Copyright © 1998, American Society for Microbiology
Characterization of the DNA-Binding Domain of the Avian Y-Box
Protein, chkYB-2, and Mutational Analysis of Its Single-Strand
Binding Motif in the Rous Sarcoma Virus Enhancer
ASHOK NAMBIAR, S. K. SWAMYNATHAN, JAGANNADHA C. KANDALA,ANDRAMAREDDY V. GUNTAKA*
Molecular Microbiology and Immunology, University of Missouri— Columbia School of Medicine, Columbia, Missouri 65212
Received 2 September 1997/Accepted 15 October 1997
chkYB-2 is a sequence-specific, single-stranded DNA binding chicken Y-box protein that promotes Rous sarcoma virus long terminal repeat (RSV LTR)-driven transcription in avian fibroblasts. The DNA-binding domain of chkYB-2 has been mapped by characterizing the DNA binding properties of purified recombinant chkYB-2 mutant polypeptides. The data indicate that the invariant cold shock domain (CSD) is necessary but not sufficient for association with DNA and suggest that another conserved region, adjacent to the carboxyl boundary of the CSD, plays a role in high-affinity DNA binding. chkYB-2 binds to a tandem repeat of the 5*-GTACCACC-3* motif on the RSV LTR. Mutational analysis of this recognition sequence revealed the requirement of an essentially unaltered template for both high-affinity binding by chkYB-2 as well as maximal transcriptional activity of the RSV LTR in vivo. The single-stranded DNA binding activity of chkYB-2 is augmented by Mg21. The possible significance of this finding for transactivation by a single-strand DNA
binding protein is discussed.
The chicken Y-box protein, chkYB-2, is a sequence-specific single-stranded DNA binding protein that binds the octanucle-otide motif 59-GTACCACC-39 present on the noncoding strand of the Rous sarcoma virus (RSV) long terminal repeat (LTR) (7). chkYB-2 is expressed abundantly in avian fibro-blasts and muscle tissue, the mesenchymal-lineage host cells most permissive to infection and tumor formation by RSV. This property combined with its ability to function as a potent activator of RSV LTR-driven transcription in avian fibroblasts suggests an important role for this protein in the virus life cycle (40). We reported earlier on the cloning and characterization of chkYB-1b, another Y-box protein that closely resembles chkYB-2 in structure as well as in its ability to interact with specific motifs in the RSV enhancer (21). Recently, we also described the cloning of chkYB-1 homology protein, a poten-tial regulator of Y-box transcription factors (31).
The Y-box proteins are a new class of DNA and RNA binding factors that have been shown to function as both tran-scriptional and translational regulators of gene expression (39, 47, 48). Genes encoding the eukaryotic Y-box proteins have been isolated from Xenopus, chicken, mouse, rat, and human cells. The modular nature of these proteins resembles that of proteins from the well-characterized families of transcription factors. What is unique, however, is the wide range of nucleic acid structures to which Y-box proteins have been reported to bind (2, 8, 9, 16, 18, 26, 34, 36, 42).
The nucleic acid binding properties of Y-box proteins are thought to reside primarily in the highly conserved cold shock domain (CSD) (14, 34, 42, 46). The CSD has been described to recognize diverse double-stranded motifs, especially sites with purine/pyrimidine asymmetry between strands, as well as
dif-ferent single-stranded DNA sequences, particularly pyrimi-dine-rich ones. The CSD also contains the RNA binding motif RNP-1 (28). While the Y-box proteins share a near identity over the CSD, they vary widely at their amino termini. The carboxyl termini, while diverse in primary amino acid se-quence, still retain the charge-zipper motif wherein acidic and basic residues are organized as alternating islands (34).
Compelling evidence linking Y-box proteins to transcrip-tional regulation is accumulating, as the reports on roles played by single-stranded DNA binding proteins in activating tran-scription. Currently, information on the domain mapping of eukaryotic Y-box proteins is largely limited to work done on the Xenopus proteins FRGY1 and FRGY2 (4, 43), where it has been demonstrated that the functions of DNA binding, trans-activation, and multimerization can be localized to different domains. In this report, we describe the DNA binding prop-erties of mutant polypeptides derived from chkYB-2. The re-sults indicate that similar to classic CSD-containing proteins, chkYB-2 has an absolute requirement of the CSD for its DNA binding ability. However, mutant proteins lacking the carboxyl-tail domain entirely or partially either were incapable of bind-ing DNA or bound with markedly lower affinity, suggestbind-ing that while the CSD is necessary, it is not sufficient for high-affinity DNA binding.
59-GTACCACC-39, the octanucleotide recognition motif for chkYB-2, is present as a nearly contiguous direct repeat on the noncoding strand of the RSV LTR. In this report, we also present the results of mutagenesis analysis of single-stranded oligonucleotides bearing this motif and correlate the nature of chkYB-2 interactions observed with these mutants in vitro with the transcriptional activity of RSV LTR reporter constructs carrying identical mutations.
MATERIALS AND METHODS
chkYB-2 mutants.Isolation of the chkYB-2 cDNA clone and its transfer into the bacterial expression vector pMAL-c2 have been described previously (7). The deletion mutants are named according to the amino acids they retain. The derivative denoted with aDlacks the segment delineated by the numbered amino * Corresponding author. Mailing address: Molecular Microbiology
and Immunology, University of Missouri—Columbia School of Med-icine, M616 Medical Sciences Bldg., Columbia, MO 65212. Phone: (573) 882-7139. Fax: (573) 882-4287. E-mail: guntaka@showme .missouri.edu.
900
on November 9, 2019 by guest
http://jvi.asm.org/
acids. Mutant 1-169 was created by the excision of a PstI-PstI fragment of pMAL-c2 YB-2 followed by religation. Mutant 1-167 was constructed by first digesting mutant 1-169 with PstI and then using T4 DNA polymerase to polish the overhang generated by PstI. Religation then induces a frameshift such that out-of-frame nonsense codons are translated after amino acid 167 until a termi-nation codon is encountered. YB-2 mutant 1-162 was generated equivalently except that digestion was with EcoNI and end filling was done with the Klenow fragment of DNA polymerase I. Religation then induces a frameshift as de-scribed above. To obtain mutant 1-155, the EcoRI-ApaI fragment from the 1-169 construct was first treated with T4 DNA polymerase to polish the ApaI 39
overhang and then subcloned in pMAL-c2.
The internal deletion mutant D158-222 was derived directly from plasmid pMAL-c2 YB-2 by cutting with the restriction enzyme ApaI, which has two sites sharing the same reading frame in the carboxyl-terminal region of the YB-2 cDNA. The N-terminal deletion mutants were constructed by first generating a PCR product. The primers used are shown in Table 1. Primers A and B were used as the forward and reverse primers, respectively, for mutant YB-2 (75-230). YB-2 (75-298) was constructed by using primers A and C. Primer pairs D-C and E-C were used for generating the YB-2 (121-298) and YB-2 (158-298) products, respectively. The PCR products were gel eluted, digested with BamHI, and cloned into the pMAL-c2 vector. All downstream PCR primers had a transla-tional stop codon incorporated in the reading frame.
Protein preparations.Proteins from all constructs were expressed in
Esche-richia coli TB1 as described previously (21). All plasmid constructs express hybrid
proteins, with the maltose binding protein (MBP) of approximately 40 kDa forming the NH2terminus in all constructs. The MBP fusion polypeptides were purified on amylose column as described earlier (21) and used in all gel shift assays. When necessary, the fusion protein was cleaved with factor Xa essentially as described by the manufacturer (New England Biolabs). All the experiments reported here have been repeated with at least two to three independent prep-arations of proteins.
DNA binding assays.Single-stranded oligonucleotides either containing the chkYB-2 wild-type binding site on the RSV LTR or carrying single- or double-point mutations or deletions in the core sequence were synthesized. These sequences are shown in Table 2. Electrophoretic mobility shift assays were carried out as described earlier (23). Binding reaction mixtures (total volume, 20
ml) included 0.1 ng of 32P 59 end-labeled synthetic oligonucleotide probe (;30,000 cpm), 2mg of poly(dI-dC)z(dI-dC), and 6 to 100 ng of partially purified Mal–chkYB-2 fusion or its mutant derivatives. The binding buffer con-sisted of 50 mM Tris-HCl (pH 8.0), 75 mM NaCl, 3 mM MgCl2, 100mg of bovine serum albumin per ml, and 5% (vol/vol) glycerol. The reaction mixtures were incubated for 20 min at room temperature, applied directly onto prerun nonde-naturing 6% polyacrylamide gels, and electrophoresed in Tris-glycine buffer as described earlier.
