Vol. 27, No. 3 JOURNALOFVIROLOGY, Sept. 1978,p.490-504
0022-538X/78/0027-0490$02.00/0
Copyright i 1978 American Society forMicrobiology Printed in U.S.A.
Collaborative
Complementation
Study
of
Temperature-Sensitive
Mutants of
Herpes Simplex Virus
Types
1
and
2
PRISCILLA A. SCHAFFER,* V. CELESTE CARTER, ANDMORAG C. TIMBURY't Divisionof BasicSciences, Sidney Farber Cancer Institute, Harvard Medical School, Boston, Massachusetts02115,andDepartment of Virology, Baylor College of Medicine, Houston, Texas 77030; and
Instituteof Virology, University of Glasgow, Glasgow G115JR,Scotland'
Received for publication 8 March1978
Twenty-three complementationgroupsof herpes simplex virustype 1(HSV-1)
and 20 of HSV-2wereidentifiedby qualitative and quantitativecomplementation
analysis from among43temperature-sensitive (ts) mutantsof HSV-1 and29ts
mutantsof HSV-2 which hadbeen isolated independently in 10laboratories.
The herpes simplex virus (HSV) genome is
large (100 x 106) and complex (5, 25, 31) and
probably encodes from 70 to80 genes. A
com-plete understanding of the structural and
func-tional organization ofthe genome will
necessi-tate theidentificationofall viral genes and their
ordering on the viral genetic map.
To date, most genetic studies of HSV have
utilized temperature-sensitive (ts) conditional
lethal mutants whichcanoccur,theoretically,in
all essential genes. Although the total number
ofessentialgenes encodedbyHSV DNA is not
known,complementation analysis of one series
of ts mutants has ledtothe identification of 15
cistrons of HSV type 1 (HSV-1) (22) and, in a
collaborative study, 18 cistrons ofHSV-2 (30).
Because it is very likely that more than this
numberofessentialcistrons is encoded bythe
HSV genome(3), a collaborative
complementa-tionstudywasundertaken toidentify additional essential cistrons from amongexistingseries of
HSV-1 and HSV-2tsmutants.
Theresultsof thisstudyestablish theidentity
of 23 cistrons among 43 ts mutants of HSV-1
and20cistronsamong29 ts mutantsofHSV-2
and demonstrate thatmostpreviously untested
t In collaboration with: S. M. Brown andJ. H. Subak-Sharpe,Institute ofVirology, University of Glasgow, Glasgow G115JR, Scotland;A.Buchan, Department of Virology, The MedicalSchool, Birmingham B15 2TJ, England; I. Hallibur-ton,Department of Microbiology, School of Medicine, Leeds LS29NL, England;R.Hughes and W. Munyon, Department ofMedical ViralOncology, Roswell Park Memorial Institute, Buffalo,NY14263; R. W. Koment and F.Rapp, Department ofMicrobiology,Milton S.Hershey Medical Center, Pennsyl-vania StateUniversity, Hershey, PA 17033;R. Manservigi, Instituto diMicrobiologia, Universita di Ferrara, 44100 Fer-rara,Italy; M. Takahashi,Research Institute for Microbial Diseases,OsakaUniversity, Yamada-Kami, Suita City, Osaka, Japan;V.Vonka,Institute of Sera andVaccines, Department ofExperimentalVirology, Praha 10, W. Pieck 108, Czechoslo-vakia; N. Zygraich and C. Huygelen, Biologics Department, RechercheetIndustrieTherapeutiques,Rixensart,Belgium.
mutants belong toalready established
comple-mentationgroups. It isclearthat the number of new complementationgroups identified in this
studyissmaller than onewould expect given the
number of independently derived mutants
tested. These findings demonstrate the
neces-sity, therefore,for(i) developingnewprocedures
for the isolation of mutants in as yet unidentified
HSV cistrons and(ii) examiningthefunctional
basis for the extensive failure to complement
observed with certain mutants.
MATERIALS AND METHODS
Celisand cellculture.Monolayercultures of
se-riallypropagatedhumanembryonic lung (HEL)
fibro-blastswere used for the preparation of virus stocks and for mixedinfectionsinthe quantitative
comple-mentation test. Vero cell monolayers were used for virusassayand for mixed infections in thequalitative complementationtest.Both HEL and Verocellswere grownat37°CinEaglemedium(Autopow,Flow
Lab-oratories,Rockville,Md.) supplementedwith 10% fetal bovine serum and containing 0.075% NaHCO3 (for
cultures in closed vessels) or 0.225% NaHCO3 (for culturesin openvesselsina5%CO2atmosphere).
