Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Interaction of Epstein-Barr Virus Nuclear Antigen Leader
Protein (EBNA-LP) with HS1-Associated Protein X-1:
Implication of Cytoplasmic Function of EBNA-LP
YASUSHI KAWAGUCHI, KAORI NAKAJIMA, MIE IGARASHI, TOMOKO MORITA,MICHIKO TANAKA, MIKIKO SUZUKI, AKIHIKO YOKOYAMA, GO MATSUDA, KENTARO KATO, MIKIKO KANAMORI,ANDKANJI HIRAI*
Department of Tumor Virology, Division of Virology and Immunology, Medical Research Institute, Tokyo Medical and Dental University, Bunkyo-ku, Tokyo 113-8510, Japan
Received 12 June 2000/Accepted 11 August 2000
Epstein-Barr virus (EBV) nuclear antigen leader protein (EBNA-LP) consists of W1W2 repeats and a unique C-terminal Y1Y2 domain and has been suggested to play an important role in EBV-induced transformation. To identify the cellular factors interacting with EBNA-LP, we performed a yeast two-hybrid screen, using EBNA-LP cDNA containing four W1W2 repeats as bait and an EBV-transformed human peripheral blood lymphocyte cDNA library as the source of cellular genes. Our results were as follows. (i) All three cDNAs in positive yeast colonies were found to encode the same cellular protein, HS1-associated protein X-1 (HAX-1), which is localized mainly in the cytoplasm and has been suggested to be involved in the regulation of B-cell signal transduction and apoptosis. (ii) Mutational analysis of EBNA-LP revealed that the association with HAX-1 is mediated by the W1W2 repeat domain. (iii) A purified chimeric protein consisting of glutathione
S-transferase fused to EBNA-LP specifically formed complexes with HAX-1 transiently expressed in COS-7 cells. (iv) When EBNA-LP and HAX-1 were coexpressed in COS-7 cells, EBNA-LP was specifically coimmu-noprecipitated with HAX-1. (v) Careful cell fractionation experiments of an EBV-infected lymphoblastoid cell line revealed that EBNA-LP is localized in the cytoplasm as well as in the nucleus. (vi) When EBNA-LP containing four W1W2 repeats was expressed in COS-7 cells, EBNA-LP was detected mainly in the nucleus by immunofluorescence assay. Interestingly, when EBNA-LP containing a single W1W2 repeat was expressed in COS-7 cells, EBNA-LP was localized predominantly in the cytoplasm and was colocalized with HAX-1. These results indicate that EBNA-LP is in fact present and may have a significant function in the cytoplasm, possibly by interacting with and affecting the function of HAX-1.
Epstein-Barr virus (EBV) is a ubiquitous human herpesvi-rus, closely associated with a variety of neoplastic diseases, including endemic Burkitt’s lymphoma, nasopharyngeal carci-noma, Hodgkin’s disease, various other lymphomas, and gas-tric carcinoma (20, 30). In vitro, EBV infection results in hu-man B-cell activation and establishment of lymphoblastoid cell lines (LCLs) with very long-term proliferation in culture (20, 30). The LCLs carry the EBV genome in an episomal form and express only several proteins, including six EBV nuclear anti-gen (EBNA) proteins (EBNA-1, -2, -3A, -3B, -3C, and -leader protein [EBNA-LP]), three latent membrane proteins (LMP) (LMP1, -2A, and -2B) (20, 30). Among these, LMP1, EBNA-2, EBNA-3A, and EBNA-3C have been shown to be essential for EBV-induced immortalization (7, 12, 19, 41). LMP1 has been shown to mimic the B-lymphocyte activation antigen CD40 by interaction with tumor necrosis factor receptor-associated fac-tors and the tumor necrosis factor receptor-associated death domain (8, 20, 26). The interactions have been shown to play roles in activation of the NF-B pathway and EBV-induced immortalization of B lymphocytes (8, 20, 26). EBNA-2 con-tributes to B-cell immortalization by regulating EBV latency gene expression and by activating the expression of cellular genes through interaction with the host cellular transcription
factor RBP-J(20). EBNA-3A, -3B, and -3C, which are con-sidered to constitute a family, also have the capability to in-teract with RBP-Jand modulate EBNA-2-mediated regula-tion of gene expression (20, 31). While other latency-related proteins, including EBNA-1 and the LMP2s, are not essential for B-cell immortalization, these viral proteins play significant roles in the maintenance of viral latency (20). EBNA-1 binds to the origin of plasmid replication and is required for mainte-nance of the viral genome during latency (20). LMP2A has been shown to block normal B-cell receptor signaling, and this signaling blockade is involved in prevention of reactivation from latency (10, 20).
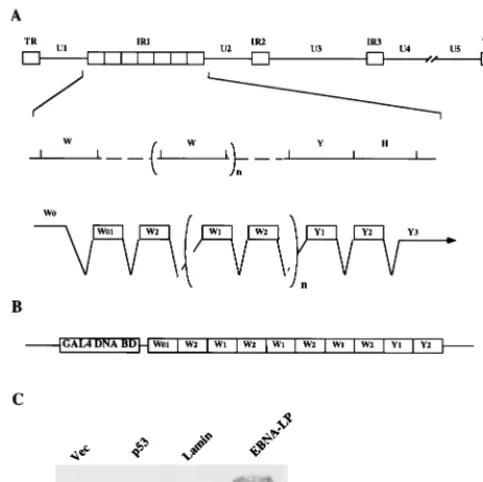
EBNA-LP, a subject of this report, is a critical regulator of EBV-induced B-cell immortalization, based on the observation that recombinant EBNA-LP mutant viruses show much less efficiency in the phenotype (2, 24). The open reading frame of EBNA-LP consists of the multiple-repeat domain encoded by the repeating W1 and W2 exons in the major internal repeat (IR1) and the unique C-terminal Y1 and Y2 exons, which are located downstream of the 3⬘ end of IR1 (see Fig. 1) (32). Together with EBNA-2, EBNA-LP is one of the earliest viral proteins expressed in EBV-infected B cells (1). In freshly in-fected B cells, EBNA-LP is expressed as a protein ladder, possibly as a result of heterogeneous polypeptides with differ-ent numbers of W1W2 repeats (9). In subsequdiffer-ently established LCLs, EBNA-LP is expressed as only one or two species and is localized in nuclear structures called ND10 (9, 37). Although the mechanism by which EBNA-LP acts in EBV-induced B-cell immortalization is unclear, several lines of evidence sug-* Corresponding author. Mailing address: Department of Tumor
Virology, Division of Virology and Immunology, Medical Research Institute, Tokyo Medical and Dental University, Yushima, Bunkyo-ku, Tokyo 113-8510, Japan. Phone: 81-3-5803-5815. Fax: 81-3-5803-0241. E-mail: [email protected].