Binding assays which examined the effects of other divalent cations like Zn21, Mn21, or Ca21 or the polyvalent cation spermidine on DNA binding were carried out similarly except for the addition of the specified ion(s) as indicated in the figure legends. Binding reactions that compared the affinities of the chkYB-2 fusion protein to different oligonucleotides with mutations in the binding site were carried out after first equalizing the specific activities of all the labeled probes.
Cell culture.Chicken embryo fibroblasts were cultured in medium 199 sup-plemented with 10% tryptose phosphate broth, 10% calf serum, and 1% chicken serum in plastic dishes at 37°C and 5% CO2in humidified air as reported earlier (21).
Reporter plasmid constructs.The reporter vectors carrying the chloramphen-icol acetyltransferase (CAT) gene under the control of the RSV LTR having point mutations in the chkYB-2 recognition site were constructed by using PCR-mediated mutagenesis as described earlier (40). The construction of E4 Del CAT, carrying a deletion in the E4 region, has also been described earlier (40). The incorporation of mutations in each of these constructs at the intended sites was confirmed by double-strand sequencing of the mutant constructs. Chicken embryo fibroblasts in the mid-log phase of growth were then transfected with 1
mg of each of these plasmids along with 1.0mg of pSVGal as an internal control, using Lipofectamine as instructed by the manufacturer (Gibco BRL). CAT assays were performed with equal amounts of extracts from these cells, normal-ized forb-galactosidase activity (11, 15). Transcriptional activity of each of the LTR constructs was quantified by scraping the silica gel corresponding to the
acetylated chloramphenicol spots seen on the X-ray autoradiogram and counting it in the scintillation liquid. All of these experiments were repeated at least three times, and the averages of the results are presented.
RESULTS
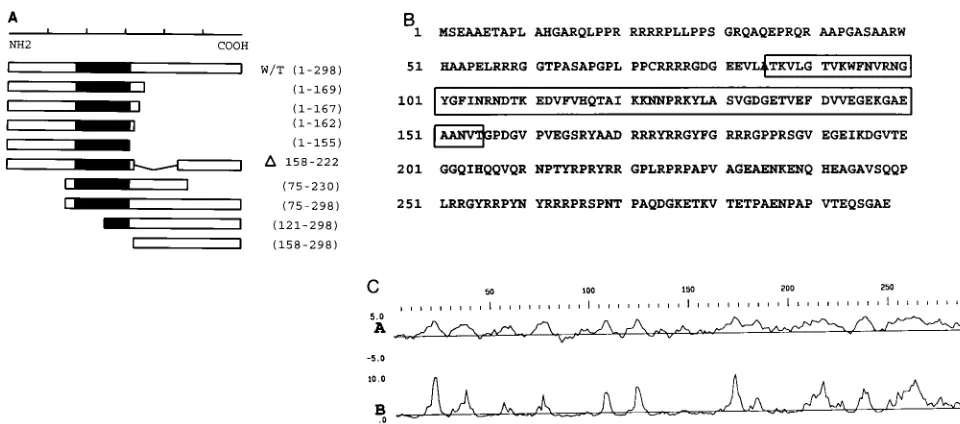
chkYB-2 binds to several single-stranded motifs on the non-coding strand of the RSV LTR and acts as a potent activator of RSV LTR-driven transcription in avian fibroblasts. To lo-calize the polypeptide region that is responsible for site-spe-cific binding to single-stranded DNA, we performed deletion mutagenesis of YB-2 cDNA. Figure 1A is a line diagram of the mutants that were created by using standard recombinant DNA methods. Except for one nested deletion, YB-2 (D158-222), all constructs were either NH2-terminal or
COOH-ter-minal deletions. The YB-2 (75-230) mutant has deletions at both termini.
Wild-type and mutant YB-2 polypeptides expressed in E. coli as MBP fusion proteins were column purified on amylose, and
TABLE 1. Sequences of primers used for PCR-generated YB-2 mutants Primer Sequence (59-39)
[image:2.612.307.549.286.551.2]A...GACATGGATCCCGCCGCAGGGGGAGACGGAGAAG B ...GACATGGATCCTTAGACCGGGGCAGGGCGTGGA C ...GACATGGATCCGCCTGAGAACCTGACATTACTC D...GACATGGATCCAAGAAGAACAACCCACGGAAGT E ...GACATGGATCCGATGGAGTCCCTGTAGAAGGCA
TABLE 2. Mutational analysis of the chkYB-2 binding motifa
Oligo-nucleotide Sequence Binding
E4C1 59-GGCACGA TCGTACCACCTT AC-39 1111
E4C2 M108 59-GGCACGA ACGTACCACCTT AC-39 11
E4C2 M109 59-GGCACGA TAGTACCACCTT AC-39 1
E4C2 M110 59-GGCACGA TGATACCACCTT AC-39 1
E4C2 59-GGCACGA TCATACCACCTT AC-39 11
E4C2 M101 59-GGCACGA TCCTACCACCTT AC-39 1111
E4C3 59-GGCACGA TCGAACCACCTT AC-39 2
E4C2 M102 59-GGCACGA TCGGACCACCTT AC-39 2
E4C2 M103 59-GGCACGA TCGTGCCACCTT AC-39 2
E4C AM3 59-GGCACGA TCGCGCCACCTT AC-39 2
E4C AM2 59-GGCACGA TCGTACGATCTT AC-39 2
E4C4 59-GGCACGA TCGTATTACCTT AC-39 2
E4C2 M104 59-GGCACGA TCGTACCGCCTT AC-39 2
E4C2 M105 59-GGCACGA TCGTACC*CCTT AC-39 2
E4C2 M106 59-GGCACGA TCGTACCATCTT AC-39 1111
E4C2 M111 59-GGCACGA TCGTACCATTTT AC-39 2
E4C2 M107 59-GGCACGA TCGTACCACCAA AC-39 1111
X2/E4C1 59-ATCGTACCACCTTAAAAATCGTACCACCTTAAA-39 1111
LTR2106/
2135 59
-ACGATCGTACCACCTTACTACCACCAATCG-39 1111
E4M103-1C 59- ATCGTGCCACCTTACTACCACCAATCG-39 11
E4M103-2C 59- ATCGTACCACCTTACTGCCACCAATCG-39 11
E4M103-3C 59- ATCGTGCCACCTTACTGCCACCAATCG-39 1/2
E4M106-1C 59- ATCGTACCATCTTACTACCACCAATCG-39 1111
E4M106-2C 59- ATCGTACCACCTTACTACCATCAATCG-39 1111
E4M106-3C 59- ATCGTACCATCTTACTACCATCAATCG-39 1111
aSingle-stranded oligonucleotides were synthesized. The first set includes all
oligonucleotides from E4C1 through E4C2 M107. E4C1 is the wild-type se-quence corresponding to the region from2103 to2123 on the noncoding strand of the RSV LTR. It contains the 12-mer, 59-TCGTACCACCTT-39motif (bold-face), a consensus sequence arrived at after aligning a panel of RSV LTR oligonucleotides that were described earlier to bind chkYB-2. Oligonucleotides E4C2-M101 to M111, E4C2, E4C3, E4C4, E4C2 AM2, and E4C2 AM3 represent the different mutations that were introduced. Mutant residues are shown with a double underline. The asterisk represents a deletion.1111binding in this set refers to binding equivalent to that obtained with the wild-type E4C1. Binding of mutants is represented relative to this level. The second set includes X2/E4C1 and the others listed below it. These oligonucleotides differ from the first set in having a tandem repeat of the binding motif. X2/E4C1 is a synthetic construct representing a perfect duplication of the 12-mer, 59-TCGTACCACCTT-39. LTR
2106/2135 represents the2106 to2135 region on the noncoding strand of the RSV LTR. There is a 59-CTACCACC-39motif (underlined) immediately down-stream of the octamer, 59-GTACCACC-39, which could serve as an additional motif for chkYB-2 binding. The oligonucleotides E4M103-1C to -3C and E4M106-1C to -3C incorporate point mutations in either the 59or the 39octamer motif or both motifs simultaneously.1111binding in this set refers to binding obtained with the wild-type LTR2106/2135 oligonucleotide. Binding of mu-tants is represented relative to this level.
on November 9, 2019 by guest
http://jvi.asm.org/
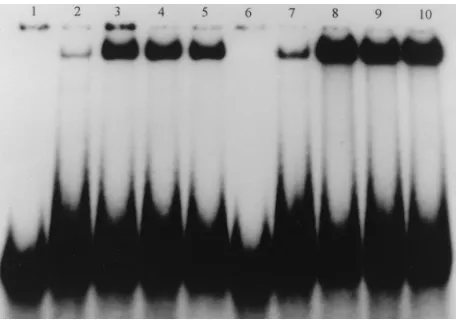
aliquots electrophoresed on sodium dodecyl sulfate (SDS)-polyacrylamide gels (data not shown). In all cases, a protein of the expected size was obtained. Wild-type and mutant YB-2 proteins were examined for DNA binding activity by carrying out gel shift assays, using a radiolabeled single-stranded re-sponse element from the RSV LTR (LTR2106/2135 [Table 2]), shown in this report to be the maximum affinity binding site of chkYB-2.