Virusesand virus assays.Propertiesof theseven
seriesof HSV-1 and five series of HSV-2ts mutants
used in thisstudyare shown inTable 1. Wild-type
virus strainsand the mutagens used for the induction ofts mutants, aswellastheviralDNAphenotypesof
the mutants, are shown. Each mutant from agiven seriesincluded in thestudyhad been shownpreviously
torepresent anuniquecomplementationgroup in that series.Onlyonememberof each groupwastested.
Virusassays wereperfornedin Verocells byusing
a 2% methylcellulose overlay (7) in CO2 incubators
(Wedco,Silver Spring, Md.)with temperature varia-tionsof+0.20C.
Preparationof virus stocks. Stocks oftsmutants
werepreparedbyinfectingconfluent HELcell
mono-layersin100-mmpetriplateswith1 mlof virus
inoc-ulum at multiplicities of0.01 to 0.1 PFU/cell. After
adsorptionat37°Cfor1h,the inoculumwasdecanted, monolayerswerewashedoncewithTris bufferatpH
490
on November 10, 2019 by guest
http://jvi.asm.org/
27,
Virus type
HSV-1
TABLE 1. Selectedproperties oftheHSV-1and HSV-2 tsmutantsused in thisstudy
Viral
Wild-type strain Mutagen' Mutant pheno- Referenceorinvestigator
typeb
KOS BUdR Al - 1,21
BUdR B2
-BUdR C4
-BUdR D9
-BUdR E6 +
NTG F18 +
BUdR G3 ±
BUdR Ill +
NTG J12 +
NTG K13 ±
NTG L14 ±
UV M19 ±
UV N20 +
UV 022 ±
AP P23 - J.Jofre, C.-T. Chu, and P. A. Schaffer,
17 BUdR
BUdR BUdR BUdR BUdR
KOS 1.1 BUdR BUdR BUdR BUdR BUdR BUdR BUdR
HFEM BUdR
BUdR BUdR NA
HFEM Spontaneous
BUdR BUdR
Spontaneous
BUdR BUdR BUdR
KOSpp60l BUdR BUdR
13 BUdR
BUdR BUdR
HSV-2 186 BUdR
BUdR BUdR BUdR BUdR BUdR BUdR UV
A +
D _
F +
G +
I +
84 478 656 661 901 822 833
Bl B5 B7 N103
LS1 LB1
LB2
LS2
LB3 LB4
LB5
3
7
D10
C4
G5
A8
B5
C2 D6 E7 F3 G4 H9
manuscript in preparation
3
14
A.Buchan +
11
NDd
ND
V.Vonka
17
+
9,20
± + + + +
4
on November 10, 2019 by guest
http://jvi.asm.org/
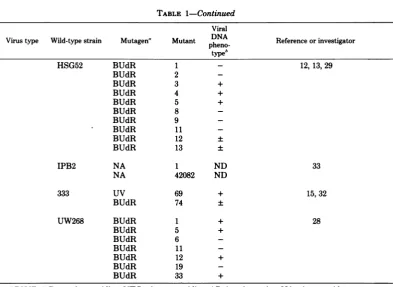
[image:2.501.53.453.83.666.2]TABLE 1-Continued
Viral
Virus type Wild-typestrain Mutagen" Mutant pDNoA Reference or investigator typeh
HSG52 BUdR 1 - 12, 13, 29
BUdR 2
-BUdR 3 +
BUdR 4 +
BUdR 5 +
BUdR 8
-BUdR 9
-BUdR 11
-BUdR 12 ±
BUdR 13 ±
IPB2 NA 1 ND 33
NA 42082 ND
333 UV 69 + 15, 32
BUdR 74 ±
UW268 BUdR 1 + 28
BUdR 5 +
BUdR 6
-BUdR 11
-BUdR 12 +
BUdR 19
-BUdR 33 +
aBUdR,5-Bromodeoxyuridine; NTG, nitrosoguanidine; AP,2-aminopurine; NA, nitrous acid.
'The viral DNA phenotype of ts mutants is based upon their ability to synthesize viral DNA at the
nonpermissivetemperature comparedwiththeability ofthe wild-type virustosynthesize viral DNA at this temperatureas100%.+,>20%;±,520%;-,nodetectableviralDNAsynthesized.
