Copyright © 1988,AmericanSocietyforMicrobiology
Identification and Characterization of
a
DNA Primase
Activity
Present in Herpes
Simplex
Virus Type 1-Infected HeLa Cells
ANDREW M. HOLMES,* STEVEN M. WIETSTOCK, ANDWILLIAM T. RUYECHAN
Department ofBiochemistry, Uniformed Services University of the Health Sciences, 4301 Jones Bridge Road,
Bethesda,
Maryland
208144799 Received 31 July 1987/Accepted 1 December 1987A novel DNA primaseactivity has beenidentified inHeLa cells infected withherpessimplex virus type 1 (HSV-1). Suchanactivity hasnot been detectedinmock-infected cells. Theprimase activitycoeluted witha
portion of HSV-1 DNA polymerase from single-strandedDNAagarosecolumnsloaded withhigh-saltextracts derived from infected cells. This DNA primase activity could be distinguished from host HeLa cell DNA primase by several criteria. First, the pH optimum of the HSV primasewasrelativelybroad andpeakedat8.2 to8.7pH units. Incontrast,thepH optimum of the HeLaDNAprimasewasverysharp and fell between pH
7.9and 8.2. Second, freshlyisolated HSV DNAprimasewasless salt sensitive than the HeLaprimaseandwas
elutedfrom single-strandedDNAagaroseathighersaltconcentrations than the hostprimase. Third,antibodies raisedagainstindividualpeptidesof the calfthymusDNApolymerase:primase complexcross-reacted with the HeLa primase but didnot react with the HSV DNA primase. Fourth, freshly prepared HSV DNA primase appeared to be associatedwith the HSV polymerase, but after storage at4°C for several weeks, the DNA primase separatedfrom the viral DNA polymerase. Separation or decouplingcould also be achieved by gel
filtration of the HSV polymerase:primase. This free DNA primase had an apparent molecular size of approximately 40 kilodaltons, whereasfree HeLaDNA primase hadan apparent molecular size of
approxi-mately 110 kilodaltons. On the basis of these data, we believe that the novel DNA primase activity in HSV-infected cellsmaybe virus coded and that thisenzymerepresentsa newand important function involved
in thereplication ofHSV DNA.
The isolation ofa virus encoded DNA polymerase from
herpes simplextype1(HSV-1)-infected cells (21, 22, 35)was
an important first step toward understanding viral DNA
replication. The DNA polymerases isolated from HSV-1-and HSV-2-infected cells show very similarproperties (26)
whichservetodistinguish the viralenzymefrom mammalian
DNApolymerases. These properties include salt optimum, inhibition by polyamines (such as spermine), inhibition by phosphonoacetate and phosphonoacetate analogs, column chromatographic behavior, sedimentation rate on sucrose
gradients,andinhibitionby 9-(2-hydroxyethoxymethyl)GTP and 9-(1,3-dihydroxy-2-propoxymethyl)GTP (10, 11, 26).
The presence of a 3' -> 5' exonuclease activity (9, 26, 35)
also serves to distinguish HSV DNA polymerase from mammalian DNA polymerases ox, 3,and-y.The size of HSV
DNApolymerase wasoriginallyestimatedtobe 180
kilodal-tons (kDa) (35), but apolypeptide with a molecular size of approximately 140 kDa has nowbeenassigned tothe DNA polymerase activity (26), and thegene for this polypeptide has been cloned and sequenced (28).
Althoughthe propertiesof HSV DNApolymerase distin-guishit from mammalian DNApolymerases, oneimportant
similarity exists between HSV DNA polymerase and
mam-malian DNA polymerase a., the enzyme believed to be
responsible forthebulkof the cellular DNA replication (see reference 16 for review). Thatpropertyis the abilitytouse a
short piece of RNAas aprimer for DNA synthesis (35). A
DNAprimasecanbecopurified from eucaryotic cells in tight association with DNA polymerase a (6, 14, 20, 34). This
primasecansynthesize oligoribonucleotides of 8to 10bases inlength, whichthe DNApolymerase canthenusein DNA
synthesis (7, 17, 32). Nascent DNA chains extracted from
*Corresponding author.
mammalian cells contain oligoribonucleotides of this size at
their 5' ends (29, 33).
Mature HSV DNA molecules contain alkali-labile sites (12, 13, 31, 36). Thesemaybe duetotheuseof RNA primers
for DNA replication, some of which may have persisted during maturation of the DNA, although alternative expla-nations for alkali lability and the presence of RNA-like material inmatureviral DNAarepossible (3,25, 38). Newly synthesized HSV DNA has been demonstrated to contain RNAcovalentlylinkedtoDNA(2, 15, 24) whichis removed
over a period of time (24). These RNA segments are esti-matedtobe 35 nucleotides inlengthandarelinkedtoDNA averaging 36 x 103 nucleotides in length (24). This would indicate amajordifference between the waythe virususes
RNA toprimeDNA synthesis and thewayeucaryoticcells replicate. The lengthof the DNA attachedtoRNAprimers in the eucaryotic cell is usually 200 nucleotidesor less (8). These differences may reflect the involvement of virus-codedproteins in the synthesis of the RNAprimers aswell
as inthesynthesis of the viral DNA.
We report hereonthe identification and characterization ofanenzymewhichcanbeclassifiedas aDNAprimasefrom HSV-1-infected HeLa cells and whoseenzymaticand
struc-tural properties distinguish it from the DNA primases iso-lated from uninfectedHeLacells.
MATERIALS AND METHODS
Chemicals and reagents. Calfthymus DNA and unlabeled deoxynucleoside and nucleoside triphosphates were ob-tained from Sigma Chemical Co. [8-3H]dATP, [methyl-3H]TTP,and [a-32P]ATPwere obtained fromNewEngland Nuclear Corp. Escherichia coli DNA polymerase I was
obtained from New England BioLabs, Inc. Sepharose 6B
was obtained from Pharmacia Fine Chemicals.