In chkYB-2, the CSD extends from amino acids (aa) 86 to 155 (Fig. 1B). The first construct that we made was YB-2 (D158-222). The internal deletion of 66 aa in the carboxyl-tail domain created a mutant with intact amino-terminal and CSDs. If the CSD is sufficient for DNA binding, this mutant polypeptide would be expected to bind DNA. As seen in Fig. 2 (lane 9), no binding to DNA was observed in an in vitro DNA binding assay. DNA binding was tested under different salt conditions, using up to 2 mg of the protein prepared from several independent clones, as well as after cleavage of the MBP fusion with factor Xa (data not shown). No binding was detected under any of these conditions. There are two possible explanations for the total abrogation of DNA binding upon the deletion of these 66 aa. This 66-aa stretch is immediate to the carboxyl side of the carboxyl boundary (aa 155) of the CSD. Given the proximity to the well-structured CSD, this deletion could have resulted in a major functional deformity in the protein leading to loss of DNA binding. An alternative expla-nation for the nonfunctionality of this mutant in terms of DNA binding is that the deleted aa 158 to 222, or at least some of them, are part of the minimum DNA binding domain, which in chkYB-2 could extend beyond the strict confines of the con-served residues of the CSD.
To map the carboxyl boundary of the DNA binding activity of chkYB-2, we constructed the YB-2 (1-169) and YB-2 (158-298) mutants and assayed their ability to bind DNA. YB-2 (158-298) expresses only the carboxyl-tail domain of YB-2, the entire CSD and amino terminus having been deleted. As seen
in Fig. 2 (lane 13), the YB-2 (158-298) mutant did not bind DNA. This result indicated that the carboxyl half of the protein cannot independently bind DNA. Although the residues in the 158–222 region are critical for DNA binding, they of them-selves do not confer DNA binding activity in the absence of the CSD.
Mutant YB-2 (1-169) retains the CSD and, unlike YB-2 (D158-222), has 14 residues to the immediate carboxyl side of the CSD intact. As seen in Fig. 2 (lane 5), this polypeptide
FIG. 1. (A) Diagram of chkYB-2 mutant constructs. Wild-type (w/t) chkYB-2 (298 aa) is shown schematically. The filled-in region represents the highly conserved CSD. The various amino-terminal and carboxyl-terminal deletion mutants, and one internal deletion mutant (see Materials and Methods), are shown schematically below the wild type. The mutants are named according to the amino acids they retain. The derivative with aDlacks the segment delineated by the numbered amino acids. (B) Predicted 298-aa sequence of chkYB-2 protein. The boxed region extending from aa 86 to 155 is the invariant, nucleic acid binding CSD. Arginine clusters are present in the hydrophilic carboxyl-tail domain, while the amino-terminal region is rich in proline. (C) Protein structure analysis of chkYB-2 carried out with the Genetics Computer Group software package. Plots A and B represent the Kyte-Doolittle hydrophilicity predictions and surface probability (Emini method) predictions, respectively.
FIG. 2. Localization of the domains responsible for DNA binding. chkYB-2 deletion mutants were expressed as MBP fusion proteins in E. coli and partially purified on amylose columns. The ability of these mutant polypeptides to bind DNA was assessed by carrying out gel shift assays as described in Materials and Methods. A single-stranded DNA oligonucleotide (LTR2106/2135 [Table 2]), corresponding to the2106 to2135 region of the noncoding strand of the RSV enhancer and bearing a chkYB-2 recognition motif, was end labeled and used as the probe. Proteins in the individual binding reactions are as follows: lane 1, probe alone; lane 2, 100 ng of MBP; lane 3, 100 ng of factor Xa-cleaved MBP–YB-2 protein. MBP fusion proteins were used in all remaining binding reactions. Lane 4, wild-type chkYB-2 (50 ng); lane 5, chkYB-2 (1-169) (200 ng); lane 6, chkYB-2 (1-167) (300 ng); lane 7, chkYB-2 (1-162) (300 ng); lane 8, chkYB-2 (1-155) (300 ng); lane 9, chkYB-2 (D158-222) (300 ng); lane 10, chkYB-2 (75-230) (300 ng); lane 11, chkYB-2 (75-298) (300 ng); lane 12, chkYB-2 (121-298) (300 ng); lane 13, chkYB-2 (158-298) (300 ng); lane 14, probe alone.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.48.528.65.277.2]bound DNA, albeit with a 10-fold-lower affinity than the wild-type YB-2 protein (lanes 3 and 4, containing factor Xa-cleaved and MBP fusion proteins, respectively). The partial restoration of DNA binding activity upon preservation of residues 156 to 169 supports our belief that the domain subserving DNA bind-ing in YB-2 extends beyond the CSD. Construction of mutants with their carboxyl termini at different positions between aa 169 and 298 is currently under way. DNA binding assays with these proteins would help in defining precisely the carboxyl boundary of the polypeptide displaying full restoration of DNA binding activity.
We were curious to know if all of the 14 residues in the short carboxyl tail of YB-2 (1-169) were necessary for DNA binding activity or whether small truncations in this region would be tolerated. Progressive C-terminal deletions of YB-2 (1-169) yielded the mutants YB-2 (1-167) and YB-2 (1-162). Gel shift assays were carried out with these mutant polypeptides to determine their ability to bind DNA. As seen in Fig. 2 (lanes 6 and 7), YB-2 (1-167) bound less avidly than YB-2 (1-169) and YB-2 (1-162) bound even less than YB-2 (1-167), showing that progressive deletions were deleterious to DNA binding. This effect was clearly seen when DNA binding was totally lost with the YB-2 155) mutant (lane 8). The deletion in YB-2 (1-155) removes the entire carboxyl-tail domain of YB-2 up to the carboxyl boundary of the CSD. The total absence of DNA binding with the YB-2 (1-155) protein is similar to the behavior of the YB-2 (D158-222) mutant. Taken together, these results indicate that in chkYB-2, the CSD is necessary but not suffi-cient for DNA binding activity.
Interestingly, in gel shift assays, the DNA-protein complexes formed by the YB-2 (1-169), (1-167), and (1-162) mutants migrated more slowly than the complexes formed by the full-size protein. The YB-2 (1-162) protein in fact forms two com-plexes, one a faster-migrating complex similar to the wild-type YB-2 protein and the other a slower-migrating complex similar to those formed by the YB-2 (1-169) and (1-167) proteins. This was surprising, considering the fact that on SDS-polyacryl-amide gels the migration of these polypeptides was propor-tional to their molecular weights. This pattern was reproduced even when DNA binding assays were carried out with factor Xa-cleaved protein, ruling out any artifacts introduced by the MBP moiety. The formation of multimeric complexes is a likely explanation for the above observation. Further experi-ments are, however, required to demonstrate unequivocally if these mutant proteins do indeed exist as multimers, either in solution or upon binding DNA.
We also constructed YB-2 mutants lacking portions of their N termini. The mutant YB-2 (121-298) disrupts the CSD and as expected showed negligible DNA binding ability (Fig. 2, lane 12). The mutant YB-2 (75-298), while carrying a large deletion at its amino terminus, still retains the entire CSD and could be expected to bind DNA. However, this mutant protein also showed minimal ability to bind DNA. A possible explana-tion for this could be the proximity of the deleexplana-tion to the CSD. Given the behavior of this mutant, it was not surprising that the double-deletion mutant YB-2 (75-230) demonstrated no ability to bind DNA (Fig. 2, lane 10). We also tested the ability of the YB-2 (1-169) mutant to bind a series of unrelated single-stranded and double-single-stranded DNA oligonucleotides to which wild-type YB-2 had not bound. No binding was observed (data not shown). We also examined the ability of this mutant to bind the different RSV LTR mutant oligonucleotides shown in Ta-ble 2. The relative binding affinity of YB-2 (1-169) to these mutants (data not shown) always paralleled the results ob-tained with the full-size protein, indicating that although YB-2 (1-169) binds with less affinity than the wild-type YB-2, there is
no relaxation in the sequence specificity. The components of the YB-2 protein molecule that are involved in sequence-spe-cific recognition probably reside within the 1–169 region.
In summary, the DNA binding studies with the chkYB-2 mutants described above indicate that the CSD is important for DNA binding and that the carboxyl-terminal charge-zipper domain has no independent ability to bind DNA. The CSD mediates sequence-specific recognition as well as binding to single-stranded DNA. It is also evident that unlike the bacte-rial cold shock proteins wherein the CSD alone is adequate for DNA binding, the residues that make up the CSD in chkYB-2 are necessary but not sufficient for DNA binding. Apparently, the residues to the carboxyl side of CSD, even if not part of the binding domain, contribute to the generation of stable com-plexes with DNA, at least in vitro.
