Copyright©1971 American Society for Microbiology PrintedinU.S.A.
Virus
Interference by Cellular
Double-Stranded
Ribonucleic Acid
P. C. KIMBALL AND P. H. DUESBERG
Department ofMolecular Biology and Virus Laboratory, University ofCalifornia, Berkeley, California 94720
Receivedforpublication8February 1971
Ribonuclease-resistant ribonucleic acid (RNA) was isolated from uridine-labeled cultures of rabbit kidney, chicken embryo, and HeLa cells. This RNA, regardless of itssource, wasfoundtoinduce interference with virus growth in either rabbit kidney or chicken embryo cultures. Nuclease-treated cellular nucleic acids exhibited interference-inducing activity which eluted witha smallfraction of RNA in the exclusion volume ofa 6% agarose gelcolumn. Besides resistance to ribo-nucleases, the interference inducer and RNA isolated from partiallydigestednucleic
acids have in common two properties of double-stranded RNA: (i) similar sharp melting profiles were obtained for inducer and ribonuclease-resistant RNA, with
TmdependentonNaCl concentration; (ii) ribonuclease-resistant inducer andRNA
banded together in Cs2SO4 density gradients at a density characteristic of known double-stranded RNA. After melting at low ionic strength, the labeled RNA shiftedto ahigher density and its capacitytoinhibit virus replication waslost.
Ve-locity sedimentation analysis of the cellular ribonuclease-resistant RNA indicated
that themajority sedimented between7and 1 S, but onlyRNAsedimentingat .8
to20Shadahigh specific activity of interference induction. Without prior
ribonu-cleasetreatment, the ribonuclease-resistant RNAcanbe precipitated with2 MLiCl
and thusappearstoexist in purified cellular nucleic acidsaspartof molecular com-plexes with both single- and double-stranded regions of RNA. Thebiosynthesis of
cellulardouble-stranded RNA is inhibited by actinomycin D.
Interferon
isinduced
by
a varietyof
viruses,often
underconditions
where little or noviral
multiplication
occurs(16).
It wassuggested,
therefore,
that some partof
the virus particleitself,
perhaps the"foreign"
nucleic acid, could bethe
stimulus for induction
(18). Early
experimentsdesigned
to testthis
theory
by
treatmentof
agiven
species of animal
orcellsin
culture with diversenucleic acids
supported
thenotion
thatnucleic
acids from
heterologous
animal species couldinduce
interferon (20-22,
27, 30), whereas thosefrom
homologous cells
were less potent orinac-tive
(20,
22,27) unless
chemically modified (20).Consequently,
thediscoveries
of severalin-ducers which
apparently
contained no nucleicacid
(15, 17)
resultedin questioning of
thetheorythat a
foreign
nucleicacid was theessentialstimu-lus
forinterferon
induction (19). Conflictingre-sults
in furthertests(3, 7)
haveleft the issue aboutthe
"foreign" origin of inducing
ribonucleic acid(RNA)
unresolved.Recently,
it becameclear thatsynthetic
ornatural double-strandedRNAspecieswhich
arenotendogenous
to the animal systemused
for
assay areextremely
potent interferon inducers(11),
whereas puredeoxyribonucleic
acid
(DNA),
DNA-RNAhybrids,
andsingle-stranded RNA were found to be
inactive
(4,
5,15).
These
findings
and thediscovery
thatfrom
about
0.01 to 1.0%of
RNA in animal cells isribonuclease-resistant (6,
24)
orreactive
withanti-bodies
specific for double-stranded
RNA(28)
led
us to suspectthattraces
of
theendogenous
ribo-nuclease-resistant RNA
could
be theinterferon
inducer in
cellular nucleic acids
previously
thought to
contain
RNAonly
insingle-stranded
form. Furthermore,
thesuspected
presenceof
double-stranded
RNA in normal cells raised thequestion
of
whetherhomologous
as well asforeign double-stranded
RNA can induceinter-feron in
agiven species of
cells. We present herefurther
characterization
of
cellularribonuclease-resistantRNAandshow that it isindeed double-stranded.Part
of
thisRNAiscapable
ofinducing
interference with virus growth ina
given species of
cells, irrespective
of whether it is isolated fromaheterologous
orhomologous
species.
MATERIALS AND METHODS
Cellcultures. For
preparation
ofrabbitkidney
pri-marycultures, mincedkidneysfrom 3-to5-week-old
New Zealand White rabbits were stirred gently at 697
on November 11, 2019 by guest
http://jvi.asm.org/
room temperature for 1 to 2 hrin a solution ofEagle's minimal essential medium (MEM; Grand Island Bio-logical Co.) containing 0.25% Pronase. The cell sus-pension resulting from one kidney pair was washed andplated in MEMcontaining 10% calf serumon a
total tissue culture dish surfacearea of about 1,000 cm2. Withamediumchange thenextday, confluency was reached in5 to 7 days after seeding. A line of HeLacells(HeLaI) from C.Colby,Jr.(Universityof
Connecticut, Storrs) and HeLa S3 cells (HeLa II) from M. Bishop (University of California, San Fran-cisco) werealso grown inMEMplus 10% calfserum.
For preparation of RNA, primary or secondary chicken embryocultures (26) weregrowninmedium
199 (GrandIslandBiological Co.) supplemented with
2%Tryptose phosphate broth and
1%
each calf and chicken sera. Periodicinspection ofallcell types by electronmicroscopy showednoevidence of contami-nation with mycoplasma.Buffers. Low-salt buffer contained 0.01 M NaCl in additionto0.01 Mtris(hydroxymethyl)aminomethane (Tris)-hydrochloride (pH 7.4) and 1 mm ethylenedi-aminetetraacetic acid (EDTA). High-salt buffer was thesameexceptfor the NaCl concentrationwhichwas 0.3 M.