Single-1038
on November 10, 2019 by guest
http://jvi.asm.org/
stranded DNA agarose was obtained from Bethesda Re-search Laboratories. Tissue culture media were obtained from GIBCO Laboratories.
Poly(dT)3500
was prepared by using terminal deoxynucleotidyltransferase as described elsewhere (5). Calf thymus DNA polymerase:primase com-plex and free calf thymus DNA primase were prepared as described previously (6). Antibodies were prepared by im-munizing individual rabbits with individual peptides from the calf thymus DNApolymerase:primase complex as described elsewhere (18). They were further purified by chromatogra-phy on protein A-Sepharose and then affinity purified by chromatography on a calf thymus DNApolymerase:primase complex Sepharose 4B column. All otherchemicals were of reagent grade.Cells and viruses. HeLa S-600cells were grownin suspen-sion culture in suspensuspen-sion-modified Eagle medium to a concentration of 1.6 x 106/ml. Infection was with HSV-1 (mP) grown in Vero cells at a multiplicity of 20 PFU per cell as described by Derse et al. (10). Cells were harvested 8 h after infection, the time viral DNA replication reached its maximum under these conditions. The cells were harvested by centrifugation and stored at -70°C or used immediately for the isolation of HSV DNApolymerase.
Isolation of HSV DNA polymerase:primase. The isolation procedure was based on a procedure described by Powell and Purifoy (27) and Ruyechan and Weir (30). The cells were suspended in extraction buffer (20 mM Tris [pH 7.5], 1.7 M KCI, 5 mM EDTA, 1 mM dithiothreitol [DTT], 0.2% Noni-det P-40, 10% glycerol), sonicated three times for 30 s in a Heat Systems Ultrasonics sonicator, andcentrifuged at 500 x g, and the supernatant was subjected to a 10% polyethyl-ene glycol phase separation. The precipitated material, in-cluding nucleic acids, was spun down. The supernatant was dialyzed against 0.15 M KCI in the buffer described above and chromatographed on a single-stranded DNA agarose column. Enzyme were eluted with a linear 0.15 to 1.0 MKCI gradient. All procedures were carried out at 4°Cin asolution containing 20 mM Tris (pH 7.5), 1 mM EDTA, 1 mMDTT, and 10% (vol/vol) glycerol. Amodified procedure ofAllenet al. (1) has also been used to isolate DNA polymerase from HSV-1-infected HeLa cells. In thisprocedure, the cells were lysed in hypotonic buffer, brought to 0.4 M potassium phosphate, as described above, and passed through DEAE-Tris-acryl. The unbound material was dialyzed against a solution containing 20 mMpotassium phosphate (pH7.5), 1 mM DTT, 20% (vol/vol) glycerol, and 0.2% Nonidet P-40 and rechromatographed on DEAE-Tris-acryl. HSV DNA polymerase bound to the column was eluted with a linear
KCI
gradient (0 to 0.5 M). Fractions containing DNA poly-merase:primase assayable at 0.2 M salt were pooled, dia-lyzed against the above buffer, and chromatographed on a single-stranded DNAagarose column underidenticalelution conditions.In some instances, the DNA polymerase:primase was concentrated by adsorption and batch elution with 0.5 M KCI from a 1-ml single-stranded DNA agarose column and stored at 4°C.
Enzymeassays. DNA polymerase activities were assayed by using three different protocols. DNApolymerase eluted from single-stranded DNA agarose columns to which high-salt extracts had been applied werecarried out in asolution containing 50 mM HEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid; pH 8.0), 8 mM MgCl,, 300
pg
of DNase I-treated DNA per ml, 0.2 mM each dATP, dCTP, and dGTP, 0.04 mM [3H]TTP at 372 dpm/pmol of 2 mM 2-mercaptoethanol, and 200 mM KCI. Aliquots (10 LI) ofindividual column fractions were added to 40 ,ul of the reaction mixture andincubatedat
35°C
for 20 min.Aliquots (40IL)
werespotted
onglass
fibersquaresandplaced
in5% trichloroaceticacid-1% sodiumpyrophosphate.
The squares werewashedtwice with1 NHCl andoncewith95%ethanol,
airdried, and counted in a
liquid
scintillation counter. One unit ofDNApolymerase
activity
in this assayis defined asthe
incorporation
of 1pmol
of[3H]TMP
into insoluble material in 1 minat35°C.
DNApolymerase
isolatedby
the modifiedprocedure
ofAllenetal.(1)
wasassayed
in 40 mM phosphatewithorwithout200 mMKCl.All otherconditions wereexactlyasdescribedabove.Aftertheinitialisolation of HSV DNApolymerase
activity
by
these twomethods,
all additionalpolymerase
assays werecarried outin a solutioncontaining
40 mMpotassium
phosphate
(pH 7.0),
8 mMMgCl2,
100 ,ugof bovineserumalbumin(BSA)
perml,
1 mM DTT, 100 ,ugofDNAase I-treated DNA perml,
0.1 mMeach dATP,dCTP,
anddGTP,
and 0.1 mM[3H]TTP
at 40to 50cpm/pmol,
with or without 0.15 M NaCl.Assays
wereincubatedat
35°C,
andat thetimesindicated,
samples
werespotted
onglass
fibersquaresandplaced
in the5% trichloro-acetic acid-1% sodiumpyrophosphate.
One unit of DNA polymeraseactivity
isdefined astheincorporation
of1nmol of[3H]TMP
into acid-insoluble material in 60 minat35°C
in this assay.DNA
primase
wasassayed
directly
ina solutioncontain-ing 50 mM Tris
(pH
8.0),
8 mMMgCl2,
100 ,ugofBSA per ml, 1 mMDTT,
10,uM
poly(dT)3,500,
and 0.1 mM[a-32P]ATP
(100to 200cpm/pmol). Assays
were done at35°C,
and
aliquots
werespotted
onWhatmanDE-81 papersprevi-ously
spotted
withanequal
volumeof5mMunlabeledATP. Thistechnique
reducescontaminationby
the[32P]ATP.
