0022-538X/87/020480-11$02.00/0
Copyright ©1987, AmericanSociety forMicrobiology
Complete Nucleotide Sequence of
a
Milk-Transmitted
Mouse
Mammary
Tumor
Virus:
Two
Frameshift
Suppression
Events
Are
Required for Translation of
gag
and
pol
R. MOORE,' M. DIXON,'R. SMITH,' G. PETERS,2 AND C.
DICKSON'*
Imperial Cancer Research FundLaboratories, London WC2A
3PX,1
andImperialCancer Research FundLaboratories, St. Bartholomew's Hospital, London ECIA 7BE,2 United KingdomReceived 1 August1986/Accepted 30 October 1986
We sequencedtworecombinant DNA clones constituting a single provirus of the milk-transmitted mouse
mammarytumorviruscharacteristic of BR6 mice. Thecomplete provirusis9,901basepairslong,flankedby 6base-pair duplications of cellularDNA atthe site ofintegration. Five extensive blocks ofopenreading frame corresponding to thegaggene, thepresumedprotease, thepolandenvgenes,and theopenreadingframeorf withinthelongterminal repeat of theproviruswerereadilydiscernible. Translationofgag, protease,andpol involved three different translationalreadingframes toproducethe threeoverlapping polyproteinprecursors
Pr77,Pr110,andPrl60found in virus-infected cells. Synthesis ofthereversetranscriptaseandendonuclease therefore requiredtwo separateframeshifts tosuppress thetermination codons at the ends of the Pr77 and PrllO domains. Direct evidence is presented for translational readthroughof bothstopcodons in aninvitro protein synthesis system.
Retroviruses are widely dispersed among vertebrate
spe-cies butareunifiedby their genomeorganization andmode of replication. An essential step in their replication is the reverse transcription of the viral RNA into a double-stranded DNA intermediate that becomes integrated as a
proviruswithin thechromosomalDNAofthehost cell. Asa
result, analyses of retroviralgenomes haverecently focused on theisolation and characterization of molecular clonesof
proviral DNA to the extent that complete nucleotide se-quences arenowavailable for several avian and mammalian retroviruses (18, 40, 42, 44, 46-49). Suchtechniquesand the
opportunity to perform site-directed mutagenesis have also
ledtotheidentification ofpreviouslyunrecognizedorpoorly
understood viral functions over and above the well-characterized components encoded bythe gag, pol, andenv genes. These includea protease, requiredforprocessing of theviralpolypeptide precursors; anendonuclease, required
for proviral integration; and specific sequences needed for
the initiation ofviral DNAsynthesis, the integration of viral
DNA, and the packaging of genomic RNA (5, 12, 16, 24, 27, 31, 32, 43, 50, 55). Whereas direct evidence for these
functionsis restricted to a few specific examples, the general
uniformity of retrovirus architecture and conservation of
particularlocalized sequence motifs suggest that most
retro-viruses conform, with only minor embellishments, to a basic prototype.
Mouse mammary tumor virus (MMTV) is unique among theRetroviridae because it represents a distinct
morpholog-icalsubclass (Btype) and is one of the causative factors in a
specific epithelial neoplasm (9, 29). However, despite being
the first mammalian retrovirus isolated, MMTV has re-mained among the most refractory to molecular analysis. Two reasons can be citedfor this: its strong tropism for the mammary alveoli, which may underlie the difficulties en-countered intissueculture manipulation of the virus, and its apparentresistance to molecular cloning as a complete DNA provirus (2, 11, 13, 25, 26, 53). Although large segments of
* Correspondingauthor.
the genome have been previously characterized (6, 11, 13, 14, 20, 25, 26, 37), here wereport the successful isolation andDNA sequencing of recombinant phage clones consti-tuting what we believe to be an entire milk-transmitted MMTVprovirus. This provides the first complete compari-sonbetween thegenome organization of MMTV and that of other retroviruses and clarifies the disposition of the open reading frames within the gag and pol domains. Previous experiments from our group and others (for a review, see reference 9) indicating threeoverlapping polyprotein precur-sorswere confirmed by the DNA sequence and extendedby the direct demonstration of two independent translational frameshifts in vitro.
MATERIALS AND METHODS
Cloningof MMTV proviralDNA. High-molecular-weight DNA from BR6 mouse mammary tumors was digested to completion with EcoRI, for which there isa single cleavage site in the MMTV provirus, and ligated into the purified armesofthe XgtWES X B vector(33).Packaged phage was plated on Escherichia coli LE392 and screened by
hybrid-izationtoaprobefor the MMTV long terminal repeat (LTR)
by standard procedures. Positive phage was then plaque
purifiedandhybridizedto5'-and 3'- specific MMTV probes
to distinguish the respective virus-cell DNA junctions (33).
Fromone particular library prepared from the tumor desig-nated W26(30), we recovered recombinants corresponding to both halves ofa single integrated provirus. These were matched initially by preparing restriction fragment probes
specificfor the cellular DNA adjacent to the viral sequences
in eachclone. The procedures used for the labeling of probes and blot hybridization have been adequately detailed else-where (33). Although the 3' junction fragment was readily transferred intoaplasmid vector to facilitate further
analy-sis, difficulties were encountered in subcloning the
corre-sponding5'junction (la).
DNA sequence analysis. The 6.7-kilobase (kb) 5' junction
fragment,containing around 1 kb of flanking cellular DNA,
and thecorresponding 6.4-kb 3' fragment were excised from 480
on November 10, 2019 by guest
http://jvi.asm.org/
their respective vectors with EcoRI and recovered by
pre-parative agarose gel electrophoresis. The fragments were
then self-ligated,randomly sheared by sonication, and
blunt-end ligated into the SmaI site of the M13 vectors mp8 and
mp9 (30). Recombinant M13 phage was grown in small-scale
liquidcultures to preparesingle-strandedDNA templatesfor
dideoxynucleotide chain termination sequencingprocedures
(30). DNA sequences were compiled by the DBUTIL com-puter program(51) and, apartfroma smallsection of the 3' LTR, were obtained for both DNA strands.
In vitrosynthesis ofMMTV-specificRNAsandproteins. A segment of MMTV proviral DNA extending from the PstI
site at nucleotide 2580 to the EcoRI site atnucleotide 5803
wasinsertedinto the polylinkerofthepSP65 plasmidvector (Promega Biotec, Madison, Wis.). Theorientationwassuch
that MMTV-specific RNA ofpositive sense could be
tran-scribed in vitro from the SP6 phage promoter with the
SP6-specific RNA polymerase (28). Conditions for RNA
synthesiswereessentiallyas describedbyMeltonetal.(28).
RNAs of different length were obtained by cleavage of the
plasmid at the specific KpnI, BglII, and EcoRI sites in the
MMTV proviralDNA(see Fig. 3). The sizeand
integrity
ofeach RNApreparation wasverifiedby Northernblot
analy-siswithanMMTV
specific probe
(notshown).
Aftersynthe-sis, the products were treated with RNase-free
DNase,
phenol extracted, and ethanol
precipitated.
Theprotein
coding capacity ofeach RNA wasthen assessed
by
transla-tion in vitro ina nuclease-treated
reticulocyte
lysate (8, 10).
The analysis of the resultant
[35S]methionine-labeled
prod-ucts by acrylamide gel electrophoresis, immune
precipita-tion, and tryptic peptide
mapping
followedprotocols
de-scribed in detail
previously
(7,8, 10).
RESULTS
Cloning and DNA sequencing of the MMTV
provirus.
Arecombinant phage library of
EcoRI-digested
DNA from aBR6 mouse mammary tumor was screened with a
probe
specific for the MMTV LTR
(33).
Since EcoRI cuts theMMTV provirus at a
single
site,
eachpositive plaque
iden-tifiedwasexpectedtorepresenta
junction
between viral andcellular DNA sequences.
During plaque
purification,
theserecombinantswere
hybridized
withprobes
specific
foreitherthe 5' or 3'
portions
oftheprovirus
(defined
relative to theEcoRIsite). In contrastto our
previous
experience
and thatof otherlaboratories in
cloning
MMTV DNA fromGR andC3H mouse mammary tumors, we recovered 5' and 3'
junction fragments
withapproximately
equal
frequency
from BR6 tumors (11,25, 26,
53). Moreover,
many of theserecombinants
represented
proviral
DNA of themilk-transmitted MMTV characteristic ofBR6 mice rather than
endogenous
sequences(35).
This wasinitially
shownby
preparing
unique
sequenceprobes
specific
for the cellularDNA
adjacent
to eachprovirus
as describedelsewhere(33,
35). Such
probes
alsopermitted
thematching
ofcorrespond-ing
5' and 3'junctions,
since theprobes
identifiedsignature
restriction
fragments
derived from theunoccupied
site innormal DNA.
Asaresultofthese
analyses,
we were in aposition
for the first time to determine thecomplete
sequence andgenomeorganization
of asingle,
potentially
infectiousprovirus
of milk-borne MMTV. Thederived sequencefortwomatchedEcoRI halves and the cellular DNA
immediately flanking
the LTRsaredepicted
inFig.
1. Six basepairs
of cellular DNA wereduplicated
at the site ofintegration,
confirming
thelinkagebetween these two
junction
fragments.
It should bestressed, however,
thatalthough
the sequencewas obtainedfor both DNA
strands,
it did not cross the EcoRI site atnucleotide
5803,
and we therefore cannotformally
excludeadditional sequence at this
position
(e.g.,
twoclosely
ap-posed
EcoRIsites).
Other features of the sequence aredescribed below and inDiscussion.
Open reading
frames in the MMTVprovirus.
From the MMTVDNAsequenceand knownfeatures of the LTRs(11,
13, 14,
20, 21,
26),
we deduced that the viral genome RNAmust extend for
8,585 nucleotides,
beginning
at nucleotide 1196(Fig.
1).
Since retroviralRNA is in thepositive
sensethe sequence as
presented
wasdirectly
translatable into the viralproteins.
Acomputed
translation of the MMTVprovirus
in the threereading
frames isdepicted
inFig.
2,
indicating
thepositions
ofallpotential
terminationcodons. It wasimmediately
apparentthatthe viral genome RNA could encompass five substantialprotein-encoding
domains. The threelargest
werepresumed
to encode the viral gag,pol,
andenv
functions,
asexpected
ofaprototype
retrovirus.Anadditional
segment
ofopenreading
frame characteristic ofMMTV and
designated
orf began immediately
proximal
to theboundary
ofthe 3' LTRaspreviously reported
(10, 11,
14, 20, 26,
37).
