0022-538X/99/$04.0010
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Genetic Analysis of a Unique Human Immunodeficiency Virus
Type 1 (HIV-1) with a Primer Binding Site Complementary to
tRNA
Met
Supports a Role for U5-PBS Stem-Loop RNA
Structures in Initiation of HIV-1 Reverse Transcription
SANG-MOO KANGANDCASEY D. MORROW*
Department of Microbiology, University of Alabama at Birmingham, Birmingham, Alabama 35294 Received 16 July 1998/Accepted 24 November 1998
Human immunodeficiency virus type 1 (HIV-1) exclusively uses tRNA3Lysto initiate reverse transcription. A
novel HIV-1 mutant which stably utilizes tRNAMetrather than tRNA
3
Lysas a primer was previously identified
[HXB2(Met-AC] (S.-M. Kang, Z. Zhang, and C. D. Morrow, J. Virol. 71:207–217, 1997). Comparison of RNA secondary structures of the unique sequence (U5)-primer binding site (PBS) viral RNA genome alone or
complexed with tRNAMet of HXB2(Met-AC) revealed structural motifs in common with the U5-PBS of the
wild-type virus. In the current study, mutations were constructed to alter the U5-PBS structure and disrupt the
U5-PBS-tRNAMet interaction of the virus derived from HXB2(Met-AC). All of the mutant viruses were
infectious following transfection and coculture with SupT1 cells. Analysis of the initiation of reverse tran-scription revealed that some of the mutants were impaired compared to HXB2(Met-AC). The genetic stability of the PBS from each virus was determined following in vitro culture. Two mutant proviral constructs, one predicted to completely disrupt the stem-loop structure in U5 and the other predicted to destabilize contact
regions of U5 with tRNAMet, reverted back to contain a PBS complementary to tRNA
3
Lys. All other mutants
maintained a PBS complementary to tRNAMetafter in vitro culture, although all contained multiple nucleotide
substitutions within the U5-PBS from the starting proviral clones. Most interestingly, a viral mutant contain-ing a 32-nucleotide deletion between nucleotides 142 and 173, encompasscontain-ing regions in U5 which interact with
tRNAMet, maintained a PBS complementary to tRNAMetfollowing in vitro culture. All of the proviral clones
recovered from this mutant, however, contained an additional 19-nucleotide insertion in U5. RNA modeling of the U5-PBS from this mutant demonstrated that the additional mutations present in U5 following culture restored RNA structures similar to those modeled from HXB2(Met-AC). These results provide strong genetic evidence that multiple sequence and structural elements in U5 in addition to the PBS are involved in the interaction with the tRNA used for initiation of reverse transcription.
A hallmark of the retrovirus life cycle is the process by which the single-stranded viral RNA genome is converted into a double-stranded DNA (1, 25). This process, called reverse transcription, uses a virus-encoded enzyme, reverse transcrip-tase (RT), and a cellular tRNA primer. The tRNA is bound to a site on the viral RNA genome, designated the primer binding site (PBS), which is complementary to the 39-terminal 18 nu-cleotides of the tRNA (4, 18). The PBS is located downstream of the 59direct repeat (R) and unique sequence (U5) in the viral RNA genome.
The tRNA primer used for initiation of reverse transcription is different for individual retrovirus groups. For example, tRNA3Lysis used by lentiviruses such as HIV-1 (human
immu-nodeficiency virus type 1) or simian immuimmu-nodeficiency virus, while tRNA1,2Lys is used by Mason-Pfizer monkey virus, visna
virus, and spumavirus; murine leukemia virus uses tRNAPro,
while avian sarcoma virus and avian leukosis virus use tRNATrp(15, 16). Why retroviruses use different but specific
tRNA primers to initiate reverse transcription is unknown. The exclusive use of specific tRNAs to initiate reverse transcription cannot be explained by incorporation into the virion particle, since a subset of cellular tRNAs, in addition to the specific
tRNA used for initiation, are encapsidated. For example, HIV-1 virions contain a subset of cellular tRNAs comprised of 30% tRNA3Lys, 60% tRNA1,2Lys, and 10% minor tRNAs (7, 29).
Recent studies have demonstrated that the composition of the tRNA species within the virion does not necessarily dictate which tRNA will be used to initiate reverse transcription. HIV-1 with genetically engineered PBS complementary to al-ternative tRNAs could use a wide variety of tRNAs for initi-ation of reverse transcription. The PBSs of these viruses were not stable and reverted to the wild-type PBS, complementary to tRNA3Lys, after a few passages of in vitro culture (3, 11, 28).
Based on these results, it was suggested that factors other than the PBS sequence specify the preferential usage of the wild-type primer for the initiation of retroviral reverse transcription. Recent studies have provided evidence of additional inter-actions between the 59region of the retroviral genome and the primer tRNA. Chemical and enzymatic footprinting of the HIV-1 genomic RNA and tRNA3Lyshas found additional sites
in U5 involved in intermolecular contacts. In one of these interactions, the anticodon loop of tRNA3Lys binds with an
A-rich loop located 12 to 17 nucleotides upstream of the PBS (5, 6, 14). Genetic evidence for a role of this A-rich loop region in interacting with primer tRNA was provided by demonstrat-ing that if both the PBS and A-rich loop in U5 are mutated so as to be complementary to the 39-terminal nucleotides and anticodon loop of tRNAHis(27) or tRNAMet(9), the resulting
viruses would stably maintain a corresponding PBS
comple-* Corresponding author. Mailing address: Department of Microbi-ology, University of Alabama at Birmingham, Birmingham, AL 35294. Phone: (205) 934-5705. Fax: (205) 934-1580. E-mail: casey_morrow @micro.micobio.uab.edu.
1818
on November 9, 2019 by guest
http://jvi.asm.org/
mentary to tRNAHisor tRNAMet, respectively, after extended
in vitro culture. In a follow-up study, we found that the tRNA composition in the virions from viruses which stably utilize tRNAHisto initiate reverse transcription was not changed
com-pared to that of the wild-type virus (29). The complexity of the U5-PBS interaction with tRNA is highlighted by the fact that not all combinations of PBS and anticodon complementary sequence in U5 (A-loop region) produce viruses which can stably maintain a PBS complementary to alternative tRNAs (8).
The development of infectious HIV-1 which stably maintain a PBS complementary to an alternative tRNA provides a unique opportunity to genetically test the importance of the U5-PBS in the selection of the tRNA used for initiation of reverse transcription. We have previously reported on the characterization of an HIV-1 which uses tRNAMetto initiate
reverse transcription (9). In contrast to our other HIV-1, which utilize an alternative tRNA to initiate reverse transcription, the virus which uses tRNAMetoccurred spontaneously in in vitro
cultures. Follow-up studies established that this virus was sta-ble if a region within U5 was engineered to contain a sequence complementary to the anticodon loop of tRNAMet. In the
current study, we found that modeling of the U5-PBS region of this virus without and with bound tRNAMet revealed RNA
structures with striking similarities to the wild-type virus (see Fig. 1). To determine a biological significance for these RNA structures, we constructed proviral clones which contain mu-tations designed specifically to disrupt the stem-loop structures in U5 of this virus. The capacity of these mutant viruses to use a tRNAMet as a primer in an in vitro endogenous reverse
transcription assay and maintain the PBS complementary to tRNAMetfollowing in vitro culture was also determined. Our
studies demonstrate that mutations upstream of the A-loop region in U5 affect reverse transcription and utilization of the tRNAMetprimer. Modeling of the RNA genomes containing
the nucleotide substitutions which arise in viruses following in vitro culture supports the conclusion that U5-PBS RNA struc-ture plays an important role in the selection of the tRNA and subsequent initiation of reverse transcription.