Preparation ofanuclease-resistant fraction of
cellu-lar 3H-RNA. Confluent monolayers were labeled by adding50 to 100,uCiof3H-uridine per 100-mmpetri
dish after 3to4hrofpreliminaryincubation in 4 ml of the freshgrowth mediumappropriatefor each cul-ture type as indicated above. Actinomycin D was added, whereindicated, at 6,ug/ml for 15 min before theadditionof radioactive precursor. After 90to120 min oflabeling, the nuclease-resistant RNA fraction
was prepared from purifiedcellular nucleic acids as
described earlier (6).Forsamplesofnucleic acidstobe treated with deoxyribonuclease alone, ribonuclease-free pancreatic deoxyribonuclease I (Worthington Biochemical Corp.) was used. Otherwise, the nucleic acids were incubated first with standard grade pan-creatic deoxyribonuclease Iand then with pancreatic
ribonuclease, following published procedures (6). When the partially digested cellular nucleic acids
werechromatographically analyzedon acolumn of 6% agarose gel, the exclusion volume (determined as in reference 6) routinelycontained the first two or three fractions of material labeled with 3H-uridine. These
were pooled andextractedtwice with phenolbefore precipitation with ethanol. At this stage, a visible precipitate often formed which madeaturbid solution whenredissolved in_1mlof low-saltbuffer(without
EDTA). This turbidity was largely removed from solutions without loss of radioactive RNA by centri-fugationat 150,000 X g for 10min, after which the supernatantfractionwas storedat -20C. This frac-tion is referredto hereafter as the nuclease-resistant RNAorcellular double-stranded RNA.
Standard nuclease treatments andthermal
denatura-tionofRNA.Thestandardribonucleasetreatmentwas with20,jgofpancreaticribonuclease per ml and0.5,ug
ofT, ribonuclease per ml (Worthington Biochemical Corp.) in0.8ml ofhigh-saltbuffer for30minat37C. The standard ribonuclease plus deoxyribonuclease
treatmentwasthesameexcept that50,ugof
deoxyribo-nuclease I perml and2 mmMgCl2 werealso present.
To standardize the ratio of enzyme to nucleic acid, yeast soluble RNA (sRNA; CalBiochem) wasadded (60 ,g/ml) to all samples with <0.01 absorbancy (A)260/ml. Thermal denaturation of ribonuclease-re-sistant RNA was done in low-salt buffer asreported
earlier (6).
Fractionation of RNA with 2M LiCl solution.
Purified total cellular nucleic acidswere treated with
deoxyribonuclease, extracted with phenol, ethanol-precipitated, and washed with ethanol as described
previously for preparation of the nuclease-resistant fraction of cellular RNA. The
deoxyribonuclease-treated nucleic acids were dissolved in low-salt buffer at<_10
A260/ml,
and sufficient LiCl insolu-tion (10M) was introducedtogiveafinal
concentra-tion of2M.After 16 hr or more at0C, the solution
was centrifuged for 15 min at 20,000 X g. Three volumes oflow-saltbufferwasaddedtotheresulting
supernatantfraction and several milliliterswasadded to thepellet. Then the nucleic acids in each fraction
wereethanol-precipitated and washed.
Virusinterference assays. Assayswere done in
dupli-catebyusing0.1 ml of medium per cellculture in
6-mm wells of microtest tissue culture plates (Falcon Plastic). The assaycultures were either 7- to 14-day-old rabbit kidney cell primarycultures prepared and grown asdescribed above or chicken embryo primary
cultures plated at 5 X 104 cells per 6-mm well and incubated for 5 to 6 days in NCI Medium (Schwarz BioResearch) supplemented with 2% Tryptose phos-phatebroth and1%S eachcalf and chicken sera. Sam-ples of RNA to be tested for interference with virus replication were precipitated in ethanol with carrier yeastsRNA anddissolveddirectlyinmedium 199with
2%o fetal calf serum and 10 ,g of
diethylaminoethyl-dextranperml (molecular weight = 2 X 106, Phar-macia) ataconcentration of<100ugof RNA perml. This concentration of sRNA in 0.1 ml failedtoinduce resistance to viruses and did not affect interference
induced by the synthetic polymer poly rI:rC (Miles Laboratories, Inc.).
After 18to24hrof exposuretonucleic acid solu-tions, chicken orrabbit cells were washed with Tris-buffered saline (TS; reference 8) and challenged,
respectively, with either Sindbis virus or vesicular stomatitis virus (VSV, Indianastrain)at amultiplicity of about5plaque-forming units (PFU) per cell. Virus
was applied in 20
Muliters
of virus growth medium (NCIplus6%calfserum forSindbis, MEMplus 10% calfserumfor VSV) which was removed aftera 1 hrperiod of adsorption. Then cells were washed twice withTSbeforebeingrefedwith0.2ml ofappropriate virus growth medium. After 18 to 24 hr of further incubation, cultures were examined for
cytopatho-logical effects (which correlated with viral growth), and theplates were frozenat-70 Cuntil the medium
wasanalyzedbyplaqueassayonchick embryo second-ary monolayers as previously described for Sindbis virus(6).
Control samples ofastandard interferon inducer, poly rI:rC, were included in all experiments when interfering activity of cellular nucleic acids was as-sayed.PolyrI:rCreducedvirus growth to titers below
on November 11, 2019 by guest
http://jvi.asm.org/
detectionataconcentration of 0.1
Ag/ml
orlesswhencultures without inducergave106to 107 PFU/ml for
either the chicken orrabbit cell system. Results are
reportedasthelog10(tothenearest0.1)of theratio of virustiter inuninduced control culturestotheaverage
titer intwoparallel culturestreatedwithagiven
sam-pletobe assayed.This ratio isreferredtoaslog10virus
titer reduction, andin practice duplicateassays gave
reductions differing by _0.5 log10 Since, ingeneral,
the log1o virus titer reduction was not found to be linear with inducer RNA concentration, no attempt
wasmadetoquantify differencesinRNA concentra-tion of compared samples when they caused titer
reductions which varied outside the range of
dupli-cates.Weused this titer-reductionassay as ameasure ofinterferon induction because it has been shownto be more sensitive than assays for extracellular
inter-feron (5, 31), even before adaptation to microassay procedure.
RESULTS
Isolation ofaribonuclease-resistant interference inducerfromcellular nucleic acids. To answer the general question of whether the cellular nucleic
acids ofa given species contain double-stranded
RNA abletoinduceinterferonin thesamespecies
orotherspeciesofcells,thefollowing experiments
werecarriedout.Thenuclease-resistantRNAwas
preparedfrompurifiedtotal cellular nucleic acids of chicken embryo fibroblasts or rabbit kidney
cells by treatment with deoxyribonuclease and
ribonuclease and chromatographic fractionation
of thedigestasoutlinedabove.Onlythatfraction
of the ribonuclease-resistant RNA which eluted in the void volume of the 6% agarose column was
thenanalyzedfor itsabilitytointerfere with virus
replication. In all four tested cell types, about 0.4% of the total 3H-uridine incorporated into RNA during a 90-min pulse was in the
ribonu-clease-resistant fraction excluded by 6% agarose
(Table 1).Thisamounted to less than 0.1% of the totalA26oin thedigest.