The papers were washed four times with 0.2 Mdipotassium
hydrogen
phosphate
andtwice withglass-distilled
waterand counted inglass-distilled
water in aliquid
scintillationcounterset onthe
3H-labeled
channel.Thecountsremaining
onthe paper
give
a direct measurement of oligoribonucleo-tidesynthesis.
One unit ofenzymeactivity
is definedastheincorporation
of 1 nmol of[32P]AMP
into oligoribonucleo-tide in 60 minat35°C.
DNA
primase
wasroutinely
assayed
by
anindirect assay. Theassayconditionswere asdescribedabove,
except
that 2 mMunlabeledATPreplaced
the 0.1mM[at-32P]ATP
and 0.1 mM[3H]dATP
at40to50cpm/pmol
and 20 UofE.coli
DNApolymerase
I per ml wasusually
present.
Assays
werecarriedout at
35°C,
andaliquots
were spottedonglass
fiberpaper and
processed
as for DNApolymerase
assays. Thiscoupled
assayprocedure
measures the extension ofprimers
synthesized
by
the DNAprimase,
and the rate of incorpo-rationof[3H]dAMP
intoacid-insolubleproduct
isdependent
on the rate of
primer
synthesis.
In the absence ofATP orDNA
primase,
noincorporation
of[3H]dAMP
is observed. This isanamplification
of DNAprimase
activity.
One unitof enzymeactivity
in this instance is defined as the incorpora-tionof 1 nmol of[3H]dAMP
intoacid-insoluble
material in 60 min at35°C.
Exonucleaseactivity
was measured in asolu-tion
containing
50 mM Tris(pH 8.0),
8 mMMgCI2,
1 mM DTT, 100,ug of BSA perml,
and 10,ug ofsingle-stranded
"4C-labeled
T7 DNA at 30 x 103cpm/,ug.
Assays
wereincubatedat
35°C,
and at the timesindicated,
aliquots
wereplacedon
glass
fiber squares and the squareswereprocessed
as for DNApolymerase
assays.Sepharose 6B
chromatography.
Sepharose
6Bchromatog-raphy
was carried out on an85-by-1.4-cm
column in asolution
containing
0.25 MNaCl,
50 mM Tris(pH
8.0),
100 ,ugofBSA perml,
1 mMDTT,
and 20%(vol/vol)
glycerol.
on November 10, 2019 by guest
http://jvi.asm.org/
Fractions (50 drops, or 1.7 ml, each) were
column was calibrated with calf thymus
catalase, rabbitIgG,E. coliDNApolymeraseI,
cytochrome c, and DTT.
Antibody-binding experiments. Antibody-binding
mentswerecarriedoutasfollows. Atotal p.l
enzyme preparation and 20,ul ofrabbit
(IgG) raised against a specific calf thymus
peptide (68 kDa,55kDa,or48kDa)inTBS
Tris, 0.15M NaCl, 2 mg of BSA per ml)
incubated for 18 h at 4°C. Samples RI
removed, and to each sample was added plI 50%
(vol/vol) protein A-Sepharose in TBS
continued for an additional 18 h. The
spundown inan Eppendorfcentrifuge, 5-pLI
the resulting supernatants were assayed
polymerase activity asdescribed above.
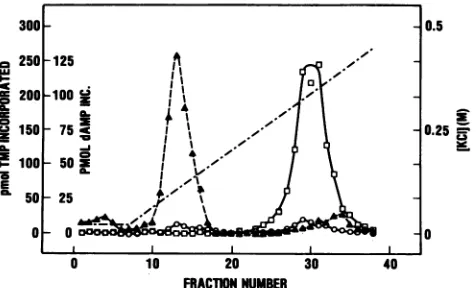
RESULTS Association of DNA primase with HSV
Whenextractsof HSV-1-infectedHeLa
graphed on single-stranded DNA agarose,
polymerase activity eluted between 0.3
With the indirect,orcoupled,DNAprimase
DNAprimase activity was detected toward
HSV DNA polymerase peak (Fig. 1).
primase assay also detected someDNA
this area of the elution profile (data
difficult to quantitate the DNA primase
fractionsby thedirectprocedure, since
[32_p]AMP incorporation, it was necessary
assaytime far beyondthatshown inFig.
of high levelsofexonuclease activitydestroyed
atlater times. Thedirectassaywasalso
presence ofaDNA-dependentATPase
H activity inthisareaoftheelutionprofile
The direct DNAprimaseassaycontained
whereas the indirect, or coupled, assay
ATP, helping to negate the effects of
ATPase. Further,thecoupledDNAprimase
theDNAprimaseactivity tosuch anextent
and the amount ofthe DNA agarose
500
=9 C1
C2
=c
E
400
300
200
100
0l
0 5 10 15
FRACTION NUMBER
20 25
FIG. 1. Chromatography ofHSV DNA
stranded DNAagarose. HSV DNA polymerase
infectedcells by themodifiedPowelland
AssaysweredoneasdescribedinMaterials
O,
DNA polymerase in the presence 0,primase in the coupled assay in the coli
polymeraseI ; A,exonuclease;
assay could be reduced so that neither
ATPase affectedthe assay. Theincorporation [3H]dAMP
shown in Fig. 1 was taken from time courses
individual fractions in which the incorporation
linearwith time. The resultshowninFig. 1
position of elution ofthe DNAprimase was
the contaminating enzyme activities. That
primase elution position was toward the
polymerase elution position. The fractions
primase activity were pooled, andthepooled
referred to as HSV DNA primase.
Uninfected HeLacells were subjected tothe
tion and chromatography procedures.
activity present in controlextracts did not
agarose column. This DNA polymerase itedby 0.15 M NaCI intheassay and could
in the absence ofsalt.This salt-inhibited
was also present in the DNA agarose column
of the infected cell extracts. The properties
polymerase are consistent with those of
polymeraseao. On application of the KCl
control (uninfected) DNA agarose column,
of DNA primase began to elute from
continued to elute uptoabout 0.4 MKCl (data shown).