'Viral DNAphenotypesof strainKOS1.1ts mutants werekindly supplied by Myron Levine, Universityof
Michigan,AnnArbor,Mich. dND, Not done.
7.4, and 7 ml ofEagle medium containing 5% fetal bovineserumand0.225%NaHCO:)wasaddedtoeach
plate.Infected cellswereincubatedat34°Cuntil
cy-topathic effects were generalized. Cells were then
scraped into the medium, and replicate suspensions
werepooled. Thesuspensionwassonically disrupted
for 45s at10kc andcentrifugedat180xg,5°Cfor10
min. The supernatant fluidwasdispensed, frozen at
-90°C,andassayedatpermissive (34°C)and nonper-missive (38°CforHSV-2 and39°Cfor HSV-1ts
mu-tants) temperatures. Virus preparations containing
significant levels of ts+ revertants were
plaque-puri-fied, and stocks were prepared from revertant-free plaques. Allvirusstocks used in thisstudy exhibited efficiencies ofplating [EOP; (PFU permilliliter,
as-sayedatnonpermissive temperature)/(PFU per mil-liliter, assayedatpermissive temperature)] of less than
orequalto10-4. Mutants withconsistentlyhigh levels ofleak and reversion (efficiency ofplating > 10-4)
werenotincluded in these studies.
Complementation tests. Two types of
comple-mentation tests were employed in this study. The
qualitativetest(4)wasused for initialscreening pur-poses, and thequantitative test (22)wasusedasthe
moredefinitive test.Thesetwo testsdifferprimarily
inthe timepermittedfor theexchangeof gene
prod-uctsandforvirusreplication duringmixed infection. Thequantitativetestinvolvesasingleround of virus
replication,whereasthequalitativetestinvolves
mul-tiple rounds ofreplication. Correlation of results ob-tained in the two types oftests hasgenerally been verygood; however, falsenegative and false positive
qualitativeresults have been described (4,30).
(i)Qualitative complementationtests.
Qualita-tivetestswereconductedasdescribedpreviously (4). Briefly,Verocellswereinfected withtwomutantsby placing filterpaperdisks saturated with mixtures
con-taining 2 x 106, 4 x 105 and 8 x 104 PFU ofeach. mutant per ml on preformed monolayers in 60-mm
petri dishes. Control disksweresaturated with suspen-sions of individual viruses at these dilutions. After incubation at the nonpermissive temperature for 5
days under 2%methylcellulose,neutral redwasadded
toeachplate.Clearingof themonolayersin theareas
ofmixed infections was comparedwith the clearing producedinareasinoculated with eachmutantalone. Ifmoremarked clearing occurred inareasof mixed infections than in areas infected with each mutant
alone,complementationwassaidtohave occurred.
(ii) Quantitative complementation tests. The
quantitative oryield-of-progeny virus test compares theyieldof virus obtained fromcells infected withtwo
492 SCHAFFER ET AL. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
tsmutantsand incubatedatthe nonpermissive tem-perature with theyield obtained from cells infected
with each mutantalone and incubated at this temper-ature. The test was carried outas previously described (22). Tubecultures of HEL cells were inoculated at a
multiplicityof 2.5PFU of each virus per cell (a com-bined multiplicity of 5), and singly infected control cultures were inoculated with 5 PFU of each virus alone per cell. Infectedcultureswereincubated for 18 h at the nonpermissive temperature in water baths
(Blue M, Rock Island, Ill.)with temperature variations of ±0.1°C, and virusyieldswere titrated in Verocell monolayers. Complementation indices (CI) were cal-culated from the formula: CI= ([A +B]s9c)/([A]is,c
+ [B]}.(-), where A and B are two mutants, and infected cells were incubated at the nonpermissive temperature.Virus yields were assayed at the permis-sivetemperature. As inprevioustests(22), a value of 2orgreater was taken to indicate positive complemen-tation.
Inthe present study each mutant pair was tested
bybothprocedures.If thequalitativetest was clearly
positiveand thequantitativetestyieldeda CI of 10 or greater, no additional tests were performed. If one typeoftestindicatedcomplementationand the other did not, both tests were repeated at least once. Unless otherwise stated, the result of the quantitative test wasconsidered to be definitive. Quantitative tests of mutant pairs which failed to complement in initial tests wererepeated three times.
RESULTS
Quantitative complementationtestswith
ts mutants of HSV-1. (i) Establishment of the standard set of HSV-1 ts mutants.