Nuclease-resistant RNA from chicken, rabbit,
or HeLa cells depressed virus growth in both rabbitkidneyand chickenembryocell
cultures-in all cases to a level lower than 2 log10 below
control titers. By contrast,the same amounts of
totalcell nucleic acids whichwereused toisolate
the interference-inducing, ribonuclease-resistant
RNA did not showanyinterfering activitybefore
digestion with deoxyribonuclease and ribonu-clease. After nuclease treatment, however, samples of unfractionated nucleic acids from all cells interferedwith virusreplicationtothesame extent (within 0.5 log10 virus titer reduction) as
thepurifiedribonuclease-resistant fraction alone. These findings are summarized in Table 1. De-oxyribonuclease treatment alone also rendered cellular nucleic acids active ininducing
interfer-ence with virus
replication (data
not shown). Furthermore, insomeexperiments,
partof the cellsuspension
from either one chicken embryo orrabbitkidneywasculturedfor interference assays, whereas the rest of the cells were grown as a source of labeled
ribonuclease-resistant
RNA. Thus induction of virus interference by the pre-sumeddouble-stranded RNAfraction of
chickenor rabbit cell nucleic acidswasdemonstrablenot
only
in cells of the sameor differentspecies, but
also
incells derived from thesamepiece of
tissueas
those
from which theribonuclease-resistant
RNAwas
prepared.
Further
nuclease
treatmentsof
the 3H-RNApurified from
degraded
cellular RNA and DNAby
chromatography
on6%o
agarose wereper-formed
under the standard conditions(see
above)at
high ionic strength.
Theresistance
toribo-nuclease
(or
amixtureof ribonuclease
plusdeoxy-ribonuclease)
of this fraction of
3H-RNA ranged from 50% togreaterthan95%0
in differentprep-arations from
eachcell type. However, nosignifi-cant change
(.0.5 log10
virustiter
reduction)
inthe
ability
of
these RNAfractions
tointerfere
withvirus
replication
wasobserved
after standardnucleasetreatments
of
sevenof
eight
RNAsam-ples
from chicken and
HeLa cells(data
notshown).
After thermaldenaturation
(Materialsand
Methods) of
thepurified
ribonuclease-resis-tantfraction
of
cellular RNAat 100C,
less than5% of
the 3H-RNA remainedresistant
to thestandard
ribonuclease
treatment and allinterfer-ing activity
waslost(Table 1).
The
finding
that thenuclease-resistant
fraction
of cellular
RNA is resistant to bothdeoxyribo-nuclease and
ribonuclease
arguesagainst
thepossibility
that thisribonuclease-resistant
3H-RNAwhich interferes with virus
replication is in
DNA-RNA
hybrids
because DNA-RNAhybrids
are
known
to besensitive
todeoxyribonuclease
(25)
or acombined
nuclease treatment(32).
However,
double-stranded
RNAwould
beex-pected
both to resist ribonuclease athigh ionic
strength
and to bemade
sensitiveby
melting atlow
ionic
strengthbefore ribonuclease
treatment(14).
Inaddition, only
such RNA is known toshowa
high
specific activity of
induction ofvirus
interference (4,
5,15).
It can be estimated fromthedata
in
Table1that less than 0.05,ug (
<0.01A26o/ml)
of ribonuclease-resistant RNA per ml reduced titers oftwodifferent virusesbyasmuchas 5
log10
in two different cellspecies.
Togetherthese observations suggest that the ribonuclease-resistant
fraction of
cellularRNAwhichinterferes
with virusgrowth is double-stranded RNA. Interference-inducing RNA appears to be
par-tially
single-stranded before ribonucleasetreat-ment. To determine whether the RNA in the
7,
on November 11, 2019 by guest
http://jvi.asm.org/
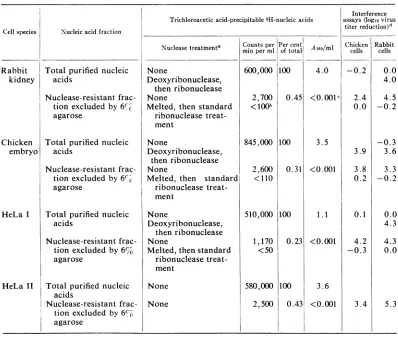
TABLE 1. Initerference with virusgrowthby cellular nucleic acids before antd after ntuclease treatments
Cellspecies
Rabbit kidney
Nucleic acid fraction
Total purified nucleic acids
Nuclease-resistant frac-tion excluded by6%c agarose
Chicken Total purified nucleic
embryo acids
Nuclease-resistant frac-tion excluded by6%C agarose
Totalpurified nucleic acids
Nuclease-resistant frac-tion excluded by6%
agarose
Total purified nucleic
acids
Nuclease-resistant frac-tion excluded by6% agarose
Trichloroacetic acid-precipitable 3H-nucleicacids Nuclease treatmenta
None
Deoxyribonuclease,
thenribonuclease None
Melted, then standard ribonuclease treat-ment
None
Deoxyribonuclease,
then ribonuclease
None
Melted, then standard ribonuclease treat-ment
None
Deoxyribonuclease,
thenribonuclease
None
Melted,thenstandard ribonuclease treat-ment
None None
Countsper Per
centi
min perml of total600,000 100
2,700
<10b
0.45
845,000
1100
2,600
<110
0.31
510,000 1100 1,170
<50
580,000 2,500
0.23
100
0.431 <0.001
aDeoxyribonuclease and ribonuclease treatments of the total nucleic acids are described for the
preparation of anuclease-resistant fraction of cellular 3H-RNA.Melting was performed at 100 C in low-salt buffer. Theprocedureis presented in Materials and Methodswiththe details of the standard ribonucleasetreatment.
bNotcorrectedforbackground.
cEstimated amount ina10-folddilution of theoriginalsolution which read<0.01A260/ml.
dDefined in Materials andMethodsunder virus interferenceassays.