This material, comprising less than 10%
eluting from the infected cell DNA agarose
pooled forenzymecomparisonpurposesand
control DNAprimase. This relatively small
primase activity identified in uninfected suggest that the much greater amount
observed in this areaof the elution profile
infected cells were subjected to the same
consequenceoftheviralinfection, although
that it is necessarily virally encoded.
By using the modified procedure of
isolate DNA polymerase from infected
tion was achieved between the HeLaDNA
the HSV DNA polymerase by the KCI
second DEAB-Tris-acryl column. The
polymerase was pooled, dialyzed, and
stranded DNA agarose as indicated in
ods. The elution profile from this DNA
shownin Fig.2. Thesalt-sensitiveHeLa ot
eluted between 0.1 and0.15 MKCI. Thesalt-activated
DNApolymerase again elutedbetween KCI.
Two peaks of DNA primase activity
major peak wasassociatedwith the HeLa
cx and the minor peak eluted at the rear
polymerasee
peak. The first peak of DNAwaspooled forenzyme comparison studies
as the HeLa DNA primase.
When this second purification procedure
uninfected HeLa cells, a salt-sensitive
meraseactivity was observed againeluting
0.15 M KCI. Salt-activated DNA polymerase not observed in this elution profile (data
peak of DNA primase activity was observed,
elution positionasthe DNApolymerase,
of DNA primase smeared off the column
concentration ofabout0.4M KCI. Again,
10% of the amount of enzyme activity
infected cell extracts were subjected RNApolymerase wasnotdetected with
procedure.
Initial characterization of the HSV
relationships betweenthedifferentDNA
100 80
40
1-20 ... .20 25
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.68.304.505.648.2]20 0 23a1
la.2ChoaorpyofHVDAplmeaeo aige
meras in~the prsnc f0.MNCl ,DN5oymrsih
z
CD~~I
~100
501 .EI
50 -25
0 10 20 30 40
FRACTION NUMBER
FIG. 2. Chromatography of HSV DNA polymerase on single-stranded DNAagarose. HSV DNApolymerase was isolatedfrom infectedcellsby themodified procedure of Allenet al. (1). Assays
were doneas described in Materials and Methods. 0, DNA
poly-meraseinthepresenceof 0.2MNaCl; 0, DNApolymerasein the absence of NaCl; A, DNA primase in the coupled assay in the
presenceofE. coliDNApolymerase I; .--, KCIgradient.
infected and uninfected cell extracts were
explored.
The effect of increasingconcentrations ofNaCl on the HSV and HeLa DNA primases was determined by using the coupled assayin the presenceofE. coli DNA polymerase I(Fig. 3). It is important to note that these primase samples also containedHSV and HeLa DNA polymerases, respectively. Freshly prepared HSV primase was less sensitive than the HeLa DNAprimase, whichshowed the same saltsensitivity as the DNA primase activity of the calf thymus DNA polymerase:primase complex. E. coli DNA polymerase I wasactive throughout therangeof salt concentrations used. SinceHSV DNApolymerase isasalt-activated enzyme(26, 35), however, the relative salt resistance of the HSV DNA primasecould have been due to the HSV polymerase begin-ning to function at high salt concentrations to efficiently elongate a reduced number of primers synthesized by a salt-sensitive DNAprimase. Accordingly,acomparisonwas madeofDNAprimase activities in the presence and absence of0.15 M NaCl and with and without the additionofE.coli125
100
75
5'
25
0
I
<:0~~~~~~~~~~~
0 .025 .05 .075 .1 .125 .15
[NaCq
(M)FIG. 3. Effects ofincreasing NaCl concentration on DNA pri-mase activity. DNA primase was assayed in the coupled assay
systemin thepresenceofE.coli DNApolymeraseI.0,HSV DNA
primase; A, HeLa DNA primase; 0, DNA primase of the calf
thymusDNApolymerase:primase complex.
TABLE 1. Effects of the additionof 0.15M NaClonthe activitiesof HeLa DNA primase and freshly
prepared HSVDNAprimase
Picomoles of[3HldAMP incorporated
per5 ,ul per60min
WithE.coli DNA WithoutE.coli polymeraseI DNApolymeraseI
No 0.15 M No 0.15 M
NaCl NaCl NaCl NaCl
HeLa 55 1 25 0
Freshly prepared HSV 180 72 105 33
DNA polymerase (Table 1). Inthe absence of bothE. coli DNA polymerase I and NaCI, the elongation of primers synthesizedby both the HeLa andHSV DNAprimases by the respective DNA polymerases was relatively efficient comparedwith theelongationof theprimersin the presence of E. coli DNA polymerase I (Table 1). At 0.15 M NaCl, there was no incorporation of [3H]dAMP with the
14eLa
DNA primase either in the presence or absence ofE. coli DNApolymeraseI, whereas therewasstillincorporationin both instanceswith theHSV DNA primase. Table 2 shows the activities of the DNA polymerase at the two salt con-centrations.
