. Many
studies of the control oftranscriptioninHSV-1-infected cells have been reported; however, to date few have addressed the factors that govern mRNA stability. The experiments reportedin thispaper concerntheregulation of viralmRNA half-livesby the virion host shutoff (vhs) function of HSV-1. One of the earliesteventsduring lyticHSV-1infections is the shutoff ofmost cellular protein synthesis (3-6, 11-13, 16-18, 20, 23, 28-30). This shutoff is initially caused by a
structural component of the infecting virion which causes
disaggregationof cellularpolyribosomes (29, 30) and degra-dation of host mRNAs (24). Mutant virus containing alter-ations in the vhs gene produce virions that are unable to
suppress host protein synthesisordegrade cellular mRNAs
(20, 28). Recently, the mutation carried by themutant vhsl hasbeenmappedtoanopen readingframe abletoencodea
protein with a predicted molecular mass of 55 kilodaltons (13, 15). However, the vhs protein has yet to be identified within virionsor infected cells.
Thefirst demonstration that the vhsl mutation affects the half-lives of viral as well as host mRNAs was provided by
the observation that, following reversal ofacycloheximide
blockpresent fromthe timeof infection, alpha (immediate-early) viral mRNAs decayed much more rapidly in cells
infected with wild-type, HSV-1 than in cells infected with vhsl (18). Subsequently, it was shown thatifvhsl-infected
cells that were already expressing alpha mRNAs were
su-perinfected with wild-typevirus in the presence of
dactino-* Correspondingauthor.
mycin, acomponentof the superinfecting wild-type virions destabilized the alphamRNAsencoded by vhsl (12).
Several studies havesuggestedthat thevhslgeneproduct
may affect the stabilities of beta (early), beta-gamma (de-layed early), and gamma(late) viral mRNAs in addition to alphaand host mRNAs. First, the vhsl mutation has been known forsome timeto affecttheaccumulation of beta and
gamma viral polypeptides within the infected cell (20). Second,atlate times after vhslinfection,thereareincreased
amountsofsomebeta andgammamRNAsinthecytoplasm
(28). Third, Kwong and Frenkel added dactinomycin to infected cultures and then pulse-labeled the cells with [35S]methionineeitherimmediatelyorafteranadditional 6 h
in thepresence ofdactinomycin (12). When they compared the proteins being synthesized before and after the 6 h incubation in thepresence ofdactinomycin, theyfound that
manyviralproteins continuedtobetranslatedlongerincells infected with vhsl than in cells infected withwild-typevirus. While the above dataareconsistent with thehypothesisthat the vhsgeneproductaffects thephysical stabilityof beta and
gamma mRNAs, they are also consistent with other
possi-bilities.
The presentstudy wasundertaken toevaluate the role of the vhs gene product in regulating the half-lives of viral mRNAs belongingto different kinetic classes andto assess
the importance of controls of mRNAstability in the overall scheme ofgeneregulation in HSV-1-infected cells. To this end, the half-lives and patterns ofaccumulation of 10 dif-ferent viral mRNAs were determined incells infected with
wild-type virus or vhsl. The results indicate that the vhs gene product significantly affects the stability of viral
mRNAs belonging to all kinetic classes. In so doing, it affects the levels and kinetics ofaccumulation ofanumber of viral mRNAs. Inparticular, the vhs proteinappearsto play
an important role, in conjunction with transcriptional
con-trols,in the normaldownregulationatlatetimes ofalphaand betageneexpression.
1897
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
Cells and virus. Vero and HeLa S3 cells were purchased
from the American Type Culture Collection and grown in Eagle minimum essential medium (MEM; GIBCO Labora-tories) supplemented with antibiotics and 10%(vol/vol) calf
serum. Stocks of wild-type HSV-1, strain KOS, and the
mutant vhsl were prepared from infected Vero cell
mono-layers asdescribed previously (20). The vhslmutantgrows
well at all temperatures from 34 to 39°C and exhibits a
defective virion host shutoff function at all temperatures (20). All experiments were performedat 34°C. In all
exper-iments, virus was allowed to adsorb for 1 h in MEM containing 1% (vol/vol) calf serum. The inocula were then
aspirated, and the cells were overlaid with fresh MEM containing 1% calfserum.
Isolation of cytoplasmicRNA. TotalcytoplasmicRNAwas
prepared from monolayer culturesof 1 x 107to2 x 107Vero
or HeLa cells as described previously (18). The cellswere
first scraped into ice-cold phosphate-buffered saline and collected by low-speed centrifugation. They werethen
sus-pended in 2.2 ml ofresuspension buffer (10 mM Tris [pH 7.9], 0.15 MNaCl, 1.5 mMMgCl2) andlysed byaddition of Nonidet P-40(Calbiochem-Behring)to0.5%. After 10minon
ice, the nuclei were pelleted by low-speed centrifugation,
and thesupernatantwasremovedtoafresh tube. The nuclei
were washed with 1.2 ml of resuspension buffer containing
0.65% Nonidet P-40, and the wash was added to the first supernatant. The combined cytoplasmic supernatants were
further clarified by centrifugation ina Sorvall SS34 rotorat 10,000rpmfor10minand then addedtoanequalvolume of
ureabuffercontaining10 mM Tris(pH 7.9),7 Murea,0.35 M NaCl, 10mMEDTA,and 1% sodiumdodecylsulfate(SDS). The mixture was extracted twice with phenol-chloroform (1:1) and twice with chloroform, and the RNA was precip-itated byaddition of 2.5 volumes of ethanol and storage of the sample at -20°C overnight. After precipitation from ethanolasecondtime, the RNAsamplesweredissolvedina
small volume of 10 mM Tris (pH 7.9)-i mM EDTA and stored frozen at -90°C.
Gel electrophoresis and Northern (RNA) blot transfer of RNAs. Samples (5 ,ug) of total cytoplasmic RNA were
denatured with glyoxal and electrophoresed through 1.2%
agarose gels cast in 10 mM sodium phosphate, pH 6.8, as
describedpreviously (21).The RNAs werethen electroblot-tedontoGeneScreenPlus membranes(DuPont NEN Prod-ucts)withatransblotapparatus (Bio-Rad Laboratories)and
a transfer buffer of12 mM Tris-6 mM sodium acetate-0.3 mM EDTA, pH 7.5. Following electrophoretic transfer of the RNA, the membranes were rinsed for 30 s in 50 mM
NaOHtoremovetheglyoxal and then neutralizedbya30-s
rinsein0.2MTris(pH 7.5)-i x SSC(0.15MNaCl,0.015 M sodium citrate). The filters were then air dried at room
temperature.
DNAprobes. DNAprobesforuseinNorthernanalysisor
pulse-chase experiments were derived from recombinant pBR325 plasmidsorM13phages containingasinserts
appro-priate fragmentsof the HSV-1genome. Theplasmids pSG28 and pSG87, which were used for Northern blot
hybridiza-tion, werekindly provided by Myron Levine and Rozanne Sandri-Goldin(8). pSG28 wasusedas aprobe todetect the mRNAs forICP4, ICPO, andICP27, whilepSG87 wasused
todetectthymidinekinase andglycoproteinH(gH)mRNAs. Bothplasmids were maintained in Escherichia coliHB101.