10104
on November 9, 2019 by guest
http://jvi.asm.org/
been identified. Thus, EBNA-LP has been shown to bind to tumor suppressor proteins, p53, and retinoblastoma protein in in vitro binding experiments (38). In vivo interaction and co-localization in ND10 of EBNA-LP and the 70-kDa family of heat shock proteins have been also demonstrated by coimmu-noprecipitation and immunofluorescence experiments (23, 25, 36). Although the biological significance of these interactions remains to be elucidated, these observations suggest that EBNA-LP interacts with and modulates the function of cellu-lar proteins during the EBV-induced immortalization process. These reports and the recent consensus that viral proteins are multifunctional proteins related to many aspects of cellular functions prompted us to hypothesize that many additional cellular targets of EBNA-LP remain and that further under-standing of EBNA-LP function requires their identification. In this study, we performed a yeast two-hybrid screen. We report here the identification of a new EBNA-LP binding partner, HAX-1 (HS1-associated protein X-1), which is a cytoplasmic protein suggested to be involved in the regulation of B-cell signal transduction and apoptosis (35). Our studies show that (i) EBNA-LP interacts with HAX-1 in yeast, in vitro, and in mammalian cells, (ii) EBNA-LP is localized in the cytoplasm of EBV-infected cells, and (iii) EBNA-LP is colocalized with HAX-1 in the cytoplasm of cells transfected with both EBNA-LP and HAX-1 expression plasmids.
MATERIALS AND METHODS
Cells.A latently EBV-infected LCL (hereafter referred to as simply LCL) was established by infection with the B95-8 strain of EBV in vitro. BJAB is an EBV-negative B-lymphoma cell line. These cell lines were maintained in RPMI 1640 medium supplemented with 10% fetal calf serum. Cells from the monkey kidney epithelial cell line, COS-7, were grown in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum.
Plasmids. To construct pGBT9-EBNA-LPR4, the entire coding sequence of EBNA-LP containing four W1W2 repeats was amplified by PCR from pZipEBNA-LP (42) (kindly provided by E. Kieff) and inserted into pGBT9 (Clontech, Palo Alto, Calif.) in frame with the DNA-binding domain of GAL4. To generate pZipEBNA-LPR1, pZipEBNA-LP was digested with ApaI and religated. The resultant plasmid contained only one W1W2 repeat. Plasmids encoding the GAL4 DNA-binding domain fused to a one-repeat EBNA-LP LPR1) or various fragments of EBNA-LP (pGBT9-EBNA-LPR4⌬Y1Y2, pGBT9-EBNA-LPY1Y2, or pGBT9-EBNA-LPY2) were obtained by PCR amplification of pZipEBNA-LPR1 or pZipEBNA-LP, respectively. Plas-mids encoding the proteins with either two (pGBT9-EBNA-LPR2) or three (pGBT9-EBNA-LPR3) W1W2 repeats linked to the Y1Y2 domain were gener-ated by cloning the 200-bpApaI fragment of pZipEBNA-LP into theApaI site of pGBT9-EBNA-LPR1. pGEX-EBNA-LPR4 was generated by cloning the EcoRI-SalI fragment of pGBT9-EBNA-LPR4 into theEcoRI andSalI sites of pGEX4T-1 (Amersham-Pharmacia, Uppsala, Sweden) in frame with glutathione S-transferase (GST).
To construct pBS-Flag, the oligonucleotide 5⬘-GCGGATCCGACTACAAG GACGACGATGACAAGTCTAGAGC-3⬘, annealed with its complement, was digested withBamHI andXbaI and inserted into theBamHI andXbaI sites of pBluescript II KS(⫹) (Stratagene, La Jolla, Calif.). Then pBS-Flag was digested withXbaI, treated with T4 DNA polymerase, and religated to yield a stop codon; the resulting plasmid was designated pBS-Flag-Stop. A PCR fragment of SR␣ -human HAX-1 (35) (kindly provided by T. Watanabe) encoding the entire
from colonies that grew on SD synthetic medium lacking histidine and expressed -galactosidase. The positive plasmids were cotransformed with pGBT9, pGBT9-EBNA-LPR4, pLAM5⬘encoding human lamin C (Clontech), or pVA3 encoding murine p53 (Clontech) into HF7c cells, and transformants were as-sayed for-galactosidase activity to eliminate false positives. The quantitative o-nitrophenyl--D-galactosidase assay was performed according to the
manufac-turer’s instructions (Clontech).
Production and purification of GST fusion proteins.GST fusion proteins were expressed inEscherichia coliBL21 transformed with either pGEX4T-1 or pGEX-EBNA-LPR4 and purified on glutathione-Sepharose beads (Amersham-Phar-macia) as described previously (16).