These results were obtained by using freshly prepared HSV DNA primase. The salt profile shown in Fig. 3 was obtainedwith HSV DNAprimasewhich had been isolateda few days before. HSV DNA primase stored at 4°C for severalweekswasalmostas salt sensitive as the HeLa and calfDNA primases were
(Table
3). Furthermore, although both the DNA polymerase and DNA primase activities of this stored enzymesamplehadnotdeclinedsignificantly (the DNA primaseactivity beingmeasuredin the presenceofE. coli DNA polymerase I by the indirect assay), the HSV DNA polymerase could now not elongate the primers pro-ducedbythe DNAprimaseineither the presenceorabsence of0. 15 MNaCl, indicatingamajor changein therelationship ofthe DNA polymerase and DNA primase in this sample. This change could be the decoupling of the two enzyme activities or the loss ofanother factor which mediates the two activities. This change in the salt sensitivity and the inability of the HSV DNA polymerase to function in the coupledDNAprimaseassayhasbeenobservedonlong-term storageofthe enzyme at4°C
orafterdialysis ofthe enzyme against50% (vol/vol) glycerol and storage of the enzyme at -20°C. Thus, these dataindicate that the relative insensitiv-ityto saltdisplayed by freshlypreparedHSVDNAprimase is not an intrinsic property of the primase but may also indicate its participationinacomplex. The uncoupledHSV DNApolymeraseis stillcapableofincorporating[3H]dAMP into product by using a poly(dT) template when given anTABLE 2. Effects oftheaddition of0.15 MNaClonthe DNA
polymerase activities of HeLaDNAprimaseand
freshly preparedHSV DNAprimase
Picomolesof[3H]dTMP
incorporatedper 5p.l
Originof DNApolymerase per 60min NoNaCl 0.15 M
NoNaCi NaCI
HeLaDNAprimase 73 10
Freshlyprepared HSV DNAprimase 92 720
21-t=
210
p
C.3
dc -e
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.63.301.64.208.2] [image:4.612.61.301.504.671.2]TABLE 3. Effects ofaddition of 0.15 M NaCl on the activities
of freshlyprepared HSV DNA primase and concentrated HSV DNA primasestored at 4°C for 35 days
% Activity
WithE.coliDNA WithoutE. coli HSVDNAprimase polymeraseI DNApQlymeraseI
No 0.15M No 0.15 M
NaCl NaCl NaCl NaCl
Freshlyprepared looa 64 76 60
Concentrated and stored 100b 3 5 3
aAnactivity of 100% for freshly prepared HSVDNAprimasewas590pmol of[3]dAMPincorporatedper 20[LIper 60min.
bAnactivity of100%forconcentrated stored HSVDNAprimasewas152
pmol of[3H]dAMPincorporatedper 1RI1per 60min.
oligo(A) initiator (data not shown), indicating that this loss of coupling ofDNA polymerase andprimase activities is not duetothe DNA polymerase losing the capability of using an oligoriboadenylate initiator.
The pHoptima of both freshlyprepared and stored HSV DNAprimase were the same, pH 8.2 to 8.7 (Fig. 4A and B). Those of the HeLa DNA primase and the DNA primaseof the calfthymus DNApolymerase:primasecomplex andfree DNAprimasefrom calf thymus were all pH 7.9 to 8.2(Fig. 5A and B). The control DNA primase (the DNA primase fromthe DNA agarose column chromatography of extracts of uninfected HeLacells) did not show a defined pH opti-mum over the pH range studied but rather indicated a gradualincrease in activitywith increasing pH (Fig. 4B).
[image:5.612.59.296.96.193.2] [image:5.612.320.556.446.609.2]Size oftheHSV DNA primase. Theresults of the chroma-tography of a stored (uncoupled) sample of HSV DNA primaseon acalibrated Sepharose 6B columnareshown in Fig. 6. The HSV DNA polymerase activity eluted in advance of beef liver catalase with an apparent molecular size in excess of250kDa, compared withthemolecularsizeof140 kDa obtainedfromsedimentation studiesandpredictedfrom the DNA sequence (28). Allen et al. (1) have previously made a similar observation with the equine herpesvirus DNA polymerase. This discrepancy in the molecular size
estimnates
is explicable on the basis that the molecule is asymmetric in shape (19).TheHSV DNAprimaseactivityeluted between the posi-tions of elution of ovalbumin and cytochrome c with an
150 125
i 100
Z 75
X50
2 5
oc 25
0
7.5 8 8.5 9 9.5 7.5
[image:5.612.63.301.522.672.2]pH
FIG. 4. The pH optima of HSV DN.
wasassayed in the coupledassaysystem DNA polymerase I in constant-molarit Stored HSVDNAprimase (O); freshlypr
(0); control DNA primase (A).
57@= _ Z75G
F - 500A
150
.100
2L 2501EC
50-0F
07.5 8 8.5 9 9.5 7.5 8 8.5 9 9.5
pH
FIG. 5. The pH optima of mammalian DNA primases. DNA primasewasassayed in the coupledassayinthepresenceof E.coli DNA polymerase I in constant-molarity Tris buffers. Symbols: HeLa DNA primase (O); calf thymus DNA polymerase primase complex (0); free calf thymus DNA primase (A).
apparent molecular size of approximately 40 kDa (Fig. 6). The material eluting between fractions 40 and 50 was not present in chromatography of other samples ofHSV DNA primase on the Sepharose 6B column and may represent aggregated material. Most of the DNA primase activity of freshlypreparedHSVDNAprimasealsoelutedfrom
Seph-arose6Bwith anapparent molecular sizeof 40 kDa. These dataindicatedthat(i) either theprimase activitywas
uncou-pled fromthe HSV DNA polymerase activityon long-term storageoruncouplingoccurred duetodilutionduring chro-matographyor(ii)itwasnotpartofatight complextobegin with. Most of the HeLa DNA primase eluted at the same
volume as E. coli DNA polymerase I, with an apparent molecular size of 110 kDa (Fig. 6). This result seems to
indicate that most of this DNA primase is also not tightly
1 234 5
111 1
125
LU
I--:f g
40
Is
E
30 b
co
20
IC
in
6 7
100-
75-50[
251
30 40 50 60 FRACTIONNUMBER
70 80
a FIG. 6. ChromatographyofHSV DNAprimaseonan
86-by-1.4-cm Sepharose 6B column. Stored HSV DNA primase was
chro-matographedon Sepharose6Basdescribed in Materials and Meth-ods. Assays were done as described in Materials and Methods.
8 8.5 9 9.5 Symbols: l, DNApolymerase activity in the presence of 0.15 M
NaCl;0,DNAprimase activityin thecoupledassayin thepresence
of E.coliDNApolymeraseI.The numbers refertostandardsrunon
A primase. DNA primase thecolumn under thesameconditions:1, Calf thymus DNA; 2,beef
inthepresenceofE. coli liver catalase; 3, rabbit IgG; 4, E. coli DNA polymerase I; 5,
ty Tris buffers. Symbols: ovalbumin; 6,cytochromec;7,DTT.Thearrowmarksthepeakof
epared HSVDNAprimase elutionof thefree HeLaDNAprimase rununder thesame
condi-tions.