Rather than undertaking the analysis of all
HSV-1 ts mutants together, complementation
tests werefirstperformed betweenmutants
de-rived from strainsKOSand 17toobtain a
stan-dard set of mutants with which to compare
mutants isolated independently from other
strains. Five ts mutants of strain 17 and 15 ts
mutantsof strainKOSweretested in all possible
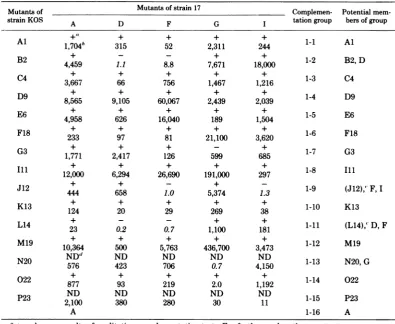
pairwisecombinations. The results of thesetests arepresentedinTable2.MutantstsD, F,
G,
andIof strain17failedtocomplementmutantsinat
least one of the strain KOS
complementation
groups. Mutants tsJ12 and L14 of strain KOS
and mutants tsD and F of strain 17 failed to complement mutants in two different
comple-mentation groups.These data suggest thattwo
or moreof thesemutants aredoublemutantsor
that thecomplementation between strain 17 ts mutants Fand I and D and F is intracistronic.
The factor which argues moststrongly against
intracistronic complementation is that if this
were the case, F, I, and D (or three of five
mutants of strain 17) would have to contain
mutations in the same gene. This situation
wouldrequireextremelystrong selectional
arti-fact andis,therefore,unlikely.Moreplausibleis
the possibility thattsJ12 and tsL14 are double
mutants.Characteristic ofapoint mutation,the
reversion frequency oftsJ12(10-4) issimilar to that of other mutants in the series, and this
mutant can be rescued by a single restriction
fragment inmarker rescueexperiments. In
con-trast, tsL14 reverts withlow frequency (<10-6)
andcannotbe rescuedbyany onefragmentin a
series of fragments representing the entire
ge-nome(unpublished data).
Mutant tsAof strain 17complemented all 15
ts mutantsofstrain KOS and wasassigned to a
separatecomplementation group.
To examine theuniformity of
complementa-tion patternsobtainedwhenmorethanone
rep-resentative of a groupwastested, two additional
mutants of the KOS series, tsA24 and tsF25,
werealsotested.tsA24and tsF25yielded results
identicaltothose of tsAl andtsF18,respectively,
in that they complemented the representative
mutantsof all five complementation groups of
strain 17efficiently (datanotshown).
The standard set of HSV-1 ts mutants was
thuscomposedofmutantsrepresenting 16
com-plementationgroups.
(ii) Complementation between mutants in the standard set and mutants in other series. Three series of ts mutants have been
derived from the KOS strain of HSV-1by
Schaf-fer et al. (22), Hughes and Munyon (14), and
Vonka (unpublisheddata). The standard set of
HSV-1 mutantswas nexttested against ts
mu-tantsin the lattertwoseries. Theresults ofthese
tests areshown inTable3.
Although tsP23 had complemented all
mu-tantsof strain17 (Table2), itfailedto
comple-mentfourofthemutantsin thetwootherKOS
series. Because of itsunusualgeneticand
biolog-ical properties which will be discussed below,
tsP23wasplacedinaseparatecomplementation
group.
In addition to the multiple negative results
obtained withtsP23,it is evident thattsP23,ts7
and ts3 did not complement any mutant well.
Mutantts3could be adouble ortriplemutant
(1-3, 1-4, and 1-14); however, the consistently
low CI obtained with this mutant makes this
conclusiontenuousin the absence ofgeneticand
physical mapping data.
With theseexceptions, allbut two other mu-tantsfell into groupsrepresented bymutantsin
the standard set. Two were shown to represent
newgroups: ts478andts7.Onlyts661exhibited
anoverlappingpatternofcomplementationnot
involving tsP23. ts661isprobablyadouble
mu-tant because the two mutantsthat it doesnot
complementareseparated by additional cistrons
(23). ts901 is asingle mutant in the tsB2 (1-2)