Theplasmid pXlrll contained a4.6-kilobase (kb) fragment
of Xenopus laevis rDNA inserted into the EcoRI site of
colicin El
(2).
The clonedfragment
contained most of the 28S rRNAcoding
region
and a smallportion
of the 18S rRNAregion. pXlrll
waskindly provided by
JeffDoering.
PlasmidDNAswereprepared
by
standardprotocols
(1).
The
probes
thatwereused forNorthernanalysis
ofICP8,
DNA
polymerase,
glycoprotein
B(gB),
ICP5,
and glycopro-tein C(gC)
mRNAs wereprepared
from recombinant M13phages
containing
asinsertsintragenic fragments
ofthe viral genes. Thesephages
werekindly
provided by
Steven Wein-heimerand have beendescribedpreviously
(31).
Thephages
contained thefollowing
inserts of HSV-1 DNA:ICP8,
a1.6-kb SalI-BamHI
fragment;
DNApolymerase,
a 0.6-kbBglII-EcoRI
fragment; gB,
a 0.6-kb PstI-SaIlfragment;
ICP5,
a0.6-kbBamHI-HindIIIfragment;
andgC,
a 1.6-kb EcoRI-BamHIfragment.
To prepare
probes
forNorthernblotanalysis,
the inserts wereexcised fromappropriate
plasmids
orM13replicative
forms. DNAfragments
wereseparated
by
agarosegel
elec-trophoresis,
and the desired inserts werepurified
fromlow-melting-temperature
agarose(21).
Theprobes
werethen labeled with 32Pby
nicktranslation withanicktranslation kit fromBethesda Research Laboratories(Rockville, Md.).
For
preparation
ofprobes
to be used in thepulse-chase
experiment,
we found it convenient to reclone the inserts fromrecombinantM13phages
intotheequivalent
sites ofthepolylinker
of theplasmid Bluescript
SK(Stratagene).
The recombinantphages
that were used for thepulse-chase
experiment
contained thefollowing
inserts: forICPO,
a 1.6-kbSstI-BamHIfragment;
forTK,
a1.1-kbBglII-HindIII
fragment
from the 3' tkdeletion mutantA3'-1.6(31).
Thesephage
were alsoprovided by
StevenWeinheimer.Hybridizations.
Membranescontaining
transferred RNAs werehybridized
tonick-translatedprobes by
theprotocol
of Graham and co-workers(9).
The filterswerefirstprehybrid-ized at
42°C
for 24 h inprehybridization
buffercontaining
50% formamide(Mallinkrodt, Inc.),
10% dextran sulfate(Pharmacia Inc.),
10%Denhardt solution(1),
Sx SSPE(1x
SSPEis 180 mM
NaCl,
10mMsodiumphosphate [pH
6.8],
1 mM
EDTA),
1% sodiumlauryl
sulfate,
and 250 ,ug of denatured salmon sperm DNA per ml.Following
prehybrid-ization,
theprobe
wasdenaturedby being placed
inboiling
waterfor10 minandwasthenaddedtotheprehybridization
buffer. The final
probe
concentrationwasgenerally
5 x107
to 10 x 107
cpm/ml. Hybridization
was then continued at42°C
for 24 h. At the end ofhybridization,
the blots werewashed twice for 45 mineach at room temperature in 2x
SSPE-0.4% SDS and then twice more for 15 min each in 0.1x SSPE at 600C. The still moist blots were then sealed inside freezer
bags
andexposed
to Kodak XAR-5 film forautoradiography.
For
quantitation
of mRNAlevels,
appropriate
exposures of theautoradiograms
were scanned with aGilford-Re-sponse
spectrophotometer
equipped
with agel-scanning
apparatus, and theareasunder the
peaks
wereintegrated.
InFig.
2through 5,
showing
the kinetics of mRNAaccumula-tion,
the amount of anygiven
mRNA isexpressed
inarbitrary
units. Thespecific
activities of theprobes
for different mRNAsvaried,
and exposures ofdifferentlength
were scanned for different mRNAs.Thus,
while the data allowedcomparison
of the levels ofaparticular
mRNA in mutant andwild-type
infections or the levels at different times afterinfection,
itwasnotpossible
todirectly
compare the level ofone typeofmRNA withthat of another.After
appropriate
exposures of theautoradiogram
had beenobtained,
theprobe
wasstripped
from the filtersby
boiling
the membranesfor 20to 30min in 10mMTris(pH
on November 10, 2019 by guest
http://jvi.asm.org/
7.9)-i mM EDTA-0.1% SDS. Complete stripping of the probe was monitored by autoradiography. Following strip-ping, the filters were ready for hybridization to another probe. This procedure allowed at least five rounds of hybrid-ization to be performed with each filter.
Measurement of mRNA half-lives with a dactinomycin chase. Vero cell monolayers were infected with 20 PFU of wild-type virus or vhsl per cell. At 6 h postinfection 5
pig
of dactinomycin per ml was added to the culture medium, and incubation was continued. Total cytoplasmic RNA was extracted at 45-min intervals, and the decay of specific mRNAs in the presence of dactinomycin was analyzed by Northern hybridization. To control for small variations in the amount of mRNA that was loaded onto different gel lanes, the results were normalized to the amount of 28S rRNAdetected by hybridization to the plasmid pXlrll.Measurement of mRNA half-lives with a pulse-chase proto-col. Measurement of mRNA half-lives by pulse-chase anal-ysis was performed as described by Pilder and co-workers (19) with minor modifications. HeLa cells infected with 20 PFU of wild-type virus per cell were pulse-labeled with MEM containing 200 ,uCi of [3H]uridine perml for 30 min beginning at 5.5 h postinfection. The label-containing me-dium was then removed, and thecellswerewashedand then overlaid with MEMcontaining 5 mMeachcold uridine and cytidine and 35 mMglucosamine. Total cytoplasmic RNAs were prepared at 45-min intervals and then subjected to exhaustivedigestionwithRQ1DNase(RNasefree;Promega Biotec), followed by phenol-chloroform extraction and eth-anol precipitation. The samples oflabeled RNA were then hybridized to specific fragments ofthe viral genome which had beenfixedto nitrocellulose filterdisks. For pulse-chase experiments, HeLa cells were used rather than Vero cells becausewe wereable toobtainamorecomplete chaseof the label with HeLa cells. Nevertheless, the patterns of viral gene expression are very similar in HSV-1-infected HeLa and Vero cells (datanot shown).
To fix the recombinant plasmid DNAs to nitrocellulose filters, the plasmids were linearized by restriction enzyme digestion, and 15-,ug portions were denatured and fixed to 25-mmnitrocellulose filterdisksasdescribedpreviously(1). Labeled RNA samples from approximately 5 x 106 cells werehybridized to the filter disks as described by Zhang and co-workers(32). Briefly,thediskswereprehybridized over-nightat 68°C in 2x SSC(lx SSC is 0.15 NaCl and 0.015 M trisodium citrate) containing 5x Denhardt solution. The labeled RNA samples werethen added to the prehybridiza-tionbuffer,andhybridizationwascontinuedat68°Cfor 48 h. The filters werethen washedtwice for30 min eachat room temperaturewith 2x SET(lx SET is 0.15 MNaCl, 20mM Tris [pH 7.8], 1 mM EDTA)containing0.1%SDS andtwice for30 mineachat60°Cwith0.1x SETcontaining0.1% SDS. They were then digested for30 min with 5 ,ug of RNase A
(Sigma Chemical Co.) perml and then washed twice more for 30 min each at 600C with 0.1x SET-0.1% SDS. The filters were then air dried and counted with a toluene-based fluor.