MATERIALS AND METHODS
Construction of vectors with mutant proviral genomes.The HXB2 molecular clone of HIV-1 was used to construct the mutant HIV-1 proviral genomes (19). We previously reported the construction of pHXB2(Met-AC) with a substitution of the nucleotide sequence TGTGAGACTG (nucleotides 167 to 176) in U5 and a PBS complementary to tRNAMet, which resulted in the virus stably maintaining a PBS complementary to tRNAMet(9). From a virus culture derived from pHXB2(Met-AC), a U5 PBS region was PCR amplified from provided DNA with three additional point mutations in U5 (145C to T,171G to A, and201G to A); this U5-PBS was isolated and used to construct HXB2(Met-AC). ABglII and BssHII DNA fragment (nucleotides 20 to 255) from the clone was first purified from a 1.2% agarose gel and cloned into shuttle vector pUC119(PBS), which contains an HpaI-to-PstI DNA fragment including the 59long terminal repeat, PBS, and leader region of thegaggene from HXB2. The resulting clone was named pUC119(Met-AC). All other mutant constructs were created by intro-ducing mutations in the U5 region of the pUC119(Met-AC) shuttle vector by using two consecutive PCR mutageneses (23). The first PCR was done to gen-erate megaprimers containing mutant sequences by using a primer in the U3 region of the 59long terminal repeat (59TTGACAGCCGCCTAGC39 [nucleo-tides 8895 to 8910]) and a mutagenic oligonucleotide. The mutations which were constructed are depicted in Fig. 1 and 2. The oligonucleotides used for construc-tion of the mutants are as follows: Met-AC(157-161), 59-CTCACAACACTCT GACAAAGGGTCTGAAG-39; Met-AC(162-164), 59-AGTCTCACAACTGAG ACTAAAAGGGTC-39; Met-AC(157-166), 59-GCTACAGTCTCACAGACAA AAGCGAAAGGGTCTGAAGGA-39; Met-AC(153-161), 59-GTCTCACAAC ACTTGTGAGACTGGTCTGAAGGATCTC-39; Met-AC(161-163), 59-AGTC TCACAACAGACACTAAAAGGGTC-39; Met-AC(143-146), 59-ACTAAAAG GGTCTGTTCCATCTCTAGTTACC-39; Met-AC(143-146,161-163), 59-AGTC TCACAACAGACACTAAAAGGGTCTGTTCCATCTCTAGTTACCA-39; and Met-AC(D142-173), 59-CCACTGCTAGAGTCTCTAGTTACCAGAGTCAC-39.
Megaprimers containing the mutant sequences were used in the second PCR
together with a primer with aBssHII site downstream of the PBS (59-GCGCG CTTCAGCAAGCCG-39[nucleotides 262 to 245]) to generate restriction en-zyme sites for cloning into pUC119(PBS). The resulting mutagenic DNA frag-ments were digested withBglII andBssHII (nucleotides 20 to 262) and cloned into pUC119(PBS), creating the corresponding mutant shuttle vectors. An 868-bp fragment ofHpaI andBssHII from each of the pUC119 plasmid con-structs containing the mutant sequences was subcloned between theHpaI and BssHII sites of HXB2. All pUC119 constructs and the resulting HXB2 mutant proviral plasmids were verified by DNA sequencing to ensure the identity of the mutated sequence (22).
Tissue culture and DNA transfections.293T and COS-1 cells were maintained in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum and 1% penicillin-streptomycin at 37°C and 5% CO2. SupT1 cells were grown in RPMI 1640 medium containing 15% fetal calf serum and 1% penicillin-strepto-mycin at 37°C and 5% CO2. 293T cells were grown overnight to about 70% confluence in a six-well plate and transfected with 10mg of HXB2 proviral DNA constructs using the Ca-PO4 method (Stratagene protocol). After overnight incubation at 37°C, the supernatant of transfected 293T cells was removed and fresh medium was added. The supernatant was collected at 3 days posttransfec-tion and filtered through a 0.45-mm-pore-size syringe filter (Nalgene). The levels of p24 capsid antigen were determined from three independent transfections by using an enzyme-linked immunosorbent assay (ELISA; Coulter Laboratories). COS-1 cells (at 60% confluence) were transfected with 5mg of wild-type or mutant proviral plasmid DNA by using DEAE-dextran as previously described (20).
Endogenous reverse transcription followed by PCR.The endogenous reverse transcription was performed by using virus particles supplied with nucleotide substrates as described elsewhere (17). Supernatants from 293T cells transfected with proviral DNAs were treated with RNase-free DNase I (Boehringer Mann-heim) at a final concentration of 20 U/ml for 1 h at 37°C in the presence of 10 mM MgCl2. Virus particles from transfected supernatants were collected by centrifugation at 27,000 rpm for 2 h in an SW28 rotor at 4°C. The pellet was resuspended in 200ml of ice-cold TEN buffer (100 mM NaCl, 10 mM Tris-HCl, pH 8.0), and aliquots of virus were kept at 270°C. For endogenous reverse transcription, aliquots of a virus suspension (equivalent to 6 ng of p24) were incubated in 60ml of the reaction mixture (0.01% Triton X-100, 50 mM NaCl, 50 mM Tris-HCl [pH 8.0], 10 mM dithiothreitol, 5 mM MgCl2, 200mM each dATP, dGTP, dCTP, and dTTP) for 0 min, 20 min, 1 h, and 2 h at 37°C. Reactions were terminated by adding 30ml of stop mix (250-mg/ml proteinase K, 5 mM EDTA, pH 8.0) and incubation at 60°C for 1 h. Reactions without deoxynucleoside triphosphate substrates were performed as a control. Reaction mixtures were boiled for 10 min to inactivate proteinase K before PCR analysis. A tRNALys-1 primer, 59-TAGCTCAGTCGGTAGAGCA-39corresponding to nucleotides 8 to 27 of tRNA1,2Lyswas used in a PCR to amplify minus-strand, strong-stop DNA [(2)ss DNA] linked to tRNA3Lys; this DNA primer did not amplify tRNAMet -extended (2)ss DNA or plasmid DNA. To amplify (2)ss DNA linked to tRNAMetpresent in the reaction mixture, a tRNAMet-1 primer was used in a PCR (59-GGAATTCGTTAGCGCAGTAGCGCGTCAGTCTCA-39, corre-sponding to nucleotides 4 to 35 of tRNAMet); this primer did not amplify tRNA3Lys-extended (2)ss DNA or plasmid DNA. Primer 1, corresponding to nucleotides 17 to 38 in the 59R of HXB2 proviral DNA was used as a 59PCR primer. A 1-ml volume of the reaction mixture (equivalent to 100 pg of p24 antigen) was subject to 30 cycles of PCR, each consisting of a denaturing step at 94°C for 1 min, annealing at 60°C for 1 min, and extension at 72°C for 40 s. Amplified products were resolved on a 1.2% agarose gel. To compare the relative amounts of (2)ss DNA linked to tRNA3Lysor tRNAMet, Southern hybridization was performed by using specific probes for tRNA species. The PCR-amplified DNA in the agarose gel was transferred to a 0.2-mm-pore-size (Protran) nitro-cellulose membrane in 203SSPE (13SSPE is 0.18 M NaCl, 10 mM NaH2PO4, and 1 mM EDTA [pH 7.7]) buffer (21). DNA was fixed to the membrane by baking at 80°C for 2 h. The probes were the following32P-end-labeled tRNA species-specific oligomers: tRNALys-2, 59-CAGACTTTTAATCTGAGGGTCC AGG-39, corresponding to nucleotides 28 to 53 of tRNA3Lys; tRNAMet-2, 59-CT CATAATCTGAAGGTCGTGAG-39, corresponding to nucleotides 32 to 53 of tRNAMet. Prehybridization was carried out at room temperature for 5 to 8 h in a buffer containing 25% (vol/vol) formamide, 63SSPE, 0.1% sodium dodecyl sulfate, 53Denhardt’s solution, and 100-mg/ml yeast tRNA. Hybridization was initiated by adding the labeled probe to the prehybridization mixture and con-tinued overnight at room temperature. The membrane was washed in washing buffer (23SSPE with 0.1% sodium dodecyl sulfate) three times for 10 min each at room temperature. Radioactivity was determined by PhosphorImager analysis of the blots.