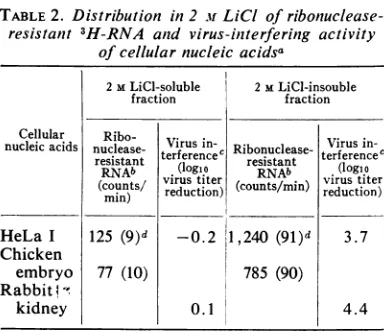
nuclease-resistant fraction of cellular nucleic acids
wascompletely oronly partiallydouble-stranded
before ribonucleasedigestion,itssolubilityin 2M
LiCl was examined. It is known that
double-stranded RNA is soluble but partially double-stranded RNA is insoluble in 2M LiCl (1). After
deoxyribonucleasetreatment,total cellular nucleic acids werefractionated with 2 M LiCl according
totheproceduredescribedabove. Then the super-natant and pelleted nucleic acids were either
assayed directly for virus-interfering activity or digested withribonuclease before isolation of the nuclease-resistant fraction of 3H-RNAbyagarose
column chromatography (see above). Over 90%0
of the total ribonuclease-resistant 3H-RNA in
HeLa Iorchicken embryo cell nucleic acidswas
found in the LiCl-insoluble pellet. Moreover, interference-inducing activitywasdetectable only
in the precipitated fraction ofHeLa I or rabbit kidney cell nucleic acids. These results are
sum-marized in Table 2. Under thesame conditions, only
12%7
(195 of 1,560 counts/min) of added influenza virus double-stranded 32P-RNA (10) was precipitated with HeLa I cellulardouble-stranded RNA. Therefore, precipitation in 2 M
LiClof theinterference-inducing activity withthe
ribonuclease-resistant RNA suggests that, in the total purified nucleic acids of the cell, the nuclease-resistant inducer could be the
double-stranded coreofan RNAcomplex thatalso has Interference assays(logiovirus
titerreduction)d
A260/ml
4.0
<O.OOlc
3.5
<0.001
1.1
<0.001
3.6 HeLa I
HeLa II
Chicken cells
-0.2
2.4
0.0
3.9
3.8 0.2
0.1
4.2
-0.3
3.4
Rabbit cells
0.0 4.0
4.5 -0.2
-0.3 3.6
3.3
-0.2
0.0
4.3 4.3
0.0
5.3
on November 11, 2019 by guest
http://jvi.asm.org/
TABLE2. Distribution in 2 MLiCI of ribonuclease-resistant 3H-RNA and virus-intterfering activity
of cellular nucleic acidsa
2MLiCI-soluble 2MLiCI-insouble fraction fraction
Cellular Ribo- Viu nVirus
in-nucleicacids nuclease- Virus Ribonuclease-
terferencec
resistant ererence resistant (oi
(logA
s RNAb (logirstie(counts/ virustiter (counts/min.,) rustiter
min) reduction) reduction)
HeLa I 125 (9)d -0.2
11,240
(91)d 3.7Chicken
embryo 77 (10) 785 (90)
Rabbit
1-kidney 0.1 4.4
aTotal purified nucleic acids were treated with deoxyribonuclease and then fractionated with 2 M LiCI.
bThe 2 M LiCi-soluble and -insoluble nucleic acids were treated with ribonuclease, and the ribonuclease-resistant fraction of RNA was puri-fied by agarose gel exclusion chromatography as described for preparation of the nuclease-resistant fraction of total cellular nucleic acids.
c After precipitation with ethanol, the nucleic
acids in theLiCl-soluble and -insoluble fractions wereassayed for interference with thereplication ofvesicular stomatitis virus on rabbit kidneycells.
dNumbers in parentheses are expressed as per cent.
single-stranded
regions
whichcauseprecipitation
at
high ionic strength.
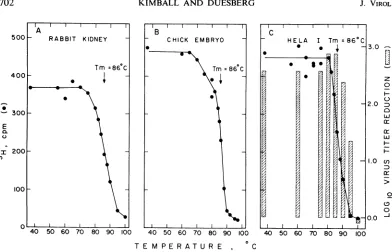
Melting
profiles ofnuclease-resistant
RNA andinterference inducer. As
expected for
an RNAduplex
(20),
melting
profiles with sharp
transi-tions
(at
86 C in 0.01 MNaCI)
from ribonuclease
resistance
toribonuclease
sensitivity
wereob-tained
for thepurified
nuclease-resistantfractions
of the different
cellular
RNAspecies (Fig. 1A,
B andC).
In the caseof
HeLa Inuclease-resistant
RNA, the
ability
to induce virus interference in rabbitkidney
cultures wascompletely
lost whenthe
3H-RNA
wascompletely
melted(Fig. IC).
Measurement of the
T,,,
of cellulardouble-stranded RNAon the basis of its
interfering
ac-tivity
in thevirus titerreduction
assaywould
re-quire both
the measurementof
atwofold
(0.3
logls)
virus titerreduction
and theassumption of
someprecise relationship between the
concentra-tion of
interference-inducing
double-strandedRNA and titer reduction. Our
experiments
werenot
designed
to determine either such a smallchange in virus titer or the exact nature of the
relation between
inducing
RNA concentration and titerreduction(see
above forthelimitsof the interferenceassay).
To test whether the
T,,,
of the material whichinterferes with virus growth is increased with in-creasing ionic strength as has been shown for double-stranded RNA (2), HeLaI ribonuclease-resistant 3H-RNA was heated at 100 C in high-salt buffer with 0.3 M NaCl (see above). It was
found that 21
%/o
(95counts/min) of the 3H-RNA and enough virus-interfering activity to give 1.5log1o
virus titer reduction remained resistant toribonuclease after heating under these conditions. By contrast, less than 3% of the 3H-RNAand no measurable interfering activity (-0.1 log10 virus titer reduction) survived the standard ribonu-clease treatment after heating to 100 C at low ionic strength (Fig. IC). This indicates that the
T,-
for ribonuclease resistanceof the interference-inducing 3H-RNA was increased with increasing ionic strength.Buoyant density in CS2SO4 of ribonuclease-resistant RNA and inducer of virus interference beforeandafter
melting.
To examine further thesuggested identity
between thepurified
ribonu-clease-resistant interference inducer and double-stranded RNA, the density distributions of 3H-RNAand inducerweredetermined
in gradients
of Cs2SO4. In all cases, theribonuclease-resistant
3H-RNA banded in a sharp peak at adensity of 1.61 to 1.63
g/ml.