However,
thepresumed
gag andpol
do-mains,
nominally
ascribedtoreading
frames 1 and2,
respec-tively,
werenotcontiguous,
and itwasclear thatcontinuity
in the
generation
ofagag-pol
precursorwouldrequire
theinclusion of amino acids encoded in
reading
frame 3. Thesituation therefore
paralleled
that describedrecently
forMason-Pfizer
monkey
virus,
bovine leukemiavirus,
andhuman T-cell leukemia virus
types
I(HTLV-I)
and II(HTLV-II)
in which the viralprotease
bridges
the gag andpol
domains in adifferentframe(38, 40, 44, 46,
49,
60;
seebelow).
The data also concur withprevious
im-munobiochemical and in vitro translation
experiments
inwhich three
overlapping
precursors were identified withantisera to the viral structural
proteins
(for
a review, seereference
9).
Designated
Pr77,
PrilO,
andPrl60,
these precursorsprobably
share the sameaminoterminus,
initiat-ing
atthemethionine codonatnucleotide1508 andterminat-ing
atnucleotides3281,
4087,
and6771,
respectively
(Fig.
2).
Synthesis
ofthePrl60
pol
precursorwould thereforeneces-sitate two
separate
-1 translationalframeshifts,
switching
from frame1to3atthePr77-PrllO
boundary
andfromframe3 to 2 atthe end of
PrilO.
Demonstration of translational
frameshifting
in vitro. We and others havepreviously
shown that rabbitreticulocyte
lysates
programmed
withpurified
MMTVgenome RNAcansupport
synthesis
ofPr77,
PrilO,
andPrl60
inroughly
thesame relative
proportions
as detected inlysates
fromin-fected cells
(8, 45).
However,
these studies could neverrigorously
exclude the presence of minor RNAspecies
invirion
preparations
whichmight
directsynthesis
of thelonger
readthrough
products.
To circumvent theseprob-lems,
wefollowedtheexample
ofJacks andVarmus(19)
andconstructed recombinant
plasmids
in whichappropriate
seg-mentsofMMTVRNAcouldbetranscribedin vitro with the
specific
RNApolymerase
ofphage
SP6
(28).
Thus,
a seg-mentof MMTVproviral
DNAextending
fromthePstI
siteatnucleotide 2580to theEcoRI site at 5803 was inserted into
the
polylinker
ofthevectorpSP65 (Fig.
3).
Linearizationof the resultantplasmid
at either theKpnI,
BglII,
orEcoRI
sites within the MMTV sequences would be
expected
toyield
RNAtranscripts
of0.9, 1.8,
and 3.2kb,
which coulddiscriminate between the
proposed
translationalframeshiftsattheendsof
Pr77,
PrllO,
andPrl60.
Although
thisfragment
lacks thenormal initiationcodonatthestartofthegaggene,on November 10, 2019 by guest
http://jvi.asm.org/
tltgtgtgtgtgtgtgtotacaccttggaggggggagcggctgegttctcctgec>catcagggggtggggtgcgggtggggtollg9t9tcccc99tctoggalgggccockottccgtcag
___1CT6CA2CASAAA7665Tt
AACTCCC6A6A6T6TCCTACAC5TA666A6AACA6CCAA6666T76TT5CCCACCAA66AC5ACCC6tC16C6CACAAAC66AT6ABCCCATCA6AC
12SAAA6ACATACICATTCTCT6CT6CAAACTT66CATA6CTCt6Cl7T76CCT6666C7AtTT6666AASTT6C66TTC6T6CTC6CASSSCTCTCACCCTT6ACTCTTTtAtA6ACTCTTCT6T6CA
250A6ATTACAATCTAAAC6ATtC66A6AACTC6ACCTTCCTCCT6A66CAAS66ACCACASCCAACTTCCICtTACAA6CC6CATCAACCTTSTCCTTCA6AAATASAAATAAZAAT6CTT6CTAAAA
375AT7AtATfTTTACCAA?AA6ACCAATCCAATA66tA6ATTATIA6TTACTAT6T1AA64AAA16AATCAT7ACCTlTTASTACIAlTTTTACTCAAATTCAAA66TTA6AAAT666AATA6AAAAT
500A6iAAA6A6ACACTCAACCTCA6TT6AA6AACA66T6CA666ACTAA666CC7CA66CCTA,6AA6TAAAAA6666AAASA66A6t6C6CrT6TCAAAATA66A6ACA66T66T66CAACCA666AC
625T7ATA6666ACCTtACACTCACA6ACCAACA6AT6CCCCCTTACC6TAtACA66AA6ATAT6ACCTAAAlTTT6ATA66TS666TCACA67CAAT6BCTATAAA6T6TTATACA166TCCCTCTCCT
750T7C6T6AAA66CTCSCCA666C7A6ACCTCCTT66T6TAT6TT6ACTCA66AA6A6AAAAAC6ACAT8AAACAACA66TACAT6iATTAtATrTATTT666AACA66AAT6A6CA6CArTT6666A
875AA8AtT7TTCATAICCAA66A6A66ACA6T66CT6CACIAATABACACTATTCT6CAAA6ACTTATSAT66At6TTATlAT6ATTA6CCTTTATTA6CCCAA7CrT6T667T=CAA66rTTAA6
l000tAS6TTCAT66TCACA6ACt6T7CrTTAAACAASSAt6t6AACAA6xT6TTC7l6ACTT66ET7667ATCAAA6nT7T6ATCTAA6CTCTAAAT6CTCTAAbCCTCCTA76TTCtTTT86ATT
112*cap
CtATMcAA6TTTTAt6tAAAt6CTTAt6tAAACCAT6AIAtAAA6A'666CtAAATTTTT6A67AAACTT6CAACA67CCTAACATTCACtCTCC6T676TT76T67CT6TTC6CCATCCC6TCT
1250pbs
MC7C67CAC6TTATtCCTTCACTTTMC*6A666TCCCCCC6CA6ACCCC66T6ACCCTCA66tC66CC6ACT6
CT66C6CCC6AACA666ACCCTC66ATAA6T6ACCCTT6tCTCTAT
1375 nTCTACTATTT76t6TTC6TCTt6TTTT67CCtACtATCtTAC66C TATTATCACAA6A6C66AAC66ACTCACCACA66SAACT6CA6tCTC6CCTACA6A6AA6A66TA66TTAC66T6A6CC 1500AT7664AAAT6666TrCTC666ATCAAAA666CA6AAACICTT76TFTCT6TtCTACAAA6ACTCCTCTCA6A6A66667CTlCAT6T6AAA6A6A6CA6T6CAATA6A6TTTTATCA6TtCCTAA
1625 I10-H 6 Y S 6 S k 6 0 K L F V S V L I R L L S E R 6 L N V K E S S A I E F Y I F L ITAAA66tC7CtCClT6rTt7CCC6AA6AA66A66ATtAAATTTACAA6ATT66AAAA666T666AA6A6A6AT6AA6A66tAC6CA6CA6AACAT666AC66AtA6tATACCAAA6CA66CTTAC
1750K V S P N F P E E 6 6 L N L I D N K R V 6 R E N K R Y A A E H 6 t D S I P K Q A Y =
CATTt66CTTCA6TT66A6A6AATACT6AtA6A6CAATCA6ACTT66TTTTr6TATCT6CA6AA6CCAA6TCC6T6ACT6AA6A66AArTA6A66AA66TTTAACC66ACTAC7ATC6ACAA6
1875P I N L I L R E I L t E I S D L V L L S A E A K S V 7 E £ E L E E 6 L 7 6 L L S t S
TTCACA66AAAAAACTTA1666ACTA66664ACA6CATAT6CA6AAA7A6ATACA6A66TA6ACAA6Ct6tCT6AACAtATTTAT6A76AACCAATA6AA6AAAA66A6AA66CA6ATAAAAAT6
2000S Q E K T Y 6 t A 6 t A Y A E I D t E V D K L S E H I Y D E P V E E K E K A D K 0 E
A66AAAA66ACCAT6TTA6AAAAATAAA6AA66TA6TACAAA6AAAA6AAAATA6T6A666TAA6A6AAAA6A6AA66AlTTCAA66CCTTTTTA6CCACA6Att66AAC6AT6AT6ACCT6TCC
2125E K D H V R K I K K V V Q R K E N S E 6 K R K E K D S K A F L A 7 8 V N D D D L S
CCT6A66ATT666AC6ATFTA6A66AACAA6%P66CACArTATCAT6A76AT6AT6A6CtAIATCCTTCCA6TAAAAA66AA66T66TTAA6AA6AAACCTCA66CACTCA6AA66AAACCCCT6CC
25 P E D 0 D D L E E O A A H Y N D D D E L I L P V K R K V V K K K P I A L R R K P LPIM66T666TTT76CA66A6C6A766CA6A66CCA666AAAAA66A6ATTT6ACTTt7AC6TTTC:T6TA6TTTTTAT666A6A6A676AT6AASAT6ACAC6CC76tTT666AACC6Ct6CCAT
2375P V 6 F A 6 A N A E A R E K 6 D L T F 7 F
fP'
V F N 6 E 6 D E D D 7 P V N E P L P Lt6AAAACCTTAAA66AATT6C^AAC66CA6TTA66ACCAT666ACCAtCT6CTCCCTACACCCT6CA66T66TA6ACAT66T66C7A6TCAAT66CTCACCCC6A6t6ACt66CACCAAACA6CC
2500K t L K E L I S A V R t N 6 P S A P Y t L Q V V D
0
V A S Q N L 7 P S D N H Q t AA6A6CTACCT76TCtCCT6A6ACTAT6tTTTAT66A6AACT6AATAT6AA6A6AAAA6tAAA6AAAT66TACAAAA466CT6CA66CAA6C6AAA666CAA66TCTCTCrT6AtAT6TTACT666
262S R A 7 L S P 6 D Y V L 0 R 7 E Y E E K 6 K E N V- Q K A A 6 K R K 6 K V S L D N L L 6CACT66CCAAltCC6TNCCCCTTCTTCTCA6ATAAAATT6TCTAA66AT6TCrTTAAA6AT6TCACCACAAAT6C76T6TTA8CAT66A666CCATTCC6CCTCCT66A6TTAAAAAA6ACT6tAT 2750
7 6 I F L S P S S I I K L S K D V L K D V t t N A. Y L A N R A I P P P 6 V K K t V L
TA6CA66ArTAAAACA666AAAT6.AA6A67CTTAt6A6,ACTTTCATTTCAA66CTC6A66AA6CT6TTTACC6AAT6AT6CCCA6A6666AA6667C66ATAtATT6ATCAAACAArT616C6T66 2675
A 6 L K Q 6 N E E S Y E t F I S R L E E A V Y R N N P R 6 E 6 S I I L I K Q L A N
B6AAT6CAAAITtATt6T6TC*6AtCtCATCC6CCCAAtACST^AAAACA66AACTATAtCA66ATTA7ArTC676CTT6TCT86AC6CTTCTCCC6CA6T767TCA1666tAT66CATAT6CA6C 3000
E A N S L C O D L I R P I A K t 6 t I D. D Y I R A C L D A S P A V V Q 6 N A
YFAA
A6CCAT6A6A66ACAAAA67T7tCtACCTTT6TAAA6CAAACAIAT66T66666AAAA66A66TCAA66A6CA6AA666CCA6TT767TTTTCCT6T667AA6ACA66ACACATCA6AAAA6ACT 312S
A N R 6 I K Y S t F V K I t Y 6 6 6 K 6 6 Q 6 A E 6 P V C F S C 6 K t 6 H I R K D C 6tAA66AT6AAAA666CTCAAAAA6666CCCICC7666CtCT6CCtCC6AT67AASAAA66CtA7CACT66AA6A6T6A6t67AAATCTAAATTT6ACAAA6AT666AATCCACTTCCTCCCTT6 325
K D E K 6 S K A A p f I L C P R C K K 6 Y H 0 K I E C K S K F D K D 6 N P L P P L
MCT*AtgATBCT7MTT6i6ACT1A6CA67tCCCCt6Ct6CCAAAA66666AT66A6TtA A466WCTP.A6AXTTZATClC1';AAGCACttCT7*TCALATA6TTT76 33 E T N A E I S K 0 L 0
31- F K K L V K I I 8 P 8 P A I K 6 D 6 V K 6 S 6 L k P I A P P F 7 la N D L
FIG. 1. Completenucleotide sequence ofamilk-transmitted MMTVprovirus.TwoEcoRIfragmentsrepresentingtheS'and 3'ends ofa singleMMTVproviruswere recoveredasrecombinant clones andsequenced by shotgun cloninginto M13 vectorsanddideoxynucleotide
chain terminationprocedures(30, 33).The sequenceacrosstheEcoRI sitewasnotdetermined, but withtheexceptionofasmallsection of the 3' LTR,allsequenceswereobtained for both strands. Shownare9,901 nucleotides ofproviralsequence,numberedfrom base 1 of the S' LTR and flankedbysegmentsof thecellular DNAadjacenttotheprovirus(show'nin lowercase).Theboundaries of the LTRsareindicated
by rectangularbrackets,and the six bases of cellular DNAduplicatedduring proviral integrationandunderlined(-).Thebeginningsof
longopenreadingframes arealso shown (D-, accompanied byanumberreferring tothetranslation frameused). Other sequence features indicatedarethe S' end of the viral genome RNA(cap);thebindingsite fortRNA31Y'(pbs);thesplicedonor(sd)andacceptor(sa)sites for
envandorfmRNAs;the knownamino-terminal sequences ofp27,p14-p30, gp52,andgp36( r' ),and thepotentialinvertedrepeatsequences distaltotranslational frameshift sites(overlined) (11, 13, 14, 17, 20, 21, 25, 26, 34, 37, 54, 57).