Analysis of virus replication.At 1 day posttransfection, COS-1 cells were cocultured with SupT1 cells (53105), which support high-level replication of HIV-1. After 48 h of coculture, the SupT1 cells were harvested by low-speed centrifugation and further cultured with fresh medium and additional SupT1 cells. The infected SupT1 cells were monitored visually for the formation of multicell syncytia and maintained by addition of fresh SupT1 cells and medium at various time intervals. For cell-free infections at 120 days postcoculture, SupT1 cells (106/ml) were infected with equal amounts of virus as measured by p24 antigen (100 ng). After allowing the virus to adsorb for 24 h, SupT1 cells were further cultured in RPMI medium. At the designated time intervals, the
on November 9, 2019 by guest
http://jvi.asm.org/
culture supernatants were collected and analyzed for p24 antigen by ELISA (Coulter Laboratories).
PCR amplification and DNA sequencing of PBS-containing proviral DNA.On designated days postcoculture, DNAs were isolated from infected SupT1 cells by using the Wizard genomic DNA purification kit and following the manufacturer’s (Promega) instructions. Approximately 1mg of cellular DNA was used to amplify the U5 and PBS regions of integrated proviral DNA sequences by using the following HIV-1 proviral-DNA-specific primers: primer 1, 59-GCTCTAGACC AGATCTGAGCCTGGGAGCTC-39(nucleotides 17 to 38); primer 2, 59-CGG AATTCTCTCCTTCTAGCCTCCGCTAGTC-39 (nucleotides 309 to 330). PCR-amplified DNA was directly ligated into the pGEM-T-easy vector (Pro-mega). Following transformation intoEscherichia coliand screening, the U5-PBS-containing plasmid DNAs prepared from individual recombinant clones were sequenced by using the primer 59-GGCTAACTAGGGAACCCACTGC-39 (nucleotides 42 to 63).
RESULTS
Construction of mutant HIV-1 proviral DNAs.In a previous
study, we demonstrated that the HIV-1 mutants with both a PBS and nucleotides in U5 complementary to the anticodon loop of tRNAMet(ACN, the A-loop region in the wild-type
genome) stably maintained a PBS complementary to tRNAMet
following extended in vitro culture; we designated this virus HXB2(Met-AC) (9). HIV-1 mutants with both a PBS and ACN complementary to tRNAIleor tRNAProwere not stable
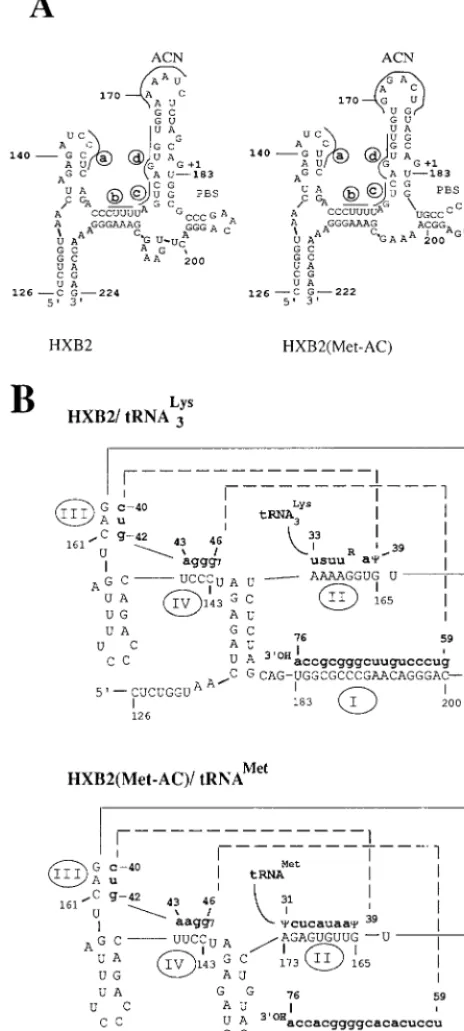
and, following in vitro culture, reverted to contain a wild-type PBS (8). The results of these studies, combined with those of others, have led to a greater appreciation of the fact that a complex RNA structure exists for both the uncomplexed and tRNA-bound U5-PBS which is important for reverse transcrip-tion. A U5-PBS structure for the wild-type genome has been elucidated from a combination of chemical and enzymatic analyses (2) (Fig. 1A). Several regions have been identified which could have importance in the initiation of reverse tran-scription. RNA modeling of the U5-PBS revealed a long stem-loop structure, from nucleotide 159 to nucleotide 186, which encompasses the initiation site for reverse transcription (nu-cleotide 182, followed by11), as well as the ACN (32). Several other stem-loop structures were also noted (designated a and b in Fig. 1A). Similar RNA modeling of the genome which stably maintains a PBS complementary to tRNAMetrevealed a
structure strikingly similar to that of the wild type (32). The only major difference was noted with respect to the nucleotides displayed on the ACN; in HXB2(Met-AC), the sequence GAG ACU was present on the loop, which is complementary to the nucleotides in the anticodon region of tRNAMet.
Previous studies by Isel et al. have used both chemical and enzymatic analyses to determine an HIV-1 tRNA3Lys binary
complex (Fig. 1B) (5). Several regions of interaction between the viral RNA genome and the tRNA were identified. The most obvious is the complementarity between the 39-terminal nucleotides of the tRNA and the PBS (designated I in Fig. 1B). Three additional, unexpected interactions between the HIV-1 U5 and tRNA were also identified (designated II, III, and IV in Fig. 1B). Again, a similar binary complex could also be drawn with the RNA genome of HXB2(Met-AC) and tRNAMet(Fig. 1B).
The conservation of RNA structures between the wild type and HXB2(Met-AC) suggested their importance for reverse transcription. To test this possibility, we targeted mutations to disrupt the stem-loop RNA structure of HXB2(Met-AC) (Fig. 1A). The mutations were selected for disruption of the pre-dicted U5-PBS RNA structure but would not be prepre-dicted to disrupt important RNA structures in the U5-PBS-tRNA com-plex (I, II, and IV in Fig. 1B). Thus, we focused our changes on nucleotides 153 to 166 in U5 to disrupt the stem-loop struc-tures in the regions designated b, c, and d (Fig. 1A and 2) and constructed HXB2(Met-AC(157-161)), HXB2(Met-AC(162-FIG. 1. RNA modeling of wild-type and mutant viral U5-PBS and U5-PBS/
tRNA. (A) The viral RNA sequence of nucleotides 1 to 230 encompassing the R-U5-PBS from the wild-type (HXB2) and mutant HXB2(Met-AC) viruses was used to predict the RNA secondary structures by using MFold (33). Only struc-tures from nucleotides 126 to 224 are shown; the original clone of HXB2(Met-AC) has a two-nucleotide deletion downstream of the PBS. The ACN sequences are marked. The regions targeted for mutations are designated a to d and underlined. The designation11 indicates the first nucleotide of the RNA tem-plate for negative-strand DNA synthesis. Nucleotides 183 to 200 are the PBS sequence. (B) RNA structure of U5-PBS/tRNA based on a model proposed by Isel et al. (5). Viral RNA sequences are shown in capital letters and tRNA sequences (only the sequences interacting with viral RNA are displayed) are in boldface lowercase letters. Region I depicts the PBS-tRNA interaction. Region II is the A-loop region in U5 complementary to the anticodon loop of tRNA. Region III is complementary to the 39anticodon stem of tRNA. Region IV is complementary to the variable loop of tRNA.