Some Y-box proteins are known to bind DNA more avidly in the presence of magnesium (18). Magnesium also appears to play a role in the nucleic acid interactions of several other RNA binding proteins (27). We carried out gel shift assays to examine the effects of different concentrations (0 to 20 mM) of several divalent cations (Mg21, Ca21, Mn21, and Zn21), as well as spermidine, a polyvalent cation. The results of these gel shift assays are shown in Fig. 3. Addition of magnesium chlo-ride to final concentrations of 3 to 10 mM in the binding reaction increased DNA binding more than 10-fold, with max-imum effect seen at 5 mM; 20 mM MgCl2, however, had an
inhibitory effect. A similar effect was noted with spermidine. While 3 or 5 mM CaCl2 stimulated binding severalfold,
con-centrations of 10 mM and above were inhibitory. MnCl2at 3
and 5 mM promoted binding, although less than for the other ions. As found for CaCl2, MnCl2concentrations of 10 mM or
more were inhibitory. In contrast to the stimulatory effects of these cations, the addition of even 3 mM ZnCl2was inhibitory
to the formation of DNA-protein complexes, with higher ionic strengths essentially eliminating binding. Figure 3 (lane 2) shows the binding of 6 ng of chkYB-2 protein to the radiolabeled LTR oligonucleotide2106/2135, in the absence of any diva-lent cation. Lanes 3 and 4 show the remarkable increase in DNA binding upon the addition of 5 mM MgCl2 or
spermi-dine, respectively. Lanes 6 and 7 show the stimulation of
FIG. 3. Effects of cations on DNA binding. The RSV LTR oligonucleotide LTR (2106/2135) was end labeled, and its binding to 6 ng of chkYB-2 fusion protein was assayed in gel shift experiments carried out as described in Materials and Methods. Binding reactions were identical except for the presence or ab-sence of the following cations: lane 1, probe alone; lane 2, no MgCl2; lane 3, MgCl2(5 mM); lane 4, spermidine (5 mM); lane 5, MgCl2and spermidine (5 mM each); lane 6, CaCl2(3 mM); lane 7, MnCl2(3 mM); lane 8, ZnCl2(3 mM); lane 9, ZnCl2and MgCl2(3 mM) each; lane 10, ZnCl2, MgCl2, and spermidine (3 mM each).
on November 9, 2019 by guest
http://jvi.asm.org/
binding in the presence of 3 mM CaCl2and 3 mM MnCl2,
respectively. ZnCl2at 3 mM inhibited binding (lane 8). This
inhibition was, however, neutralized upon the addition of ei-ther 3 mM MgCl2(lane 9) or 3 mM each MgCl2and
spermi-dine (lane 10) to the reaction.
The exact significance of the effect of a cationic environment on chkYB-2–DNA interactions is not known. We are not aware of any specific metal ion binding motifs on the chkYB-2 protein. We were curious to know if these results could be reproduced with any of the YB-2 mutants that we have made. We tested the DNA binding activity of the YB-2 (1-169) mu-tant protein either in the absence of cations or in the presence of MgCl2, spermidine, or CaCl2. As shown in Fig. 4, the
addi-tion of 5 mM MgCl2(lane 3), 5 mM spermidine (lane 4), or 3
mM CaCl2(lane 5) significantly promoted DNA binding
com-pared to DNA binding carried out in the absence of any of these ions (lane 2). Lanes 2 to 5 contained 10 ng of the protein. The same effect was repeated when 50 ng of the protein was used in each binding reaction (lanes 7 to 10).
Effects of point mutations in the core binding site for chkYB-2.We have shown earlier that the E4 region in the RSV LTR is important for maximal enhancer activity (40). We also reported that the recognition motif for chkYB-2, the octamer 59-GTACCACC-39, is located in this region. Also, transfection experiments using E4-deleted LTR constructs and chkYB-2 antisense oligonucleotides had demonstrated that the ability of chkYB-2 to act as an activator was mediated primarily through this octanucleotide motif. Our earlier work had shown that this protein bound with various affinities several different single-stranded oligonucleotides spanning the RSV LTR. We aligned the sequences of all these oligonucleotides to which chkYB-2 had bound and looked for a consensus sequence. This com-parison revealed that the 12-mer 59-TCGTACCACCTT-39is the common motif. This is essentially the previously described octamer 59-GTACCACC-39extended by two nucleotides each in the 59and 39directions.
The 21-mer oligonucleotide E4C1, bearing this motif and corresponding to the region from2103 to2123 on the
non-coding strand of the RSV LTR, was hence used as the wild-type binding motif, and systematic point mutations spanning the entire motif were introduced (Table 2). End-labeled oli-gonucleotides, adjusted for specific activity, were then used in gel shift assays. A summary of the binding results is presented in Table 2. The gel shift assay shown in Fig. 5 is representative of some of the oligonucleotides used. It is evident from these results that the binding of YB-2 to its recognition motif was abolished upon the introduction of any mutation in the core octamer, except for the mutant oligonucleotide E4C2 M101, where replacement of G with a C at position 3 appeared to be well tolerated. The other exception was the mutant E4C2 M106, where replacement of C with a T at position 9 did not affect binding. However, when the adjacent C was also re-placed by a T, to yield the double mutant E4C2 M111, binding was abolished. Nucleotides at positions 11 and 12 did not appear to be critical, as shown by binding equivalent to wild-type binding by the mutant E4C2 M107. Binding to oligonu-cleotides with mutations at positions 1 and 2 (E4C2 M108 and E4C2 M109, respectively) was significantly less than binding to E4C1. These results indicate that the single-stranded DNA binding protein chkYB-2 binds its ligand in a sequence-specific manner and that maximum binding affinity requires the pres-ence of at least the 59-TCGTACCACC-39decamer motif.
In our earlier report (40), we had remarked on the ability of chkYB-2 to bind more than one site on the RSV LTR and had suggested that the appearance of multiple, slower-migrating complexes in gel shift assays carried out with the full LTR as the probe was probably due to occupancy of the other sites by additional molecules. To confirm this effect directly, we
[image:5.612.56.284.68.228.2]de-FIG. 4. Effects of cations on chkYB-2 (1-169) binding to DNA. The oligo-nucleotide LTR2106/2135 was end labeled, and DNA binding assays were carried out with the chkYB-2 (1-169) mutant polypeptide. Binding mixtures were incubated at room temperature for 20 min, loaded onto prerun, nondenaturing 6% polyacrylamide gels, and electrophoresed in Tris-glycine buffer. Binding reactions for lanes 2 to 5 and lanes 7 to 10 were carried out with 10 and 50 ng, respectively, of the chkYB-2 (1-169) protein. The binding buffer always con-tained 75 mM NaCl. The concentrations of divalent and polyvalent cations included in the different reactions are as follows: lanes 1 and 6, no protein added, 5 mM MgCl2; lanes 2 and 7, no MgCl2; lanes 3 and 8, MgCl2(5 mM); lanes 4 and 9, spermidine (5 mM); lanes 5 and 10, CaCl2(3 mM).
FIG. 5. Mutational analysis of the chkYB-2 binding motif in RSV LTR. A series of point mutations was introduced in the single-stranded DNA oligonu-cleotide E4C1 (see Table 2 for sequences of oligonuoligonu-cleotides). E4C1 represents the2103 to2123 region in the noncoding strand of the RSV LTR and contains the octamer 59-GTACCACC-39, previously described as the recognition motif for chkYB-2. All oligonucleotides were end labeled and equalized for specific activity, using appropriate amounts of corresponding unlabeled DNA. Gel shift assays were carried out as described in Materials and Methods. chkYB-2 fusion protein (6 ng) was used in each binding reaction. The autoradiogram is repre-sentative of some of the oligonucleotides used. The binding results are also summarized in Table 1. Lane 1, wild type, E4C1; lane 2, E4C2 M103; lane 3, E4C2 AM2; lane 4, E4C2 M109; lane 5, E4C2 M107; lane 6, E4C2 M106. Two oligonucleotides were synthesized to examine the nature of protein-DNA inter-action in the presence of a double motif for chkYB-2 binding. The 33-mer oligonucleotide X2/E4C1 (lane 7) is a synthetic construct carrying a perfect repeat of the 12-mer 59-TCGTACCACCTT-39motif. The oligonucleotide LTR
2106/2138 (lane 8) is a 30-mer corresponding to the2106 to2135 region on the RSV enhancer and contains a tandem repeat of the 59-GTACCACC-39
octamer. Lane 9 represents binding to another 30-mer oligonucleotide having the same sequence as LTR2106/2135 described above but synthesized in the anti-parallel direction, i.e., 59-GCTAACCACCATCATTCCACCATGCTAGCA-39.