on November 10, 2019 by guest
http://jvi.asm.org/
PRST BSAS DLSS KDLI SLED YSLV
TTASTLAPCC
ECC6MATAE'7
MtC6A66CACCCCTS6A6T6C:E6rTT7ACCT6TCATCACASAAZ6A6ATTTCTCC7tCTCtAA6AA66AT6ATATCArT6tSACCCAC6t6AA6ACCCTCCt
666.G P A 6 t P 6 S A 6 L I L S 6 I K D L I L S L E D 6 V S L V P T L V K 6 t L P E 6 T 7T66ACTAATAATABSTA6AA6TTCTAATTATAAAAA666ACT76A66TTTTACCA66A6TCATT6ACTCC6ATTTCCAA6b6ASAMtCA66T7AT66TTAA66M6CiAAAAAT6C66TCATCA
3625 6 L I 1 6 R S SUY K k 6 L E V L P 6 V I D S D F 0 6 E I K V N V K A A K N A V I I TTCACAAA86A6AAA6^AtA6CACAACcT676T76jC76CCATATTTAAMTTACCCMATCCT6TATCSsAA66AASACA66CTCA6AS6CTTC66ATCAACAA67CAT6T6cATT666T6CA6
3750N K 6 E R I A I L L L L P Y L K L P I P V 1 K I EfR 6 S E 6 F 6 S T s H V H U v Q
6AAATAA6T6ACTCCA6ACCCA76CTTCACATTTACTTMAT66AA6
6ATTM7C66TCtCT766A7ACC6666CA6ATAAACTT6CATA6CA66CA6A6ACT6.6cCSCTAATT66CCCAT 3875 E I S D S A P N L H I Y L N 6 R R F L 6 L L D T 6 A D K 7 C I A 6 R I N P A N N P ITCACCAAAC76A6A6TTCTCtTtCAA66T76CA766CCTS6T666ST66C6C6TA6TASTCA6CCACTCC6TT66CAACAT6A66AT^AAATCA66AATTATACATCCUTT6tT6ATCCCTACAC
4000 N O 7 E S S L Q 6 L 6 N A C 6 V A R S S I P L R V Q N E I t S 6 1 I N P f V I P t L 76CCTTTCACCTTAT6666AA6A6A7ATTAT 6AAA6ATATA AA66TCAatTTAA6AMACTACATATCACCB%nAlTTA16AT46CM16A6CAAI
TC CTTT6CA6ACCAAA 4125P F T L 1 6 R 9 I N K D I I t A L # 7 I S P I D S I D L # #
210-F T S F N I 6 A I E S N L F A D Q I TATCTT66AA6TCAAC6AMSATJsJ*T#ntBXAT
ASTt=
ArAA6AAAtTTAACA66CTTTACAAC6TATACASAtACTTACAAT6666C
ACTTAAA6A6A6CAATA6C 4250S N K S D I t V N L N I V P L K Q E K L I A L I I L V t E Q L I L 6 H L E E S k S
CCtT6AATAC6CC76tTTTT6TCATTAAAAAA6AA1CA66AAAAT66A6ACT6TTACAA6ACC7AC6T6CA6TTAAT6CTACAAt6CAC6ATAt666A6CATTACAACCC66CTT6CC6TCCCC
4375 P H N 7 P V F V I K K K S 6 K N R L L I I L R A V N A 7 H H I N 6 A L I P 6 L P S PT67A6CA6T6CCCTAAA66AT666AAATAATCATAATA6ATCTACAA6ATT6CTfTTTTAAATAAAAACT6CATCCTSAA6AltTTAATM
6AtTT6CtTTTA676T6CCCTMCCCTAATTTTAA6A 4500 V A V P K 6 N E I I I I D L I D C F F N I K L N P E D C K R F A F S V P S P N f K R6ACCrTATCAAA6ATTCCAAt66AAA6tTTTTCCCCA666TAT6AAAAATA6CCCTACTTTATSTCAAAAArTT6T66ACAAA6CTATATT6ACT6TAA666ATAAATACCAA6ACTCATATATT
4625P Y I R F I 9 K V L P O 6 N K N S P T L C I K F V D K A I L 7 Y R D K Y I D S Y I
6T6CATtACA166AT6ACATtCtTtT66CACACCCATCAAi6AtCCATTSTC6AT6AAATACrTAC7?CCAT6ATACAS6CCCTTAACAAACAT66CCT?6tA6TAICCACA6A6AA6ATTCAAAA
4750 V N Y N D D I L L A N P S R 6 I V D E I L 7 S N I I A L N K H 6 L V V S T E K I O KtAtA6AtAATCTCAAATAtTTT666AACTCAtATACA666T6ATTC46T6tCTTATCAAAAATTACA6ArTA66ACA6ATAAArTAA6AACCTTAAAT6ATTTCCAAAA46CTArTA66AAATATTA
4675 Y D N L K Y L 6 t H I I 6 D S V S V I K L I I 0 t I K L A T L k D F Q K L L 6 N I NAT766ATAC61CCTTTCTTAAAATTAACTAC666A6A6TTAMACCTCTCrTT6AAATTCTTAAT66A6A77CTAATCC6ATCTCAACAA6AAAACTtAC7CCT6A66CAT6CAAA6rTCttCAA
5"00N I R P F L K L t T 6 E L K P L F E I L N I D S N P I S t R K L t P E A C K A L Q
TTAAT6AAT6A6A6ACTATCTACC6CTC66ETAAA6A66CtA6ATTTATCACA6CCTTB6tCTCTAT6tAtATTAAA6ACT6AATATACCCCCACA6CAT6CCTCT66CA66AT66A6TTSTA6A
5125 L N N E R L S T A R V K R L D L S I P V S L C I L K 7 E Y t P 7 A C L V Q D 6 V V EAT66AtACATTTSCCTCATATTTCACCAAA66T6ATTACTCCTTAT6ATATCrTrTTTTACA%AACTTATTATTAA666CC64CACC6tCTCAAA6AATTAtTTA6TAAA6ACCCT6ATTATATTS
5250 N I H L P N I S P K V I t P Y D I F C t I L I I K 6 R N R S K E L F S K D P D Y I VT76T6CCCTACACCAAA6rTCAATT76ATCTCCtATtACAA6AAAA66AA6ATT66CCTATt7CTTlATTA666TTCTT666A6A66TTCArTTCCATCTTCCAAAA6AcCCCT7T6CTTACATTT
5msV P Y 7 K V I f D L L L O E K E D I P I S L L 6 F L 6 E V H F N L P K D P L L t f
ACCCTACAAACT6CCtAlTlTATTCCTCACAt6ACCTCTACCACACCACTASA6AAA66AAT76T6ATTTTTACA6AC666TCA6CAAAt66CC6TTC66TAACATATATACAA66AA666A6CC
5500 t L I t A I I f P N N t S 7 t P L E K 6 I V I F t D 6 S A 1 6 R S V t Y I Q 6 R E PTATAATTAAA6AAAATACACAAAACACA6CCCAACA66CT6AAATT6T66CASTCATTACA6CCYTT6A66AAT6A66TCAACCCTTTAATTT6ATAACT6ATTCTAAATAT6T6ACA666TT67
5625I I K E 0 t I N T A O Q A E I V A V I t A f E E V S I P f N L Y T D S K Y V t 6 L f
TTCCC6AAATC6AAACT6CAACTTT6TCACCCA6AACAAAAATTTACACA6AACT6AAACATTTACAAA66TTAATCCACAA6A6ACAA6AAAAATtT7ACATT66TCATATCA6A66ACACACT
5750P E I E t A t L S P R t K I Y t E L K N L O R L I N K R I E K F Y 1 6 N I R 6 N t
68ACTTCCC667CCTTT66CACA666AAAT6CCTAT6CA6ATTCTTTAACAA6AATTCT6ACC6CTTTA6A6TCA6CTCAA6AAABCCAC6CACTACATCATCAAAAT6CC6:66CCTTA66TT
5075 6 L P 6 P L A I 6 N A V A D S L T R I L t A L E S A O E S H A L H H I N A A A L R fTCA6TTTCACATCACTC6T6AACAA6C6C6A6AAATA6TAAAATTAT6TCCCAATTSCCCC6ACT66666CAC6C6CC6CAATTA66667AAACCCCA6666CCTTAA6CCCC6A6TTCTAT66C
6000O F H I t R E I A R E I V K L C P N C P D 9 6 R A P I L 6 V N P R 6 L K P R V L k O
AAAT6,6AYT7tACTCAT6TTTCA6AATTTSSAAAATTAAAAtAT61ACAT6T6ACA6T66ATACTlATTCtCATTTTACTT7C6CIACC6CCC6AAC666C6AA6CAACCAA66ATT61TTACAA
t125N D V 7 H V S E f 6 K L K Y V N V T V D 7 Y S H F t F A 7 A N t 6 E A 7 K D V L O
UCATT66CTCAAA6CTTT6CATACAT666CATTCCTCAAAAAATAAAAACA6ATAAT6CCCCT6CATAT761TCTC6TTCAATACAA6AATTTCT66CCA6AT664AAAAIATCTCAC6TCAC666
in0 N L A I S F A Y N 6 1 P I K I K t D N A P A V V S R S I I E F L A R H K I S H V t 6CATCCCCTACAATCCCCAA66ACA66CCATT6TT6AAC6AAC6CACWCAAAATATAAA66CACA6CTlAAIAAACTTCAAAA66CT66AMATAAC7ATACACCCCATCATC76TT66CACAC6CTC
6375I P Y N P Q 6 Q A I V E R t N O N I K A O L N K L Q K A 6 K Y Y T P N N L L A N A L
tTTTTT6TCT6AATCAT6TAAATAT66ACAATCAA66CCATACA6C66CC6AAA6ACAtTT6666TCCAiATCTCA6CC6ATCCAAAACCTA766TCAtT666AAA6ACCTTCTCACA666TCCT6)6
6500F V L N N V N N D N I 6 H t A A E A H H 6 P I S A D P K P N V N V K I L L T 6 S V
AAA66ACCC6AT6TCCTAATAACA6CC66AC6A66C7AT6CTT6T6TTTTTCCACA6iAt6CC6AAACACCAAICT666TCCCC6ACC6ATTCATCC6ACCTTTTACT6A6C66AAA6AA6CAAC
"2 K 6 P D V L I t A 6 A 6 V A C V FR~~