on November 9, 2019 by guest
http://jvi.asm.org/
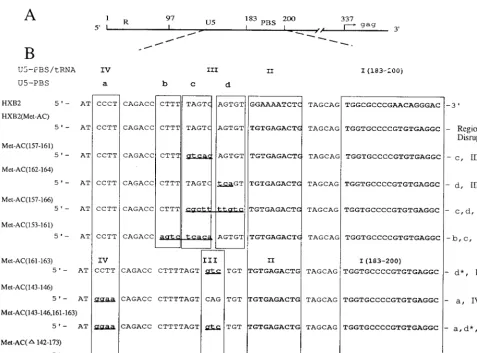
[image:3.612.59.291.80.596.2]164)), HXB2(Met-AC(157-166)), and HXB2(Met-AC(153-161)). The nucleotides were chosen such that the predicted RNA structures with the mutations would be sufficiently dif-ferent from the original predicted RNA structure of HXB2(Met-AC) (in the secondary-structure analysis using the MFold algorithm). The mutations were named in accordance with the following convention: HXB2(Met-AC(Y-Y)), where Y identifies the nucleotides changed from the starting genome, that of HXB2(Met-AC).
We also introduced a second set of mutations into regions III and IV to disrupt the U5-PBS-tRNA interactions (Fig. 1B and 2), creating HXB2(Met AC(161-163)), HXB2(Met-AC(143-146)), and HXB2(Met-AC(143-146, 161-163)); some of these mutations would also be predicted to disrupt RNA secondary structures in regions a and d of Fig. 1A. Finally, we also constructed a proviral genome which contained a deletion of the U5 region from nucleotide 142 to nucleotide 173, HXB2(Met-AC(D142-173)). Although we anticipated that this virus would be viable since previous studies by Vicenzi et al. had demonstrated that deletion of a similar region in U5 did not result in a noninfectious wild-type virus, we predicted that
the PBS of this virus would rapidly revert to using tRNA3Lysas
the primer for reverse transcription (26). In addition, it has previously been reported that mutations in this region do not effect encapsidation of the genome RNA (26).
Analysis of the infectious potential of mutant proviruses.
The proviral clones were transfected into 293T cells, and p24 capsid antigen levels in the culture supernatants were mea-sured at 48 h posttransfection to determine the effect of mu-tations on viral protein production and virion release. The levels of p24 antigen released in the supernatant were similar in all of the clones tested (data not shown).
To determine the infectious potential of the mutant proviral genomes, COS-1 cells were transfected with the proviral ge-nomes and then cocultured with SupT1 cells, consistent with previous studies (9, 20, 28, 29). At various times postcoculture, the production of both the wild-type and mutant viruses was monitored by visual inspection for syncytia; virus production was also quantitated by levels of viral capsid (p24) antigen in the supernatants from the cultures (Fig. 3). Although the ki-netics of appearance for all mutant viruses was delayed com-pared with that of AC), the viruses from HXB2(Met-FIG. 2. Expanded view of the 59region of the HIV-1(HXB2) RNA genome and nucleotide sequences of the U5-PBS region in wild-type and mutant proviruses. (A) Schematic location of the R-U5-PBS region in HIV-1 genomic RNA. R is the direct repeat sequence at the 59and 39ends of the viral RNA genome. (B) Nucleotide sequences of wild-type (HXB2) and mutant proviruses constructed from HXB2(Met-AC). a, b, c, and d are regions involved in the stem-loop structure in U5 as described in Fig. 1A. I, II, III, and IV are regions of viral RNA positioned to interact with the tRNA primer as described in Fig. 1B. Regions targeted for mutation are aligned and boxed along with the ACN sequence (II) and the PBS (I). Asterisks indicate partial disruption of the marked region, and dots indicate deletion. Designations: AC(157-161), HXB2(AC(157-161)); AC(161-164), HXB2(AC(161-164)); AC(157-166), HXB2(AC(157-166)); Met-AC(153-161), HXB2(Met-AC(153-161)); Met-AC(161-163), HXB2(Met-AC(161-163)); Met-AC(143-146), HXB2(Met-AC(143-146)); Met-AC(143-146, 161-163), HXB2(Met-AC(143-146, 161-163)); Met-AC(D142-173), HXB2(Met-AC(D142-173)).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.62.539.71.424.2]AC(157-166)), HXB2(Met-AC(153-161)), HXB2(Met-AC(143-146)), and HXB2(Met-AC(143-146, 161-163)) showed the greatest delay.
To understand how the mutations in U5 affect viral replica-tion, we analyzed the initiation of reverse transcription of the mutant viruses obtained following transfection of proviral DNA containing the designated mutations. For this analysis, we utilized an in vitro endogenous RT PCR method to detect (2)ss DNA since virion particles contain all of the necessary viral components for reverse transcription (17). The products for endogenous reverse transcription were subjected to PCR amplification using a tRNA-specific primer, followed by South-ern hybridization with a32P-labeled tRNA-specific probe. In
preliminary experiments, we analyzed the wild-type (2)ss DNA extended from the tRNA3Lys [39R-U5(DNA)-tRNA3Lys
-59]. The amount of (2)ss DNA increased for approximately 120 min of in vitro incubation time before plateauing (data not shown). For the mutants then, we used a 120-min incubation time for the reverse transcription reaction and a tRNAMet
-specific PCR primer with a tRNAMet-specific probe
comple-mentary to tRNAMetto detect (2)ss DNA produced during
endogenous reverse transcription. The amounts of (2)ss DNA product [39R-U5(DNA)-tRNAMet-59] were compared for each
[image:5.612.58.293.69.441.2]of the mutants (Fig. 4A and B). There was a general correla-tion between the amounts of (2)ss DNA detected and the appearance of the viruses following transfection. That is, we detected lower amounts of the (2)ss DNA product from vi-ruses derived from AC(157-166)), AC(153-161)), AC(143-146)), and HXB2(Met-AC(143-146, 161-163)) than that from HXB2(Met-AC). The most severely replication-compromised virus, HXB2(Met-AC-(157-166)), had the lowest amount of (2)ss DNA product detected from the in vitro reactions (Fig. 4A). The correlation was not perfect, however, as evidenced by the analysis of HXB2(Met-AC(D142-173). In this case, we detected less re-verse transcription products from HXB2(Met-AC(D142-173)) than from other viruses ([e.g., HXB2(Met-AC(143-146)). However, the virus derived from HXB2(Met-AC(D143-172))
[image:5.612.318.541.352.579.2]FIG. 3. Appearance of infectious virus after transfection of mutant proviral genomes. Plasmids containing the wild-type or mutant proviral genomes were transfected into COS-1 cells and cocultured with SupT1 cells (53105) 24 h later. After 48 h, the SupT1 cells were isolated by centrifugation, washed once, and further cultured with additional SupT1 cells and medium (day 0). At various intervals postcoculture, culture supernatants were collected and the p24 antigen was quantitated by ELISA. (A) Mutant viruses designed to affect the secondary structure in the context of RNA alone: Met-AC(157-161), HXB2(Met-AC(157-161)); Met-AC(162-164), HXB2(Met-AC(162-164)); Met-AC(157-166), HXB2(AC(157-166)); AC(153-161), HXB2(AC(153-161)); Met-AC, HXB2(Met-AC). (B) Mutant viruses designed to affect the contacting re-gions in the complex of U5-PBS and tRNA: Met-AC(143-145, 161-163), 146, 161-163)); Met-AC(143-146), HXB2(Met-AC(143-146)); Met-AC(161-163), HXB2(Met-AC(161-163)); Met-AC(D142-173), HXB2(Met-AC-(D142-173)).