Affinity precipitation with GST-EBNA-LP fusion protein.COS-7 cells were transfected with pME-HAX-1(F) according to the DEAE-dextran method as described previously (33), with minor modifications. Briefly, near-confluent COS-7 cells on 100-mm-diameter dishes were washed with Opti-MEM (Gibco BRL, Gaithersburg, Md.), and plasmid DNAs in 6 ml of MEM–DEAE-dextran solution (400 mg of DEAE-MEM–DEAE-dextran/ml, 0.1 mM chloroquine in Opti-MEM) were added to the cells. At 4 h after transfection, the cells were treated with 6 ml of 10% dimethyl sulfoxide in phosphate-buffered saline (PBS) for 2 min, and then fresh medium was added. At 3 days after transfection, the cells were lysed in HEPES buffer (50 mM HEPES [pH 7.4], 250 mM NaCl, 10 mM MgCl2, 1 mM phenylmethylsulfonyl fluoride [PMSF]) containing 1% Triton
X-100. The cell lysate was incubated with GST or GST–EBNA-LP containing four W1W2 repeats immobilized on glutathione-Sepharose beads. After the beads were washed extensively with HEPES buffer containing 1% Triton X-100, the bound protein complexes were subjected to electrophoresis on a 10% poly-acrylamide gel containing sodium dodecyl sulfate (SDS), transferred to a nitro-cellulose sheet, and reacted with a mouse monoclonal antibody to the Flag epitope M2 (Sigma, St. Louis, Mo.).
Coimmunoprecipitation.Immunoprecipitation experiments were done as de-scribed previously (18). Briefly, COS-7 cells transfected with appropriate expres-sion plasmids by the DEAE-dextran method as described above were lysed in NP-40 buffer (50 mM Tris-HCl [pH 8.0], 150 mM NaCl, 0.5% Nonidet P-40, 1 mM PMSF). The supernatants obtained after centrifugation of cell lysates were precleared by mixing them with protein G-Sepharose beads (Amersham-Phar-macia) for 30 min and were reacted with the M2 antibody for 2 h at 4°C. Protein G-Sepharose beads were then added and allowed to react for an additional 1 h. Immunoprecipitates were collected by brief centrifugation, rinsed extensively with NP-40 buffer, subjected to electrophoresis on a 10% polyacrylamide gel containing SDS, transferred to a nitrocellulose sheet, and reacted with a mouse monoclonal antibody to EBNA-LP (JF186; provided by G. Klein) and the M2 antibody.
Immunoblotting. The electrophoretically separated proteins transferred to nitrocellulose sheets were reacted with appropriate antibodies as described pre-viously (17). Briefly, the nitrocellulose sheets were blocked with 5% skim milk in T-PBS (PBS containing 0.1% Tween 20) for 1 h or overnight, rinsed twice, washed once for 15 min and twice for 5 min, and reacted for 2 h with primary antibodies in T-PBS containing 1% bovine serum albumin (BSA). The blots were then washed as before, reacted for 1 h with peroxidase-conjugated goat anti-mouse immunoglobulin G (IgG) (Amersham-Pharmacia) in T-PBS containing 3% skim milk, rinsed twice, washed once for 15 min and four times for 5 min each time in T-PBS, and developed using the enhanced chemiluminescence reagent (Amersham-Pharmacia).
Cell fractionation.Nuclear and cytoplasmic fractions of LCL were obtained by the method described previously (16). Briefly, LCL cells were harvested, washed with PBS, and resuspended in buffer A (10 mM HEPES [pH 7.4], 1.5 mM MgCl2,
10 mM NaCl, 1 mM PMSF). The cells were lysed by addition of Nonidet P-40 to 0.1%, and the nuclei were pelleted by centrifugation at top speed in a micro-centrifuge. The supernatant (cytoplasmic fraction) was transferred to a new tube. The pellet was washed with buffer A twice, resuspended in buffer C (10 mM HEPES [pH 7.4], 1.5 mM MgCl2, 420 mM NaCl, 1 mM PMSF), and sonicated
briefly. The remaining debris was removed by centrifugation as described above, and the supernatant (nuclear fraction) was collected. These fractions were then subjected to electrophoresis on polyacrylamide gels containing SDS, transferred
on November 9, 2019 by guest
http://jvi.asm.org/
to nitrocellulose sheets, and reacted with mouse monoclonal antibody to EBNA-LP (JF186) or EBNA-2 (DAKO, Kyoto, Japan).
Immunofluorescence.Approximately 5⫻104COS-7 cells were seeded onto
glass slides and transfected with appropriate expression vectors using DOTAP (Roche Molecular Biochemicals, Tokyo, Japan) according to the manufacturer’s instructions. The cells were fixed in ice-cold methanol and acetone (1:1) at 3 days after transfection, blocked for 30 min in PBS containing 20% goat serum and 1% BSA at room temperature, rinsed once in PBS, reacted for 2 h with the primary antibodies in PBS containing 10% goat serum and 0.2% BSA, rinsed three times in PBS, reacted for 1 h with secondary antibodies in PBS containing 10% goat serum and 0.2% BSA, rinsed as before, and mounted in PBS containing 90% glycerol. The mouse monoclonal antibody to EBNA-LP (JF186) and/or a rabbit polyclonal antibody to the Flag epitope (sc-807; Santa Cruz Biotechnology, Inc., Santa Cruz, Calif.) was used as a primary antibody. A goat-anti mouse IgG conjugated to fluorescein isothiocyanate (Sigma) and a goat-anti rabbit IgG conjugated to Texas red (Molecular Probes, Eugene, Oreg.) were used as sec-ondary antibodies. The slides were examined under a Zeiss fluorescence micro-scope.
RESULTS
EBNA-LP interacts with HAX-1 in a yeast two-hybrid sys-tem.The results of the two-hybrid screen described in Mate-rials and Methods were as follows. Of 2.6⫻106colonies, 22
were histidine positive, and of these, 3 colonies expressed -ga-lactosidase activity. Three cDNAs isolated from the positive colonies were positive for interaction with EBNA-LP and neg-ative for interaction with human lamin, murine p53, or the GAL4 DNA-binding domain alone (Fig. 1C). The sequences of the 5⬘ ends of the cDNAs were determined, and all were
found to completely match the published sequence of human HAX-1 (35).