I E t P I I V P D A f I R P F t E R K E A tsa-3I, P K H O S 6 S P 7 D S S I L L L S 6 K K I R
IICCCACACCT66CACT6C66A6AAAAC6CCSCC6C6A6AT6A6AAA6ATCAACA66AAA6TCCCAA6AATSAATCTA6TCCCCATCAAAMA6A6AC66CtTT6CMCATCT6CA66C6TT6ATC
6750 P 7 P 6 7 A E K t P P R D E K D I O E S P K N E S S P N I R E D 6 L A t S A 6 Y D LP H L A L R R K A R R E N R K I N R K V P A N N L V P I K E K 7 A V I N L O A L I
tcC6AA6C66A66A66TCCTTAAAACCT>CACAACCCCCAAACCTCTTTACCTTATTlCT76CtT76TT6TCT6TCCTC66cCCCCCC6CCT6t6ACA6666A6A6TTATT666CCTACCTACC
6875 A S 6 6 6 P #6 E A E E V L K t S I t P I t S L 7 L F L A L L S V L 6 P P P V E S A LP
TAAACCACCTATTCTCCATCCC6T666AT6666AA67ACA6ACCCCAT7A6A6TTCT6ACAAATCAAACCAT6ATTTi666766TTC6CCT6ACTTTCAT666TTCA6AAATAT67TCt66AAT6
7"00
K P P I L H P V 6 V 6 S T D P I R Y L t 0 I t N V L 6 6 S P D F H 6 F R N N S 6 N V
7ACAlTTT6A6666AA6TCT6ATAC6CTCCCCATTT6CCTTTMTCTCTCClTTTCtACCCCCAC666CT6ClTTCAA67A6ACAA6CAA6TATTTCTTTCT6ATACACCCAC66TTlATAATAAT
7125 H F E 6 K S I t L P I C L S F S F S t P t 6 C F I V D K Q V F L S D t P t V D N NAAACC766666AAA666T6ATAAAA66C6TAT6T666AAClTt6tT76ACTACCTT6666AACTCA6666CCMTACAAMAC766TCCCTATMAAAASAA6TT6CCCCCCAAATATCCTCACT6
7250 K P 6 6 K 6 D K R R N V E L V L 7 t L 6 N S 6 A N t K L V P I K K K L P P K Y P H CMAAbTC6CCtTTAA6AA66AC6CCT7CT666A666A6AC6A6TCt6CTCCTCCAC66T66TT6CCTT6CBCCttCCCT6ACCA6666T6t6ATTTTCTCCAAAA6666CCCTTS666TTACTTT
7ms
O I A f K K D A F V E 6 D E S A P P R V L P C A F P D C 6 V S F S P K 6 A L 6 L L V
666ATTTCTCCCTTCCCTC6CCTA6T67A6ATCA6tCA6ATCA6ATTAAAA6CAAAAA6AATCTATTT66AAATTATACTCCCCC76TCAATAAAGA66TTCATC6AT66TAT6AA6CA66AT66
7500 D F S L P S P S V D Q S D Q I K S K K N L f 6 N Y t P P V N K E V H R v Y E A 6 k6tA6AACCTACTT66TTCT666AAAATTCtCCTAA66ATCCCAAT6ATA6A6ATTTCACT6CACTA6TCCCCCATAAAA776TT7CSCTTASTCSCA6CCTCAA6ACATCTTATTCTCAAAAS
7625 V E P t V f N E N S P K D P N D R D F t A L V P H t E L F R L V A A S R N L I L K RFIG. 1-Continued.
on November 10, 2019 by guest
http://jvi.asm.org/
BCCA66ATTTCAASAACA16A6AT6AlXCCTACATCT6CCT6T61TACTTACCCTTAT6CCATATIATTA66AlT£CTCiA6TTiAA76TATA6A6AAAA6A66ATCTACTTTTCATATTTCCT
P 6 F O E H E H I P T S A C V T Y P Y A I L L 6 L P I L I D I E K A 6 S T F N I S C
6TTCTTCrT67A6ATT6ACTAATl6TTTA6ACTCTTCT6CCTAC6ACTAT6CA6C6AICATA6TCAA6A66CC6CCATAT676CT6CTACCT6TSA6AATT66T6ATBAACCAT66TTT6AT6AT
S S C N L 7 N C L I S S A Y D Y A A I I V K A P P Y V L L P V D 1 6 D E P N F D D S A I I T F R Y A t LL I R A K RJTV A A I I L 6 1 9 A L I A I T7 S F A V AT T A
CT7A6T1AA66A6A76CAAACT6CIACST6TTAArtTAATCTTCAtA66AA76TTACAT7A6CCTTATCT6AACAAC66ATAATA6AMAAATTA6AA6CIAMCTTAAT6CTTTA6A6AA6AA
L V K E N I 7 A T F V 0 k L N R N V 7 L A L 6 E Q R I I D L K L E A R L N A L E E V TASTfT7tA6ATT666ACAA6At6t66CCAATTTAAA6ACCA6AATSTCCACTA667T6CAT6CAAATTA76ACTTTA7Ct6C6T7ACACCTT7ACCCTAtAAT6CtACT6A6AACT666AAA6A
V L E L 6 I I V A N L K 7 A N 6 7 P C H A 0 V D F I C V t P L P V H A 7 E N N E R ACCA666CTCATTTATT666CAlTT66AAT6AtAAt6A6AlTTCA7AIAACATACA66A6T7AACCAACCT6AlTA616A7AT6A6CAAACAACATAT76AT6CAST66ACCTTA6T66CTT66C t R A H L L 6 1 1 N I H E I S Y 0 I 1 E L t N L I C I N 6 K O H I D A V I L 6 6 L A
7750
7,75
3000 8125 8250 .375
ttCATCTTTT6CCAAT66A6T6AA6CTTTA)AATCCATTA6ATT66ACACAATATTTCATTTTTAtA6676TT66A6CCCT6CTTTTA67CAT76tACTTAt6ATTTtMCCATT6TTTTCCA6T 0500
0 S F A N S V K A L N P L I U T C Y F I F 1 6 V 6 A L L L V I V L N I F P I V F I9 a
KCCTT6C6AA6A6CCTT6ACCAA6T6CA6TCA6ATCT7AACT6CTTCTTTTAAAAAA6AAAAAA66666AAlE6CC'6C6CCT6CA6CA6AAATBST766
ACTCCCBA6A6T6TCCTACACTTAiU625
L A K S L I I V I S I L N V L L L K K K K S6 6 A A P A A E H V E L P RVSYYT* 11H P R L 09 K I L N S R E C P t L R
S6A6AA6CA6CCCUS666TT6TTTCCACCAA66AC6ACCE6TCTZCGEACZAACG6AT6A6ECCATCACACAAA6ACATACTCATTC7CTCt6CCAAACTt66CATA6CTC76CTTTGCT66 1750 6 E A A K 6 L F P T K D D P 6 A N K A N 6 P 6 D K D I L I L C C K L 6 1 A L L C L 6 66CtATT66666M6TT6C66TTC676CTCBCA666CTCTCACCCTT6ACTCTTTttAAA6CTCrTCT6T6CAA6ATTACAATCTAASC6ATTC66A6AACTC6AeCTTCCTCCt6A66CM66A
L L 6 E V A V R A N R A L T L D S F N S 6 S V I D Y N L N D S E N S T F L L N Q 6
POPT6SYHWUWW% 16161I 0P6ECIRgHLSI EIYINoT"N""I"LAKNYIFTK"""NMIIn IN III III T K166ITN SPODPRL I MI I RDL LV TN1 I
P Q P 7 6 S V K P H I P C P S E I E I N N L A K N Y I F 7 N K 7 N P I 6 R L L V T I ctTTAAAAYuTrATTArrzrT=TACriAsTTTTirTr^aATir^ALcctT^crAA^t=cAAAAAcAaTcanasrarrArrTrAhrTPACTTcAA&ArAnyacr=rrTAAcalrrT
mtv
3575
9000
aIJr,
a*IIwINR"fDMlo In"l IIIInI§WI INII IIRL ILA" II1IL"I"ARRDI iR"W""""lbombIM6Lbo iboIRAI IWRIk"bAII nb"RLMV " NMLI VBo"I TIAD
L A I E I L t F 1 7 I F 7 O I I R L E N 6 I E N R K A N 6 7 6 V E E O V I 6 L R A 6 MC68CT*'6AA61AAAAASCSAAABA66A6T6C6CTT6TCAAAATA66A6ACA66T66766CAACCA666ACTTATA6666ACCT7ACATC7ACA6ACCAACA6,AT6MCCCTTACC6tATACA 9250
6 L E V K R 6 K A 6 A L V K J 6 1 N I N Q P 6 t V N 6 P Y I Y R P 7 0 A P L P Y t
66AA6AIAT6ACC7AAA7TTT6ATA66T666TCACA67CAA766C7ATAAA6T6TTATACA66TCCCTC7CCTTTC6T6AAA66CTC6CCA666CTABCC7CCTT66T6TAT6TT6ACTCA66 9m 6 A Y b L N F I N H V T V N 6 Y K V L Y R S L 6 F A E N L A N A R P P N C N L t I E A6A6AAAAAC6ACAT6AAACAACA66TACAT6ATTA7AlTTATTT666AACA66AAT6A6CA6CAtTTT6666AAA6ATrTTTCATACCAA66A6A66ACA6766CT6CAIC7AAIABA6CACTATT "00
E K H I 0 K I I V H D Y I Y L 6 T 6 P S S I 9 6 K I F H t K E A T V A A L I E N Y 6 CT6CAAA6ACTTA166CA76A6TTATlA16AT7A6CCTTTAlT7ACCCAATCT76T66TTCCCA66T7AA67AA6TTCA766TCACA6ACT61TCTTAAAACAA66AT6T6A6ACAA6T66T 9US
A K T 6 N S6 Y YDI
Mt6ACT766CT766TATCMAAt67tT6ttbCCTAACCTAAAT6CTCIAACTCCtAT6T7CTtTt76ATTCTAtCC6tTttAT67AAAT6CttAT6tAAACCAt6ATATAAAAST6rKT 9750
975 AC*CCTCAC*6TC86CC8ACTCstcctcgag_
&ccaccgFtgagttag
-gCogagtincgtuedg.ccggtgcttcgtcgcgggttcgctcgctagctatctggatcactccc
FIG. 1-Continued.