Single-stranded tobacco
mosaic virus (TMV) RNA marker hadadensity
of 1.68 g/ml, and salmon sperm DNA had adensity
of less than 1.50 g/ml under the same conditions. Interference assays on rabbitkidney
cells showedhigher
levels of antiviralactivity only in
thegra-dient fractions
containing
thepeak of
thenuclease-resistant
3H-RNA(Fig. 2A,
B,and
C).The
density of
thebiologically active
RNAfrac-tion
is ingood
agreement with thatof known
double-stranded
RNA(29)
and is much greaterthan those
reported
for DNA-RNA hybrids (1.49 to 1.54 g/ml;references
9, 29). When asample of ribonuclease-resistant
3H-RNAfrom
eachcell typewasmeltedat100Cbefore
analysis,
the 3H-RNA and TMV RNA marker
sub-sequently
bandedtogether
at 1.68g/ml
(Fig.
2D,E,andF), and
interference-inducing capacity
waslost.
Similardensity
shifts and lossof
activity
werenoted after treatment of ribonuclease-resistant
RNA with
97%NO
dimethylsulfoxide (data
notshown).
Thetemperature-dependent
density
tran-sition of
theribonuclease-resistant
3H-RNA isconsistent with the melting of an RNA
duplex
(29),asistheloss ofability to induce virus inter-ference
(6).
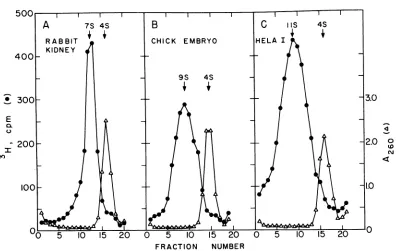
Sedimentation
properties
of the interference-inducing nuclease-resistantRNA before and afterstandard nuclease treatments. The ribonuclease-resistant 3H-RNA
separated
from cellular nucleicacid
digests
by agarosegel chromatography
sedi-mentedthroughsucrose
gradients
inabroadbandon November 11, 2019 by guest
http://jvi.asm.org/
[image:5.493.44.236.76.243.2]CHICK EMBRYO
-Tm =86°C
40 50 60 70 80 90 100 40 50 60 70 80 90 100
[image:6.493.63.455.58.310.2]T E M PER ATUR E C
FIG. 1. Meltinigprofiles of niuclease-resistanit RNAantd virus interference iniducerpurified from ntucleic acid
digests byexclusionchromatographyoni6%agarose. Samples ofRNA wereheatedattheindicatedtemperatures anidassayedfortrichloroaceticacid-precipitable radioactivity afterthe standardribonucleasetreatment.ForpartC, samples of niuclease-resistanit HeLa cell RNA were also analyzed for interference inducer. After thermal de-niaturation andtreatment with ribon2uclease unider thestandardconditions, theRNA wasphenolextractedthree
timesbeforeprecipitationi with carrier sRNAbyadditionoftwovolumesofethanol. TheRNAwasthenidissolved
directly inthe mediumu*sedjbrviruisiniterferenceassayswithrabbitkidnzeycellcultures. with a major peak occurring at about 7S for
rabbitkidney, 9Sforchickenembryo,and11S for HeLa I cells whencomparedwith4S yeast sRNA marker(Fig. 3).Sedimentation coefficientsranged from6to8S forrabbit cellribonuclease-resistant
RNA (seven gradients of fourindependent
prep-arations), 7 to 9S for chicken cell RNA (six gradients offour preparations), and from 10 to 11S for HeLa I cell ribonuclease-resistant RNA
(four gradients oftwopreparations). Foragiven preparation, little change in sedimentation be-havior of the ribonuclease-resistant 3H-RNAwas
observed after the standard nuclease treatments,
althoughstillmoreribonucleasedigestionreduced
the averageS valuebymorethan 15 S (datanot shown). Similar results were reported for de-pendenceofsedimentationon ribonuclease treat-ment in the case of MS2 double-stranded RNA
(2).
The lowest sedimentation coefficient obtained inourexperiments after the standard deoxyribo-nuclease plus ribonuclease treatment of purified ribonuclease-resistant RNA ispresentedforeach cell species in Fig. 4. A shoulder (Fig. 3B) or
distinct secondcomponent (Fig.4B)with alower
sedimentationcoefficient than the mainpeakwas
observed in most preparations of chicken cell
RNA. We did not examine the possibility that
the physiological state of the cells in culture
determines the sedimentation characteristics of
different isolates of cellular double-stranded RNA. Nor was it shown whether the variations
reflect differences in the original size or in the
ribonuclease sensitivityofthebase-paired portion
oftheundigestedmolecules. In allcases,however, it wasfoundthat the purified ribonuclease-resis-tant RNA which induced virus interference was
distributed insucrosegradientsfromover7Sto at least20S. Most of the active fractions (15to40% of thetotal) weresedimented faster than the main peaksofcellularnuclease-resistant 3H-RNA (Fig. 4).
Usingthe formula of Franklin for the
molecu-lar weight of double-stranded RNA (molecular weight = 2.42S; 16; reference 13), one can
esti-mate average molecular weights for cellular double-strandedRNA of 5 X 104daltons for 7S rabbit kidney cell RNA,2 X 105for9S chicken cellRNA,and 6 X 105 for11S HeLa I RNA. The
interference-inducing double-stranded RNA, with
sedimentation coefficients from 8 to 20S, canbe estimatedtopossess aminimum molecularweight
of 105daltons andamaximum of several millions.