on November 10, 2019 by guest
http://jvi.asm.org/
494 SCHAFFER ET AL.
TABLE 2. Complementation between ts mutants of HSV-1 strains KOS and 17
Mutants of Mutants of strain 17 Complemen- Potential
mem-strain KOS A D F G tationgroup bers ofgroup
Al + + + + 1-1 Al
1,704b 315 52 2,311 244
B2 4,459 1.1 8.8 7,671 18,000 1-2 B2, D
C4
~+
+ + + + 13 CC4 3,667 66 756 1,467 1,216 1-3 C4
D9
~+
+ + + + 14 DD9 8,565 9,105 60,067 2,439 2,039 1-4 D9
E6
~+
+ + + + 15 EE6 4,958 626 16,040 189 1,504 1-5 E6
F18
~
+ + + ++16
F8F18 233 97 81 21,100 3,620 1-6 F18
G3 1,771 2,417 126 599 685 1-7 G3
+ + + + + il
12,000 6,294 26,690 191,000 297 1-8
Ill
J12 + +
-+
1- J12,F444 658 1.0 5,374 1.3 1-9 (J12),' F, I
K13
~
+ + + + + 11 1K13 124 20 29 269 38 1-10 K13
L14 23 0.2 0.7 1,100 181 1-11 (L14),' D, F
M19
~
+ + + + + 11 1M19 10,364 500 5,763 436,700
3,473
1-12 M19N20
ND"
ND ND NDND113
N0G576 423 706 0.7 4,150 1-13 N20,G
022
~
+ + + + + 11 2022 877 93 219 2.0 1,192 1-14 022
P23
ND
ND ND ND ND 1-15 P232,100 380 280 30 11
A 1-16 A
+and-areresults ofqualitative complementationtests.For furtherexplanation,see text.
bNumbersareCIobtained in quantitative complementation tests. Pairs with indices of 10 or greater were not
retested. Pairs which failed tocomplement, yieldedCI of less than 10, oryielded disparate results in the two typesof test wererepeatedtwo tofive times. In these cases valuesrepresent the average ofalltests; results of
quantitativetests wereregardedasdefinitive.Numbers initalics indicatenegativeindices.
'Potentialdouble mutantsareshown inparentheses.
dND, Not done.
cistron. BecausetsL14 also containsadefectin
cistron 1-2, itwouldnotbeexpectedto
comple-mentts901.
Theresults ofcomplementationtests
compar-ing mutants in the standard set with mutants
derived from strains HFEM and13 areshown in
Table 4. Again, tsP23 yielded negative or low,
positive indices with nearly half the mutants
tested. Excluding results with tsP23, mutants
tsBl, B7, LB4, LB5, and D10 were shown to
constitute new groups, whereas all other
mu-tants wereassignedtoexisting groups. Overlap-pingpatterns ofcomplementationnotinvolving
tsP23wereevidentinthe case of tsLB2 andLS2.
Because tsL14 is probably a double mutant, tsLB2wasassignedtogroup 1-2withtsB2. The
failure of tsLS2 to complement two mutants
whichliesomedistanceapart on theKOS
link-age map (23) suggests that tsLS2 is a double
mutant. The mutation in tsLB3 is probably in
the F cistron of strain 17 (i.e., cistron 1-11),
otherwise it wouldnothavecomplemented tsB2.
Theoverlapping patterns of complementation
noted in Tables2, 3, and4 suggest thattsL14
and tsB2 shouldnothavecomplementedin
pre-vious studies (22). By present standards,
how-ever, they did complement, although
ineffi-ciently (CI =3.6). Such inefficient
complemen-tation in retrospect may have been
intracis-tronic.