we noticed a fortuitous ATG codon 36 nucleotides distal to thePstI site, which was likely to act as an initiator under the
more promiscuoustranslation conditions of the reticulocyte
lysate. The 0.9-kb RNA species should therefore be
trans-lated into a 25-kilodalton (kDa) protein which terminates at
theTAAcodon at the end of Pr77 (nucleotide 3281), whereas
readthrough of this codon should yield a 28-kDa species
terminated at theKpnIsite marking the end of the transcript
(Fig. 3a).Similarreasoning for the 1.8-kb RNA would predict
naturally terminated 25- and 51-kDa species and a 64-kDa
readthrough product ending at the
BglII
site, whereas the3.2-kb RNA should yield 25-, 51-, and 92-kDa products.
Theresultsobtainedby translating these RNAs in a rabbit
reticulocyte lysate and by analyzing the
[35S]methionine-labeledproducts by polyacrylamide gel electrophoresis (7, 8)
are shown inFig. 3b. Densitometric scanning of the
autora-diographs, with corrections for the relative contents of
methionine, indicated that the predicted products were
pre-sentin proportions consistent with approximately 15 to 25% readthrough of the successive termination codons. With RNA terminating at the EcoRI site, the low level of the
92-kDagag-pol product presumably reflected the
vulnerabil-ity of this longer RNA to degradation in the lysate or premature termination of the translation machinery. A sim-ilar explanation could be proposed for the background in these gels, since breakage of RNA chains permits initiation
atessentiallyanymethionine codon encountered close to the
5' ends of the resultant RNAfragments (8, 10).
Neverthe-less,theindicatedproductswerespecificallyprecipitated
by
antiseraagainst themajorgagdeterminant, p27 (not
shown).
Partial amino acidsequencingof thevirionproteins p27and p14 has established their amino termini at nucleotides2315
and2976, respectively (N. Totty, M. Waterfield,R.
Moore,
M. Dixon,R.Smith, G. Peters,andC. Dickson) sothatthe
in vitro translation products predicted in Fig. 3a should include approximately 60% of theprimarysequenceof
p27.
Trypticpeptide analysis of in vitro readthroughproducts.
To verify the identity of the in vitro translation products
detected in Fig. 3, we prepared fingerprints of the
methio-nine-labeled tryptic peptides from the 25-, 51-, and 92-kDa
proteins(Fig. 4). Previous reportsfrom this laboratory have
described two-dimensional tryptic-peptide maps of
individ-ualMMTVproteinsandofPr77, Prl1O, and Pr160produced
either in vivoorin vitro(7, 8). A schematic representationof thepattern of spots that weobtained from Prl60expressed inareticulocytelysate(8) is shown in Fig. 4d. In thisearlier work, it was established that peptides 1 through 8 were present in Pr77 and peptides 9 and 10 were additionally present in PrllO, whereas only peptide 11 distinguished Prl60 from Prl1O. Although not all of these peptideswere represented in the present work (because of the artificial initiation site generated in the SP6 transcripts), it was clear that peptides 9 and 10 were again acquired in the readthrough from the 25- to 51-kDa species and that peptide AAATTTTT6ASTAAACTTBCAACASTCCTAACATTCACCTCTCSTSTSTTTST6TCTSTTC6CCA7CCC6TCTCC6CTC6TCACTIATCCTTCACTTTCCA6A666TCMCCCSCA64CCM66T
w_,....
on November 10, 2019 by guest
http://jvi.asm.org/
a
LTR gag pol
°
1 2 11 1111 11 11 kb11111111
21
2 3
b Pr77 0.
Prl6O *..:..j *.. ...
Pr731
FIG. 2. Dispositionof openreading frames in the MMTV proviral sequence. (a) Computed translation oftheMMTV DNA sequence of Fig. 1in the three possible reading frames. The positions of all potential termination codons are indicated relative to the linear map from nucleotides 1 to 9901. (b) Open reading frames shown in panel a directly correlated with the precursor polyproteins identified in MMTV-infectedcells andcharacterizedbyimmuneprecipitationandtryptic peptide mapping (9). Thedifferentreading frames are depicted by thesymbols: a,frame 1; [lii,frame 3;mm, frame 2. Thus Pr77 is translated in frame 1,PrllOspans frames 1 and 3, andPrl60uses frames 1, 3, and 2. Theopen reading frame orf in the LTR is not included in this figure, since the predicted protein has not been detected in MMTV-infectedcells (8, 10, 45).
11 was acquired in the readthrough from the 51- to 92-kDa
products. Moreover, theothermajor peptides were entirely
consistentwith the initiation of translation within the coding region of virion protein p27.
DISCUSSION
The majority of previous studies on the MMTV provirus
and sequence have concentrated on the milk-borne viruses
characteristic of the GR and C3H strains of mice or on
endogenouselements. Although there have been numerous
reportsonthe sequenceoftheLTRs, theenvgene,and parts
ofthepol gene,noneofthese hasprovidedacomprehensive
picture ofthe MMTV genome (6, 11, 13, 14, 20, 25, 26, 37).
Oursuccessinobtaining recombinantclones encompassing the 5' half of the genome mayin some measure reflect our
choice ofthe BR6 mouse strain. The exact origin of the
milk-transmitted MMTV in these mice remains unclear,
since they were derived initially from a cross between an
RIII male and a nonviremic C57BL female (15), but we assume that this virus represents the onethat is character-isticofRIIImice. However, theso-called poisonsequences which have hampered many studies on MMTV are not
completely
abrogated in thisstrain,
since we encountereddifficultieswhenever attemptswere madetotransfer cloned
5' junction fragments from A vectors into
plasmids (la).
Itwas therefore impracticable to reconstruct a complete
provirus byjoiningthetwoEcoRI
junction
fragments,whichthus precluded any tests on the
biological
activity
ofthisprovirus. Nevertheless, all thefeatures of the derived DNA
sequencewouldfulfilthe
expectations
foraninfectious virus andwould be entirely consistent with the knownbiochemi-cal data.
The5'LTRandleadersequences. The LTR of theprovirus shown inFig. 1 extends for 1,328 base pairs and is bounded
bytheexpected 6-base-pairinverted repeats (11, 14, 20, 25).
The various elements required for initiation of transcription
and glucocorticoid regulationhave been described in detail
by others(for a review, see reference 39) and are maintained in our sequence with only scattered base changes. Flanking the LTR to the 5' side is the hexanucleotide CGTCAG
duplicated upon integration of the provirus into cellular
DNA(11,20,25), andatthe3' boundary is the18-nucleotide
bindingsite for the DNAsynthesis primertRNA-ys(25, 34).
The sequences distal to the primer binding site are loosely
described asleader sequences, since they precede thestart ofthe gag gene, buttheyencompassthesplicedonor site for
envmRNAat nucleotide1484(Fig.1) andpresumablyallor
partofthe
packaging
signal forMMTV genomeRNA(13, 26,27, 54, 55).
gag gene. Identification of the ATG codon at nucleotide
1508astheinitiator for the gag gene productsis based on a number ofconsiderations: it is the first ATG encountered
afterthe LTR
(Fig.
1),itconformstotheGNNATGconsen-sus(23), itis followedby590 codonsin frame
(Fig.