FIG. 4. Analysis of initiation of reverse transcription. Virus (equivalent to 6 ng of p24 antigen) produced from transfection of 293T cells were used in an endogenous reverse transcription reaction (37°C for 2 h). Endogenous reaction products (equivalent to 100 pg of p24 antigen) were subjected to 30 cycles of PCR to amplify the extended tRNAMet linked to (2)ss DNA by using a tRNAMet-specific primer. No PCR products were observed from viruses incu-bated without deoxynucleoside triphosphates (data not shown). Southern hybrid-ization was used to detect cDNA products of tRNAMetlinked to (2)ss DNA by using a 32P-labeled tRNAMet-specific probe. Radioactivity was quantified by using a PhosphorImager. (A) Mutant viruses designed to affect the secon-dary structure in the context of RNA alone. Met-AC(157-161), HXB2(Met-AC(157-161)); Met-AC(162-164), HXB2(Met-AC(162-164)); Met-AC(157-166), HXB2(AC(157-166)); AC(153-161), HXB2(AC-(153-161)); Met-AC, HXB2(Met-AC). (B) Mutant viruses designed to affect the contacting regions in the binary complex of HIV-1 RNA and tRNA. Met-AC(143-146,161-163), HXB2(Met-AC-(143-146, 161-163)); Met-AC(143-146), HXB2(Met-AC(143-146)); Met-AC(161-163), HXB2(Met-AC(161-163)); Met-AC(D142-173), HXB2(Met-AC(D142-173)).
on November 9, 2019 by guest
http://jvi.asm.org/
grew faster than the virus from HXB2(Met-AC(143-146)). Al-though the exact reason for this discrepancy is not clear, it is possible that the virus derived from HXB2(Met-AC(D 143-172)) had undergone rapid mutation as a result of limited tissue culture, which facilitated virus replication (see the next section).
DNA sequence analysis of the U5-PBS region from
inte-grated proviruses obtained after extended in vitro culture.To
ascertain the stability of the PBS in the mutant viruses, after 120 days in culture we harvested the culture supernatants and analyzed the replication of the viruses by infecting SupT1 cells with equal amounts of virus as measured by p24 antigen. Ex-cept for strain HXB2(Met-AC(143-146, 161-163)), all of the
mutant viruses replicated similarly to the wild-type (HXB2) and HXB2(Met-AC) (data not shown); the replication of HXB2(Met-AC(143-146,161-163)), however, was still delayed compared to that of the wild-type and mutant viruses.
One of the hallmarks of HIV-1 with a PBS complementary to an alternative tRNA is the propensity of the PBS to revert to complementarity to tRNA3Lysfollowing in vitro culture.
Pre-viously, we have found that even subtle mutations within U5 of viruses with a PBS complementary to tRNAHis or tRNA
1,2 Lys
resulted in reversion back to the wild-type PBS following in vitro culture (30). To determine if this was also the case for the mutants with a PBS complementary to tRNAMet, we analyzed
the U5-PBS from integrated proviruses. Since the RT copies
TABLE 1. DNA sequences of U5-PBS regions from integrated proviruses in infected cells at day 120
Strain and sequencea Frequencyb
HXB2(Met-AC) ACN (167–176)c PBS (183–200)d
59-ACTAGAGATCCtTCAGACCCTTTTAGTCAGTGTTGTG AGACTGTAGCAG TGGTGCCCCGTGTGAGGC-39... Input
59-******A****************************** ********A*** *****************A-39... 5/9
59-******A**********************A******* ********A*** ******************-39... 2/9
59-******A****************************** ************ *****************A-39... 2/9
1e
HXB2(Met-AC(162–164))
59-ACTAGAGATCCtTCAGACCCTTTTAGTCtcaG TTGTGAGACTGTAGCAG TGGTGCCCCGTGTGAGGC-39... Input
59-******A************************* ***************** ******************-39... 2/6
1TTATGAGACTGTAGTAG
59-******A************************* A*A************** ******************-39... 4/6
1f
HXB2(Met-AC(157–166))
59-ACTAGAGATCCtTCAGACCCTTTcgcttttgtcTGTGAGACTGTAGCAG TGGTGCCCCGTGTGAGGC-39... Input
59-******A******************A************A*********A TGGCGCCCGAACAGGGAC -39... 2/6
1g
59-******A******************A*A**********A*********A TGGCGCCCGAACAGGGAC -39... 1/6
1g
59-******A******************A************A*********A TGGCGCCCGAACAGGGAC -39... 3/6
1h
HXB2(Met-AC(143–146))
59-ACTAGAGATggaaCAGACCCTTTTAGTCAGTGTTGTGAGACTGTAGCAG TGGTGCCCCGTGTGAGGC-39... Input
59-*...*CCCT**********************A************* ******************-39... 4/7
59-*...*CCCT**********************A************* *****************A-39... 3/7
HXB2(Met-AC(143–146, 161–163))
59-AGATggaaCAGACCCTTTTAGTgtcTGTTGT GAGACTGTAGCAG TGGTGCCCCGTGTGAGGC AAA-39... Input
59-************************AC****C *********A**A TGGCGCCCGAACAGGGAC ..*-39... 3/5
1i
59-************************A****A* *********A**A TGGCGCCCGAACAGGGAC ..*-39... 2/5
1i
HXB2(Met-AC(D142–173))
59-AC TAGAGA... ...CTCTAGCAG TGGTGCCCCGTGTGAGGA-39... Input
59-** **********************************************A ******************-39... 5/6
1TAGAGACTCTAGCAGAAAT
59-** **********************************************A ****************** -39... 1/6
1TAGAGACTCTAGCAGAAAT AGTGAGGC1
aAsterisks indicate identity with the input sequence. Lowercase boldface letters denote initial nucleotide mutations in U5 (the lowercase t at nucleotide 145 indicates
a C-to-T mutation in the parent HXB2(Met-AC) virus genome. Dots indicate deletions. Arrows indicate insertions.
bFrequency of the DNA sequence of the PBS region obtained from independent clones. cNucleotides in the A-loop region which constitute a ACN sequence.
dPBS located at nucleotides 183 to 200 in the HIV-1 HXB2 isolate. The sequence TGGTGCCCCGTGTGAGGC is complementary to tRNAMet. The sequence TGGCGCCCGAACAGGGAC is complementary to tRNA3Lys.
eInsertion of AGACTGTAACAGTGGTGCCCCGTGTGAGGCAAAAGCGAAAGGG (the sequence underlined is complementary to tRNAMet).
fInsertion of TTGTGAGACTGTAGTAGTTGAGACTGTAGCAGTGGTGCCCCGTGTGAGGCAAAAGCGAAAGGGAAA (the sequence underlined is
com-plemetary to tRNAMet).
gInsertion of TATAGCAGTGGTGCCCCGTGTGAGGC (the sequence underlined is complementary to tRNAMet).
hInsertion of TATAGCAGTGGTGCCCCGTGTGAGGCAAAGCGAAAGGGAAGAGACTGTAGCAG (the sequence underlined is complementary to
tRNAMet).
iInsertion of GAGACTGTAGCAGTGGTGCCCCGTGTGAGGCAAAAGCGAAAGGGAA (the sequence underlined is complementary to tRNAMet).
on November 9, 2019 by guest
http://jvi.asm.org/
the 39 18 nucleotides of the tRNA used as the primer for reverse transcription during positive-strand synthesis, the pro-viral PBS sequence reflects which tRNA species was used to initiate reverse transcription (24). Isolation of the high-molec-ular-weight DNAs from the infected cells at 120 days of culture was followed by PCR amplification of the U5-PBS region, and the DNA sequences of individual subclones were determined (Table 1). Viruses derived from HXB2(Met-AC), which have an ACN in U5 to stably utilize tRNAMet(9), maintained the
PBS complementary to tRNAMet, as expected. Two G-to-A
point mutations were found at nucleotides 140 and 179 up-stream of PBS in most clones; these mutations result in an A-U
instead of a G-U base pair in the stem-loop structure of Fig. 1A. Interestingly, a few clones (two of nine) were obtained which contain two PBSs complementary to tRNAMetwith
ad-ditional nucleotides consistent with a duplication of the 13 nucleotides downstream of PBS and the 12 nucleotides up-stream of PBS.