RESULTS
Effect ofvhsl mutation on accumulation ofdifferent viral mRNAs. In an earlier study, we showed that, following reversal ofacycloheximide block present from the time of infection and addition of dactinomycin, alpha mRNAs de-cayed more rapidly in cells infected with wild-type HSV-1 than in cells infected with the vhsl mutant (18). This
sug-A
WT VHS1
IcP0 *~
ICP27 6
*
* - *o mit
TK * a
gH - is..
1 2 3 4 5 6
7* 914011w
7 8 9 10 1112
B
WT
DNA Pol
ICP 8
gB ICP 5
gC w...
1 2 3 4 5 6 7 8
VHS 1
,...sw..
___
"-3,aw W
9 1Q 1 11 2 1 3 14 1516
FIG. 1. Accumulation of HSV-1mRNAsduring wild-type (WT) and vhsl infections. (A) Verocells infectedwith20PFU of either wild-type virus (lanes1 to 6) or vhsl(lanes7 to12)percell.Total cytoplasmicRNA wasextracted at 2.5 h(lanes1and7),4.5h(lanes 2and8),6.5h(lanes3and 9),8.5 h(lanes4and10),10.5(lanes5and 11),and 12.5 h(lanes6and12). (B)Vero cellswereinfectedwith 20 PFUof either wild-typevirus(lanes1to8)orvhsl (lanes9 to16) per cell. TotalcytoplasmicRNA wasextractedat2 h(lanes 1and9),4 h(lanes 2 and 10), 6 h(lanes3and 11), 8.25 h(lanes4and12), 10 h (lanes 5 and 13), 12 h(lanes6and14), 14h(lanes 7and15),and 16 h (lanes 7 and 16). Samples (5 ,ug) of RNA were analyzed by Northernblotting. DNAPol,DNApolymerase.
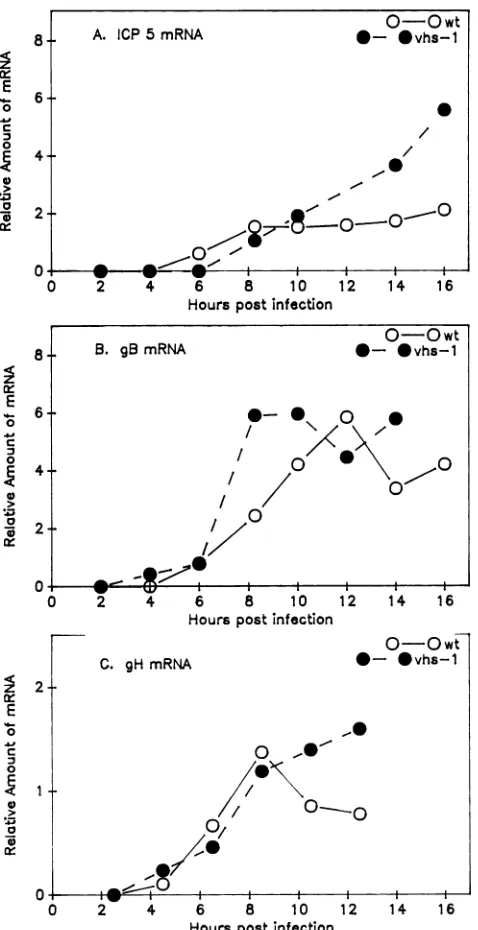
gested that,atleast under the conditions ofacycloheximide reversal experiment, the vhs gene product negatively regu-lates the stabilityofalpha mRNAs. If the Mhs gene product also regulates the half-lives of alpha mRNAs during lytic infections in the absence ofdrugs,onewould expect the vhsl mutationtoaffect theaccumulationofalphamRNAswithin infected cells. To test this prediction, Vero cells were infected with 20 PFU of eitherwild-type or vhsl virus per cell andcytoplasmicmRNAswerepreparedat2-hintervals. The levelsof selectedalpha mRNAswere thenanalyzedby Northernblottingandquantitated bydensitometricscanning ofappropriately exposed autoradiograms. Atthesametime, theaccumulation kinetics of selectedbeta, beta-gamma,and gammamRNAswerealsodetermined. The results shown in Fig. 1through 5 indicate that the vhsl mutation affected the accumulationkinetics ofanumberofdifferent viral mRNAs belonging todifferent kinetic classes.
Alpha mRNAs.The vhslmutation affected the accumula-tion kinetics of thealphamRNAsfor both ICP27 and ICPO. In cells infected with wild-type virus, ICP27 mRNA ap-peared earlyininfection, peakedin abundanceatabout 6h, and then declined (Fig. 1 and 2). By late times, only 40 to
50% of the peakamount ofmRNA was still present in the cytoplasm. Similar accumulationprofileshave beenreported previously for the alphamRNAsencodingICP22 and ICP47 (31) and ICP4 (10,31) with theexceptionthat in thesestudies the time of peak alpha mRNA accumulation was slightly earlierthan wasobservedin ourexperiments. Itislikelythat this difference wasdue to the fact that the present
experi-ments wereperformedat34°Cwhile the earlier studieswere
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.321.540.74.300.2]z
E
0 6
0 ~ ~
0
Z~~~~~~~
0 2 4 6 8 10 12 14 16
[image:4.612.325.561.74.534.2]Hourspost infection
FIG. 2. Accumulation ofICP27mRNAduring wild-type (wt)and
'h*sl infections. Autoradiograms of the gel shown in Fig. 1 were
scanned in ordertoquantitate the levels ofICP27mRNA.
performed at 37°C. As expected, the vhsl mutation led to significant overaccumulation ofICP27mRNA.Thus,incells infected withvhsl,the level of ICP27mRNA increased until 8 h and then remained high until at least 12.5 h after infection. At late times there was at least twice as much
ICP27mRNA in cells infected with vhsl as in cells infected
withwild-type virus.
In wild-type infections ICPO mRNA appeared in the cytoplasm at the same time as ICP27 mRNA and initially increased in abundance with similar kinetics (Fig. 1).
How-ever,althoughthe diffusenatureof the band for ICPO mRNA madeprecise quantitation difficult,it isapparentthatat late times the level of ICPO mRNA didnot decline tothe same
extent as did the level of ICP27 mRNA. Similarly, both Harris-Hamilton and Bachenheimer (10) and Weinheimer (31) have reported that at late times ICPO mRNA did not
decline in abundance as much as did the mRNAs encoding ICP4, (10, 31), ICP22, and ICP47 (31). In contrast to wild-type infections,in cells infected with i'hsl the level of ICPO mRNAcontinued to increase throughoutthe first 12.5 h of infection. Thus,atlatetimes therewastwo tothree timesas
much ICPOmRNA in vhsl-infected cells asin cellsinfected
with wild-type virus.