A A~~~~~~~
L
i
on November 10, 2019 by guest
http://jvi.asm.org/
associated with DNA polymerase under these elution con-ditions. It is important to note that free calf thymus DNA primase elutes at the same position and sediments with the sameS value as E. coli DNA polymeraseI, indicating that it is a globular molecule with an approximate molecular size of 110 kDa (18). This does not necessarily indicate that the apparent molecular size of 40 kDa observed for the HSV DNAprimase is the true molecular size of this enzyme (see Discussion).
Reactivity with anti-calf DNA primase IgGs. Rabbit IgGs raised against individual peptides of the calf thymus DNA polymerase:primase complex and previously used to assign functions to these individual peptides (18) were used in antibody-binding experiments to ascertain the relationship of the HSV, HeLa, and calf DNA primases. Previously, by using both an immunoblotting procedure and enzyme neu-tralization, peptides of 55 and 48 kDa had been assigned to thefreeDNAprimase ofcalf thymus and peptides of 68 kDa, 55kDa, and 48 kDa had been assigned to the DNA primase of the calf thymus DNA polymerase:primase complex, with the 48-kDa peptide being derived, perhaps proteolytically, from the 68-kDa peptide (18). The results of the antibody-binding experimentsarepresented in Table 4. All threeIgG samples reacted with the HeLa DNA primase, but none of themreacted with the HSV DNAprimase. Threetimes the amount of calf thymus DNA primase showed the same reactivity with these antibodies as the HeLa enzyme did. Thus, although cross-reactivity of these antibodies with the HeLa DNA primase was not 100%, the results show a significant difference between the HeLa and HSV DNA primases. Immunoblots with the anti-calf DNA primase antibodiesshowedcross-reactivitybetween theseantibodies and the HeLa DNA primase and indicated that the HeLa DNA primase has a peptide composition similar to that of the calf DNA primase. No cross-reactivity with the HSV DNA primase wasobserved (datanot shown).
DISCUSSION
[image:6.612.60.299.603.680.2]A new DNA primase activity has been identified in ex-tractsderivedfrom HSV-1-infectedHeLacells. Thisactivity appearstobecoupled with the HSV DNApolymerasewhen freshly isolated, but it appears to decouple from the poly-merase activity upon long storage and is separable by gel filtration chromatography. Severalpropertiesof thisprimase activity distinguish it from the host HeLa cellprimase. First, freshly prepared HSV primase retains approximately 50% activityatsaltconcentrationsupto125to150mM,although this salt resistance is lost when the primase is uncoupled
TABLE 4. Percentage of DNAprimase activity remaining in solution after treatment withanti-calfpeptide IgGsandafter binding of the antibody-antigen complextoprotein
A-Sepharose"
%Activitywith antibodiesagainst Enzyme the following calf peptides:
68 kDa 55kDa 48kDa
CalfthymusDNApolymerase: 41 74 25
primase complex
HeLa DNAprimase 54 73 31
HSV DNAprimase 96 96 118
a An activityof100%wasthatobtained aftertreatmentof enzymes with controlIgGand was34, 13,and 14pmolof[3H]dAMPincorporationper 5 ,ul per 60minforthecalfthymusDNApolymerase:primase complex,the HeLa DNA primase, and HSV DNA primase, respectively, with E. coli DNA
polymerase I presentin the assays.
from the polymerase. In contrast, HeLa primase is ex-tremely salt sensitive both when it is coupled to the host polymeraseand when it is free. Second,thepHoptimumfor the HSV primase is higher and much broader than that determined for the HeLa primase. Third, the apparent molecularweight of the HSVprimaseas determined by gel filtration isapproximatelyone third that determinedforthe HeLaprimase. Fourth, antibodiesraisedagainstsubunits of the calf thymus DNA primase complex which bind to the HeLaprimaseactivitydonotshow anycross-reactivitywith theHSVprimase.
The question then arises as to the origin of this primase activity. Severalpossibilitiesmust beconsidered. First, the putativeHSV
primase
couldsimplybe HeLaprimasewhich hasbecome associated with the HSVDNApolymerase
and is used in viral DNAreplication. Such anassociationcould alter the pH optimum and saltsensitivity
of the HeLa primase. Thefact that thepHofuncoupled
HSVprimase
is the same as that offreshly prepared (coupled) HSV DNA primase would argue that thisactivity
is not the HeLa primase.Thisfact,inconjunctionwith thedifferentapparent molecularweightsdetermined for thetwoprimase
activities by gelfiltration,
appears to eliminate thispossibility.
Sec-ond,
the HSVprimase
could be a modified(proteolytic
fragment) ofthe HeLa
primase.
Inthis case, the salt andpH
optima
of the enzyme couldagain
be alteredsignificantly
and would not necessarily change upon decoupling ofthe pri-mase from thepolymerase. Theevidencewe havegathered
argues
against
thispossibility.
Thecross-reactivity
of anti-bodies to thepeptides
of the calf DNAprimase
with the HeLa DNAprimase
and the elutionposition
ofthe HeLa primasefromSepharose6B suggestastructuralsimilarity
of the DNAprimases
from these two sources. The free calf DNAprimase
has beendemonstratedtocontainpeptides
of 55 and 48 kDa(18).
If the size of the HSV DNAprimase
is indeed 40 kDa, it ispossible
that it is derived from thecatalytic
subunit of the HeLaprimase.
In this case, how-ever,itwould beexpected
thatsomeepitopes
recognized by
thesepolyclonal
antibodies raisedagainst
the calf DNAprimase
would be retained and that somereactivity
with these antibodies would have been observed with the HSV primase. No suchreactivity
was observed.Thus,
while wecannot
absolutely
ruleoutthissecondpossibility,
webelieve itto beunlikely
on the basisof the above lineofreasoning.
A third
possible
source of the HSVprimase
could be mitochondrial DNAprimase,
which is solubilized and be-comesfortuitously
associated with HSV DNApolymerase
during
the extractionprocedure.