1and2),and itpredictstheaminoterminalMet-Glysequence consis-tent with the observed
myristylation
of the MMTV gag precursor(13, 41). However, weasyethavenoinformation that would allow us to align the amino-terminal virionprotein,
plO,
with the DNA sequence. Partial amino acidsequencinghas allowed ustomatch the N terminus of
p27,
themajorcore
protein,
with theproline
codonatnucleotide2315, and upstreamofthis
point
aretwodomains ofaround20 amino acids, one of which is
composed
of 50% acidicresidues, the other being 50% basic. This latter
region
isreminiscent ofthe
highly
basic virionprotein, p8,
describedenv
LTR
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.131.475.70.315.2]a
P K Bg E
i ~~i
[111111:
_SP6
K . 0.9kb
2s :.-~I
Q8
7 6
00
90
203
O'2
11 01c)lo
O5
0
4 [image:7.612.65.311.57.510.2]01
FIG. 4. Tryptic peptide maps of in vitro translation products. Two-dimensional fingerprints were prepared of tryptic peptides from the 35S-labeled in vitro products depicted in Fig. 3b (7, 8). Shown are the 25-kDa (a), the 51-kDa (b), and the 92-kDa (c) productsand aschematicdiagram of thepreviously reported finger-print of in vitro synthesized Pr160 (d) (8). Numbered peptides correspondtothose discussed in earlierpublications andare repre-sentedaccordingtothe framesymbols used in Fig. 2. The symbols A and A indicate additional peptides found in the 51-kDa and 92-kDaproducts, respectively.
464
_M 451
"428
a
425 *425_451
25
_, 2 5
FIG. 3. Translationalframeshiftingin vitro.(a) Partial restriction
mapof MMTVproviral DNA. The positionsofsignificant (butnot
all) PstI(P), KpnI (K), BgIII (Bg),and EcoRI(E)sitesareindicated.
The segment from PstI to EcoRI depicted by the bold line was
subcloned into the polylinker of the pSP65 vector such that
tran-scription of the resultant plasmid DNA by the SP6 phage RNA polymerase initiated at the specific promoter (*) and copied the MMTVsequences into positive-sense RNA(19, 28). Linearization of the plasmid at the KpnI, BglII, orEcoRI sites yielded specific
RNAtranscripts (.--) of 0.9, 1.8, and 3.2 kb, respectively. When thesewereusedtodirect protein synthesis inarabbit reticulocyte
lysate (8, 10), the expected products had the sizes (in kilodaltons) and composition shown in the diagram. The symbols for each translationalframe correspondtothose usedin Fig. 2. Termination occurred eitherat the normal stopcodons oratthe 3' end ofthe specific RNA transcripts. (b) In vitro translation productsgenerated fromRNAs terminating attheKpnI (K), BglII (Bg), orEcoRI (E) sites in the MMTV sequence. These products were analyzed by electrophoresis in 10% polyacrylamide gels (7, 8), labeled by the addition of[35S]methionine tothereticulocyte lysates, and visual-ized by autoradiography. The positions and molecular sizes (in kilodaltons) of the relevant products are indicated. The No RNA
lane shows the background of protein synthesis observed in the
previously (7, 9). The amino-terminal sequences ofvirion protein p14 and the minorprotein p30 both align perfectly with the DNA sequence beginning at nucleotide 2996 (N. Totty, M. Waterfield, R. Moore, M. Dixon, R. Smith, G. Peters, and C. Dickson, unpublished results). P14 is the majornucleic-acid-binding protein ofMMTV, a conclusion that issupported bythepresenceoftwoof the motifs(N,N + 3, N+ 13)ofcysteineresidues identified in othersystems (1, 4). However, any function ofp30 remains a mystery, since to our knowledge, no equivalent protein has been described for other retroviruses. As p30represents a
trans-frameprotein, sharingan amino terminus withp14 in frame 1 andacarboxyterminus withPrllOinframe3,it willclearly be interesting to determine the complete amino acid
se-quence of p30 to define exactly where in the nucleotide
sequence the frameshiftoccurs.
Protease. One consequence ofexpressing viral structural proteins in the form ofprecursors is the requirement fora
specific protease to process these polypeptides into the mature forms. It is therefore hardly surprising that
retrovi-ruses encode theirown enzymes toperformthis role, butit has become apparent that they have adopted various strat-egiesforimplementingthisfunction. Avianleukosisviruses, for example, include the protease within the gag gene; in murine andfeline leukemiaviruses, the enzymeisformally
nuclease-treatedlysate. We notedthat several of the major in vitro products migratedastriple rather than single entities in acrylamide
gels,butwehavenoexplanation for this phenomenon,norhavewe
been abletodistinguishdifferences between the various species.
a
_ ... ...
b
Bg 1..kb
25 ..-..
C d
E 3.2kb
25
b
NO
RNA K Bg E
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.330.565.70.299.2]VOL. 61, 1987
MMTV
aMPMV
RSV MoMLV HTLV-1 VTS N A
MYTVt MPMV
R SIV"
MoMLV HTLV-1 VISNA
MMTV
MPMV RSV MoMLV HTLV-1
VISNA MIVI MPMV RS, V MOML.V MT L V-1 VISNA
b
MNTV% MPMVMloM LV HTL'V- 2.
VISNA
M TV MPMV PRSV MoMLV MTLV -1 VISNA
MMTV MPMV
MoMLV H L -A VISNA
MMTV DNA SEQUENCE
DPLTNDKLAAAJLV QZQMEASITKSs$ L
DUWLPEGILV<WV EKEL4I£PS
LISW
IVWVZRKA WSYALLHRAVU4
KQYRVSAR LGSIKPHI Q L.LEQtrLVPCQSMU LLPV*PGNNYRPVGS
.JEVN
QPVPFPgUlQHL RKALEAQUIXYT G?GN?VFPVPLWA NGTISF TM
AQI WTUEGtKEIVDRLEKEGKVGF¶AFPHWTCWNTIIFCZIW 151"MLLIDFREL"U
.AV
Kf$KlflsvnSPM.AlVrarJ
AKLTVPF AVQQAPVLSAPA WPLMVLD&KCWSIPLAEQORE A PAPTVfNQA
LRvED:MrilHPTV PN PYNLLSGLPS FPS HQWYVLDLKIDAPFC LRF,L SQF' nEWR D?EMS:' S LT'IDLSSSSPQPPDLSSLPrTLAH LQTWb>A"QI PLPKQFQPYIWTVW rQYG KQTEDLAEAQLSHPGGLQ RKKHVTILRIGDAYVTIPLYEPYFQY'TCFTMLPN'ULG
487
Vt
flQKFVP%ALLTVWfDKYQWYlSWYXPZLL&MFWtZVUZ
P!siMJIKn?QAIbtonYvAsATA;HA kn AWkQMY-IwLIAGK:;GQQVLQ
PARVtKVLJT I0TVVGQ\ EPLRLKHF..CML*I£LLAASSH LEA
SGQLT W#TRRf$
S
C;QT
~~~~~~~~~~~~bT
DAHl~:RQHPODLILLYvRL4AtSEI.DFRI?EP>LTC _L ; F E LFAaXv4 SE }i
PGNAYWKtWAH.L Z, P R- PWf,vFI].L4wA-V
PCvatYYIKVL WRLS#A;YQFTMQIK:TLF.JIEErPRT W M : QFGWY:GV DLLEA-, L,3S EHF.
CFDQLKQEL"T AAHI
APS.,VQLQEDP
YTYWFELNGP AG-E E VISSTLE RA TI SPDKVQREEPS VQflYKLGSTGTR ALL TLG NLRx'AA KKAQICQQVKIITtLAY KEG LSEATMASLI SDSLPVIENKT,QQZKTPC;M-KFIAQIZSPN
GI7TLVN F LASYrAQYSFlMPyE.CQEJYP AKWLSFL H?E
ZLT*AIEBSAQKSEALHQNAAALRFQFHITREQAREIKVKLCPNCPDWGRAPQL GVNPR INfNL&% A4Hr, NAC77L T-TMP .1IFIt
MAR
I VW-YCP Cp'9 L GVNPRQAYErL?EvAKLMTALML 3PF;.ALSKAThNIZXQARE V&?CPF CNS^ PAL.EA GVNPIR THLSF S.MKA ..EFSWHS2r'lYMLNFDFNI..C>CAASA.^G'P. P P L, L.;S; AE:RESXTEB ALTTLQ,a A T:TT:& C AeC fH.- w =^A.G7@ -FWR
WP ALrSFEAHFI H TA L.LIECriIGT?A- 5C sQ ' FR
GLKPRVL WQMDVTHVSEPGKL KYVHVTVDTYSHFTFATARTGQATKDVLQJLAQSFA QLFPM' WQMDVTh"'SEflNL KYtNV:H;: DTwS PAT1G"TrTK .T HL.z-;V: Fs GLGPLIWT-T-DFT FF3 WLA.-,?Y£VTVDTA.SJw s/W}T';- -;. fi;HA?A
GHRPPS-TH WE DFTE IF-n3L_Y frKYI.VFY D--FIDTFS A E -KvTrKLYF GLS1P H 9WQ D :TN F;-.IV K NTr L FHVAVDT?S AIX.T K 3:5JLA:A
GSN DxR=ID1WQ.DYTEx.rf-, LP TNSs' .AerG: .- rhVxMKWYA
YMGIPQKZKTDNAPAYVSRSIQEFLARWKXSHVTGI PYNPQOQAIVERTHQNIKAQLNKL :AIBLPKQW1KTDlM PGP-YTYSVVrQEFW½CI2--eVI-VH:TGITPYNPQQQ:I--VLRAHLKT??:E K: VLG^RPKAIKTDNIGJ>_W-LSKS? EnW-ARW 2It*^H.TGi;P *eNLQGQAMVKRCEA\Esi .K21F IRVL
1-. PKAI NTnw DNff.:3`_Y-FSKS.--E'r..-LARW I P. TGyI P:: N,- .'*vRIJsVK
F FGMPQ r_TD-3PAN'-AFVSi` I It.-.C YPQ'QKLVJ - IKL'
HL_GKP'SYINTDNSPAYSCW..'HKT>PYN?`x3/K.
lgFAPrK.3EZDXN.3PA FV AEES', .J--;-. ..iTGIPA"NP.QA"H VKRTHQt-'K\ BK
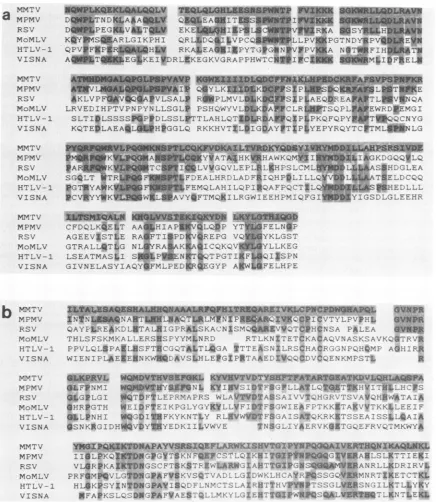
FIG. 5. Amino acid sequence homologies amongretrovirus polgenes. Sequence comparisons are presented between thepolgenes of
MMTV(thispaper), Mason-Pfizer monkeyvirus(49), Roussarcomavirus (42), Moloney murine leukemia virus(47),HTLV-I(44),andVisna virus(48). Residues homologous to the MMTV sequence are shaded. The distinction between polymerase (a) and endonuclease (b) domains isbasedonthefeaturesof these proteins in Rous sarcoma virus and Moloney murine leukemia virus.
partofthepol gene, occurringdownstream of the termina-tion codon separating gag and pol; and inHTLV-I,
HTLV-II, bovine leukemia virus, and Mason Pfizer monkey virus, the proteaseis encoded in a separate reading frame between the gag and pol genes (5, 18, 19, 38, 40, 42, 44, 46, 47, 49,
58-60).