Beta mRNAs. In wild-type virus infections, the beta mRNAs encoding ICP8, thymidine kinase (TK), and DNA polymeraseall accumulated with similar kinetics(Fig. 1 and 3). Eachappeared slightly later than the alphamRNAs and increased topeak levels by 6to 8 h after infection. mRNA levels then declined sharply, so that by 12.5 h very little mRNAremained inthecytoplasm. In ourhands the decline
inabundance ofthe beta mRNAswasnoticeably sharperand morecompletethan itwasforICP27mRNA. Thispatternof accumulation was similar to those reported previously for various beta mRNAs byother investigators (10, 25, 31).
The vhsl mutation had a striking effecton the
accumula-tion of all three beta mRNAs (Fig. 1 and3). ICP8, TK, and DNA polymerase mRNAs all overaccumulated in mutant
infections. In contrast to the wild-type accumulation pat-terns, characterized by a peak and subsequent decline, in
vhslinfections the betamRNAsreachedpeak levelsat 10to
12 hafterinfection and thenleveled toaplateaurather than
declining. Thus,vhsl is defective inafunction thatplaysan
important role in the normal turning-down of beta gene expression.
Beta-gamma mRNAs. In wild-type virus infections, the beta-gamma mRNAs encoding ICP5, glycoprotein B (gB),
E
0
c
t:3
4,
0
E
0
c
-a5
E
0
0 0
*45
tp
0-Owt
5- A. TK mRNA *- *vhs-1
4-.
3
/-/
2-
/
1 -/0 0
10
1
to\°\
/
iii0 2 4 6 8 10 12 14 16
Hours postinfection
8
6
4
2
2 4 6 8 10 12 14 16
Hours postinfection
_ _ I a I
0 2 4 6 8 10 12 14 16
Hourspostinfection
FIG. 3. Accumulation of beta mRNAsduringwild-type (wt)and
i'hsl infections. Autoradiograms of thegels shown in Fig. 1 were
scanned inordertoquantitate the levels of beta mRNAs.(A) TK; (B) ICP8; (C)DNA polymerase.
and glycoprotein H (gH) all appeared in the cytoplasm at
about the sametime asthe beta mRNAs (Fig. 1, 3, and 4). However, unlike the betamRNAs,thebeta-gammamRNAs did not decline in abundance at late times. Instead,
signifi-cant amountsof thesemRNAsremainedin thecytoplasmas
late as 12to 16 h after infection.
The v'hsl mutation affected the accumulation of the three beta-gamma mRNAs to different extents (Fig. 1 and 4). However, inno instancewas the overaccumulation as dra-matic asthat observed for the beta mRNAs. At late times,
ICP5 mRNAwasoverproducedtothegreatest extentof the three mRNAs. In wild-type infections, the levels of ICP5 mRNA remainedrelatively constant at8 h postinfection. In contrast, in i'hsl infectionsthelevel of ICP5 mRNA
contin-O-Owt
8- B. ICP 8 mRNA 0- *vhs-1
6--01.
6-.
,@~-
/ *2
0i a
0- wt
C. DNA PolmRNA 0- *vhs-1
. -
_-
-/
0'0 /
_
A _ . . . i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.68.304.75.226.2]O-Owt
8-- A. ICP 5 mRNA 0- *vhs-1
6
2-- 0
0~~~~
O /~~~~~
0~~~0~
0 2 4 6 8 10 12 14 16
Hours post infection
O-Owt
8 B.g.9mRNA 0- *vhs-1
6
0-0
*/
4
/
0 0/O
2
/0
_/-0i
0 2 4 6 8 10 12 14 16
Hours post infection
O-Owt
C. gH mRNA 0- *vhs-1
2
_-0
o
0~~
1 /
/
0
A-0-
1-z-
I0 2 4 6
8
10 12 1416
[image:5.612.54.293.75.540.2]Hourspostinfection
FIG. 4. Accumulation ofbeta-gamma mRNAs during wild-type
(wt) and 'hsl infections. Autoradiograms ofthe gels showninFig.
1 were scanned in order to quantitate the levels of beta-gamma mRNAs. (A) ICP5; (B) gB; (C) gH.
uedto increase until 16 h, by which time therewas atleast 2.5timesasmuchmRNApresentasinwild-type infections.
In addition, although the amounts ofICP5 mRNA in vhsl-infected cells eventually surpassed the wild-type levels, at
early times accumulation ofICP5 mRNA in vhsl-infected cells actually lagged slightly behind that observed in cells infected with wild-type virus.
The mRNA encoding glycoprotein H accumulated with similar kinetics in vhsl and wild-type infections forthefirst 8.5 hofinfection (Fig. 1and 4).However, afterthattimethe levels ofgHmRNAin wild-type infections either plateaued
or decreased slightly, while they continued to gradually increase in vhsl infections. By 12.5 h after infection, there
z :
E
0
c
J
Q) 0
E
0
._:
0
u 2 4 6 a U0 12
Hourspostinfection 14 16 FIG. 5. Accumulation of thegammamRNAencoding glycopro-tein Cduring wild-type(wt)andvhslinfections. Autoradiogramsof the gels shown in Fig. 1 were scanned in orderto quantitate the levels ofgCmRNA.
was approximately twice as much gH mRNA in cells
in-fected with vhslasin cells infected withwild-type virus. The
vhsl mutation had the leasteffectonthe accumulationof the
mRNA forglycoproteinB, causing onlyamarginal increase
in the gB mRNA levels at intermediate and late times after infection (Fig. 1 and4).
Gamma mRNAs. In cells infected withwild-type virus, the
gammamRNA for glycoprotein C begantoaccumulate by 6
hafter infection(Fig. 2 and5). It had reached high levelsby 8 h, and large amounts of gC mRNA persisted in the cytoplasm until at least 16 h afterinfection.
The vhsl mutation led to some overaccumulation ofgC
mRNA, but only at late times afterinfection. By 16 h there
was more than twice as much gC mRNA in vhsl-infected
cellsasincells infected with wild-type virus. However,prior to 12 h, the levels of gC mRNA in vhsl infections lagged behind the levels in cells infected withwild-type virus.
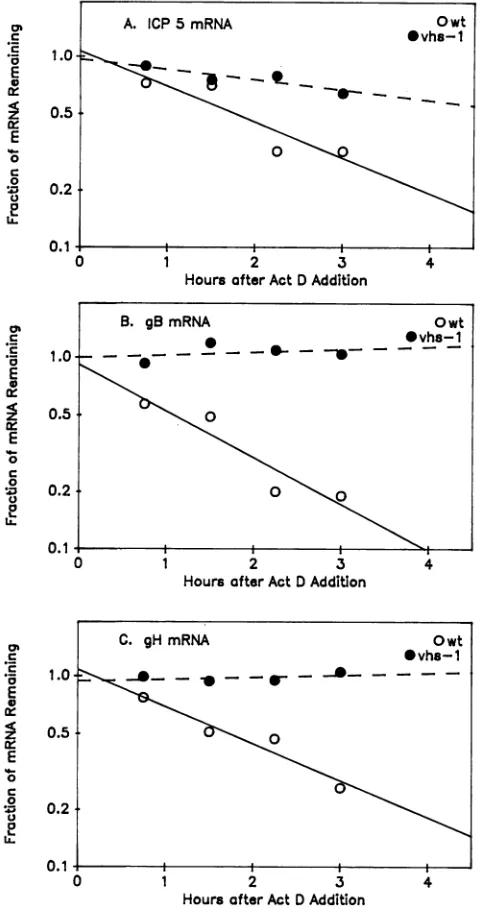
Measurement of mRNA half-lives with a dactinomycin
chase. We nextturnedourattentiontothe specificity of the lhs gene product; that is, are some mRNAs intrinsically more sensitive or resistantto vhs-induced degradation than others? In cells infected with wild-type virus, beta-gamma
andgamma mRNAs continued to accumulate at late times
when alpha and beta mRNAs were declining in abundance.