W1W2 repeat domain of EBNA-LP interacts with HAX-1. To map the domain of EBNA-LP responsible for the interac-tion with HAX-1, a series of deleinterac-tion clones of EBNA-LP were constructed (Fig. 2) and tested for interaction with HAX-1 in the yeast two-hybrid system. As shown in Fig. 2, EBNA-LP from which Y1Y2 was deleted interacts with HAX-1 as effi-ciently as wild-type EBNA-LP, while the Y1Y2 or Y2 domain alone was not able to interact with HAX-1. Two further EBNA-LP mutants were generated to determine whether the number of copies of W1W2 repeats of EBNA-LP influences the interaction with HAX-1. The interactions of EBNA-LP species with the two or three copies of the W1W2 repeat with HAX-1 were as significant as those with four copies of the repeat (Fig. 2). The results were confirmed by quantitative
o-nitrophenyl--D-galactosidase assay (data not shown). These results indicated that HAX-1 interacts with the W1W2 domain of EBNA-LP but that the strength of the interaction does not depend on the number of W1W2 repeats.
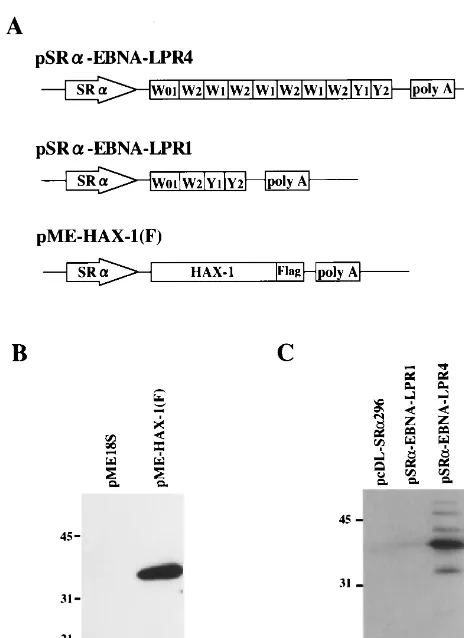
GST–EBNA-LP specifically forms complexes with HAX-1 in vitro.To verify and extend the binding data obtained in yeast, a GST–EBNA-LP fusion protein was expressed inE. coli. The GST–EBNA-LP or GST protein bound to glutathione-Sepha-rose beads was reacted with an extract from COS-7 cells trans-fected with pME-HAX-1(F). In COS-7 cells transtrans-fected with the expression vector, Flag epitope-tagged HAX-1 was ex-pressed, and anti-Flag monoclonal antibody specifically de-tected the protein (Fig. 3B). After extensive rinsing, protein complex captured on the beads was solubilized, subjected to electrophoresis in a denaturing gel, transferred to a nitrocel-lulose sheet, and reacted with monoclonal antibody to the Flag epitope. As shown in Fig. 4, the GST–EBNA-LP protein was able to pull down HAX-1 (lane 2), whereas GST alone did not (lane 1). The electrophoretic mobility of the HAX-1 that com-plexed with GST–EBNA-LP fusion protein (lane 2) was similar to that found in the whole-cell extracts of the transfected cells (lane 3), while the monoclonal antibody to the Flag epitope did not detect any GST–EBNA-LP fusion proteins (lane 4). These results indicated that EBNA-LP can complex with a full-length sequence of HAX-1 in vitro.
EBNA-LP interacts with HAX-1 in mammalian cells. To determine whether EBNA-LP associates with HAX-1 at the cellular level, extracts from COS-7 cells transfected with the FIG. 1. (A) Schematic diagram of the sequence of the EBV genome and
[image:3.612.317.541.72.223.2]location of the EBNA-LP gene. The top line is a linear representation of the EBV genome. The unique sequences are represented as U1 to U5. The terminal and internal repeats flanking the unique sequences are shown as open rectangles with their designations given above. The middle line shows an expanded section of the domain encoding the EBNA-LP gene. The exons of EBNA-LP open reading frames are derived from theBamHI W and Y fragments. The bottom line shows the structures of the EBNA-LP transcript and coding regions. (B) Schematic diagram of the bait plasmid, pGBT9-EBNA-LPR4, for yeast two-hybrid screening. (C) Specific interaction between EBNA-LP and HAX-1 in the yeast two-hybrid system. Yeast strain HF7c that was cotransfected with the GAL4 activation domain fused to cDNA encoding HAX-1 in combination with the GAL4 DNA-binding domain alone, murine p53, human lamin, or EBNA-LP was assayed for-galactosidase activity as described in Materials and Methods.
FIG. 2. Mapping of the domain of EBNA-LP that interacts with HAX-1 and influence of the number of W1W2 repeats on the interaction with HAX-1. The EBNA-LP deletion mutants and the level of-galactosidase activity detected for each construct in the presence of HAX-1 in the yeast two-hybrid system are shown.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.53.295.73.314.2]indicated expression vectors (Fig. 5) were immunoprecipitated with the anti-Flag epitope antibody. Immunoprecipitates were solubilized, separated on a denaturing gel, transferred to a nitrocellulose membrane, and reacted with the monoclonal antibody to EBNA-LP. As shown in Fig. 5A, the anti-Flag epitope antibody coprecipitated EBNA-LP with Flag epitope-tagged HAX-1 when EBNA-LP and HAX-1 were coexpressed in COS-7 cells (lane 1). In contrast, when EBNA-LP was ex-pressed by itself (lane 2) or EBNA-LP and an unrelated Flag epitope-tagged protein (BMAL1) were coexpressed (lane 3), EBNA-LP was not coprecipitated by the antibody. The levels of expression of EBNA-LP in the whole-cell extracts of the transfected cells were equivalent (lanes 4 to 6), and the elec-trophoretic mobilities of the proteins were similar to that co-immunoprecipitated with HAX-1. Figure 5B shows the results of immunoblotting of the same nitrocellulose sheet depicted in Fig. 5A but reprobed with the anti-Flag epitope antibody. The results indicated that each protein tagged with the Flag epitope was appropriately expressed in COS-7 cells and immunopre-cipitated by the antibody to the Flag epitope. These observa-tions indicated that EBNA-LP interacts with HAX-1 in mam-malian cells.