A minimum molecular weight of 1.5 X 105 to
B c
HHE LAI Tm =86°C
*
0I
0!
l
E
UL
3.0
--z
0 0 2.0 a 0
U
cr-1.0 e)
o
0
0 0.0
Ij
r-L-i
L
on November 11, 2019 by guest
http://jvi.asm.org/
RNA
703
1.8
1.7
1.6
1.5 1.4 1.3
1.8
-1.7 -1.6
-1.5
-1.4
0
E
"I
en z 0 z
0
0
()
JI.3
1.5
1.0
0
0
cD
cx
I
5.0 0Z
H
4.0 2
0
3.0 n 2.0 f 1.0 ci
0.0
>
-0.1 £
0
0
-J 1.5
1.0
0.5
0
FRACTION NUMBER
FIG. 2. Density distributionz of cellular nuclease-resistant 3H-RNA anid virus initerferenice in2ducerin Cs2SO4
gradienztsbefore antdafterthermal deniaturation. ForpartsA, B,andC, samples ofinticlease-resistantRNAfractions
wereanalyzed afterafurther stantdardnucleasetreatment,followed bythreepheniolextractions anidethaniol
pre-cipitationiinlthepresence of onteA260 unit eachoftobaccomosaicvirusRNAandsalmoniDNA. ForD, E,andF, sampleswereheated inlow-saltbufferat100 Cfor3miti,quenchedinmeltinigice,andprecipitatedwithmarker
niucleicacidsasabove. Eachsample wasthendissolved in 3.8 ml ofTris-hydrochloride buffer (0.05 mi,pH 7.4) containinlgsuifficient Cs2S04togiveafinalaveragedensity ofabout 1.58g/ml. Centrifugation wasat20 Cantd
31,500rev/mi,zfor3daysinaSpincoSW 50.1 rotor.Gradientswerefractioniated bycollecting dropsfroma
pin-hole in the bottomof the centrifuge tubes.Samples of gradient fractionswere takentup immediatelyin0.100-ml
taredmicropipettes whichwereweighedtomeasuresolutiondensity. Toeachfractioni anequal volume ofwater wasaddedbefore readingA2M0,andsampleswereassayedeitherfortrichloroacetic acid-insolubleradioactivityor,
afterethantolprecipitation, for initerference inlducerinrabbit cells. The latterassays were nlotperformed forparts D, E, anid F becausemelting destroys the inducer. Theslight interference inductioniobservedunderthe tobacco mosaic virus RNAmarkerforthegradientinpart Bprobablyindicates nuclease-resistantRNAco-precipitating
withsinigle-strandedRNA duetoincompleteremovalofsingle-stranded regions fromthenuclease-resistantcoreof
theRNAcomplex.
1.6 X 105 daltons for interferon induction has been determined similarly for the synthetic polymer duplexrI:rC (12).
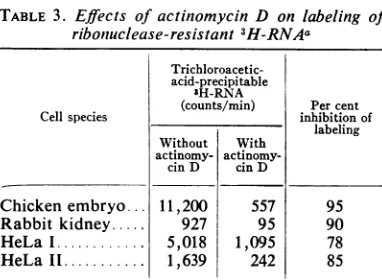
Effects of actinomycin D onthe biosynthesisof cellular double-stranded RNA. Incorporation of 3H-uridine into the double-stranded RNA frac-tion of cellular nucleic acids was inhibited by actinomycin D, as reported previously (24). In-hibitationwas 90%c ormoreinthetwospeciesof cellsgrowninprimary cultures, whereas the
syn-thesis of ribonuclease-resistant 3H-RNA in the
two HeLa cell lines was reduced by 78 to 85% (Table 3).Totalincorporationof3H-uridine into cellular RNA was inhibitedby over 98% under
thesameconditions(datanotshown).The inhibi-tion of incorporation into cellular double-stranded RNAbyactinomycinDsuggeststhatits
synthesismaybedependentonDNAtemplate. DISCUSSION
Experiments described in this paper indicate
that animal cellscontain afraction of RNA that
0
E
I
on November 11, 2019 by guest
http://jvi.asm.org/
[image:7.493.45.442.58.384.2]500 II I
A 7S 4S B C iis 4S
RABBIT CHICK EMBRYO HELA I
KIDNEY 400
~~~~~~9545
_ 300 3.0
E
0.
0
4~~~~~~~~~~~~~~~~~~~~-.
-200
2.00
100 1.0
0 5 10 15 20 0 5 10 15 20 0 5 10 15 20
FRACTION NUMBER
FIG. 3. Sedimentationanalysis of purified cellular ribonuclease-resistant RNA. Samples were precipitated wit/l twovolumesof ethanol in the presence of 50
lug
of 4S yeast sRNA marker. Pellets were then dissolved in 0.15 ml of buffer containing0.01 M Tris-hydrochloride (pH 7.4), 0.1 m LiCl, Imm EDTA, and0.1%1,
sodium dodecyl sulfate and layered on 5-mllineargradienitsof sucrose (10 to25%) in the samebuffer.A0.3-ml cushion of65% sucrose in D20 was included at the bottom of each gradient. Centrifugation was for 315 min at 20 C and 65,000 rev/min in a SpincoSW 65rotor.Gradients were fractionated as in Fig.2and weresimilarly assayedforA260 and trichloroaceticacid-precipitable radioactivity.A,I II,
RABBIT KIDNEY dsRNA CHICK EMBRYO dSRNA HELA I dsRNA
75 4S 7S 4S 7S 4S
700 -SS7
600 5.
1
I A X0oJo
0.5 -4.0
400
10015
20 0 5 10 15451 2 -3.0300
-~~~~~~~~~~~~O3 2.0wu
0
] H
I0.2 1.0 U
100
~~~~~~~~~~~~~~~~~~0.1
-0.0
>0
0 5 10 15 20 0 5 10 15 20 0 5 10 IS 20
-Ol
FR ACT I ON NU M BE R
FIG.4. Sedimentation distribution of interference-inducing activityandradioactivityafternuclease treatment of cellular double-strandedRNAlabeled with 3H-uridine. Aftertheribonuclease-resistant RNAfractionwasgiven the standard ribonucleaseplus deoxyribonuclease treatment, it wasphenol-extractedandprecipitatedwithethanol. Velocity sedimentationanalysison sucrosegradientsisdescribedinFig.3. In thiscase,afterA260readings, partof eachfractionwasassayedfor trichloroacetic acid-insolubleradioactivity, and the remainderwasethanol-precipitated forvirusinterferenceassayswith rabbitkidneycultures.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:8.493.62.461.70.321.2] [image:8.493.68.454.400.595.2]TABLE 3. Effects ofactinomycin D on labeling of
riboniuclease-resistant sH-RNAa
Trichloroacetic-acid-precipitable
3H-RNA
(counts/min) Per cent
Cell species inhibition of
labeling Without With
actinomy-cinD cin D
Chicken
embryo
11,200
557 95Rabbit
kidney
927 95 90HeLa. 5,018 1,095 78
HeLaII... 1,639 242 85
aNuclease-resistant fraction of cellular
3H-nucleic acids was prepared from parallel cultures with and without actinomycin D.
is
double-stranded
and induces interference withvirus
replication.