Sequence analysis of the proviral PBS from viruses HXB2(Met-AC(157-161)), HXB2(Met-AC(153-161)), and HXB2(Met-AC(161-163)) revealed that all maintained a PBS complementary to tRNAMet, although we did not recover the
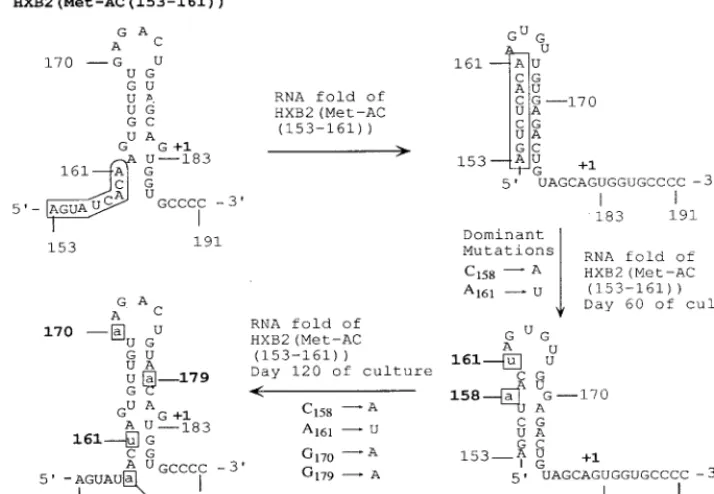
[image:7.612.124.481.80.327.2]starting proviral sequence in any of the clones analyzed. Most of the nucleotide changes were one- or two-base changes con-FIG. 5. Effects of additional mutations on recovery of the RNA stem-loop structure upstream of PBS in viruses derived from HXB2(Met-AC(153-161)). The secondary structures of RNA encompassing nucleotides 126 to 222 with input mutations and with additional mutations following in vitro culture were predicted by using the RNA folding algorithm MFold (32, 33). Only the nucleotides (153 to 191) forming a stem-loop structure upstream of PBS are shown. The input mutations between nucleotides 153 and 161 are in the rectangle. Individual mutations following in vitro culture are in lowercase and boxed.
FIG. 6. Additional mutations in U5 restore the secondary structures of RNA alone in viruses derived from HXB2(Met-AC(D142-173)). Dots shows deleted nucleotides 142 to 173 in the context of secondary structure of HXB2(Met-AC). The secondary structure with input deletion mutations and with the additional mutation of a 19-nucleotide insertion was predicted by the MFold RNA folding algorithm (32, 33). Inserted nucleotides are in boldface lowercase letters with line drawing. Boxed nucleotide 182 indicates a point mutation.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.55.553.539.691.2]sisting mainly of a G-to-A change (data not shown). Analysis of the proviral clones of the U5-PBS from viruses derived from HXB2(Met-AC(143-146)) revealed that all of the clones con-tained a deletion of nucleotides 135 to 141 with a change of nucleotides 143 to 146 (GGAA to CCCT). This reversion would restore the wild-type sequence for this virus similar to that found in HXB2(Met-AC) (CCUU versus CCCU). RNA modeling suggests that the CCUU sequence is critical for the interaction of the U5-PBS with tRNAMet.
Analysis of the U5-PBS from several of the viruses revealed many times two distinct PBSs complementary to tRNAMet. For
example, the virus derived from HXB2(Met-AC(162-164)) in four of six clones contained an insertion of a 68-nucleotide sequence which is composed of 34 nucleotides upstream, an 18-nucleotide sequence complementary to tRNAMet, and 16
nucleotides downstream; all of these inserted nucleotide se-quences were duplicated from the sequence upstream or
downstream of the PBS. All of the clones recovered from HXB2(Met-AC(143-146, 161-163)) at day 120 contained dual PBSs; the upstream PBS was complementary to tRNAMet, and
most importantly, the downstream PBS was a wild-type PBS complementary to tRNA3Lys. A two-nucleotide deletion
imme-diately downstream of the wild-type PBS suggested that a mis-match repair might have occurred during the second template switch in which a tRNA3Lyswas positioned at the PBS
comple-mentary to tRNAMet. In a previous study, we have shown that
both of the PBSs from viruses with dual PBSs can be utilized for initiation of reverse transcription (12, 13). It is possible, then, that the virus derived from HXB2(Met-AC(143-146, 161-163)) grows slower than the other mutant viruses because of interference between two different PBSs for reverse transcrip-tion. All of the clones recovered from the virus derived from HXB2(Met-AC(157-166)), with mutations encompassing the stem region of the stem-loop structure upstream of the PBS, contained a wild-type PBS complementary to tRNA3Lysat day
120 with eight nucleotides duplicated from the sequence up-stream of PBS (Table 1). Half of the clones examined also contained additional insertions of a PBS complementary to tRNAMetand 27 duplicated nucleotides (14 nucleotides
up-stream and 13 downup-stream of PBS). Again, mutations pre-dicted to completely disrupt the stem-loop structure in U5 upstream of PBS resulted in viruses reverting to contain a PBS complementary to tRNA3Lys.
A completely unexpected result was obtained from the anal-ysis of the virus derived from HXB2(Met-AC(D142-173)), which contained a 32-nucleotide deletion in U5 (Fig. 2). Upon extended in vitro culture, we found that this virus still stably maintained a PBS complementary to tRNAMet. All of the
clones recovered, however, had an additional 19 nucleotides inserted upstream of the PBS; 15 of these nucleotides were duplicated from nucleotides immediately upstream of the PBS of the input proviral genome. This is the first time that we have obtained viruses which contain a substantial change in U5 that were still able to maintain the use of an alternative tRNA as a primer for initiation of reverse transcription.
DISCUSSION
In this study, we have utilized a unique HIV-1 strain which stably maintains a PBS complementary to tRNAMetto
charac-terize U5-PBS interactions with the tRNA primer used for initiation of reverse transcription. RNA modeling of the U5-PBS alone and complexed with tRNAMet revealed striking
similarities to models of the wild-type U5-PBS alone and com-plexed with tRNA3Lys. To investigate the potential significance
of these RNA structures, we constructed mutations designed to disrupt critical elements within the secondary structures. All of the mutations resulted in viruses which initially had delayed replication compared to the parental virus, HXB2(Met-AC). By using an endogenous reverse transcription-PCR method, we found that the delays in replication correlated with a re-duced capacity for initiation of reverse transcription as deter-mined by the ability to synthesize (2)ss DNA primed with tRNAMet. Analysis of the U5-PBS following long-term in vitro
culture revealed that while most mutants were stable and maintained a PBS complementary to tRNAMet, others had
reverted to a virus with a PBS complementary to tRNA3Lys. In
some instances, viruses with complex genomes containing dual PBSs were observed. Interestingly, we found that viruses which initially contained a deletion of 32 nucleotides of the U5 region still maintained a PBS complementary to tRNAMetfollowing
in vitro culture. Sequence analysis of the U5-PBS of this virus FIG. 7. Additional mutations in U5 restore the secondary structure of RNA
complexed with tRNAMet. The models of viral RNA secondary structure com-plexed with tRNAMetwere predicted in the viruses derived from HXB2(Met-AC(D142-173)). tRNA sequences are in boldface lowercase letters.D32 indicates a 32-nucleotide deletion between nucleotides 141 and 174. (A) Input mutant showing enough complementary nucleotide interactions in regions II and III in U5. (B) Additional mutations caused by inserting 19 nucleotides upstream of PBS following culture, allowing formation of the secondary structure of viral RNA complexed with tRNAMetmore like HXB2(Met-AC), restoring the impor-tant interaction in region IV, as shown in the box (Fig. 1B). The insertion of 19 nucleotides between nucleotides 135 and 136 is in shadowed uppercase letters.
on November 9, 2019 by guest
http://jvi.asm.org/
revealed the presence of 19 new nucleotides in place of the original deletion.