on November 9, 2019 by guest
http://jvi.asm.org/
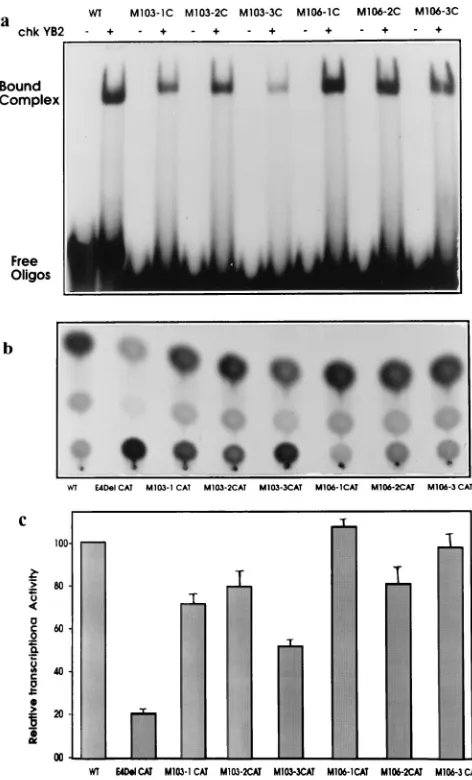
signed the 30-mer oligonucleotide X2/E4C1 (Table 2), which is a direct repeat of the YB-2 binding motif. As shown in the gel shift assay (Fig. 5, lane 7), chkYB-2 bound avidly to this oli-gonucleotide, forming two DNA-protein complexes. The slow-er-migrating complex is a minor component and probably rep-resents more than one molecule of YB-2 complexed to DNA. An examination of the sequence of the noncoding strand of the RSV LTR, immediately to the 39side of the octamer motif 59-GTACCACC-39(2112 to2119), revealed the presence of an almost identical 59-CTACCACC-39(2123 to2130) motif. Also, the gel shift assays described above had shown that the replacement of the nucleotide G in the motif with a C, as in the mutant E4C2 M101, did not decrease the affinity of chkYB-2 binding (Table 2). Hence, the2112 to2130 region of the RSV LTR can be viewed as providing two potential sites for high-affinity binding by chkYB-2. To examine the high-affinity of chkYB-2 to DNA bearing such a double motif, we synthesized the oligonucleotide LTR 2106/2135 (Table 2). Unlike the oligonucleotide E4C1 (extending from2103 to2123), which has only the first octamer motif, this new oligonucleotide, by extending from2106 to2135, incorporates both the 59and 39 octamer motifs. The gel shift assay (Fig. 5) showed that the ability of chkYB-2 to bind LTR2106/2135 (lane 8) was sev-eralfold greater than that observed with E4C1 (lane 1). As found for X2/E4C1, another slower-migrating complex was also seen as a minor component. Surprisingly, the size of the major protein-DNA complex formed with the double-motif oligonucleotide was comparable to the one found with the single-motif DNA. This finding suggests that only a monomer was complexing with the LTR2106/2135 oligonucleotide to form the major retarded species. These results suggest that it would be more accurate to consider these 59-GTACCACC-39 repeats as providing two half-sites, rather than two full motifs, for chkYB-2 binding. It is also evident that binding to this native double-octamer motif on the RSV LTR is comparable to the binding observed with the synthetic construct X2/E4C1, carrying a perfect repeat of the 12-mer 59-TCGTACCACCT T-39motif (lane 7). These experiments with X2/E4C1 and LTR 2106/2135 were done with limiting amounts (6 ng) of the chkYB-2 protein. When these experiments were repeated with progressively larger amounts of protein, there was only a slight increase in the amount of the slower-migrating complex (data not shown). Since the formation of the faster-migrating species is an apparent prerequisite for the formation of the second, more slowly migrating species, these probably represent one and two molecules of chkYB-2, respectively. However, consid-ering the low rate at which the slower-migrating complex forms, the binding of the second molecule is apparently not cooperative in nature. Curiously, chkYB-2 demonstrated good binding to another 30-mer oligonucleotide that had the same sequence as the oligonucleotide LTR2106/2135 except that it was synthesized in the antiparallel direction (Fig. 5, lane 9). We are not aware whether this apparent ability to bind the recognition motif without regard to its polarity has been re-ported earlier for other single-stranded DNA binding proteins. Considering the above results, which we obtained with the LTR 2106/2135 oligonucleotide, we wished to explore by mutational analysis whether point mutations introduced in one of the motifs adversely affected the ability of chkYB-2 to in-teract with the second motif. Toward this end, we designed mutant oligonucleotides (E4M series [Table 2]) representing the2109 to2135 region, such that identical point mutations were introduced in either the 59 or 39 octanucleotide motif alone or in both motifs simultaneously. One set of mutant oligonucleotides was designed based on the negligible protein binding that we had observed with the E4C2 M103 mutant
[image:6.612.310.546.67.457.2](Fig. 5, lane 2). The same A3G change was made in either the 59 or 39 motif or both motifs (E4M103-1C, E4M103-2C, or E4M103-3C, respectively). The other set of mutants was based on the E4C2 M106 mutant, wherein a C3T change apparently did not affect the formation of DNA-protein complexes (Fig. 5, lane 6). The oligonucleotides E4M106-1C, E4M106-2C, and E4M106-3C thus represent a C3T change in the 59, 39, and both 59and 39octanucleotide motifs, respectively. The results of chkYB-2 binding to these mutant constructs compared to the wild type are presented in Fig. 6a. chkYB-2 showed re-duced affinity for both M103-1C and M103-2C, indicating the contribution of both motifs for maximal binding. Not surpris-ingly, negligible binding was observed with the double mutant M103-3C. In contrast, binding to the E4M106 set of mutants was essentially unaffected.
FIG. 6. Contribution of both chkYB-2 recognition octamers to RSV LTR-driven transcription. (A) All mutant oligonucleotides (see Table 2 for nomen-clature and sequences) were end labeled and equalized for specific activity. Gel shift assays were carried out using 6 ng of chkYB-2 fusion protein in each reaction. Binding to the wild-type (WT) is shown for comparison. (B) A repre-sentative autoradiogram showing the relative transcriptional activity of a wild-type RSV LTR-CAT reporter construct in chicken embryo fibroblasts compared to constructs carrying either a deletion or point mutation in the chkYB-2 binding site on the RSV LTR. See Materials and Methods for details of mutations. (C) Quantitative data from three identical transient transfection experiments con-ducted independently. Details of constructs and quantitation are described in Materials and Methods.
on November 9, 2019 by guest
http://jvi.asm.org/
The experiments described thus far helped in defining the requirements of the chkYB-2 binding motif. However, the exact relevance of results obtained from in vitro DNA binding assays to chkYB-2 interactions with the RSV LTR in vivo had to be determined. One approach was to correlate the muta-tions that abrogated protein binding in vitro with alteramuta-tions in the in vivo transactivating potential of RSV LTR constructs carrying the same mutations in the YB-2 binding motif. To-ward this end, we constructed a series of mutant RSV LTR reporter vectors. Mutations were made only within the2112 to 2130 region described earlier as the site of two motifs for chkYB-2 binding. Furthermore, the point mutations intro-duced in these six constructs reflect exactly the changes made in either the 59 or 39 octamer motif or both binding motifs while designing the E4M series of mutant oligonucleotides.
Chicken embryo fibroblasts were chosen for the transfection experiments, as chkYB-2 is expressed abundantly in these cells and has been demonstrated to activate RSV LTR-driven tran-scription in these cells, primarily through its interaction with recognition motifs present within the 2112 to 2130 region (40). Cells in the mid-log phase of growth were transfected with 1mg of each of these plasmids, along with 1mg of the internal control plasmid, pSVGal. CAT assays were performed with protein extracts normalized tob-galactosidase activity. A representative autoradiogram (Fig. 6b) and the average results of three identical experiments (Fig. 6c) are presented. These results reveal that point mutations in either the 59or 39motif (M103-1CAT or M103-2CAT, respectively) reduced the tran-scriptional activity of the RSV LTR by about 20 to 25%. The decrease in transcriptional activity upon the introduction of point mutations in the two separate motifs is synergistic, as evidenced by the much greater reduction in transcriptional activity of the double mutant (M103-3CAT) compared with either single mutant alone. The activity of this double mutant (mutations are at positions 2114 and 2125) was, however, much higher than that observed with E4 Del CAT, an RSV LTR construct carrying a 14-nucleotide deletion (2114 to 2127) that encompasses both chkYB-2 binding motifs. A pos-sible interpretation of this finding is that although in vitro chkYB-2 bound negligibly to this double mutant (M103-3C [Fig. 6a]), in vivo, low-affinity interactions with this mutant YB-2 motif probably occur and contribute to transactivation of the LTR, albeit less efficiently. Even these low-affinity interac-tions apparently cannot take place when both motifs are com-pletely deleted, as in E4 Del CAT. Alternatively, this result could reflect the fact that protein-DNA interactions, other than those mediated by chkYB-2, contribute to transactivation from this deleted region. Figure 6c also shows that compared to the M103-CAT constructs, the M106-CAT constructs did not show significant reductions in transcriptional activity, with even the double mutant M106-CAT displaying transcriptional activity comparable to wild-type RSV LTR.