In this study, it has not beendemonstrated
directly
that interference with thegrowth
ofSindbis
virus or VSV by cellularribo-nuclease-resistant
RNA ismediated by interferon.Butexperiments carried out under similar
condi-tions
inthis
(5, 6)
andother
(12,
13)laboratories
have
suggested
thatinduction
of interferon bydouble-stranded
RNAis the
basis of suchinter-ference.
Cellular double-stranded RNA
andinterfer-ence-inducing
activity purified from cellular
nucleic
acids sharethese
properties: they are bothresistant
toribonuclease and
deoxyribonuclease,they
havethe
samedistinctive
solubilities
insalt
solutions
ofdifferent concentrations,
and the Tmvalues
fortheir
thermal denaturation
areco-ordinately affected by changes in ionic
environ-ment.
Also
they have the same buoyantdensity inCs2SO4.
However,the
distributions
of
ribonu-clease-resistant interference-inducing activity
andRNA were
only partially coincident after
frac-tionation
by
velocity
sedimentation: inducing
RNA
sedimented
faster
than the average-sizedmolecule of cellular double-stranded
RNA.Therefore,
it
may beconcluded
that thehigh-molecular-weight double-stranded
RNA incells
is
the mostefficient
component ofcellular
nucleicacids reported
to induceinterferon.Earlier
reports onribonuclease-resistant
cellu-lar
RNAdid
not showinduction
ofvirusinter-ference(6,
24),
butpresumably
thenegative result(6)
was due to lowersensitivity of
theassay sys-tem whichemployed cultures
100 timeslarger
than those
used
here. Theobservation
inthis workthat
total cellular nucleic
acids had nointerfer-ence-inducing
action but were rendered potentafter
deoxyribonuclease
treatment cannot beex-plained
from ourexperiments.
This work shows that
cells of
agiven animal
produce double-stranded RNA which induces
virus interference
inother cells of the sameani-mal, as well as in those of another species of animal. This evidence indicates that double-strandedness may be the onlyessential property
for interferon
induction bynatural
RNA, ratherthan the source of the RNA as postulated pre-viously
(18).
Such aconclusion raises the question of whyresident cellular double-stranded RNA, asit
exists in the cell, does not induce detectable interference with virus growth. Perhaps thein-tracellular concentration
ofdouble-stranded
RNAis
too low for interferon induction. Fromlong-term
labeling experiments
with32P-phosphate, it
ispossible
toestimate
that of thetotal
RNA(10-
jig
per
chicken cell) only about
0.01% (~-10" Mg)
isdouble-stranded
(ourunpublished
data),
inagree-ment
with
immunochemical
measurements(28).
The minimumlevel of poly
rI
:rC forinterference induction was found to be 10-8Mig
per chickencell
(5). Assuming
thatpoly
rI:rCand
cellular
dou-ble-stranded
RNA havesimilar specific
activities
of
interferon
induction,then it may beestimated
that about
10times
moreextracellular
double-stranded
RNA thanis
present in thecell
isnecessary for induction of virus growth
interfer-ence.
It
is
possible, however,
thatenough
double-stranded RNA is present in the
cell
to causeinter-ference
with viruses, but the cellular
double-stranded
RNA insitu
may notbe able
tointeract
with
components necessary forinterferon
induc-tion.
It isalso
possible
that in thecell less
RNAexists in the
double-stranded
form beforede-proteinization
which occursduring isolation.
The
biological
role
andorigin of the cellular
double-stranded
RNA arestill
open tospecula-tion: this
RNAmay begenerated by
alatent virusinfection
orit
could be
atruecellular
component.The
general
occurrence
of
double-stranded
RNAin different
primary cells (chicken,
rabbit, mouse)
and
cell lines (human, hamster;
comparethis
report,24,
28,
and ourunpublished observations)
argues
against
aviral
origin of this
RNA. More-over,the
finding
that thebiosynthesis
ofcellular
double-stranded
RNAisinhibited
by
actinomycin
Dfurther
dispels
thesuspicion
thatthis
RNAmay represent thereplicative
intermediate of
alatent
RNA
virus,
becauseviral
RNA-dependent
RNAsynthesis
isgenerally
known to be resistant toactinomycin
D. However, we cannotexclude
thepossibility
that the double-stranded RNAmight
derive from infection
by
aDNAviruswhich,
like vaccinia virus(6),
iscapable
ofsynthesizing
double-stranded
RNA.It seems more
likely,
however,
that theinter-ference-inducing,
double-stranded
RNAde-scribed
here istranscribed
fromcomplementary
7,
705on November 11, 2019 by guest
http://jvi.asm.org/
[image:9.493.46.237.75.215.2]regions ofcellular DNA. This idea is compatible with the ubiquity ofthis RNA speciesin animal
cellsandwiththesensitivity of itsbiosynthesisto
actinomycin D. It is also conceivable that some
cellular RNA-dependent RNA synthesis exists,
since we found that the biosynthesis of cellular double-stranded RNAappearsto be less sensitive toinhibition by actinomycinD than total RNA synthesis (alsomentioned in24).
Furthercharacterization of the cellular
double-strandedRNA will be necessary to speculate on
its biological role.
ACKNOWLEDGMENTS
We thank Harry Rubin forgenerouscooperation andG. S. Martin,M.Bissel,and Bart Sefton for useful criticism of the
manu-script.Furthermoreweappreciatethepleasant exchangeofresults
before publication with Edward De Maeyer, Jacqeuline De Maeyer-Guignard, and Luc Montagnier, who haveshown that
the double-strandedRNA from rat cellsinduces interferon
pro-duction incellsof thesamespecies.
This investigation was supported byPublic Health Service
grantCA 11426 fromthe National Cancer Institute andtraining
grantGM 01389 from the National Institute of General Medical
Sciences.
ADDENDUM IN PROOF
Results similar tothosereported byus have been
published byE. DeMaeyer,J.DeMaeyer-Guignard,
and L. Montagnier [Nature New Biology (London)
229:109-110, 1971].
LITERATURE CITED
1. Baltimore,D. 1966.Purification and properties of poliovirus
double-strandedribonucleicacid. J. Mol. Biol.18:421-428.