In previous reports, we have described the effects that mu-tations in U5 have on the subsequent capacity of HIV-1 to stably utilize tRNAHisor tRNA
1,2
Lysto initiate reverse
transcrip-tion (9, 10, 29, 30, 31). Mutatranscrip-tions within the U5 region of these viruses all had drastic effects on the subsequent capacity to maintain a PBS complementary to these alternative tRNAs (30). One of the major differences between the virus which utilizes tRNAMetto initiate reverse transcription and the other
viruses which use other tRNAs to initiate reverse transcription is the ability of HXB2(Met-AC) to accommodate several dif-ferent types of mutations within U5 without reverting to a PBS complementary to tRNA3Lys. It is possible that this feature is
due to the fact that RNA structures of the U5-PBS of HXB2(Met-AC) alone and complexed with tRNAMetare
sim-ilar to that of the wild-type virus and, thus, stabilize the use of tRNAMetto initiate reverse transcription. Several of the results
from our analysis of different virus mutants support this idea. Analysis of the virus derived from HXB2(Met-AC-(143-146)) revealed that all of the clones had restored the CCCT nucle-otides which could potentially interact with the variable loop of tRNAMet(region IV in Fig. 1B). Previous studies in our
lab-oratory have shown that even in the wild-type genome, muta-tion in this region resulted in a virus which reverts to restore the CCCT motif (31). What is the importance of this interac-tion? One explanation could be that this interaction is required to maintain the disrupted tRNA structure necessary to form an initiation complex similar to what has been found by Isel et al. (5) (Fig. 1B). However, the CCCT reversion would not be predicted to completely restore the region IV interaction in HXB2(Met-AC(142-146)). Thus, it is possible that this motif is involved in other, as yet undefined, RNA-RNA interactions within the viral genome that result in additional selective pres-sure for the reversion to CCCT. The further analysis of the role of the CCCT motif in reverse transcription might be compli-cated because we found that in contrast to strain HXB2(Met-AC(143-166)), double mutant HXB2(Met-AC(143-146, 161-163)) did not contain this reversion following in vitro culture. Rather, this virus had mutated to contain a PBS complemen-tary to tRNA3Lys positioned downstream from the PBS
com-plementary to tRNAMet. In a related set of experiments, we
found that the wild-type virus containing a similar set of mu-tations also did not restore the CCCT motif (31). Why the virus derived from HXB2(Met-AC(143-146, 161-163) mutated to contain two PBSs is not clear. Virus HXB2(Met-AC(143-146,161-163)) maintained a slow-growth phenotype, even after extended culture (data not shown). It is possible that the po-sitioning of a PBS complementary to tRNA3Lys downstream
from the PBS complementary to tRNAMetinterfered with the
reverse transcription process. Previous studies in this labora-tory have shown that if the upstream PBS in mutant viral genomes with dual PBSs was complementary to tRNA3Lys, this
PBS was used predominantly for reverse transcription (12). Furthermore, viruses with this dual-PBS configuration did not show a delay in replication compared to the wild-type virus; this result was similar to what was found for HXB2(Met-AC(157-166)), in which a PBS complementary to tRNA3Lys is
positioned upstream of a PBS complementary to tRNAMet. In
contrast, the viruses derived from HXB2(Met-AC(143-146, 161-163)) contain an upstream PBS complementary to tRNAMet positioned in front of a PBS complementary to
tRNA3Lys. The combination of both PBSs occupied by
tRNAMetand tRNA 3
Lys, respectively, could affect the capacity
of the virus to undergo reverse transcription (12, 13). One of the hallmarks of the viruses with a PBS
complemen-tary to an alternative tRNA is the generation of numerous nucleotide changes in U5 following in vitro replication. Based on the results of our previous studies, we suggested that these mutations might be the result of selection of viruses which can undergo a more efficient initiation of reverse transcription (27). The results of our current study also support this idea. It was clear from our analysis of the mutant viruses constructed from HXB2(Met-AC) that mutations in the U5 region affected reverse transcription. Not surprisingly, those viruses derived from HXB2(Met-AC(157-166)), in which the mutants most severely affected the initiation of reverse transcription, also exhibited the slowest initial replication. Similar results have been obtained from analysis of the wild-type virus with muta-tions in U5 (31). Deletion of the four consecutive A’s in the U5 stem-loop of HIV-1 did not affect tRNA3Lysplacement on the
genomic RNA; however, viruses with the deleted A nucleo-tides exhibited decreased reverse transcription and slower rep-lication kinetics (6, 14). What was most interesting, however, was that the A loop was restored over time during in vitro culture (14). Analysis of the U5 region from the virus derived from HXB2(Met-AC(153-161)) revealed a similar situation after in vitro culture. Modeling of the RNA stem-loop of the U5-PBS from nucleotide 126 to nucleotide 222 revealed that the initial mutations would have resulted in a stem-loop struc-ture containing different nucleotides in the region postulated to interact with the anticodon loop of tRNAMet(Fig. 5). RNA
modeling of the U5 of viruses obtained after in vitro culture revealed an apparent evolution toward recovery of the stem-loop structure seen for the U5 derived from HXB2(Met-AC). Most importantly, the region of U5 complementary to the anticodon of tRNAMetcan be displayed on the loop region of
a stem-loop structure similar to that for HXB2(Met-AC). In contrast to HXB2(Met-AC(153-161)), viruses derived from HXB2(Met-AC(157-166)), with mutations predicted to com-pletely disrupt this stem-loop structure, displayed a greater defect in initiation of reverse transcription and eventually re-verted to contain the wild-type PBS. Why then do some mu-tations lead to reversion of the PBS to complementarity to tRNA3Lys? One possibility is that mutations that disrupt the
U5-PBS structure result in an RNA genome encapsidated in the virus without a tRNA positioned at the PBS complemen-tary to an alternative tRNA. If this were to happen, upon maturation of the virus particle, the nucleocapsid protein (p7) and/or RT might force tRNA3Lys to be positioned at the PBS
complementary to the alternative tRNA. If initiation of reverse transcription occurred, the virus would have a potential for two PBSs, one complementary to tRNAMet (for negative-strand
DNA) and one complementary to tRNA3Lys(for positive-strand
DNA), which would lead to viruses with dual PBSs. Further replication of these viruses might lead to a stable virus with dual PBSs (which was observed) or even resolution of the dual PBSs to a single wild-type PBS complementary to tRNA3Lys
(12).
Finally, compelling evidence for a general role of RNA structures in the selection of the tRNA and reverse transcrip-tion comes from the analysis of the virus which contained a 32-nucleotide deletion in U5. Remarkably, this virus was still able to undergo reverse transcription and exhibited replication kinetics similar to that of the parental virus. Molecular mod-eling of the RNA structures of the virus containing the addi-tional 19-nucleotide insertion revealed a similar overall struc-ture for the U5-PBS genome alone (Fig. 6), as well as in the presence of tRNAMet(Fig. 7). The additional 19 nucleotides in
the virus derived from HXB2(Met-AC-(D142-173)) were es-sential to restore similar stem-loop RNA structures modeled from HXB2(Met-AC) (Fig. 1A). The additional nucleotides
on November 9, 2019 by guest
http://jvi.asm.org/
would also be predicted to restore the interaction at region IV of the U5-PBS complexed with tRNAMet(Fig. 7). Taken
to-gether, the results of our study support the concept that RNA structures of the U5-PBS are important for the interaction with the tRNA used for initiation of reverse transcription.