Thispossibility
also does not seemlikely.
TheratlivermitochondrialDNApolymer-ase hasa
pH
optimum
of9.3,
with very littleactivity
below pH8.5(23).Further,the humanmitochondrialDNAprimase
hasbeen showntobe boundto
RNA,
which,
whenremoved,
leaves an enzyme that sedimentsatthe same rate astheE. coli
polymerase
Ilarge
fragment (37).
If theprotein
isspherical,
it wouldcorrespond
to a molecular size ofabout 70kDa. In thelight
of theseconsiderations,
the control DNAprimase
we have observed may represent a mixture of mitochondrialDNAprimaseandHeLa DNAprimase
onthe basis of the lack of apH
optimum
(Fig. 4B)
and a small amount ofreactivity
with the antibodies raise'dagainst
the calfDNAprimase peptides (data
notshown).
Thefinal
possibility
tobeconsidered is that the HSV DNA primaseactivity
is in fact a virus-coded enzyme. Thecou-pling
of theprimase
withtheviralDNApolymerase
in fresh extracts,theunique pH
profile
of itsactivity,
and its lack ofcross-reactivity
with antibodiesagainst
the calfprimase
allon November 10, 2019 by guest
http://jvi.asm.org/
argue that this may be the case. Challberg (4; personal communication)hasrecentlydemonstrated that thereare at least seven openreadingframes presentonthe HSV genome whose products are essential for the replication of HSV DNA.Thegeneproducts potentiallyencodedbythese open reading frames range in size from 140 to 51 kDa. The two largestopenreadingframescorrespondto the genesforthe viral DNA polymerase and the HSV major DNA-binding
protein,
ICP8. The geneproducts from theother five open reading framesarealllargerthan the apparentmolecularsize of40kDa which wehavedetermined forthe HSVprimase.
Thiswouldimplythat if the HSVprimase isencodedbyone ofthese openreading frames,theactivitywhichwedetected represents a posttranslationally modified protein. Whether this modification is required for the activity of the HSV primaseorrepresents nonspecific proteolytic degradationof theprimase during extraction and partial purification ofthe enzyme is notknown.
Furtherexperiments needto be carriedout to purify the HSV DNA primase to homogeneity, to characterize its enzymatic activity in depth, and to demonstrate that the primase can synthesize RNA primers of the size seen in newly replicating HSV DNA, both in the presence and absence of HSV DNA polymerase and deoxynucleoside triphosphates. These last experiments would demonstrate theeffectsof the DNApolymeraseonprimer synthesisand theuseofsuchprimers bythe DNApolymerase. However, suchproductcharacterization is meaninglesswith the HSV DNAprimase at its present level ofpurification because of the presence of other activities, such as RNase H, which affect the size of the product or the ability of the DNA polymerase to use these products. Additional work also needstobe donetodefinitively show that the DNAprimase is partofanHSV DNApolymerase:primasecomplex andto discover other proteins which may be involved in holding enzymatically active peptides together or in coupling the DNA primase activity with the DNA polymerase activity. Theseexperiments areunder way.
ACKNOWLEDGMENTS
The expertassistance of Steven C.Maresinthepreparationand
chromatography of cell extracts is gratefully acknowledged. We
thankF. J. Bollum, L.M. S. Chang,and M.Challberg for helpful discussionsduringthecourseof this work.
This workwassupported by Public Health ServicegrantA122468 fromtheNationalInstitute forAllergy and InfectiousDiseases (to W.T.R.)and UniformedServices Universityof the Health Sciences grant R07156(toA.M.H.).
LITERATURE CITED
1. Allen, G. P., D. J. O'Callaghan, and C. C. Randall. 1977.
Purification and characterisation ofequine herpesvirus-induced
DNApolymerase. Virology76:395-408.
2. Biswall, N., B. K. Murray, and M. Benyesh-Melnick. 1974.
Ribonucleotidesin newly synthesizedDNAofherpes simplex virus. Virology61:87-99.
3. Blair, D.G., D.J. Sherratt, D. B.Clewell,and D. R. Helinski. 1972. Isolation of supercoiled colicinogenic factor E1 DNA sensitivetoribonucleaseandalkali. Proc. Natl. Acad. Sci. USA 69:2518-2522.
4. Challberg,M.D. 1986. Amethod foridentifyingthe viral genes
required for herpesvirus DNA replication. Proc. Natl. Acad.
Sci. USA83:9094-9098.
5. Chang,L.M.S.,and F.J.Bollum.1971.Enzymatic synthesisof oligodeoxynucleotides. Biochemistry10:536-542.
6. Chang,L. M. S., E. Rafter, C. AugI, and F. J. Bollum. 1984.
PurificationofaDNApolymerase:DNA primase complexfrom calfthymusglands. J. Biol. Chem. 259: 14679-14687.
7. Conaway, R. C., and I. R. Lehman. 1982. A DNA primase
associated with DNApolymerase ao from Drosophila melano-gasterembryos. Proc.Natl. Acad. Sci. USA 79:2523-2527. 8. DePamphilis, M., and P. Wasserman. 1980. Replication of
eukaryotic chromosomes: a close-up look at the replication
fork. Annu. Rev. Biochem.49:627-666.
9. Derse,D.,and Y.-C.Cheng. 1981.Herpessimplex virus type 1 DNApolymerase. J. Biol. Chem. 256:8525-8530.
10. Derse, D., Y.-C. Cheng, P. A. Furman, M. H. St. Clair, and G. B. Elion. 1981. Inhibition of purified human and herpes
simplexvirus-induced DNApolymerases by 9-(2-hydroxyethoxy-methyl)guanine triphosphate.J. Biol. Chem. 256:11447-11451. 11. Frank, K. B., J.-F.Chiou,andY.-C.Cheng.1984. Interaction of
herpes simplex virus-induced DNApolymerase with 9-(1,3-di-hydroxy-2-propoxymethyl)guaninetriphosphate. J. Biol. Chem. 259:1566-1569.