In summary, the results presented above show a correlation between the transcriptional activities of RSV LTR constructs carrying point mutations in the chkYB-2 binding motifs and the relative affinities of the corresponding mutant oligonucle-otides as assayed by DNA-protein complex formation in vitro. Additionally, our results also show that (i) both of the chkYB-2 recognition motifs contribute to RSV LTR-driven transcrip-tion and (ii) the interactranscrip-tion of chkYB-2 with these adjacent motifs is probably just additive and not cooperative in nature, since the reduction in transcription observed from the double mutant (M103-3CAT) was not more than the cumulative re-duction in transcriptional activities of the single mutants. These results are also in agreement with our interpretation of the chkYB-2 binding assays with LTR2106/2135, which
sug-gested that the2112 to2130 region is best viewed as provid-ing a sprovid-ingle high-affinity bindprovid-ing site for chkYB-2, with each octamer motif behaving as a half-site.
DISCUSSION
chkYB-2 was originally isolated by screening a chicken em-bryo fibroblast cDNA expression library by using a probe cor-responding to the U3 enhancer region of the RSV LTR (7). We have since demonstrated that purified, recombinant chkYB-2 specifically recognizes the 59-GTACCACC-39 single-stranded motif on the noncoding strand of the RSV enhancer and acts as an activator of RSV LTR-driven transcription in avian fibroblasts (40). An understanding of the mechanism by which a factor like chkYB-2, which binds only single-stranded DNA templates yet acts as a transcriptional activator, would be facilitated by studies delineating the functional domains in the protein involved in DNA binding, multimerization, and trans-activation. As a first step toward this, we report here the DNA binding properties of bacterially expressed and partially puri-fied, recombinant chkYB-2 mutant polypeptides to a single-stranded response element (2106 to2135) of the RSV en-hancer.
The specific DNA binding activities of several transcription factors have been localized to relatively small domains consist-ing of 60 to 100 aa. chkYB-2, an avian Y-box protein, is a 298-aa polypeptide. Like other Y-box transcription factors, chkYB-2 is characterized by the presence of the invariant CSD located in an intermediate position and flanked by variable amino and carboxyl-terminal domains. Wistow (46) initially proposed that the CSD is a structural motif involved in pro-tein-nucleic acid interactions. The CSD is 43% identical to CS7.4, the 70-aa major cold shock protein of E. coli (14). Unlike the Y-box proteins, the E. coli cold shock proteins do not have the additional tail domain. CS7.4, nevertheless, binds DNA in a sequence-specific manner, suggesting that this do-main is adequate for DNA binding. Studies on CspB (38), a cold shock protein of Bacillus subtilis, revealed the three-di-mensional structure of its nucleic acid binding domain and suggested that the structural organization of the CSD was best suited for interaction with single-stranded nucleic acids.
Mutagenesis analysis of Xenopus Y-box proteins had shown the CSD to be the primary domain responsible for both DNA and RNA binding (43). However, there are few data on the domain analysis of other eukaryotic Y-box proteins. Notwith-standing the strong conservation of the CSD, a remarkable feature of the Y-box proteins characterized thus far has been their ability to bind diverse double- and single-stranded DNA sequences, as well as RNA. It is hence likely that although the highly conserved amino acids in the CSD confer a structural framework for DNA binding, the determinants of binding specificity reside in the variable amino and carboxyl regions of the protein. The highly hydrophilic, carboxyl-tail domain (Fig. 1B) is a more likely candidate because, despite the divergence in primary amino acid sequence (e.g., only 50% identity be-tween chkYB-2 and chkYB-1b), the organization of the resi-dues into alternating clusters of acidic and basic resiresi-dues to create a charge-zipper motif is a feature that is conserved among all Y-box proteins.
Similarities between classic Y-box proteins and chkYB-2 predicted that DNA binding would be mediated by CSD. How-ever, a mutant chkYB-2 protein, lacking a 60-aa region in the carboxyl-tail domain, was found incapable of binding DNA. It therefore became interesting to determine the potential con-tribution of the carboxyl domain for interactions with DNA. The data presented in this study provide evidence that even in
on November 9, 2019 by guest
http://jvi.asm.org/
chkYB-2, the CSD is indeed indispensable but not sufficient for DNA binding. While low-affinity, site-specific binding is obtainable with carboxyl-domain truncation mutants that have an intact CSD and as few as 10 to 14 adjacent residues of the carboxyl tail, high-affinity binding requires that apart from the CSD, larger segments of the carboxyl domain remain intact in the protein.
Tafuri and Wolffe (43) had reported that progressive dele-tion of the carboxyl terminus of FRGY2, a Xenopus Y-box protein, resulted in a reduction in the number of complexes formed with DNA. Binding to DNA, however, remained spe-cific, even where the entire carboxyl-tail domain was removed. Removal of the CSD, however, led to a loss of specific DNA binding. They obtained similar results with the closely related FRGY1 protein and concluded that the CSD was essential for specific DNA binding, whereas the hydrophilic carboxyl-tail domain facilitated the formation of multiple protein-DNA complexes. It was also demonstrated that in both FRGY1 and FRGY2, the CSD was adequate for stimulation of transcrip-tion, both in vitro and in vivo (35, 42, 43).
In chkYB-2, the CSD extends from aa 86 to 155. However, we detected no DNA binding with the mutant chkYB-2 (1-155), which has an intact CSD but no carboxyl tail. Low-affinity DNA binding ability returned incrementally upon the progres-sive lengthening of the carboxyl tail, as evidenced by the com-plexes formed by the chkYB-2 mutants (1-162), (1-167), and (1-169). Significantly, we found no evidence of a relaxation in site specificity when the chkYB-2 (1-169) mutant was tested for its ability to bind several unrelated oligonucleotides, both sin-gle stranded and double stranded. While the lack of binding of the YB-2 (1-155) mutant could be explained by the proximity of the truncation to the carboxyl boundary of the CSD, the fact that even the chkYB-2 (1-169) mutant bound with only a 10-fold-lower affinity than the wild type suggests that the residues in the carboxyl domain beyond the CSD either are part of an extended DNA binding domain or contribute indirectly to the ability of chkYB-2 protein to complex with DNA. This possi-bility is further strengthened by our finding that the strong amino acid sequence conservation between eukaryotic Y-box proteins was not confined to the 70-aa CSD alone. A compar-ison of the sequence of the carboxyl-tail domain of chkYB-2 with those of other vertebrate Y-box proteins revealed an additional 32-aa conserved region contiguous with the carboxyl boundary of the classic CSD. Except for a single amino acid change, the aa 156–187 region showed 100% identity to human dbpA and greater than 70% identity to the Xenopus, chicken, mouse, and human YB-1 proteins.
The basic islands in the carboxyl-tail domain contain argi-nine clusters, which are frequently found in several RNA bind-ing proteins and are thought to increase the potential for nucleic acid binding (5). We have recently demonstrated that chkYB-2 binds single-stranded RNA in a sequence-specific manner and that YB-2 mutants [including YB-2 (1-169)] that lack the carboxyl tail fail to bind RNA (unpublished results). These results are in agreement with the recent report by Bou-vet et al. (4) which shows that RNA binding by FRGY2 is facilitated by both the amino- and carboxyl-terminal regions flanking the CSD, indicating contributions from regions be-yond the CSD for optimal nucleic acid interactions.
The chkYB-2 (158-298) mutant did not bind DNA, showing that the carboxyl-tail domain had no independent ability to bind DNA. This is similar to the results obtained by Murray (30), who found that recombinant FRGY2 proteins expressing only the carboxyl-tail domain were incapable of binding DNA. In contrast to FRGY2, wherein the presence of the carboxyl-tail domain facilitated the formation of multiple protein-DNA
complexes (43), we found that chkYB-2 mutants lacking the carboxyl-tail domain appeared to multimerize upon DNA binding.