EBNA-LP was detected in the cytoplasmic fraction of EBV-infected LCL.EBNA-LP is thought to be primarily a nuclear
detected only in the nuclear fraction but not in the cytoplasmic fraction of LCL, indicating that EBNA-LP in the cytoplasmic fraction was not due to leakage from the nuclei during the experimental procedure. These results indicate that EBNA-LP is in fact a cytoplasmic protein in EBV-infected cells.
[image:4.612.55.287.69.388.2]EBNA-LP with a single W1W2 repeat transiently expressed in COS-7 cells was localized predominantly in the cytoplasm of transfected cells and is colocalized with HAX-1.Next, we investigated the localization of EBNA-LP in transfected COS-7 cells. Slide cultures of COS-7 cells were transfected with EBNA-LP expression vectors, fixed at 3 days posttrans-fection, and reacted with the EBNA-LP monoclonal anti-body (JF186). As shown in Fig. 7A, diffuse staining of EBNA-LP was detected mainly in the nuclei in COS-7 cells transfected with the expression vector of EBNA-LP containing four copies of the W1W2 repeat. Similar results were obtained with expression vectors of EBNA-LP containing two or three copies of the W1W2 repeat (data not shown). Although cyto-plasmic EBNA-LP with multiple copies of W1W2 repeats was difficult to detect by immunofluorescence analysis, EBNA-LP with four W1W2 repeats was clearly detected in cytoplasmic fractions of transfected COS-7 cells by cell fractionation anal-ysis (data not shown). When the expression vector containing
FIG. 4. Photograph of an immunoblot of transfected cell proteins bound to GST or chimeric GST–EBNA-LP with four W1W2 repeats, electrophoretically separated in a denaturing gel, and reacted with mouse monoclonal antibody to the Flag epitope (M2). Lysates of COS-7 cells transfected with pME-HAX-1(F) were reacted with GST or GST–EBNA-LPR4 chimeric protein immobilized on glutathione-Sepharose beads. The beads were pelleted, rinsed extensively, sub-jected to electrophoresis on a denaturing gel, transferred to a nitrocellulose sheet, and reacted with the anti-Flag epitope antibody. Lanes 1 and 2, whole-cell extracts (WCE) from COS-7 cells transfected with pME-HAX-1(F) bound to GST and GST–EBNA-LPR4, respectively; lane 3, whole-cell extracts (WCE) from COS-7 cells transfected with pME-HAX-1(F); lane 4, GST–EBNA-LPR4. Molecular weights (in thousands) are shown on the left.
FIG. 3. (A) Schematic diagrams of the expression plasmid containing EBNA-LP with four W1W2 repeats (top line) or a single W1W2 repeat (middle line) or HAX-1 tagged with the Flag epitope (bottom line). (B and C) Photo-graph of an immunoblot of electrophoretically separated lysates of COS-7 cells transfected with the indicated plasmids, which were probed with mouse mono-clonal antibody to the Flag epitope (B) or mouse monomono-clonal antibody to EBNA-LP (JF186) (C). Molecular weights (in thousands) are shown on the left.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.315.553.473.627.2]only one copy of the W1W2 repeat was transfected into COS-7 cells, EBNA-LP was evident as punctate staining predomi-nantly in the cytoplasm (Fig. 7B). These results indicated that transiently expressed EBNA-LP is in fact localized in the cy-toplasm of cells and that the cycy-toplasmic localization of EBNA-LP detected by immunofluorescence analysis depends on the W1W2 repeat copy number.
As the pattern of cytoplasmic EBNA-LP staining was rem-iniscent of that of HAX-1 reported elsewhere (35), double-labeling immunofluorescence assays were performed to de-termine whether the EBNA-LP is colocalized with HAX-1.
COS-7 cells that were cotransfected with expression constructs for EBNA-LP (with one copy of the W1W2 repeat and Flag epitope-tagged HAX-1) were fixed at 3 days posttransfection and reacted with the anti-EBNA-LP monoclonal antibody (JF186) and a polyclonal antibody to the Flag epitope (sc-807). As shown in Fig. 7C, EBNA-LP was colocalized with HAX-1 in the cytoplasm of transfected COS-7 cells, further supporting the finding that EBNA-LP interacts with HAX-1.
DISCUSSION
We report here the identification of a cellular protein, HAX-1, as a new EBNA-LP binding protein in the yeast two-hybrid system, in in vitro biochemical assays, and in coimmu-noprecipitation experiments. The salient features of this study were as follows.
(i) In the yeast two-hybrid system, EBNA-LP interacted with HAX-1 (Fig. 1). The interaction of HAX-1 with EBNA-LP was also demonstrated by using the GST–EBNA-LP fusion protein in pull-down experiments (Fig. 4), which reinforced the evi-dence of physical interaction between the two proteins. Fur-thermore, consistent with the binding data obtained in yeast and in vitro, EBNA-LP containing four W1W2 repeats was coimmunoprecipitated (Fig. 5) and the protein with the single W1W2 repeat was colocalized (Fig. 7) with HAX-1 when both of the proteins were transiently coexpressed in COS-7 cells, indicating that the interaction between EBNA-LP and HAX-1 occurs at the cellular level.
[image:5.612.104.501.71.307.2](ii) HAX-1 was originally identified by the yeast two-hybrid screen as a protein that associates with HS1 (35). HS1 contains an Src homology 3 domain, an acidic␣helix structure, and a basic motif resembling the DNA-binding motif of the helix-loop-helix protein family (21, 22, 40, 43). A mouse cell line, WEHI-231, is known to undergo apoptosis after B-cell recep-tor cross-linking (3, 14). A variant of the cell line in which the level of HS1 expression is significantly reduced has been shown FIG. 5. Interaction between EBNA-LP and HAX-1 in mammalian cells. COS-7 cells transiently expressing the indicated protein(s) were immunoprecipitated with the mouse monoclonal antibody to the Flag epitope (M2). The immunoprecipitates were subjected to electrophoresis on a denaturing gel, transferred to a nitrocellulose sheet, reacted with mouse monoclonal antibody to EBNA-LP (JF186) (A), and then reprobed with anti-Flag epitope antibody (M2) (B). Molecular weights (in thousands) are shown on the left.