3. Billeter, M. A., C. Weissmann, and R. C. Warner. 1966. Replication of viral ribonucleic acid. IX. Properties of double-stranded RNAfromEscherichia coli infected with bacteriophageMS2.J. Mol. Biol.17:145-173.
3. Billiau, A., andE.Schonne. 1970. Inductionof the interferon mechanism by natural RNA.LifeSciences 9:69-78.
4. Colby,C. 1970. The induction ofinterferonby natural and
synthetic polynucleotides, p. 1-32. In W. Cohn and N.
Davidson (ed.), Progress in nucleic acid research. Aca-demic Press Inc.,NewYork.
5.Colby, C., and M. J. Chamberlin. 1969. The specificity of interferon induction in chick embryo cells by helical RNA. Proc. Nat. Acad. Sci. U.S.A. 63:160-167.
6.Colby, C.,and P. H.Duesberg.1969. Double-stranded RNA invaccinia virus infected cells. Nature (London) 222:940-947.
7. Colobert, L.,and P. Louisot. 1968. Inhibition of the
multipli-cation ofmyxovirusand arbovirusby chemicallymodified
ribonucleic acids from the host cells. Biochem. Biophys.
Res. Commun.30:148-152.
8. Duesberg, P. H. 1969. Distinct subunitsof the ribonucleo-proteinof influenzavirus. J. Mol. Biol. 42:485-499. 9. Duesberg,P.H., and E. Canaani. 1970. Complementarity
be-tweenRoussarcoma virus (RSV) RNA andthe in vitro-synthesizedDNAof thevirus-associated DNApolymerase. Virology 42:783-788.
10. Duesberg, P. H., and W.S.Robinson. 1967.On thestructure
andreplication of influenza virus. J. Mol. Biol. 25:383-405.
11. Field, A. K., A. A. Tytell, G. P. Lampson, and M. R.
Hilleman. 1967. Inducers of interferon and host resistance. II. Multistranded synthetic polynucleotide complexes. Proc. Nat. Acad. Sci. U.S.A. 58:1004-1010.
12. Field, A. K., A.A.Tytell,G. P.Lampson, M. M. Nemes, and M. R. Hilleman. 1970. Double-stranded polynucleo-tidesasinterferoninducers.J. Gen.Physiol.56:90S-96S. 13. Franklin, R. M. 1967. Replication of bacteriophage
ribo-nucleicacid:somephysical properties of single-stranded, double-stranded, and branched viral ribonucleic acid. J. Virol. 1:64-75.
14. Geiduschek, P. E., J. W. Moohr, andS. B. Weiss. 1962. The secondarystructure ofcomplementary RNA. Proc. Nat. Acad. Sci. U.S.A. 48:1078-1086.
15. Hilleman, M. R. 1968. Interferon induction and utilization.
J.Cell.Physiol. 71:43-60.
16. Ho, M. 1966. The production of interferons, p. 21-52. In N. B.Finter (ed.), Interferons. North-Holland Publishing Co., Amsterdam.
17. Ho, M., K. H. Fantes, D. C. Burke, and N. B. Finter. 1966. Interferons or interferon-like inhibitorsinduced by
non-viral substances, p. 181-201. In N. B. Finter (ed.), Inter-ferons.North-HollandPublishing Co., Amsterdam. 18. Isaacs, A. 1961. Mechanisms of virus infections. Nature
(London) 192:1274.
19. Isaacs,A. 1965. Studies on interferon. Aust. J. Exp. Biol.
Med. Sci. 43:405-412.
20. Isaacs, A.,R. A.Cox, andZ.Rotem.1963. Foreign nucleic
acidsasthe stimulus tomakeinterferon. Lancet2:113-116.
21. Jensen, K. E.,A. L.Neal, R. E. Owens, and J. Warren. 1963. Interferonresponsesof chickembryo fibroblaststonucleic
acids and relatedcompounds.Nature (London)
200:433-434.
22. Kohlhage, H., and D. Falke. 1964. Vermehrungshemmung des herpes-simplex-virus durch ribonukleinsaiuren. Arch.
Gesamte Virusforsch.14:404-409.
23. Louisot, P., L. Colobert,C. Bollack,J. H. Weil, and J. P. Ebel. 1968. Inhibition dela multiplication viraleal'aide
d'acides ribonucleiques chimiquement modifies.I.
Inhibi-tion de la multiplication de myxo',irusparainifluienzae I (virus sendai) surcellules de rein deveau.Biochim. Bio-phys. Acta 155:38-50.
24. Montagnier, L. M. 1968.Presence d'un acideribonucleique
endouble chaine dan descellulesanimales. C. R. Acad.
Sci.Paris 267:1417-1420.
25. Nygaard,A.P., andB.D. Hall.1964. Formation and
proper-ties of RNA-DNAcomplexes.J.Mol. Biol. 9:125-142. 26. Rein, A.,and H. Rubin. 1968.Effects of localcell
concentra-tionsuponthegrowthof chickembryo cellsintissue
cul-ture.Exp. Cell Res. 49:666-678.
27. Rotem, Z.,R. A.Cox, andA.Isaacs.1963.Inhibition ofvirus multiplication by foreign nucleic acid. Nature (London) 197:564-566.
28. Stollar, V., andB. D.Stollar. 1970.Immunochemical
meas-urementof double-stranded RNA of uninfectedand arbo-virus-infected mammalian cells. Proc. Nat. Acad Sci.
U.S.A. 65:993-1000.
29. Szybalski, W. 1968. Use of cesium sulfate forequilibrium density gradient centrifugation,p.330-360.InL. Grossman
andK.Moldane (ed.), Methodsinenzymology, vol. 12B. Academic Press Inc., New York.
30.Takano, K.,J.Warren,K. E.Jensen,and A. L.Neal. 1965.
Nucleic acid-induced resistance to viral infection. J.
Bac-teriol. 90:1542-1547.
31. Vilcek, J.,M. H.Ng,A.E.Friedman-Kien, andT.Krawciw.
1968. Induction of intetferonsynthesis by synthetic
double-strandedpolynucleotides.J. Virol. 2:648-650.
32.Yankofsky,S. A.,and S.Spiegelman.1962. Theidentification
ofthe ribosomal RNA cistron by sequence
complemen-tarity. I. Specificity of complex formation. Proc. Nat.
Acad. Sci.U.S.A. 48:1069-1978.