(iii)Complementationtestsamong ts
mu-tantsof HSV-1 representing putativenew
complementation groups. Eight of the 43
HSV-1tsmutants werefoundtocomplementall
mutants (except tsP23) in the standard set of test mutants and were, therefore, assigned to
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
COMPLEMENTATION WITH ts MUTANTS OF HSV 495
TABLz 3. ComplementationbetweenHSV-1ts mutants in the standard set and other tsmutantsof strain KOSa
Mutants Mutantsofstrains:
Mutan Comple- Potential
mem-84 478
| . KOS1.1 KOSpp601 mentation1
ber
ofgroupset 84 478
1
6561
6611
901 822 833 3 7Al 27,000 5,294 1.1 9,600 343 17 42,727 39 13 1 Al65
B2 + + + + - + + + - 12 B,0
B2
1,210 3,111 622 653 1.9 30 18,235 25 121-2
B2,
901
-4
+ + + + - + - A 61,3,3C4
40 63 34,400 0.8 980 225 0.4 0.6 6.81-3
C4,(661),833,(3)
D9 + + + + + - - + + 14 D984 3
D9
|
1.9
113
34,800
4.2
564
100
13
0.6
17
4 D9,8,(3)-6
+ + + + + + + - - EE6
4.0 469 1,520 12 1,494 230 19,166 357.3
1-5
F18
+
+
+
+
+
+
+
+
+
1-6 F1846 465 661 2,333 376 72 14,285 32 21
G3
+ + + + + + + + 1-7 G338,000 28 2,890 424 183 45 4,181 4.2 3.0
I1l
+ + + + + + + + + 1-8 ill340,000 944 24,400 2,000 565 59 3,181 30 6.8
J12 + + + + + - + + + 1-9 (J12),822
13,000 1,062 17,600 1,273 240 0.6 15,000 14 6.4
K13 + + + + + + + + + 1-10 K13
786 344 76,000 154 238 51 1,791 30 4.0
L14 18 1,000 3,080 35 0.4 2.8 9,523 19 2.8 1-11 (L14),901
M19
+ + + + + + + + +1-12 M19
8,000
17,500 144,000
520
518
15
15,384
8.4
14
N20
+ + + + + + + + +1-13 N20
682
421
376
169
197
22
9,393
33
21
02 + + + + + + + + + 11 2,(6) 3
022
2144
1,289
1.3
352
34
435
1.2
8.2
1-14
022,1(61),(3)
P23
0.9
4.6
35
5.1
230
11
1.0
0.02
0.7
1-15 P23A
+ + + + + + + + +A
24,000
1,062
1,880 8,800
1,877
26
7,000
18
7.6
1-16
478 1-17 478
1
1
1 1
1
1
1
I
1 ~~~~7 1-18 7aSeefootnotes toTable2forexplanationofresults.
putative
newcomplementation
groups.To con- Mixed infectionsresulting
inequivocal
firmthe
uniqueness
oftheirfunctions, they
were and anomalous patterns ofquantitative
tested among themselves
by
thequantitative
complementation.
CI inpositive quantitative
complementation
test.Theresults of thesetests testsranged
from 2.0(e.g.,
Table 4, tsK13 +are shown in Table 5.
Complementation
wastsLB3)
to440,000(e.g.,
Table4, tsMl9+tsG5);
demonstratedto occur between mutantsin all 72% ofpositive indicesweregreaterthan50.Of
pairwise
combinations.Therefore,
eachof these the 123positive
indiceswithvalues of lessthanmutantswasshowntorepresenta new
comple-
50,nearly
halfwereproduced
inmixed infectionsmentation group. Mutantpairs ts7 + tsBl, ts7 with5 mutants (e.g.,tsP23, tsLB2, tsC4 [strain
+tsLB5,ts7+tsD10 andtsLB4 + tsLB5yielded 13],
ts3,
andts7).
Negativeindicesrangedfromlow,
butpositive
indices. Similar results were 0.02(e.g.,Table3,tsP23+ts3)
to1.9(e.g., Tableobtained inthree separatetestsofthesemutant 3, tsD9 +ts84).
pairs. The low indices obtained with ts7 are Althoughthe results of mostquantitative tests
consistent withpreviousfindings(Table 3). were unequivocal, certain mutants exhibited
Quantitative
complementationtestshave thus equivocaloranomalous behavior inquantitativeresulted in theidentificationof23complemen- tests. A careful survey of results
indicates
thattationgroups from among43ts mutantsofHSV- such behavior isaconsequence of the properties
1
(Tables
2 to5). ofindividualmutantsratherthan thepropertieson November 10, 2019 by guest
http://jvi.asm.org/
J. VIROL.
cq m
O-"Z N
q m I" LO CO 000 C I" Lo co
+ ^+ + N+ + + + O+ + + + +N +
+t- + P ++P- ++ + >+;+ -4+ t-+.+ 4+ +q++ 4 + 04 + Lq
+ ~ ~t~ ~~' + r+° +° + +C,,+ +_+ _+
+_+O+U+ + +°++ +U+ s|>+ +: + 8 + +
C+ + c ++ + +m+ + +< +N+° :+s+ O+N+
+0 + + +
'o
+ +CS+
+ +8
+_+ +uN+°+ Ci++~~~ ~N-+st- ult+ + N+ + +-5 + +t-4 +O + ul
m-
t;SXx c: _ ; X.9 9co
1.
I
i
.4.1
1
:a
e
I
1
11-1.