Thesituationin MMTV most closely parallels that of the last virus group in that the sequence LLDTGADK, which is homologous to the consensus active site of retrovi-ral and some cellular acid proteases (52), occurs in frame 3(centered onnucleotide 3821)in thebridgebetween the gag
and poldomains (Fig. 1and 2). Although we are not yet in a
positiontodefine the boundariesor exactsize ofthe MMTV
protease, it is clear from these data that its expression
requiresatleast onetranslational frameshift.
Polymerase and endonuclease. Characterization of the MMTV reverse transcriptase has been and remains a con-tentious topic, with conflicting reports ofa 100-kDa mono-mer or 85- and 55-kDa dimers as the active moiety (for a
review, seereference 9). Now that the fullDNAsequenceis
known, it should bepossibletoresolvethis issueby
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.86.524.67.569.2]a
Frame 1
Frame 3
b
Frame 3
Frame 2
Pr77 /PrllO READTHROUGH
3260
GCTGAAAATTCAAAAAACTTGTAAAGGGGCA
A E N S K N L
* K F K K L V K G Q
PrllO/Prl6O READTHROUGH
4069 4
GATGATTCACAGGATTTATGATAGGGGCCAT
D D S Q D L
* F T G F M I G A I
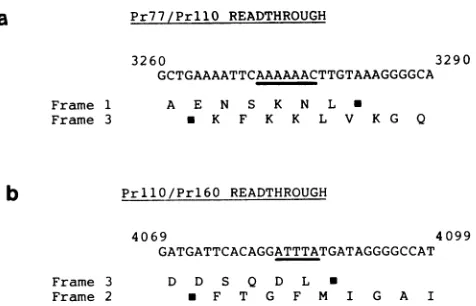
FIG. 6. Sequences involved in translational frameshifting. The
DNA sequences spanningthe boundaries between Pr77 and PrllO (a)and between PrllO and Prl6O(b) are displayed in a simplified
form.Theaminoacidsencoded in therelevant frames areindicated by the single letter code, with stop codons also shown (i). The sequence motifs discussed inthe textare underlined.
ing and aligning terminal amino acid sequences. A similar strategy will be required to characterize the endonuclease domain, since the presence of poison sequences has pre-cluded the types of mutagenesis studies applied to other
retroviruses (12, 16, 32, 43). Asnoted by others, the coding domains ofpolymerase and endonuclease are nevertheless readilydiscernible intheMMTVsequence simplyby homol-ogiestootherknown sequences (3, 6, 38,40, 42, 44,46-49).
The degree of conservation is most striking within the polymerase itself(Fig. 5), particularly between MMTV and
Mason-Pfizervirus,wherethehomologyintheregionshown
is around65% (Fig. 5) (49).
envgene.Wepreviously reportedtheenvgene sequenceof
the GR strain of MMTV and discussed its implications in
terms of the viral glycoproteins and their precursor Pr73 (37).Thenucleotide sequenceof thecorresponding region of
the BR6provirus differs at60positions, reflectedin only10
aminoaciddifferences. Five ofthese changesarelocalized
withinthe leaderprecedingthegpS2domain(17, 26, 37), but
theirsignificanceisunclear,since wedonotyetknowwhich
ATGcodonserves astheinitiatorforPr73(26, 37). None of
theessentialfeatures ofthe two matureglycoproteins, such as the number and position of glycosylation sites or the
natureofthe transmembraneanchor, isaffected.
The3'LTRandorf.The mostsignificant featureof the3'
LTRofMMTVistheexistenceofasubstantialopenreading
frame designated orf, which begins at an A residue in the
polypurine tractimmediatelypreceding the boundary of the
LTRandcontinuesfor962residuesinthe U3 portionof the genome(8, 10,11, 14, 20, 26, 45). Wepreviouslyargued that
themaintenanceoforfinseveralendogenousand milk-borne
strains ofMMTVindicates it hasafunctional role (9). The sequence presented in Fig. 1 extends this argument, since
the integrity ofthe reading frame is maintained despite 94 single-base differences between this sequence and the GR
virus LTR. Moreover, the putative splice acceptor site mapped approximately50nucleotidesupstreamofthe LTR isalsorepresented in oursequence (54, 57) (Fig. 1).
Suppression by translational frameshifting. Although it is
stillunclearhoworwhy orfmay beexpressed, synthesisof theenvgenefromaspliced, subgenomicmRNApresumably reflects the need for high levels ofthe viral glycoproteins, roughly equivalent to the levels ofthe gag structural
pro-teins. In contrast, the protease,
polymerase,
andendonucle-ase are nonstructural components, and
although
they
may occurinvirions,
they
arerequired
inonly
catalytic
amounts. It has long beenrecognized
that infected cells express thegag-pol
precursor atonly around 5% ofthe level of thegag precursor, and it hasrecently
become apparent thatretrovi-ruses haveadoptedanunusual strategy,
namely
suppression
of termination
condons,
forcontrolling
the levels ofthesefunctions (19, 36, 58-60). InMMTV, there arethree
precur-sors, nowidentifiedas
pr77gag
prllogagpro,
andPr160gag
propol,so that two termination codons must be
bypassed
during
synthesis ofthe
pol
gene.Thisrequired
twoframeshifts(Fig.
2),and bothevents couldoccurinrabbit
reticulocyte
lysates
primed withdefinedsegmentsof MMTV RNA
(Fig.
3and4).
From the ratios of readthrough
products
to terminatedproducts, we estimated that each frameshift occurred with
an efficiency of around 15 to 25% in
vitro,
which isreason-ably consistent with the levels of Pr77,
PrilO,
andPrl60
observed for infected cells (7, 9). However, these studies cannot formally exclude the
possibility
ofself-splicing
orsplicing catalyzedby components ofthe
reticulocyte
lysate.
The sequences at which frameshifting occurs are dis-played in more detail in Fig. 6. At present, we have insuffi-cient information to determine where within the
overlaps
between frames1 and 3in
Fig.
6aandbetween frames 3and2 in Fig. 6b the actual switches occur. Resolution ofthis issue awaits protein sequence data for the virion
p30
and protease, the twolikely candidates for products which span these frameshifts. However, wenote thatboth theseframe-shift sequences comply with precedents set by other retro-viruses, in that one occurs at or near the sequence ATTTA, as in avian sarcoma virus (19,
42),
and the other probablyinvolves the AAAAAACsequence identifiedasthe potential frameshift site in bovine leukemia virus,
HTLV-I,
andHTLV-IIand paralleled in some procaryote systems (22, 38,
46, 56, 60). Moreover, both frameshift sites are closely
followed by potential stem-loop structures in the virion
RNA. Although we have not assessed the thermodynamic
stability of such secondary structural features relative to
othersin thesequence, it is interestingtospeculatethatthey maycontribute in cis toframeshifting (38, 46, 60). Whether suchprocesseshavewider significance in the generalcontrol
ofgene expression remains an intriguing possibility.
ACKNOWLEDGMENTS
We thank S. Brookes for additional technical help; L. Crawford, J. Wyke, and J. Witkowski for comments on the manuscript; and A. Kessler for its preparation.
LITERATURE CITED
1. Arthur, L. O.,C.W. Long, G. H.
Smith,
and D. L. Fine. 1978. Immunological characterization of the low molecular weight DNA binding protein of mouse mammary tumor virus. Int. J. Cancer 22:433-440.la.Brookes, S., M. Placzek, R. Moore, M. Dixon, C. Dickson, and G. Peters. 1986. Insertion elements and transitions in cloned mouse mammary tumour virus DNA: further delineation of the poison sequences. Nucleic Acids Res. 14:8231-8245.
2. Buetti, E., and H. Diggelmann. 1981. Cloned mouse mammary tumor virus DNA is biologically active in transfected cells and its expression is stimulated by glucocorticoid hormones. Cell
23:335-345.
3. Chiu,I. M., R. Callahan, S. R. Tronick, J. Schlom, and S. A. Aaronson. 1984. Major pol gene progenitors in the evolution of oncomaviruses. Science 223:364-370.
4. Copeland, T. D., S. Oroszlan, V. S. Kalyanaraman, M. G.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.72.310.66.219.2]Sarngardharan, and R. C.
Gallo.
1983. Complete amino acid sequence of human T-cell leukemia virus structural proteinp15. FEBS Lett. 162:390-395.5. Crawford, S., and S. P. Goff. 1985. A deletion mutation in the 5' part of thepolgene of Moloney murine leukemia virus blocks proteolytic processing of the gag andpolpolyproteins. J. Virol. 53:899-907.
6. Deen, K. C., and R. W. Sweet. 1986. Murine mammary tumor virus pol-related sequences in human DNA: characterization and sequence comparison with the complete murine mammary tumor viruspolgene. J. Virol. 57:422-432.
7. Dickson, C., and M. Atterwill. 1979. Composition, arrangement and cleavage of the mouse mammary tumor virus polyprotein precursorPr77gagand pllOgag. Cell 17:1003-1012.
8. Dickinson, C., and G. Peters. 1981. Protein-coding potential of mouse mammary tumor virus genome RNA as examined by in vitro translation. J. Virol. 37:36-47.
9. Dickson, C., and G. Peters. 1983. Proteins encoded by mouse mammary tumour virus. Curr. Top. Microbiol. Immunol. 106:1-34.
10. Dickson, C., R. Smith, and G. Peters. 1981. In vitro synthesis of polypeptides encoded by the long terminal repeat region of mouse mammary tumour virus DNA. Nature (London) 291:511-513.
11. Donehower, L. A., A. L. Huang, and G. L. Hager. 1981. Regulatory and coding potential of the mouse mammary tumor virus long terminal redundancy. J. Virol. 37:226-238.
12. Donehower, L. A., and H. E. Varmus. 1984. A mutant murine leukemia virus with a single missense codon inpol is defective in a function affecting integration. Proc. Natl. Acad. Sci. USA 81:6461-6465.
13. Fasel, N., E. Buetti, J. Firzlaff, K. Pearson, and H. Diggelmann. 1983. Nucleotide sequence of the 5' noncoding region and part of the gag gene of mouse mammary tumor virus; identification of the 5' splicing site for subgenomic mRNAs. Nucleic Acids Res. 11:6943-6955.