12. Frenkel, N.,and B. Roizman. 1972.Separation of the herpesvi-rus deoxyribonucleic acid duplex into unique fragments and intact strand on sedimentation in alkalinegradients. J. Virol.
10:565-572.
13. Gordin, M.,U. Olshevsky, H. S. Rosencrantz,and Y. Becker.
1973. Studies on herpes simplex virus DNA: denaturation properties. Virology 55:280-284.
14. Gronostajski,R.M., J.Field,andJ.Hurwitz.1984. Purification
ofaprimase activity associated withDNApolymerasea from
HeLacells. J.Biol. Chem. 259:9479-9486.
15. Hirsch,I., andV.Vonka. 1974.Ribonucleotideslinked to DNA ofherpes simplex virustype 1. J. Virol. 13:1162-1168.
16. Holmes,A.M.,F.J. Bollum,and L. M. S. Chang. 1983. DNA
polymerases ofeukaryotes: Isozymes. Curr. Top. Biol. Med.
Res.7:277-279.
17. Holmes,A.M.,E.Cheriathundam, F.J. Bollum, and L. M. S.
Chang. 1985. Initiation ofDNA synthesis by the calfthymus DNApolymerase-primase complex. J.Biol. Chem. 260:10840-10846.
18. Holmes,A. M.,E. Cheriathundam,F.J. Bollum,and L. M.S.
Chang. 1986. Immunological analysisofthepolypeptide struc-tureof calfthymusDNApolymerase-primase complex.J. Biol. Chem. 261:11924-11930.
19. Holmes,A.M.,I. P.Hesslewood,and I. R.Johnston.1974.The
occurrence ofmultiple activities in the high-molecular-weight
DNApolymerase fraction of mammalian tissues. Eur. J.
Bio-chem. 43:487-489.
20. Kaguni, L. S., J.-M. Rossignol, R. C. Conaway, and I. R. Lehman. 1983. Isolation of intact DNA polymerase-primase fromembryos ofDrosophila melanogaster. Proc. Natl. Acad. Sci. USA 80:2221-2225.
21. Keir,H. M., J. Hay, J. Morrison,andH.Subak-Sharpe. 1966.
Altered properties of deoxyribonucleic acid nucleotidyl-trans-ferase after infection of mammalian cellswith herpes simplex virus. Nature (London)210:369-371.
22. Keir,H.M.,H.Subak-Sharpe,W.I. H.Sheddon,D. H.Watson,
and P.Wildy.1966.Immunologicalevidence foraspecificDNA
polymerase produced afterinfectionby herpes simplex virus. Virology 30:154-157.
23. Ledwith,B.J.,S.Manam,andG. C. VanTuyle. 1986. Charac-terisation ofaDNAprimase fromratlivermitochondria.J.Biol.
Chem.261:6571-6577.
24. Muller,W. E.G.,R. K.Zahn, J. Arendes,and D. Falke. 1979. Oligonucleotideinitiators forherpessimplexvirus DNA synthe-sis in vivo and in vitro.Virology98:200-210.
25. Nass, M. M. K. 1969. Mitochondrial DNA II. Structure and
physicochemical propertiesof isolated DNA. J. Mol. Biol. 42: 529-545.
26. Ostrander, M.,Y.-C.Cheng.1980.Propertiesofherpessimplex type 1and type 2 DNA polymerase. Biochim. Biophys. Acta 609:232-245.
27. Powell, K. L., and D. J. M. Purifoy. 1977. Nonstructural proteinsofherpes simplexvirus. I. Purification oftheinduced DNApolymerase. J.Virol. 24:618-626.
28. Quinn,J. P.,and D.J. McGeoch. 1985. DNA sequence in the genome ofherpessimplexvirus type 1containingthegenes for DNApolymeraseand themajorDNAbinding protein. Nucleic
on November 10, 2019 by guest
http://jvi.asm.org/
Acids Res. 13:8143-8163.
29. Reichard, P., R. Eliason, and G. Soderman. 1974. Initiator RNA in discontinuous polyoma DNA synthesis. Proc. Natl. Acad. Sci. USA 71:4901-4905.
30. Ruyechan, W. T.,andA. C. Weir.1984.Interaction with nucleic acids and stimulation of the viral DNA polymerase by the herpes simplex virus type 1 major DNA-binding protein. J. Virol. 52:727-733.
31. Sheldrick, P., M. Laithier, D. Lando,andM. L. Ryhiner. 1973. Infectious DNA from herpes simplex virus: infectivity of double stranded and single stranded molecules. Proc.Natl. Acad. Sci.
USA70:3621-3625.
32. Tseng, B. Y., and C. N. Ahlem. 1982. DNA primase activity from human lymphocytes. Synthesis of oligoribonucleotides that prime DNA synthesis. J. Biol. Chem.257:7280-7283. 33. Tseng, B. Y., J. M. Erickson, and M. Goulian. 1979. Initiator
RNA of nascent DNA from animal cells. J. Mol. Biol.
129:531-546.
34. Wang, T. S.-F., S.-Z. Hu, and D. Korn. 1984. DNA primase fromKB cell. J. Biol. Chem.259:1854-1865.
35. Weissbach, A., S.-C. Hong, J. Aucker, and R. Muller. 1973. Characterisation of herpes virus-induceddeoxyribonucleic acid polymerase. J. Biol. Chem. 248:6270-6277.
36. Wilkie, N. M. 1973. Thesynthesis andstructureofherpesvirus DNA:thedistribution of alkali-labile single strand interruptions inHSV-1 DNA. J. Gen. Virol.21:453-467.
37. Wong,T.W.,and D. A. Clayton.1986. DNA primase of human mitochondria is associated with structural RNA that is essential forenzymatic activity. Cell 45:817-825.
38. Wong-Staal, F., J. Mendelsohn, andM.Goulian. 1973.
Ribonu-cleotides in closed circular mitochondrial DNA from HeLa
cells. Biochem. Biophys. Res. Commun. 53:140-148.