O
-S a
A
:3e
12
it
9:
-S
t
d
Po PA m
49
E-,
4)
r.
t
0d
IzLO
0
m
1.4 u
0
V-4
0
LO
5
9
u
2 cli
5
1. m
C9
m
0 1
2
-4
5
P-4
2
a
P-4
z
t--a m
2
m 0
m
1-4
m
C)1.6-ill
-10 &
-SI
ET AL. 496 SCHAFFER
.8
I
I
l
@.
;R
on November 10, 2019 by guest
http://jvi.asm.org/
of the testitself.
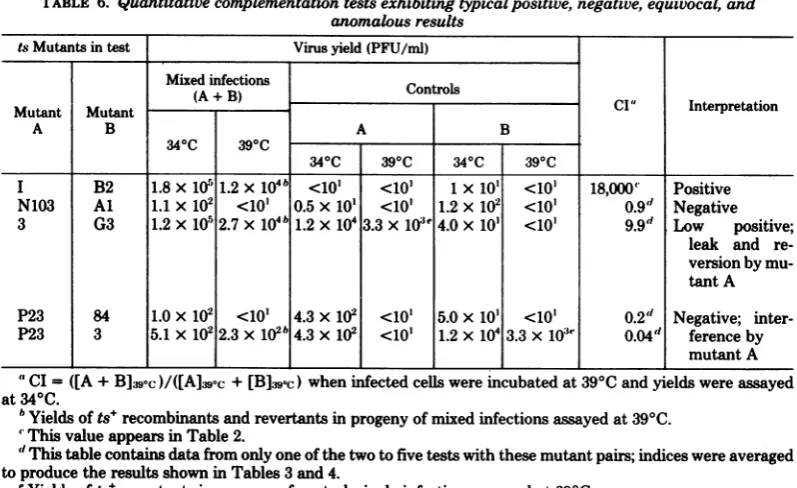
Toillustrate the anomalous behavior of these
mutants,the derivation ofCI for five
represent-E8
ative mutantpairs
is shown in Table 6. Datafroma
typical
positive
test areillustratedby
thepair
tsI
+ tsB2. In this test, the yield from themixedinfection was clearly greater than the sum
ofyieldsincontrol, single infections. Such
effi-cient complementation is in sharp contrast to
the typical negative test illustratedin Table 6
eo
No
eq eq eq bytsN103
+tsAl in whichtheyieldof the mixedinfection was
slightly
less than the sumof theyields
from control infections.ull<
Despite
the fact that all mutant stocks usedin these tests exhibited low levels of leak and
reversioninplaqueassays,severalmutants,
no-tably ts3, 7, LB2, and C4 (strain 13), yielded
higher levels ofleakand/orreversion than other
mutants afterincubation for 18 h at39°C which
oo complicated the interpretation of results. As S
a shown in Table6 (ts3+ tsG3),thebackground
levels of leak and reversion produced by ts3
Lo
mresulted
in a reduction in the CI in this test.Whether the index would have been higher if
leak were suppressed by incubation at higher
temperature
is notknown.
Itwas not
surprising,
therefore,
thatmutantswith thehighest
levels ofleak andreversionyielded consistently lower CI than mutants which did not leak or revertsignificantly. Consequently,
these mutantsyielded
agreater
number ofequivocal
andneg-C4
.lX
>ative
results.Anadditionalcauseof
equivocal
andnegative
e results in quantitative tests is illustrated in
Ta-ble 6 by pairs tsP23 + ts84 andtsP23 + ts3. In
these teststhe yieldsof mixed infectionswere
significantly lower than the sum of the yields of
controlinfections. InthecaseoftsP23+ts3, this reduction was nearly 1.5 logs. tsP23 also
sup-pressedthe
replication
ofts+ revertantsof ts3 inthis test. In contrast to other mutants which
consistently yielded low CI, tsP23 did not exhibit
unusually highlevelsofleak and/or reversion.
0 u i3 These data indicateastrong
tendency
by
tsP23to suppress the productionof ts virusthrough
<:-~complementation mechanisms and ts+
revert-ants andrecombinants. Theability of tsP23 to
suppress the growth of the wild-type virus by SE > >-= greater than 90% when equal
multiplicities
oftsP23 and wild-type virus are used to infect cells has been noted by J. Jofre (personal
communi-°X
XM4
cation). In a total of 28 quantitative tests in thepresent study,tsP23failedtocomplementeight
mutants and produced indices of less than 10
with four othermutants. Itis noteworthy that
five ofeight of the negative quantitative tests
.2 6^3 8 X z
Swere
positive
inqualitative
tests,whichinvolvemultiple rounds of replication.