FIG. 6. Photograph of immunoblots of electrophoretically separated cell fractions of BJAB cells and LCL infected with EBV, which were probed with mouse monoclonal antibody to EBNA-LP (A) or to EBNA-2 (B). Nuclear fractions (Nu) (lanes 1 and 3) and cytoplasmic fractions (Cy) (lanes 2 and 4) were prepared as described in Materials and Methods, subjected to electrophoresis on denaturing gels, transferred to nitrocellulose sheets, and reacted with mouse monoclonal antibody to EBNA-LP (A) or to EBNA-2 (B). Molecular weights (in thousands) are shown on the left.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.78.267.509.659.2]to be resistant to B-cell receptor-induced apoptosis (4, 15). Interestingly, exogenous expression of HS1 in the variant cell line restores the sensitivity of the cell line to B-cell receptor-mediated apoptosis (11). These data indicate that HS1 plays a role in signal transduction from antigen receptors.
HAX-1 mRNA is expressed ubiquitously among tissues, and the protein is localized in the cytoplasm (35). The predicted amino acid sequence of HAX-1 shows similarity to that of Nip3 (5, 35). Nip3 was identified as an adenovirus E1B 19-kDa-interacting protein using a yeast two-hybrid system and was also shown to interact with several antiapoptotic viral and cellular proteins, including Bcl-2 and BHRF-1 (an EBV Bcl-2 homolog) (5). Nip3 is classified in the Bcl-2 family based on limited sequence homology to the Bcl-2 homology 3 domain and the carboxyl-terminal transmembrane domain (5, 6, 29, 44). Studies of Nip3 revealed that the protein is a mitochon-drial protein that induces apoptosis and enhances apoptosis
though the biological significance of the interaction remains to be elucidated. Additionally, our preliminary study showed that HAX-1 interacts with BHRF-1, which is an EBV homolog of Bcl-2 and is known to promote survival of EBV-infected cells (20), further supporting the hypothesis that the HAX-1– EBNA-LP interaction is involved in the regulation of apoptosis in EBV-infected cells (K. Nakajima, Y. Kawaguchi, and K. Hirai, unpublished data).
[image:6.612.54.292.69.440.2](iii) EBNA-LP accumulates in both the nuclei and cytoplasm in EBV-infected LCL. If EBNA-LP in fact interacts with HAX-1, it would be necessary for EBNA-LP to be localized in the cytoplasm because HAX-1 is a cytoplasmic protein (35). However, until this study, EBNA-LP was believed to be a nuclear protein (28). In this study, we carefully separated nu-clear and cytoplasmic fractions of LCL and examined the ex-istence of EBNA-LP in each fraction. Although the results (Fig. 6) clearly showed evidence of native EBNA-LP in the cytoplasmic fraction of LCL, one would argue that EBNA-LP detected in the cytoplasmic fraction is due to leakage from the nucleus during the experimental procedure. Our conclusion is supported by the following observations. First, EBNA-2, known to be sequestered only in the nuclei of LCL, was de-tected in the nuclear fraction but not in the cytoplasmic frac-tion of LCL. Second, under the experimental condifrac-tions used in this study, leakage was not observed even from fragile nu-clei, such as those in herpes simplex virus-infected cells as was reported elsewhere (16). Third, EBNA-LP containing four W1W2 repeats was coimmunoprecipitated with HAX-1 in COS-7 cells transfected with the corresponding expression vec-tors. As reported elsewhere (35) and shown in Fig. 7, HAX-1 expressed in COS-7 cells was localized only in the cytoplasm. Further, the localization of HAX-1 was not affected by EBNA-LP with four W1W2 repeats in COS-7 cells transfected with both of the expression vectors (data not shown). These results indicated that EBNA-LP coimmunoprecipitated with HAX-1 was derived from the cytoplasm. Fourth, as shown in Fig. 7, EBNA-LP containing only one W1W2 repeat was de-tected in the cytoplasm by immunofluorescence assay. These results indicated that EBNA-LP has the ability to accumulate in the cytoplasm of mammalian cells.
(iv) The pattern of localization of the single-repeat EBNA-LP in COS-7 cells was clearly distinct from those of EBNA-LP with more than two W1W2 repeats (Fig. 7). The EBNA-LP with a single repeat showed a punctate staining pattern predominantly in the cytoplasm of COS-7 cells and was colocalized with HAX-1. In contrast, nuclear EBNA-LP with a single repeat was barely detectable, and proteins with more than two W1W2 repeats were mainly localized in the nucleus. These observations suggest that the EBNA-LP with the single repeat has a specific function in the cytoplasm, possibly by interacting with HAX-1. This speculation is supported by the previous report that expression of EBNA-LP with only one FIG. 7. Localization of EBNA-LP or HAX-1 in COS-7 cells that were
trans-fected with pSR␣-EBNA-LPR4 (A) or pSR␣-EBNA-LPR1 (B) or cotransfected with pSR␣-EBNA-LPR1 and pME-HAX-1(F) (C and D). The singly transfected cells (A and B) were fixed and reacted with the EBNA-LP antibody (JF186) and anti-mouse IgG conjugated to fluorescein isothiocyanate (green fluorescence). The cotransfected cells (C and D) were fixed and reacted with mouse monoclo-nal antibody to EBNA-LP (JF186) and rabbit polyclomonoclo-nal antibody to the Flag epitope (sc-807) and then reacted with anti-mouse IgG conjugated to fluorescein isothiocyanate and anti-rabbit IgG conjugated to Texas red (red fluorescence). Single-color images were photographed separately and are shown for EBNA-LP (C) and HAX-1 (D).
on November 9, 2019 by guest
http://jvi.asm.org/
copy of the W1W2 repeat was in fact observed in primary B cells and in an EBV-negative Burkitt’s lymphoma cell line freshly infected with EBV (9). Our results also indicate that the tendency of EBNA-LP to be localized in the cytoplasm de-pends on the number of W1W2 repeats. To our knowledge, this is the first report demonstrating that different species of EBNA-LP show biological differences. These observations may explain why EBNA-LP is expressed as multiple protein species with various numbers of W1W2 repeats. Furthermore, our results may account for the observation by Nitsche et al. (27) that EBNA-LP cooperatively induced LMP1 expression with EBNA-2 in transient transfection systems, while EBNA-LP with a single repeat was inactive for this phenotype. Conceiv-ably, single-repeat EBNA-LP is localized only in the cytoplasm and does not encounter EBNA-2, resulting in inability to mod-ulate EBNA-2-mediated regulation of gene expression.