The ability of Y-box factors to bind both single- and double-stranded DNA distinguishes them from other transcription factors. Impressive evidence implicating Y-box proteins in transcriptional regulation has accumulated in recent years. Re-ports of Y-box factors transactivating from viral promoters include the role of EF1A (12, 17) and chkYB-2 (40) in RSV LTR transcription and that of YB-1 in stimulating transcrip-tion from the human T-cell lymphotropic virus type 1, human immunodeficiency virus, and JCV virus promoters (22, 24). Y-box factors are also involved in the regulation of cellular genes. For example, YB-1 represses transcription of major histocompatibility complex class II genes (44) and acts as an activator of the MDR1 gene (1), and FRGY2 promotes tran-scription from the Xenopus hsp70 promoter (43).
Our earlier experiments have shown that chkYB-2 binds the single-stranded motif 59-GTACCACC-39 on the noncoding strand of the RSV enhancer and promotes RSV LTR-driven transcription. In this study, we present the results of systematic mutational analysis of this motif. Gel shift assays using a series of oligonucleotides carrying point mutations spanning the en-tire 59-GTACCACC-39 motif revealed that almost every nu-cleotide in the octamer was absolutely essential for high-affin-ity binding. A closer examination of the RSV LTR sequence showed that this octanucleotide motif (59-[G/C]TACCACC-39) was in fact present as a tandem repeat in the2112 to 2130 region. DNA binding assays carried out with a single-stranded oligonucleotide spanning this region showed a severalfold aug-mentation of DNA binding compared to oligonucleotides bearing a single motif. Also, the nature of complexes formed on gel shift assays suggested that the repeats behave more like two half-sites rather than two independent binding motifs. The importance of this region for chkYB-2 binding was further confirmed by introducing point mutations in the two half-sites in RSV LTR reporter constructs. Our results clearly demon-strate that mutations that decreased binding affinity in vitro also led to a decrease in the transcriptional activity of the corresponding mutant RSV LTR constructs.
chkYB-2 binds single- but not double-stranded DNA. The B.
subtilis Y-box protein CspB has been shown to be capable of
binding single-stranded but not double-stranded DNA (38). Also, NSEP-1 binds pyrimidine-rich single-stranded DNA (26), and YB-1 binds single-strand motifs with greater affinity than double-strand ones (29, 33). Apart from the Y-box pro-teins, the characterization of several other eukaryotic single-stranded DNA binding proteins has been reported (37, 41, 45). Other members of this growing family include the single-strand binding protein that complexes with the noncoding strand of the TSH receptor gene promoter and stimulates transcription (32), a pyrimidine single-strand-specific protein (ssPyrBF) that interacts with the androgen receptor gene promoter (6), and FUSE-binding protein, a single-stranded DNA binding protein whose role in transcription regulation of the c-myc gene has been well documented (10).
Experiments carried out to determine the effect of magne-sium on DNA binding revealed the remarkable increase in affinity upon the addition of 3 to 5 mM Mg21. Enhancement of DNA binding upon the addition of MgCl2has been reported
for two closely related Y-box proteins, human dbpA and dbpB (18). The effect of magnesium on FRGY2 binding to RNA has also been described (27). While the authors found Mg21to interfere with binding by the CSD over a range of 1 to 5 mM, it appeared to favor binding by the tail domains. Unr is a recently described DNA and RNA binding protein that is
on November 9, 2019 by guest
http://jvi.asm.org/
acterized by the presence of a fivefold repeat of the CSD but with no tail domains (20). Interestingly, the interaction of Unr with either DNA or RNA was very sensitive to even low con-centrations of magnesium. Addition of Mg21 markedly de-creased the affinity of binding, which was in contrast to the results we observed with chkYB-2. In fact, even chkYB-2 (1-169), a mutant with a large truncation in the tail domain, demonstrated increased DNA binding in the presence of Mg21.
Magnesium is the most abundant intracellular divalent cat-ion and is known to be required for the activity of several DNA repair enzymes. A recent report (13) that describes the effect of Mg21 on protein binding and structural transitions in a retroviral promoter, however, suggests that Mg21may also be physiologically relevant for the optimum activity of single-stranded DNA binding transactivators. The authors reported that in the presence of Mg21, the binding of a nuclear factor to a polypurine/polypyrimidine DNA sequence element (NRE1) in the mouse mammary tumor virus LTR led to the appearance of single-stranded regions upstream. Also, factor binding to single-stranded DNA was facilitated by the presence of Mg21. As the authors pointed out, in order for single-strand binding proteins to successfully bind and regulate transcription, there needs to be a mechanism that will expose single-stranded re-gions in the promoters of genes. Binding of a protein factor(s) that induces Mg21-dependent structural transitions that allows a second, Mg21-dependent single-stranded DNA binding fac-tor(s) to make contact with the exposed single strand is a possible solution to this problem.
chkYB-2 binds selectively to the pyrimidine-rich strand of the 2112 to 2130 region of the RSV LTR. The only other region (2142 to2161) in the LTR exhibiting a greater degree of purine/pyrimidine strand asymmetry is located about 10 bp upstream. Regions of strong purine/pyrimidine strand asym-metry that can assume an H-DNA conformation have been identified in the promoter elements of the c-myc andg-globin genes (19, 25). The pyrimidine-rich single strand is apparently accessible for interactions with proteins in these H-DNA re-gions. YB-1 has been shown to induce or stabilize single-stranded regions in a major histocompatibility complex class II gene promoter (29), and FUSE-binding protein has been re-ported to induce targeted melting of c-myc promoter prior to binding its single-strand recognition element on the noncoding strand (3). The regions of strand asymmetry in the RSV LTR may be too short to form H-DNA. However, localized melting of double-stranded DNA could expose the CT-rich strand for chkYB-2 binding.
In summary, we have characterized the functional domains in chkYB-2 responsible for its DNA binding activity, examined the effects, both in vitro and in vivo, of mutations in its binding motif, and discussed the possible relevance of Mg21 -depen-dent binding for transcriptional activation by a sequence-spe-cific, single-stranded DNA binding protein.
ACKNOWLEDGMENTS
We thank K. Krishnaveni for technical assistance and the University of Missouri DNA Core Facility for automated sequencing. We also thank D. Pintel for comments and Karen Ehlert for help in preparation of the manuscript.
This work was supported by a NIH grant 1 RO1 CA54192 to R.V.G.
REFERENCES
1. Bargou, R. C., K. Jurchott, C. Wagener, S. Bergmann, S. Metzner, K.
Bommert, M. Y. Mapara, K. J. Winzer, M. Dietel, B. Dorken, and H. D. Royer.1997. Nuclear localization and increased levels of transcription factor YB-1 in primary human breast cancers are associated with intrinsic MDR1 gene expression. Nat. Med. 3:447–450.
2. Bayarsaihan, D., B. Enkhmandakh, and L. N. Lukens. 1996. Y-box proteins interact with the S1 nuclease-sensitive site in the chickena2(I) collagen gene promoter. Biochem. J. 319:203–207.
3. Bazar, L., D. Meighen, V. Harris, R. Duncan, D. Levens, and M. Avigan. 1995. Targeted melting and binding of a DNA regulatory element by a transactivator of c-myc. J. Biol. Chem. 270:8241–8248.
4. Bouvet, P., K. Matsumoto, and A. P. Wolffe. 1995. Sequence-specific RNA recognition by the Xenopus Y-box proteins. J. Biol. Chem. 270:28297–28303. 5. Burd, C. G., and G. Dreyfuss. 1994. Conserved structures and diversity of
functions of RNA binding proteins. Science 265:615–621.
6. Chen, S., P. C. Supakar, R. L. Vellanoweth, C. S. Song, B. Chatterjee, and
A. K. Roy.1997. Functional role of a conformationally flexible homopurine/ homopyrimidine domain of the androgen receptor gene promoter interact-ing with Sp1 and a pyrimidine sinteract-ingle-strand DNA bindinteract-ing protein. Mol. Endocrinol. 11:3–15.
7. Cleavinger, P. J., B. A. Shin, J. C. Kandala, A. Nambiar, S. K. Swamynathan,
and R. V. Guntaka.1996. Cloning of Rous Sarcoma virus enhancer factor genes II. RSV-EF-II, abundantly expressed in fibroblasts and muscle tissue, binds to an octamer sequence, 59-GTACCACC-39in the noncoding strand of RSV enhancer. Virology 222:133–143.
8. Cohen, I., and W. F. Reynolds. 1991. The Xenopus YB3 protein binds the B-box element of the class III promoter. Nucleic Acids Res. 19:4753–4759. 9. Didier, D. K., J. Schiffenbauer, S. L. Wolffe, M. Zacheis, and B. D. Schwartz. 1992. Characterization of the cDNA encoding a protein binding to the major histocompatibility complex class II Y-box. Proc. Natl. Acad. Sci. USA 85: 7322–7326.
10. Duncan, R., L. Bazar, G. Michelotti, T. Tomonaga, H. Krutzsch, M. Avigan,
and D. Levens.1994. A sequence-specific, single-strand binding protein activates the far upstream element of c-myc and defines a new DNA binding motif. Genes Dev. 8:465–480.
