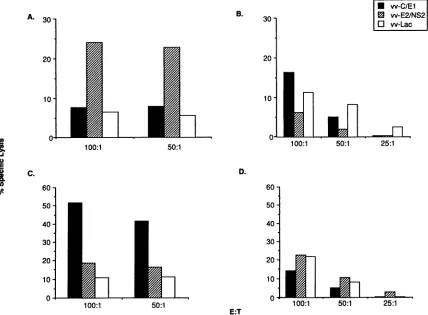
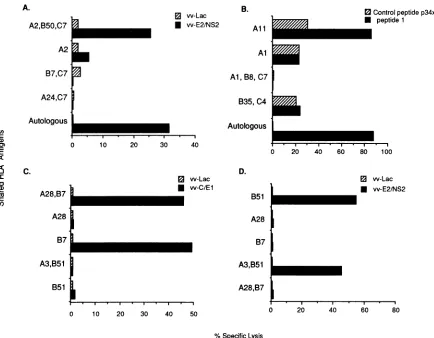
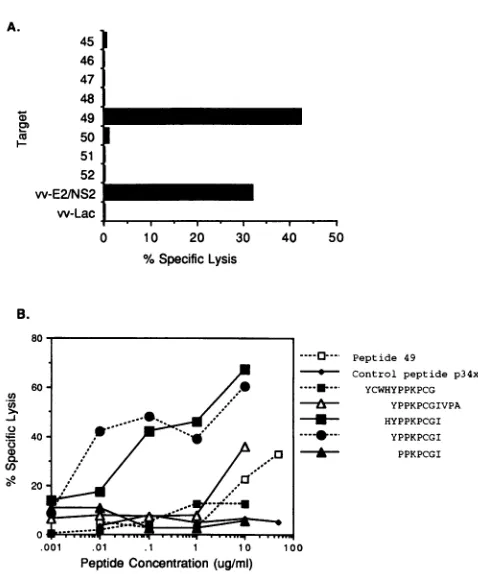
Hepatitis C virus (HCV)-specific cytotoxic T lymphocytes recognize epitopes in the core and envelope proteins of HCV.
11
0
0
Full text
Figure


+3

Related documents