ACKNOWLEDGMENTS
We thank Zhijun Zhang and Qin Yu for helpful comments and Dee Martin for preparation of the manuscript. C.D.M. thanks MAR for continued support.
Virus culture was carried out in the UAB AIDS Center Virus Cul-ture Core (AI 27767). This work was supported by grants AI-34749 and GM 56544 to C.D.M.
REFERENCES
1.Baltimore, D.1970. Viral RNA-dependent DNA polymerase. Nature (Lon-don)226:1209–1211.
2.Baudin, F., R. Marquet, C. Isel, J.-L. Darlix, B. Ehresmann, and C. Ehres-mann.1992. Functional sites in the 59region of human immunodeficiency virus type 1 RNA form defined structural domains. J. Mol. Biol.229:382– 397.
3.Das, A. T., B. Klaver, and B. Berkhout.1995. Reduced replication of human immunodeficiency virus type 1 mutants that use reverse transcription primer other than the natural tRNA3Lys. J. Virol.69:3090–3097.
4.Gilboa, E., S. W. Mitra, S. Goff, and D. Baltimore.1979. A detailed model of reverse transcription and tests of crucial aspects. Cell18:93–100. 5.Isel, C., C. Ehresmann, G. Keith, B. Ehresmann, and R. Marquet.1995.
Initiation of reverse transcription of HIV-1: secondary structure of the HIV-1 RNA/tRNALys,3(template/primer) complex. J. Mol. Biol.247:236– 250.
6.Isel, C., J.-M. Lanchy, S. F. J. L. Grice, C. Ehresmann, B. Ehresmann, and R. Marquet.1996. Specific initiation and switch to elongation of human immunodeficiency virus type 1 reverse transcription require the post-tran-scriptional modifications of primer tRNALys,3. EMBO J.15:917–924. 7.Jiang, M., J. Mak, A. Ladha, E. Cohen, M. Klein, B. Rovinski, and L.
Kleiman.1993. Identification of tRNAs incorporated into wild-type and mutant human immunodeficiency virus type 1. J. Virol.67:3246–3253. 8.Kang, S.-M., J. K. Wakefield, and C. D. Morrow.1996. Mutations in both the
U5 region and primer binding site influence the selection of the tRNA used for the initiation of HIV-1 reverse transcription. Virology222:401–414. 9.Kang, S.-M., Z. Zhang, and C. D. Morrow.1997. Identification of a sequence
within U5 required for human immunodeficiency virus type 1 to stably maintain a primer binding site complementary to tRNAMet. J. Virol.71:207– 217.
10. Kang, S.-M., Z. Zhang, and C. D. Morrow.Identification of a human im-munodeficiency virus type 1 which stably uses tRNA1,2Lysrather than tRNA3Lys for initiation of reverse transcription. Submitted for publication. 11.Li, X., M. Johnson, E. J. Arts, Z. Gu, L. Kleiman, M. A. Wainberg, and M. A.
Parniak.1994. Effects of alterations of primer-binding site sequences on human immunodeficiency virus type 1 replication. J. Virol.68:6198–6206. 12. Li, Y., S.-M. Kang, and C. D. Morrow.1997. Stability of HIV-1 proviral
genomes which contain two distinct primer binding sites. AIDS Res. Hum. Retroviruses13:253–262.
13. Li, Y., Z. Zhang, S.-M. Kang, J. L. Buescher, and C. D. Morrow.1997. Insights into the interaction between tRNA and primer binding site from characterization of a unique HIV-1 virus which stably maintains dual PBS complementary to tRNAGlyand tRNAHis. Virology238:273–282.
14.Liang, C., X. Li, L. Rong, P. Inouye, Y. Quan, L. Kleiman, and M. A. Wainberg.1997. The importance of the A-rich loop in human immunodefi-ciency virus type 1 reverse transcription and infectivity. J. Virol.71:5750– 5757.
15.Mak, J., and L. Kleiman.1997. Primer tRNAs for reverse transcription. J. Virol.71:8087–8095.
16. Marquet, R., C. Isel, C. Ehreshmann, and B. Ehreshmann.1995. tRNA as primer of reverse transcriptase. Biochimie77:113–124.
17. Masuda, T., V. Planelles, P. Krogstad, and I. S. Y. Chen.1995. Genetic analysis of human immunodeficiency virus type 1 integrase and the U3 att site: unusual phenotype of mutants in the zinc finger-like domain. J. Virol.
69:6687–6696.
18. Peters, G., and J. E. Dahlberg.1979. RNA-directed DNA synthesis in the Moloney murine leukemia virus: interaction between the primer tRNA and the genome. J. Virol.31:398–407.
19. Ratner, L., A. Fisher, L. Linda, J. Agodzinski, H. Mitsuya, R.-S. Liou, R. C. Gallo, and F. Wong-Staal.1987. Complete nucleotide sequences of func-tional clones of the AIDS virus. AIDS Res. Hum. Retroviruses3:57–69. 20. Rhim, H., J. Park, and C. D. Morrow.1991. Deletions in the tRNALysprimer
binding site of human immunodeficiency virus type 1 identify essential re-gions for reverse transcription. J. Virol.65:4555–4564.
21. Sambrook, J., E. F. Fritsch, and T. Maniatis.1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
22. Sanger, F., S. Nicklen, and A. R. Coulson.1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463–5467. 23. Sarker, G., and S. S. Wimmer.1990. The “megaprimer” method of
site-directed mutagenesis. BioTechniques8:404–407.
24. Taylor, J. M., and T. W. Hsu.1980. Reverse transcription of avian sarcoma virus RNA into DNA might involve copying of the tRNA primer. J. Virol.
33:531–534.
25. Temin, H. M., and S. Mizutani.1970. RNA-directed DNA polymerase in virions of Rous sarcoma virus. Nature226:1211–1213.
26. Vicenzi, E. D., S. Dimitro, A. Engleman, T. Migone, D. F. J. Purcell, J. Leonard, G. Englund, and M. A. Martin.1994. An integration-defective U5 deletion mutant of human immunodeficiency virus type 1 reverts by elimi-nating additional long terminal repeat sequences. J. Virol.68:7879–7890. 27. Wakefield, J. K., S.-M. Kang, and C. D. Morrow.1996. Construction of a
type 1 human immunodeficiency virus that maintains a primer binding site complementary to tRNAHis. J. Virol.70:966–975.
28. Wakefield, J. K., A. G. Wolf, and C. D. Morrow.1995. Human immunode-ficiency virus type 1 can use different tRNAs as primers for reverse tran-scription but selectively maintains a primer binding site complementary to tRNALys,3. J. Virol.69:6021–6029.
29. Zhang, Z., S.-M. Kang, A. LeBlanc, S. L. Hajduk, and C. D. Morrow.1996. Nucleotide sequences within the U5 region of the viral RNA genome are the major determinants for a human immunodeficiency virus type 1 to maintain a primer binding site complementary to tRNAHis. Virology226:306–317. 30.Zhang, Z., S.-M. Kang, Y. Li, and C. D. Morrow.1998. Genetic analysis of
the U5-PBS of a novel HIV-1 reveals multiple interactions between the tRNA and RNA genome required for initiation of reverse transcription. RNA4:394–406.
31. Zhang, Z., S.-M. Kang, and C. D. Morrow.1998. Genetic evidence for the interaction between tRNALys,3and U5 facilitating efficient initiation of re-verse transcription by human immunodeficiency type 1. AIDS Res. Hum. Retroviruses14:979–988.
32.Zuker, M.1989. On finding all suboptimal foldings of an RNA. Science
244:48–52.
33. Zuker, M., and P. Stiegler.1981. Optimal computer folding of large RNA sequences using thermodynamics and auxiliary information. Nucleic Acids Res.9:133–148.