11. Eustice, D. C., P. A. Feldman, A. M. Colberg-Poley, R. M. Buckery, and R. H.
Neubauer.1991. A sensitive method for the detection of beta-galactosidase in transfected mammalian cells. BioTechniques 11:739–742.
12. Faber, M., and L. Sealy. 1990. Rous sarcoma virus enhancer factor I is a ubiquitous CCAAT transcription factor highly related to CBF and NF-Y. J. Biol. Chem. 265:22243–22254.
13. Giffin, W., and R. J. G. Hache´. 1995. Nuclear factor binding to a DNA sequence element that represses MMTV transcription induces a structural transition and leads to the contact of single-stranded binding proteins with DNA. DNA Cell Biol. 14:1025–1035.
14. Goldstein, J., N. S. Pollitt, and M. Inouye. 1990. Major cold shock protein of
Escherichia coli. Proc. Natl. Acad. Sci. USA 87:283–287.
15. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant ge-nomes which express chloramphenicol acetyl transferance in mammalian cells. Mol. Cell. Biol. 2:1044–1051.
16. Grant, C. E., and R. G. Deeley. 1993. Cloning and characterization of chicken YB-1: regulation of expression in liver. Mol. Cell. Biol. 13:4186–4196. 17. Gruel, B. T., L. Sealy, and J. E. Majors. 1990. Transcriptional activity of the
Rous sarcoma virus long terminal repeat correlates with binding of a factor to an upstream CCAAT box in vitro. Virology 177:33–43.
18. Hasegawa, S. L., P. W. Doetsch, K. K. Hamilton, A. M. Martin, S. A.
Okenquist, J. Lenz, and J. M. Boss.1991. DNA binding properties of YB-1 and dbpA: binding to double-stranded, single-stranded, and abasic site con-taining DNAs. Nucleic Acids Res. 19:4915–4920.
19. Horwitz, E. M., K. A. Maloney, and T. J. Ley. 1994. A human protein containing a cold shock domain binds specifically to H-DNA upstream from the human gamma-globin genes. J. Biol. Chem. 269:14130–14139. 20. Jacquemin-Sablon, H., G. Triqueneaux, S. Deschamps, M. LeMaire, J.
Doniger, and F. Dautry.1994. Nucleic acid binding and intracellular local-ization of unr, a protein with five cold shock domains. Nucleic Acids Res.
22:2643–2650.
21. Kandala, J. C., and R. V. Guntaka. 1994. Cloning of Rous sarcoma virus enhancer factor genes. 1. Evidence that RSV-EF-I is related to Y-box (in-verted CCAAT) binding proteins and binds to multiple motifs in the RSV enhancer. Virology 198:514–523.
22. Kashanchi, F., J. F. Duvall, J. Dittmer, A. Mireskandari, R. L. Reid, S. D.
Gitlin, and J. N. Brady.1994. Involvement of transcription factor YB-1 in human T-cell lymphotropic virus type I basal gene expression. J. Virol.
68:561–565.
23. Kenny, S., and R. V. Guntaka. 1990. Localization by mutational analysis of transcription factor binding sequences in the U3 region of Rous sarcoma virus LTR. Virology 176:483–493.
24. Kerr, D., C. F. Chang, N. Chen, G. Gallia, G. Raj, B. Schwartz, and K.
Khalili.1994. Transcription of a human neurotropic virus promoter in glial cells: effect of YB-1 on expression of the JC virus late gene. J. Virol.
68:7637–7643.
25. Kinniburgh, A. J. 1989. A cis-acting transcription element of the c-myc gene can assume an H-DNA conformation. Nucleic Acids Res. 17:7771–7778. 26. Kolluri, R., T. A. Torrey, and A. J. Kinniburgh. 1992. A CT promoter
element binding protein: definition of a double-strand and a novel single-strand DNA binding motif. Nucleic Acids Res. 20:111–116.
27. Ladomery, M., and J. Sommerville. 1994. Binding of Y-box proteins to RNA:
on November 9, 2019 by guest
http://jvi.asm.org/
involvement of different protein domains. Nucleic Acids Res. 22:5582–5589. 28. Landsman, D. 1992. RNP-1, an RNA binding motif is conserved in the
DNA-binding cold shock domain. Nucleic Acids Res. 20:2861–2864. 29. MacDonald, G. H., Y. Itoh-Lindstrom, and J.-P. Y. Ting. 1995. The
tran-scriptional regulatory protein, YB-1 promotes single stranded regions in the DRA promoter. J. Biol. Chem. 270:3527–3533.
30. Murray, M. T. 1994. Nucleic acid binding properties of the Xenopus oocyte Y-box protein mRNP 314. Biochemistry 33:13910–13917.
31. Nambiar, A., J. C. Kandala, J. Svoboda, and R. V. Guntaka. Cloning of a novel Y-box homology protein (chk-YB-1HP) cDNA lacking the cold-shock domain. Biochim. Biophys. Acta, in press.
32. Ohmori, M., M. Ohta, H. Shimura, Y. Shimurat, K. Suzuki, and L. D. Kohn. 1996. Cloning of the single-strand DNA binding protein important for max-imal expression and thyrotropin (TSH)-induced negative regulation of the TSH receptor. Mol. Endocrinol. 10:1407–1424.
33. Ohmori, M., H. Shimura, Y. Shimura, and L. D. Kohn. 1996. A Y-box protein is a suppressor factor that decreases thyrotropin receptor gene ex-pression. Mol. Endocrinol. 10:76–89.
34. Ozer, J., M. Faber, R. Chalkley, and L. Sealy. 1990. Isolation and charac-terization of a cDNA clone for the CCAAT transcription factor EF1A re-veals a novel structural motif. J. Biol. Chem. 265:22143–22152.
35. Ranjan, M., S. Tafuri, and A. P. Wolffe. 1993. Masking mRNA from trans-lation in somatic cells. Genes Dev. 7:1725–1736.
36. Sakura, H., T. Maekawa, F. Imamoto, K. Yusuda, and S. Ishii. 1988. Two human genes isolated by a novel method encoding DNA-binding proteins containing a common region of homology. Gene 73:499–507.
37. Santoro, I. M., T. M. Yi, and K. Walsh. 1991. Identification of single-stranded DNA binding proteins that interact with muscle gene elements. Mol. Cell. Biol. 11:1944–1953.
38. Schindelin, H., M. A. Marahiel, and U. Heinemann. 1993. Universal nucleic acid-binding domain revealed by crystal structure of the B. subtilis major
cold-shock protein. Nature 364:164–168.
39. Sommerville, J., and M. Ladomery. 1996. Masking of mRNA by Y-box proteins. FASEB J. 10:435–443.
40. Swamynathan, S. K., A. Nambiar, and R. V. Guntaka. 1997. Chicken YB-2, a Y-box protein, is a potent activator of Rous sarcoma virus long terminal repeat-driven transcription in avian fibroblasts. J. Virol. 71:2873–2880. 41. Tada, H., and K. Khalili. 1992. A novel sequence-specific DNA binding
protein, LCP-1, interacts with single-stranded DNA and differentially regu-lates early gene expression of the human neurotropic JC virus. J. Virol.
66:6885–6892.
42. Tafuri, S. R., and A. P. Wolffe. 1990. Xenopus Y-box transcription factors: molecular cloning, functional analysis, and developmental regulation. Proc. Natl. Acad. Sci. USA 87:9028–9032.
43. Tafuri, S. R., and A. P. Wolffe. 1992. DNA-binding, multimerization and transcription stimulation by the Xenopus Y-box in vitro. New Biol. 4:349– 359.
44. Ting, J.-P. Y., A. Painter, N. J. Zeleznik-Le, G. MacDonald, T. M. Moore, A.
Brown, and B. D. Schwartz.1994. YB-1 DNA binding protein represses interferon gamma activation of class II major histocompatibility complex genes. J. Exp. Med. 179:1605–1611.
45. Wang, Z. Y., X. H. Lin, M. Nobuyoshi, and T. F. Deuel. 1993. Identification of a single-stranded DNA binding protein that interacts with an S1 nuclease-sensitive region in the platelet-derived growth factor A-chain gene promoter. J. Biol. Chem. 268:10681–10685.
46. Wistow, G. 1990. Cold shock and DNA binding. Nature 344:823–824. 47. Wolffe, A. P. 1994. Structural and functional properties of the evolutionarily
ancient Y-box family of nucleic acid binding proteins. Bioessays 16:245–251. 48. Wolffe, A. P., S. Tafuri, M. Ranjan, and M. Familari. 1992. The Y box factors: a family of nucleic acid binding proteins conserved from Escherichia
coli to man. New Biol. 4:290–298.