The salient new contribution presented in this report is that EBNA-LP may have a significant function in the cytoplasm of infected cells, possibly by interacting with and affecting the function of HAX-1. Further experiments to unveil the biolog-ical significance of the interaction between EBNA-LP and HAX-1 are needed and are under way in our laboratory.
ACKNOWLEDGMENTS Y.K. and K.N. contributed equally to this work.
We thank E. Kieff for the EBNA-LP cDNA, G. Klein for the mouse monoclonal antibody to EBNA-LP, T. Watanabe for the HAX-1 cDNA, K. Maruyama for pME18S, and R. Furuya for technical assis-tance.
This study was supported in part by grants for scientific research (Y.K. and K.H.) and a grant for scientific research in priority areas (K.H.) from the Ministry of Education, Science, Sports, and Culture of Japan. Y.K. was supported by a grant from the Inamori Foundation.
REFERENCES
1.Alfieri, C., M. Birkenbach, and E. Kieff.1991. Early events in Epstein-Barr virus infection of human B lymphocytes. Virology181:595–608.
2.Allan, G. J., G. J. Inman, B. D. Parker, D. T. Rowe, and P. J. Farrell.1992. Cell growth effects of Epstein-Barr virus leader protein. J. Gen. Virol.73: 1547–1551.
3.Benhamou, L. E., P.-A. Cazenave, and P. Sarthou.1990. Anti-immunoglobu-lins induce cell death by apoptosis in WEHI-231 B lymphoma cells. Eur. J. Immunol.20:1405–1407.
4.Benhamou, L. E., T. Watanabe, D. Kitamura, P. A. Cazenave, and P. Sarthou.1994. Signaling properties of anti-immunoglobulin-resistant vari-ants of WEHI-231 B lymphoma cells. Eur. J. Immunol.24:1993–1999. 5.Boyd, J. M., S. Malstrom, T. Subramanian, L. K. Venkatesh, U. Schaeper, B.
Elangovan, C. D’Sa-Eipper, and G. Chinnadurai.1994. Adenovirus E1B 19 kDa and Bcl-2 proteins interact with a common set of cellular proteins. Cell 79:341–351.
6.Chen, G., R. Ray, D. Dubik, L. Ahi, J. Cizeau, R. C. Bleackley, S. Saxena, R. Dan Gietz, and A. H. Greenberg.1997. The E1B 19K/Bcl-2-binding protein Nip3 is a dimeric mitochondrial protein that activates apoptosis. J. Exp. Med. 186:1975–1983.
7.Cohen, J. I., F. Wang, J. Mannick, and E. Kieff.1989. Epstein-Barr virus nuclear protein 2 is a key determinant of lymphocyte transformation. Proc. Natl. Acad. Sci. USA86:9558–9562.
8.Damania, B., J.-K. Choi, and J. U. Jung.2000. Signaling activities of gam-maherpesvirus membrane proteins. J. Virol.74:1593–1601.
9.Finke, J., M. Rowe, B. Kallin, I. Ernberg, A. Rosen, J. Dillner, and G. Klein. 1987. Monoclonal and polyclonal antibodies against Epstein-Barr virus nu-clear antigen 5 (EBNA-5) detect multiple protein species in Burkitt’s lym-phoma and lymphoblastoid cell lines. J. Virol.61:3870–3878.
10. Fruehling, S., R. Caldwell, and R. Longnecker.1997. LMP2A function in EBV latency. Epstein-Barr Virus Rep.4:151–159.
11. Fukuda, T., D. Kitamura, I. Taniuchi, Y. Maekawa, L. E. Benhamou, P. Sarthou, and T. Watanabe.1995. Restoration of surface IgM-mediated apoptosis in an anti-IgM-resistant variant of WEHI-231 lymphoma cells by HS1, a protein-tyrosine kinase. Proc. Natl. Acad. Sci. USA92:7302–7306. 12. Hammerschmidt, W., and B. Sugden.1989. Genetic analysis of
immortaliz-ing functions of Epstein-Barr virus in human B lymphocytes. Nature340: 393–397.
13. Harada, S., and E. Kieff.1997. Epstein-Barr virus nuclear protein LP stim-ulates EBNA-2 acidic domain-mediated transcriptional activation. J. Virol. 71:6611–6618.
14. Hasbold, J., and G. G. B. Klaus.1990. Anti-immunoglobulin antibodies induce apoptosis in immature B cell lymphomas. Eur. J. Immunol.20:1685– 1690.
15. Hibner, U., L. E. Benhamou, M. Haury, P.-A. Cazenave, and P. Sarthou. 1994. Signaling of programmed cell death induction in WEHI-231 B lym-phoma cells. Eur. J. Immunol.23:2821–2825.
16. Kawaguchi, Y., R. Bruni, and B. Roizman.1997. Interaction of herpes simplex virus 1␣regulatory protein ICP0 with elongation factor 1␦: ICP0 affects translational machinery. J. Virol.71:1019–1024.
17. Kawaguchi, Y., C. Van Sant, and B. Roizman.1997. Herpes simplex virus 1 ␣regulatory protein ICP0 interacts with and stabilizes the cell cycle regulator cyclin D3. J. Virol.71:7328–7336.