Onepossible explanation was that beta-gammaand gamma
mRNAs areintrinsically more resistantto the effects ofthe vhsgeneproduct than arealpha and betamRNAs.
Alterna-tively, all viral mRNAs may be equally sensitive tothe vhs protein, butsomemaypersistathigh levelsbecause theyare
continuously replenished by new transcription, while
tran-scription of othergenesis turnedoff. Aswasdiscussed in the
preceding section, the vhs mutation affected the accumula-tion of different viral mRNAs to different extents. Thus, whilethebeta mRNAs forICP8, TK, and DNApolymerase
were affected dramatically, accumulation of some
beta-gammamRNAs, suchasgB mRNA,wereaffectedtoamuch
lesser extent. This may reflect differential sensitivities of
different mRNAs to vhs-induced degradation.
To resolve these questions, the half-lives of 10 different viralmRNAsweremeasured beginningat6hafter infection. Wedecided that the best way to determine the question of differentialtarget sensitivitywastomeasurethe half-livesof
differentmRNAsexposedtothesamecytoplasmicfactorsat thesametime.Wetherefore choseatimeatwhich members of allkinetic classes of mRNAwerepresentinthecytoplasm O-Owt
8 j- gC mRNA *- -@vhs-1
6
0
2
/-0
S
.-i----;
ri n A e- O d^ e n AA 4.
t-xE
0
c
I
0 0
a
XI
0
E
0 ;&j
C :3
0
0
E
0
:3
.
-o
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.311.550.79.232.2]1.0
0.5
0.2
0.1
Hoursafter Act DAddition
B. ICP0mRNA Owt
0vhs-1
|~~~~~
a;X- - _0 0
---0 1 2 3 4
Hours after Act D Addition
0 2 3 4
Hoursafter Act DAddition
FIG. 6. Decay ofalpha mRNAs.Verocellswereinfected with 20 PFU ofeither wild-type (wt) virus or vhsl percell. At 6 h after
infection, 5 p.gof dactinomycin (Act D) per mlwas addedto the
medium,andincubationwascontinued.Totalcytoplasmic mRNAs
wereisolatedat45-min intervals, and 5-,ug samples wereanalyzed byNorthernblotting.Theblots wereprobedforviral mRNAs and
for28SrRNA. The 28S rRNAwasusedas anormalization standard
toensurethatequalamountsoftotalcytoplasmicRNAwereloaded
ontoeachlane of thegel. Autoradiogramsof theresulting gelswere
scanned in orderto quantitate the levels ofspecific mRNAs. (A) ICP4; (B) ICPO; (C) ICP27.
toanappreciableextent. Cellswereinfected with 20 PFU of
eitherwild-typeorvhs 1 viruspercell. At6hpostinfection, dactinomycin wasaddedtothe cultures, and total cytoplas-mic mRNAswereprepared at45-minintervals. The levels of different viral mRNAs were analyzed by Northern blotting
andquantitatedbydensitometric scanning of the
autoradio-grams. Following the analysis of viral mRNAs, the blots
were stripped and then rehybridized to aprobe specific for
C
c
.5
E
0
a:
E
0 c 0
0
.
E
0
E
0 c 0
c
10
Lt
CA ._.Ec a:
E
0
c
0
0
2.0
1.0
0.5
0.2
0.1
1.0
0.5
0.2
0.1
2 3
Hours after Act DAddition
0 1 2 3 4
Hours after Act DAddition
O 1 2 3 4
[image:6.612.281.558.73.539.2]Hours after ActD Addition
FIG. 7. Decayof beta mRNAs. Verocellswereinfected with 20 PFU of either wild-type (wt) virus or vhsl per cell. At 6 h after
infection, 5 ,ugofdactinomycin (Act D) perml was addedto the
medium,andincubation wascontinued. Totalcytoplasmic mRNAs
were isolatedat45-min intervals,and 5-p.gsamples wereanalyzed
byNorthern blotting. The blots were probedfor viral mRNAs and for28SrRNA. The28SrRNAwasusedas anormalization standard
toensurethatequalamountsof totalcytoplasmicRNAwereloaded
ontoeach laneof thegel. Autoradiogramsof theresultinggelswere
scannedinordertoquantitatethelevelsofspecificmRNAs.(A) TK; (B) ICP8; (C)DNApolymerase.
28S rRNA. This allowed us to normalize the results to
control for small differences in the amounts ofcytoplasmic RNA that were loaded ontothe different lanes ofthegel.
The results of thisexperiment andasecondsetofhalf-life determinations are shown in Fig. 6 through 9 and
summa-rized in Table 1. The vhsl mutation hadadramatic effecton
the half-lives ofall mRNAs examined. The half-life of each mRNAwasatleastseveralfoldgreater incellsinfected with
cr c
.E
0
E
0
C
0
c
2
to
L.;
0 C
._
E
o
c
._
a:
E
0
C 0
E
0
c0
C
.5 E
0
z
0 C 0
0
LL
B. ICP 8 mRNA Owt
*vhs-1
_- ~- - t-- -
-0
0~~~
b=&__<~~~~
0~
C. DNA Pol mRNA Owt
*vhs-1
- - - - _. _
,
-1
i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.86.388.79.525.2]Vb A. IGr aMRNA uwt
c *vhs-1
.0 1
E
0.5
-E
0
c
0
02
0.1eI,...
O1 2 3 4
Hours afterAct D Addition
ot B. gB mRNA Owt
*vhs-1
,C 1.0-- -a-- ____
E
0
a:
1
0.5o:
E
0 c 0
0.2
0.1
0 1 2 3 4
[image:7.612.324.544.75.207.2]Hours afterAct D Addition
FIG.o8.
Decay gHbeta-gammRNa mRNs.VercllsweeOwtwith 20 FU f ethr wld-ypevius r vsl ercel. A6vhs-1
E
i t 5 o
al:
E
0 C 0
0.2
-HoursafterAct DAdditizon
FIG. 8. Decayofbeta-gammamRNAs.Vero cellswereinfected
with20 PFUof eitherwild-type virusorvhsl per cell. At 6 hafter
infection', 5 p.g ofdactinomycin (Act D) per ml was added to the
medium,and incubationwascontinued. TotalcytoplasmicmRNAs wereisolatedat45-mmn intervals, and5-p.g sampleswere analyzed byNorthern blotting.The blotswereprobedfor viral mRNAs and
for 28S rRNA. The28S rRNAwasusedas anormalization standard
toensurethatequalamountsof totalcytoplasmicRNAwereloaded
ontoeachlane of thegel. Autoradiogramsof theresulting gelswere
scanned in order to quantitate the levels ofspecific mRNAs. (A)
ICP5;(B) gB; (C) gH.
vhsl than incells infected withwild-type virus.Infact,many
mRNAsdecayedsoslowly in vhsl-infected cells that precise
determination of the half-lifewas difficult.