14. Fasel, N., K. Pearson, E.Buetti, and H. Diggelmann. 1982. The region of mouse mammary tumor virus DNA containing the long terminal repeat includes a long coding sequence and signals for hormonally regulated transcription. EMBO J. 1:3-7.
15. Foulds, L. 1949. Mammary tumours in hybrid mice: the pres-ence and transmission of the mammary tumour agent. Br. J. Cancer3:230-239.
16. Grandgenett, D., T.Quinn, P. J.Hippenmeyer,and S. Oroszlan. 1985. Structural characterization of the avian retrovirus reverse transcriptase and endonuclease domains. J. Biol. Chem. 260:8243-8249.
17. Henderson, L. E., R. Sowder, G. Smythers, and S. Oroszlan. 1983. Terminal amino acid sequences and proteolytic cleavage sites of mouse mammary tumor virus env gene products. J. Virol. 48:314-319.
18. Herr, W. 1984. Nucleotide sequence of AKV murine leukemia virus. J. Virol. 49:471-478.
19. Jacks, T., and H. E. Varmus. 1985. Expression of the Rous sarcoma virus pol gene by ribosomal frameshifting. Science 230:1237-1242.
20. Kennedy, N., G. Knedlitschek, B. Groner, N. E. Hynes, P. Herrlich, R. Michalides, and A. J. J. van Ooyen. 1982. Long terminal repeats of endogenous mouse mammary tumourvirus contain a long open reading frame which extendsintoadjacent sequences. Nature (London) 295:622-624.
21. Klemenz, R., M. Reinhardt, and H. Diggelmann. 1981. Sequence determination of the 3' end of mouse mammary tumor virus RNA. Mol. Biol. Rep. 7:123-126.
22. Kohno, T., and J. R. Roth. 1978. A salmonella frameshift suppression that acts at runs of A residues in the messenger RNA. J. Mol. Biol. 126:37-52.
23. Kozak, M. 1984. Compilation and analysis of sequences up-stream from the transcriptional start site ineucaryotic mRNAs. Nucleic Acids Res. 12:857-872.
24. Lobel, L. I., and S. P. Goff. 1985. Reverse transcription of retroviral genomes: mutations in the terminal repeat sequences. J. Virol. 53:447-455.
25. Majors, J. E., and H. E. Varmus. 1981. Nucleotide sequences at host-proviral junctions for mouse mammary tumour virus. Na-ture (London) 289:253-258.
26. Majors, J. E., and H. E. Varmus. 1983. Nucleotide sequencing of an apparent proviral copy of env mRNA defines determinants of expression of the mouse mammary tumor virus env gene. J. Virol. 47:495-504.
27. Mann, R. S., R. C. Mulligan, and D. Baltimore. 1983. Construc-tion of a retrovirus packaging mutant and its use to produce helper-free defective retrovirus. Cell32:871-879.
28. Melton, D. A., P. A. Krieg, M. R. Robagliati, T. Maniatis, K. Zinn, and M. R. Green. 1984. Efficient in vitro synthesis of biologically active RNA and RNA hybridization probes from plasmids containing a bacteriophage SP6 promoter. Nucleic Acids Res. 12:7035-7056.
29. Moore, D. H., C. A. Long, A. A. Vaidya, J. B. Sheffield, A. S. Dion, and E. Y. Lasfargues. 1979. Mammary tumor viruses. Adv. Cancer Res. 29:347-418.
30. Moore, R., G. Casey, S. Brookes, M. Dixon, G. Peters, and C. Dickson. 1986. Sequence, topography and protein coding poten-tial of mouse int-2: a putative oncogene activated by mouse mammary tumour virus. EMBO J. 5:919-924.
31. Panganiban, A. T., and H. M. Temin. 1984. Circles with two tandem LTRs are precursors to integrated retrovirus DNA. Cell 36:673-679.
32. Panganiban, A. T., and H. M. Temin. 1984. The retrovirus pol gene encodes a product required for DNA integration: identifi-cation of a retrovirus int locus. Proc. Natl. Acad. Sci. USA 81:7885-7889.
33. Peters, G., S. Brookes, R. Smith, and C. Dickson. 1983. Tumorigenesis by mouse mammary tumor virus: evidence for a commonintegration region for provirus integration in mammary tumors. Cell 33:369-377.
34. Peters, G., and C. Glover. 1980. tRNA's and priming of RNA-directed DNA synthesis in mouse mammary tumor virus. J. Virol. 35:31-40.
35. Peters, G., M. Placzek, S. Brookes, C. Kozak, R. Smith,and C. Dickson. 1986. Characterization, chromosomalassignment, and segregation analysis of endogenous proviral units of mouse mammary tumor virus. J. Virol. 59:535-544.
36. Philipson, L., P. Anderson, U. Olshevsky, R. Weinberg, D. Baltimore, and R. Gesteland. 1978. Translation of MuLV and MSV RNAs in nuclease-treated reticulocyte extracts: enhance-mentof the gag-pol polypeptide with yeast suppressor tRNA. Cell 13:189-199.
37. Redmond, S. M. S., and C. Dickson. 1983. Sequence and expression of the mouse mammary tumour virus env gene. EMBO J. 2:125-131.
38. Rice, N. R., R. M. Stephens, A. Burny, andR.V. Gilden. 1985. The gag and pol genes of bovine leukemia virus: nucleotide sequence and analysis. Virology 142:357-377.
39. Ringold, G. M. 1983. Regulation of mouse mammary tumor virus gene expression by glucocorticoid hormones. Curr. Top. Microbiol. Immunol. 106:79-103.
40. Sagata, N., T. Yasunaga, J. Tsuzuku-Kawamura, K. Ohishi, Y. Ogawa, and Y. Ikawa. 1985. Complete nucleotide sequence of the genome of bovineleukemia virus: itsevolutionary relation-ship to other retroviruses. Proc. Natl. Acad. Sci. USA 82:677-681.
41. Schultz, A. M., and S. Oroszlan. 1983. In vivo modification of retroviral gag gene-encoded polyproteins by myristic acid. J. Virol. 46:355-361.
42. Schwartz, D. E., R. Tizard, and W. Gilbert. 1983. Nucleotide sequence of Rous sarcoma virus. Cell32:853-869.
43. Schwartzberg, S., J. Colicelli, and S. P. Goff. 1984. Construction and analysis of deletion mutations in thepol gene ofMoloney murine leukemia virus: a new viralfunction required for pro-ductive infection. Cell37:1043-1052.
44. Seiki, M., S. Hattori, Y. Hirayama, and M. Yoshida. 1983. Human adult T-cell leukemia virus: complete nucleotide se-quence of theprovirus genomeintegratedin leukemia cell DNA. Proc. Natl. Acad. Sci. USA 80:3618-3622.
45. Sen, G. C., J. Racevskis, and N. H.Sarkar. 1981. Synthesis of
on November 10, 2019 by guest
http://jvi.asm.org/
murine mammary tumor viral proteins in vitro. J. Virol. 37:963-975.
46. Shimotohno, K., Y. Takahashi, N. Shimizu, T.Gojobori, D. W. Golde, I. S. Y. Chen, M. Miwa, and T. Sugimura. 1985. Complete nucleotidesequence of an infectious clone of human T-cell leukemia virus type II: an open reading frame for the protease gene. Proc. Natl. Acad. Sci. USA82:3101-3105. 47. Shinnick, T. M., R. A. Lerner, and J. G. Sutcliffe. 1981.
NucleotidesequenceofMoloneymurine leukemiavirus. Nature (London) 293:543-548.
48. Sonigo, P., M. Alizon, K. Staskus, D. Klatzmann, S. Cole, 0. Danos, E. Retzel, P. Tiollais, A. Haase, and S. Wain-Hobson. 1985.Nucleotide sequence ofthevisna lentivirus: relationship totheAIDS virus. Cell 42:369-382.
49. Sonigo, P., C. Barker, E. Hunter, and S. Wain-Hobson. 1986. Nucleotide sequence of Mason-Pfizer monkey virus: an im-munosuppressive D-typeretrovirus. Cell 45:375-385.
50. Sorge, J., and S. H. Hughes.1982. Polypurinetractadjacentto the U3 region of the Rous sarcoma virus genome provides a cis-acting function. J. Virol.43:482-488.
51. Staden, R. 1980. A new computermethod for the storage and manipulation of DNA gel reading data. Nucleic Acids Res. 8:3673-3694.
52. Toh, H., M. Ono, K. Saigo, and T. Miyata. 1985. Retroviral protease-like sequence in the yeast transposon Tyl. Nature (London) 315:691.
53. Ucker, D. S., S. R. Ross, and K. R.Yamamoto. 1981. Mammary
tumorvirusDNAcontainssequencesrequiredfor its hormone regulatedtranscription. Cell27:257-266.
54. Van Ooyen, A. J. J., R. J. A. M. Michalides, and R. Nusse. 1983. Structural analysis of a 1.7-kilobase mouse mammary tumor
virus-specificRNA.J. Virol. 46:362-370.
55. Watanabe,S.,andH. M. Temin.1982.Encapsidationsequences for spleen necrosisvirus,anavianretrovirus,arebetweenthe5' longterminalrepeatand the startof thegag gene. Proc. Natl. Acad. Sci. USA 79:5986-5990.
56. Weiss, R. B. 1984. Molecularmodel of ribosomeframeshifting. Proc.Natl. Acad.Sci. USA81:5797-5801.
57. Wheeler, D. A., J. S. Butel, D. Medina, R. D.Cardiff,andG. L. Hager. 1983. Transcription of mouse mammary tumor virus: identificationofacandidatemRNAfor the long terminalrepeat geneproduct.J. Virol.46:42-49.
58. Yoshinaka, Y., I. Katoh, T. D.Copeland, and S. Oroszlan. 1985. Murineleukemia virusproteaseis encodedby thegag-pol gene and issynthesized throughsuppressionofanambertermination codon. Proc.Natl. Acad. Sci. USA82:1618-1622.
59. Yoshinaka, Y.,I.Katoh, T. D.Copeland, and S. Oroszlan. 1985. Translationalreadthroughof an amberterminationcoding dur-ing synthesis of feline leukemia virus protease. J. Virol. 55:870-873.
60. Yoshinaka, Y.,I. Katoh, T. D.Copeland, G. W. Smythers, and S.Oroszlan. 1986.Bovineleukemia virus protease: purification, chemical analysis, and in vitro processing of gag precursor polyproteins.J. Virol.57:826-832.