18. Kawaguchi, Y., C. Van Sant, and B. Roizman.1998. Eukaryotic elongation factor 1␦is hyperphosphorylated by the protein kinase encoded by the UL13
gene of herpes simplex virus 1. J. Virol.72:1731–1736.
19.Kaye, K. M., K. M. Izumi, and E. Kieff.1993. Epstein-Barr virus latent membrane protein 1 is essential for B-lymphocyte growth transformation. Proc. Natl. Acad. Sci. USA90:9150–9154.
20. Kieff, E.1996. Epstein-Barr virus and its replication, p. 2343–2396.InB. N. Fields, D. M. Knipe, P. Howley, R. M. Chanock, M. S. Hirsch, J. L. Melnick, T. P. Monath, and B. Roizman (ed.), Virology, 3rd ed. Raven Press, New York, N.Y.
21. Kitamura, D., H. Kaneko, Y. Miyagoe, T. Ariyasu, and T. Watanabe.1989. Isolation and characterization of a novel human gene expressed specifically in the cells of hematopoietic lineage. Nucleic Acids Res.17:9367–9379. 22. Kitamura, D., H. Kaneko, I. Taniguchi, I. Akagi, K. Yamamura, and T.
Watanabe.1995. Molecular cloning and characterization of mouse HS1. Biochem. Biophys. Res. Commun.208:1137–1146.
23. Kitay, M. K., and D. T. Rowe.1996. Protein-protein interactions between Epstein-Barr virus nuclear antigen-LP and cellular gene products: binding of 70-kilodalton heat shock proteins. Virology220:91–99.
24. Mannick, J. B., J. I. Cohen, M. Birkenbach, A. Marchini, and E. Kieff.1991. The Epstein-Barr virus nuclear protein encoded by the leader of the EBNA RNAs is important in B-lymphocyte transformation. J. Virol.65:6826–6837. 25. Mannick, J. B., X. Tong, A. Hemnes, and E. Kieff.1995. The Epstein-Barr virus nuclear antigen leader protein associates with hsp72/hsc73. J. Virol. 69:8169–8172.
26. Mosialos, G.1997. Signal transduction by the Epstein-Barr virus latent membrane protein-1. Epstein-Barr Virus Rep.4:121–126.
27. Nitsche, F., A. Bell, and A. Rickinson.1997. Epstein-Barr virus leader pro-tein enhances EBNA-2-mediated transactivation of latent membrane propro-tein 1 expression: a role for the W1W2 repeat domain. J. Virol.71:6619–6628. 28. Petti, L., C. Sample, and E. Kieff.1990. Subnuclear localization and
phos-phorylation of Epstein-Barr virus latent infection nuclear proteins. Virology 176:563–574.
29. Ray, R., G. Chen, C. Vande, J. Cizeau, J. H. Park, J. C. Reed, R. D. Gietz, and A. H. Greenberg.2000. BNIP3 heterodimerizes with Bcl-2/Bcl-XLand
in-duces cell death independent of a Bcl-2 homology 3 (BH3) domain at both mitochondrial and nonmitochondrial sites. J. Biol. Chem.275:1439–1448. 30. Rickinson, A. B., and E. Kieff.1996. Epstein-Barr virus, p. 2397–2446.In
B. N. Fields, D. M. Knipe, P. Howley, R. M. Chanock, M. S. Hirsch, J. L. Melnick, T. P. Monath, and B. Roizman (ed.), Virology, 3rd ed. Raven Press, New York, N.Y.
31. Robertson, E.1997. The Epstein-Barr virus EBNA 3 protein family as reg-ulators of transcription. Epstein-Barr Virus Rep.4:143–150.
32. Sample, J., M. Hummel, D. Braun, M. Birkenbach, and E. Kieff.1986. Nucleotide sequence of mRNAs encoding Epstein-Barr virus nuclear pro-teins: a probable transcriptional initiation site. Proc. Natl. Acad. Sci. USA 83:5096–5100.
33. Seed, B., and A. Aruffo.1987. Molecular cloning of the CD2 antigen, the T cell erythrocyte receptor, by a rapid immunoselection procedure. Proc. Natl. Acad. Sci. USA84:3365–3369.
34. Sinclair, A. J., I. Palmero, G. Peters, and P. J. Farrell.1994. EBNA-2 and EBNA-LP cooperate to cause G0 to G1 transition during immortalization of resting human B lymphocytes by Epstein-Barr virus. EMBO J.13:3321–3328. 35. Suzuki, Y., C. Demoliere, D. Kitamura, H. Takeshita, U. Deuschle, and T. Watanabe.1997. HAX-1, a novel intracellular protein, localized on mito-chondria, directly associates with HS1, a substrate of Src family tyrosine kinases. J. Immunol.158:2736–2744.
36. Szekely, L., W.-Q. Jiang, K. Pokrovskaja, K. G. Wiman, G. Klein, and N. Ringertz.1995. Reversible nucleolar translocation of Epstein-Barr virus-encoded EBNA5 and hsp70 proteins after exposure to heat shock or cell density congestion. J. Gen. Virol.76:2423–2432.
37. Szekely, L., K. Pokrovskaja, W.-Q. Jiang, H. D. The, N. Ringertz, and G. Klein.1996. The Epstein-Barr virus-encoded nuclear antigen EBNA-5 ac-cumulates in PML-containing bodies. J. Virol.70:2562–2568.
38. Szekely, L., G. Selovanova, K. P. Magnusson, G. Klein, and K. G. Wiman. 1993. EBNA-5, an Epstein-Barr virus-encoded nuclear antigen, binds to the retinoblastoma and p53 proteins. Proc. Natl. Acad. Sci. USA90:5455–5459. 39. Takebe, Y., M. Seiki, J.-I. Fujisawa, P. Hoy, K. Yokota, K.-I. Arai, M. Yoshida, and N. Arai.1988. SR␣promoter: an efficient and versatile mam-malian cDNA expression system composed of the simian virus 40 early