Incellsinfected withwild-typevirus, the mRNA half-lives ranged from approximately 1 to 2.5 h. While it is possible that small actual differences in stability did exist between different mRNAs, the most striking result was that the
half-lives of all 10 mRNAs were remarkably similar. The
0'
c
E
0
z
: E 0
c
0
u
1.0o
0.51
0.2
0.1
Owt
0 *vhs-1
0 0
0 1 2 3
Hours After ActD Addition 4
FIG. 9. Decay of the gamma mRNA encoding glycoprotein C. Vero cells wereinfected with 20 PFU of either wild-type (wt) virus orvhsl per cell.At6hafterinfection,5,gofdactinomycin (Act D) per ml wasadded to the medium, and incubation wascontinued. Totalcytoplasmic mRNAs were isolated at45-min intervals, and 5-,ug sampleswere analyzed byNorthern blotting. The blots were probed for viral mRNAs and for 28S rRNA. The 28S rRNA was used as anormalization standard to ensurethat equal amounts of total cytoplasmic RNA were loaded onto each lane of the gel. Autoradiograms of the resulting gels were scanned in order to quantitate the levels ofgCmRNA.
correlation coefficient relating the fit of the data to acurve with the calculated half-life was determined for each of the decay curves summarized in Table 1. For ICP8 and gC mRNAs in experiment 1, the correlation coefficients were 0.804 and 0.83, respectively. For ICP27 mRNA in experi-ment2, thecorrelation coefficient was 0.84. The correlation coefficients for all of the otherdecay curves exceeded 0.90. In addition, the half-lives of five of the mRNAs were measured in twoseparate expeiiments. The average values for theresulting half-lives were: ICP27, 1.4 h; TK, 1.7 h; gB, 1.6 h; gH, 1.4h; andgC, 1;95 h. In each case the half-life values in each experiment varied from the average by at pnost 0.5 h. This gives an indication of the variability in half-life values that were obtained when half-life measure-ments were performed on different days with different batches ofcells and virus.
[image:7.612.58.298.76.532.2]In spite of small differences that were observed in the half-lives for different mRNAs in wild-type infections, it is clear that the differences between theaccumulation kinetics of, for example, DNA polymerase and ICP8 mRNAs on the
TABLE 1. Half-lives of HSV-1 mRNAs inthe presence ofdactinomycina
Half-life(h)
mRNA Wild-type virus vhsl
Expt1 Expt2 Expt1 Expt2
ICP4 1.4 Sb
ICP0 1.1 4.4
ICP27 0.9 1.9 3.3 4.4
TK 1.5 1.9 S 4.4
DNApolymerase 2.5 S
ICP8 2.0 S
lCP5 1.6 5.5
gB 1.3 1.9 S S
gH 1.6 1.2 S S
gC 2.1 1.8 S S
IDetermined fromdatainFig.6through9.
bS,Half-life greater than 8 h.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.314.553.567.710.2]one hand and gB or gC mRNAs on the other cannot be explained bydifferential mRNA half-lives. The data indicate that the vhs gene product causes degradation of many different viral mRNAs in a rather nonselective fashion. Differences in the kinetics of accumulation of different mRNAs musttherefore beprimarily duetodifferences in the rates of transcription or processing and transport to the cytoplasm.
Measurement of mRNA half-liveswith apulse-chase proto-col. The procedure of determining mRNA half-lives by measuring their decay in the presenceofdactinomycin has been used widely and produced meaningfulresultsfor many different mRNAs. However, the drug does have secondary effects on processes other than transcription, and these in someinstances may complicate half-life measurements. Fur-thermore, in some instances, turning off transcription may affect the half-lives of mRNAs that already exist in the cytoplasm. Therefore, to confirm the results ofourhalf-life measurements with dactinomycin, we determined the half-lives of selected viral mRNAs with a pulse-chase protocol.
Cellswereinfected with 20 PFU ofwild-typevirus per cell and incubated until 5.5 h postinfection. At this time the infected cells were pulsed for 30 min with
[3H]uridine.
The label was then removed, and the cellswerewashed and then incubated in medium containing an excess of unlabeled uridine as well as cytidine and glucosamine. Cytoplasmic RNA samples were prepared at 45-min intervals after re-moval of the label, and the amount of label remaining in specific viral mRNAs was determined by hybridization to filter disks containing an excess of specific viral DNA fragments. The decay curves for ICPOand TK mRNAsare shown inFig. 10. The resulting half-liveswere:ICPO, 2.4h; TK, 1.5 h. These values, obtained with the pulse-chase procedure, were verysimilar to thoseobtained by measuring mRNAdecayin the presenceofdactinomycinandtherefore confirm the earlier results.DISCUSSION
The experiments reported here demonstrate that the vhsl mutationdramatically affects thephysical stabilities of many viral mRNAs representing all kinetic classes. The data suggest that the vhs protein plays an important role, in conjunction with transcriptional controls, indeterminingthe abundance levels and kinetics of accumulation ofanumber of viral mRNAs. These studies constitute the firstsystematic measurement,in eitherwild-typeor mutantinfections, ofthe half-lives of HSV-1 mRNAs belongingtoall kinetic classes. The vhs protein is responsible for thedegradation of cellular mRNAs and the concomitant shutoff of host polypeptide synthesis that occurs early during lytic infections. The present experiments suggest that anotherimportant role of theprotein is the regulation of viral mRNA stability.
A central question addressed by these studies was the specificity or selectivity of the vhs gene product. Immedi-ately after infection, copies of the vhs protein thatenterthe cellascomponents of the infecting virioncausedegradation ofavariety of preexisting cellular mRNAs atthe sametime that viral mRNAs begin to accumulate (24, 28). However, cellular mRNAs are not necessarily intrinsically more sen-sitive to
vhs-induced
degradation than are viral mRNAs, as shown by the data of Smiley and co-workers (26). These investigators found that when a rabbit3-globin
gene was inserted into the viral genome, accumulation oftheresulting3-globin
mRNA wasregulated in afashion similar to that of members of the beta class of viral mRNAs. In the presentc
a,
a
E
a)
z
wE
0 c 0
aI 4)
1.00o
0.50
0.20
0.100
0.05
0.02
0.01
c
._
E
a)
z
E
4J
0 C: 0
0
U-
1.00-0.50
0.20
0.10+
0.05
0.02
0.01
Hours After Start ofChase
0 0
0
0
B. ICP 0
1 2 3 4 5 6
Hours After Start of Chase
FIG. 10. Measurementof mRNAhalf-livesby pulse-chase anal-ysis. HeLacellswereinfected with 20 PFU ofwild-typevirus. At 5.5 h postinfection, they were pulse-labeled for 30 min with [3H]uridine,after which the label was removed and thecells were
incubatedin mediumcontainingan excessofcolduridine, cytidine, andglucosamine.TotalcytoplasmicRNAwasextractedatvarious timesafter thestartofthechase,andtheamountof labeledICPO (B)
orTK(A)mRNAwasdeterminedbyhybridizationto an excessof specific viral DNA fragments immobilized on nitrocellulose filter disks.
studies, the vhsl mutationwas found todramatically affect the half-lives of all viral mRNAs thatwereexamined,andin cells infected with wild-type virus the vhs-regulated half-lives of all 10 viral mRNAs were very similar. Thus, the accumulated data strongly suggest that the vhs protein negativelyregulates the stability ofmany,ifnotall, mRNAs with little or no target selectivity.
As would be expected if it lengthened mRNA half-lives, the vhsl mutation affected the abundance levels and accu-mulation kineticsofanumber of viral mRNAsbelongingto different kinetic classes. The effect was most dramatic for the alpha mRNA encodingICP27 and the beta mRNAs for ICP8, TK, and DNA polymerase. Thus, these mRNAs accumulated to higher levels in v,hsl-infected cells than in cells infected with wild-type virus. In wild-type infections the mRNAsencodingICP27, ICP8, TK, andDNA
polymer-aseallappearedatearlytimes,increasedtopeaklevels,and then declined in abundance. The decline at late times was sharper and more complete for the beta mRNAs than for ICP27 mRNA. In contrast, in vhsl infections the levels of these four mRNAs failed to decline after reaching peak values and instead leveled to a plateau. An attractive hy-pothesistoexplaintheseaccumulationcurvesis thatmRNA levels continued to increase for as long as the rates of transcription, processing, and transportof the mRNA tothe cytoplasmremainedhighandthat,followingadecline in any or all ofthese rates, the levels ofalpha and beta mRNAs
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.334.547.72.352.2]declined in wild-type infections as a result of vhs-induced mRNAturnover.
Whether the normal decline in the rate of appearance of alphaand beta mRNAs in thecytoplasm results exclusively fromtranscriptional controls is unclear. In studies in which transcriptionrates were measured by pulse-labelingcells in vivowith [3H]uridine, Zhang and co-workers observed that the rate of transcription of the beta gene for dUTPase increased from1 to2 hafterinfectionand thendeclined (32). Ifthe transcription patternsforTK, ICP8, and DNA
poly-merase are similar, then the accumulation curvesforthese three mRNAs could be explained as the result of specific transcriptional controls acting in conjunction with the vhs-mediatedturnoverofthecytoplasmic mRNAs. However,in nuclear runoffmeasurementsofthetranscriptionratesofthe TK, ICP8, and DNA polymerase genes, Weinheimer and McKnight were unable todeterminewhether the decline in the steady-state cytoplasmic levels of the mRNAs resulted from transcriptional or posttranscriptional controls (31). Interpretation of theirresultswashampered by the observa-tion that significant amounts ofsymmetric, promoter-inde-pendent transcription ofthese geneswasobservedbeginning
at 4 h after infection and increased thereafter. In other studies
utilizing
thenuclear runofftechnique, GodowskiandKnipe
obtainedsimilarresultsfortherateofICP8 transcrip-tion(7). Thus,atpresent, one cannotexclude thepossibility that the normalshutoff ofbeta geneexpressionmay involve controlatthe levelofprocessingand transport of the mRNA tothe cytoplasmaswell asregulation of transcription rates and the cytoplasmicstabilities of the mRNAs.The vhsl mutation ledtooveraccumulation ofsome beta-gammaand gammamRNAs,butnot tonearlytheextentthat was observed for the beta mRNAs. The differences that were observed inwild-type infections between the
accumu-lationpatterns ofthe beta mRNAson theone hand and the
beta-gamma
and gamma mRNAsontheotherwereparticu-larly striking
in view of the fact that the half-lives of the mRNAswere very similar.The datatherefore indicatethat these differences in accumulation kinetics wereprimarily
duetocontrolsatthe levels of
transcription, processing,
or mRNA transport ratherthandifferentialmRNA stabilities.The effect of the vhsl mutation on the accumulation of
some mRNAscould notbe explained by simply postulating that themutation altered mRNA half-lives whileleavingthe rates
of
transcription, processing, andtransportunchanged. This was especiallytrue forthe mRNAforglycoprotein C. In vhsl infections, accumulation of gC mRNA actually lagged behindthat observed in wild-typeinfections until 12h,
at which point the vhsl mRNA levels finally surpassed thoseinwild-type
infections. Thereasonfor this behavior is notknown. However,overexpressionofone or moreearlier geneproductsmaydelayorreducetranscriptionortransport ofsome mRNAs that are normally expressed later in the viruscycle.The experiments presented here suggest that inwild-type virusinfections, the vhs gene productcauses rapidturnover
ofmanydifferentviral andcellularmRNAs. Thenetresultof
such a wild-scale decrease in mRNAhalf-lives would be to accentuate the
importance
andflexibility
oftranscriptional controls. It isinteresting
that HSV apparently encodes aprotein
which isinvolved in the regulationofmRNA stabil-ities and does not rely on unmodified cellular functions fordetermining
the rates ofmRNA turnover. A similar obser-vation has recently been made by Kwong and co-workers (12).The
importance
to virus growth of the vhs-mediatedregulation of mRNA levels is unclear. The fact that vhsl is a viable, nonconditionally lethal mutation indicates that a certain amountofoverexpression of many gene products can be tolerated, at least in cell culture. Nevertheless, the vhs protein is probably important to optimal virus growth, as evidenced by thefacts that the vhsl mutation resulted in a
severalfold reductioninvirus burst size insingle-step growth experiments(20) and wild-type virus rapidlyoutgrows vhsl inmixed infections(13). Furthermore, fine tuning of mRNA levels may be more criticaltogrowth ofthevirus in animals than it is incell culture.
Finally,themechanismof vhs-induced mRNA turnoveris unknown. The vhs protein may be a virus-encoded RNase or,alternatively,mayactivateapreexistingcellular enzyme. Recentdata fromourlaboratorysuggest that at earlytimes afterinfection, changes in the structure of messenger ribo-nucleoprotein particles correlate with a wild-typevhs func-tion (C. R. Krikorian and G. S. Read, unpublished data). Thus, by inducing changes in messenger ribonucleoprotein particle structure, the vhs product might render mRNAs more susceptible to digestion by RNases thatare constitu-tively present within the cell. Elucidation of the exact mechanism of vhs-induced mRNAturnoverwill await exper-iments with in vitro mRNA degradation systems.
ACKNOWLEDGMENTS
We thank Kim Knight for excellent technical assistance and AnnetteSchmidtandCharlie Krikorian forhelpful discussions.We alsothankSteve Weinheimer forprovidingtheM13 clonesandfor helpful discussions.
This workwassupportedby Public Health ServicegrantA121501 fromthe National Institute ofAllergyandInfectious Diseases.
LITERATURE CITED
1. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, and J. A.Smith. 1987.Currentprotocols in molecularbiology.John Wiley&Sons, New York.
2. Dawid,I.B.,and P. K. Wellauer. 1976. Areinvestigationofthe 5' to3' polarity in 40S ribosomal RNA precursorofXenopus laevis. Cell 8:443-448.
3. Fenwick, M. L. 1984. The effects ofherpesviruses oncellular macromolecular synthesis, p. 359-390. In H. Fraenkel-Conrat and R. R. Wagner (ed.), Comprehensive virology, vol. 19. PlenumPublishing Corp.,New York.
4. Fenwick,M.L., andJ.Clark. 1982. Earlyanddelayed shut-off of host protein synthesisin cells infected withherpes simplex virus.J. Gen. Virol. 61:121-125.
5. Fenwick, M. L., and M. M. McMenamin. 1984. Early virion-associated suppression ofcellular protein synthesis by herpes simplex virus isaccompaniedby inactivation ofmRNA.J. Gen. Virol. 65:1225-1228.
6. Fenwick, M. L., and M. J. Walker. 1978. Suppression of synthesisofcellularmacromoleculesby herpessimplexvirus. J. Gen. Virol. 41:37-51.
7. Godowski,P.J.,and D. M.Knipe.1986.Transcriptionalcontrol of herpesvirus gene expression: gene functions required for positiveand negative regulation. Proc. Natl. Acad. Sci. USA 83:256-260.
8. Goldin, A. L., R. M. Sandri-Goldin, M. Levine, and J. C. Glorioso. 1981. Cloning of herpes simplex virus type 1 se-quencesrepresentingthe wholegenome. J. Virol.38:50-58. 9. Graham, D. E., D. Medina, and G. H. Smith. 1984. Increased
concentrationofanindigenous proviralmousemammarytumor
virus long terminal repeat-containing transcript is associated withneoplastictransformation of mammaryepitheliumin C3H/ Sm mice. J. Virol. 49:819-827.
10. Harris-Hamilton, E.,and S.L.Bachenheimer. 1985. Accumula-tionofherpes simplex virus type 1 RNAs of different kinetic classes in thecytoplasmof infected cells. J. Virol. 53:144-151.
on November 10, 2019 by guest
http://jvi.asm.org/
11. Hill, T. M., R. K. Sinden, and J. R. Sadler. 1983. Herpes simplextypes1and 2 induce shutoff of hostproteinsynthesis in Frienderythroleukemia cells. J.Virol.45:241-250.
12. Kwong, A. D., and N. Frenkel. 1987. Herpes simplex virus-infected cells containafunction(s) thatdestabilizes both host and viralmRNAs. Proc. Natl. Acad. Sci. USA84:1926-1930. 13. Kwong, A. D., J. A. Kruper, and N. Frenkel. 1988. Herpes
simplex virus virionhostshutoff function.J.Virol.62:912-921. 14. Marsden, H. S., L. Haarr, and C. M. Preston. 1983.Processing ofherpes simplexvirusproteins and evidence that translation of thymidine kinase mRNA is initiated at three separate AUG codons.J. Virol.46:343-355.
15. McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan, M.C. Frame, D. McNab, L. J. Perry, J. E. Scott, and P. Taylor. 1988. The complete DNAsequenceof the long unique region in
the genome of herpes simplex virus type 1. J. Gen. Virol.
69:1531-1574.
16. Nishioka, Y., and S. Silverstein. 1977. Degradation of cellular mRNAduring infection with herpes simplex virus. Proc. Natl. Acad. Sci. USA74:2370-2374.
17. Nishioka,Y., andS. Silverstein. 1978. Requirements of protein synthesis forthedegradation ofhost mRNA inFriend erythro-leukemia cells infected with herpes simplex virus type 1. J. Virol.27:619-627.
18. Oroskar, A. A., and G. S. Read. 1987. A mutant of herpes simplex virustype1 exhibits increased stability of immediate-early (alpha)mRNAs. J. Virol. 61:604-606.
19. Pilder, S., M. Moore, J. Logan, and T. Shenk. 1986. The adenovirusE1B55Ktransforming polypeptidemodulates
trans-portorcytoplasmicstabilization of viral and host cell mRNAs. Mol. Cell. Biol.6:470-476.
20. Read, G. S., and N. Frenkel. 1983. Herpes simplex virus
mutants defective inthevirion-associated shutoff ofhost
poly-peptide synthesis and exhibiting abnormal synthesis of alpha (immediate-early) polypeptides. J.Virol. 46:498-512.
21. Read, G. S., and W. C. Summers.1982.In vitrotranscription of thethymidine kinasegeneof herpes simplex virus. Proc.Natl.
Acad. Sci. USA 79:5215-5219.
22. Roizman, B., and W. Batterson. 1986.Herpesviruses and their replication, p.607-636. In B. N. Fields and D. M.Knipe (ed.), Fundamental virology. Raven Press, New York.
23. Roizman, B., G.S.Borman,and M. Rousta.1965. Macromolec-ular synthesis in cells infected with herpes simplex virus. Nature(London) 206:1374-1375.
24. Schek, N., and S. L. Bachenheimer. 1985. Degradation of cellular mRNAs induced by avirion-associated factor during herpes simplex virus infection of Vero cells. J. Virol. 55: 601-610.
25. Sharp, J. A., M. J. Wagner, and W. C. Summers. 1983. Transcription of herpes simplex virus genes in vivo: overlap of alate promoter with the 3'end oftheearlythymidine kinase gene.J. Virol. 45:10-17.
26. Smiley, J. R., C. Smibert, and R. D. Everett. 1987. Expression of acellular gene clonedinherpessimplex virus: rabbit,B-globin is regulatedas anearly viral gene in infected fibroblasts.J.Virol. 61:2368-2377.
27. Smith, I. L., and R. M. Sandri-Goldin. 1988. Evidence that transcriptional control is the major mechanism ofregulation for theglycoproteinDgenein herpessimplex virus type 1-infected cells.J.Virol.62:1474-1477.
28. Strom, T.,and N. Frenkel. 1987.Effects of herpessimplex virus on mRNAstability.J. Virol. 61:2198-2207.
29. Sydiskis, R.J.,and B. Roizman. 1966. Polysomes and protein synthesis in cells infected withaDNAvirus.Science 153:76-78. 30. Sydiskis, R.J., and B. Roizman. 1967. The disaggregation of host polysomesinproductiveandabortiveinfection with herpes simplex virus. Virology32:678-686.
31. Weinheimer,S.P., and S. L.McKnight. 1987. Transcriptional andposttranscriptional controls establish the cascade of herpes simplex virusprotein synthesis.J. Mol. Biol. 195:819-833. 32. Zhang, Y.-F., G. B. Devi-Rao,M. Rice,R. M. Sandri-Goldin,
and E. K.Wagner.1987.Theeffect of elevated levels ofherpes simplex virus alpha-gene productsontheexpression of model
early and late genesinvivo.Virology 157:99-106.



![FIG. 10.5.5timesorandincubatedysis.specific[3H]uridine, TK Measurement of mRNA half-lives by pulse-chase anal- HeLa cells were infected with 20 PFU of wild-type virus](https://thumb-us.123doks.com/thumbv2/123dok_us/1325852.86426/8.612.334.547.72.352/timesorandincubatedysis-specific-uridine-measurement-hela-cells-infected-virus.webp)