Furthermore,
theon November 10, 2019 by guest
http://jvi.asm.org/
TABLE 5. Complementation tests with HSV-I ts mutants representing putative new complementation groups
CIfrommixed infections with ts mutant: Mutant
478 7 Bi B7 LB4 LB5 D1O
478 16 166 174 1,322 4,167 253
7 5.4 64 18 5.7 3.2
Bi 159 34,286 13,000 1,287
B7 62 30 47
LB4 2.6 86
LB5 210
[image:9.501.59.458.212.456.2]D10
TABLE 6.
Quantitative
complementationtestsexhibiting
typical positive, negative, equivocal, and anomalousresultstsMutantsin test Virusyield(PFU/ml)
Mixed infections Controls
(A+B) ci" Interpretation
Mutant Mutant
A B A B
34"C
390C 340C390C 34°C 390C
I B2 1.8x10' 1.2x 104b <101 <101 1x 101 <10o 18,000' Positive N103 Al 1.1 x 102 <10' 0.5 x101 <101 1.2 x102 <10' 0.9" Negative 3 G3 1.2xlOr1 2.7 x 104b1.2x1043.3x 10'e4.0 x10' <10' 99d Low positive;
leak and re-versionby
mu-tantA
P23 84 1.0X 102 <10' 4.3x 102 <10' 5.0x 10' <10' 0.2" Negative;
inter-P23 3 5.1x1022.3x 102b4.3 x 102 <10' 1.2x 1043.3x 1o3e 0.04d ference by
mutantA
"CI= ([A+
B]e,'c
)/([A]39,c
+[B]:j'sc)
wheninfected cellswereincubatedat390C
andyields
wereassayed
at34°C.
'Yields ofts+recombinants and revertants in progeny of mixed infections assayed at39°C.
'This value appears inTable2.
" This table contains datafromonlyone ofthe two to five tests with these mutant pairs; indices were averaged toproduce the resultsshown inTables3and 4.
'Yields ofts+revertants in progeny of control, single infections assayed at390C.
reasons for the efficient
complementation
ob-served between tsP23 and mutants
tsA,
D,
F(Table 2),901(Table 3),and B5 and LB4(Table 4),areunclear.
Ofequalsignificanceinthe
interpretation
of the results ofquantitative testsis theidentifi-cationofmutantswithoverlappingpatterns of
complementation, i.e., those with multiple ts
defectsasdescribed above.
Qualitative complementation tests with
tsmutantsof HSV-1.Althoughthe
quantita-tivetest wasconsideredtobe definitive in these
studies, excellent agreement was observed
be-tweentheresults ofqualitativeandquantitative complementation tests. With ts mutants of
strainsKOS and 17
(Table
2),nofalsepositive
qualitativetests(testsyielding positive
qualita-tive and negative
quantitative
results)
andtwofalsenegativetests(testsyielding negative
qual-itative and positive quantitative results) were
observed.
Intestsbetween mutants in the standard set
and other ts mutants ofHSV-1 (Tables3and 4),
agreement between the two types of tests was
alsogood,albeitlessso. Thirteenfalse negative
qualitative results (e.g., Table 3, tsC4 [strain
KOS] +ts84) and 12 false positive results (e.g.,
Table3, tsD9 +ts84)wereobtained. Of noteis
the observation that 5 of the 12 false
posi-tive results occurred in mixed infection with
tsP23. Thus, in atotal of433qualitative tests,
15 false negative (3.4%) and 12 false positive
tests(2.8%)wereobserved.
Quantitative complementationtestswith
ts mutants of HSV-2: the standard set of
HSV-2tsmutants.Inacollaborative
comple-mentation study involving 20 ts mutants of
HSV-2 strains HGS52 and 186,Timbury etal.
(30) identified18complementationgroups.
Rep-resentatives ofthese 18 groups were therefore
used in thisstudyasthestandardsetofHSV-2
tsmutants.
498
SCHAFFER ET AL. J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
Complementation between the standard
setand other HSV-2tsmutants.Theresults
of complementationtestsbetweentsmutantsin
the standard set and those derived from
wild-typestrainsIPB2, 333,andUW268areshownin
Table 7. Themoststrikingfeature oftestswith
HSV-2mutantswasthe fact that CI were
con-sistently 10-foldlower thanindices obtained in
tests with HSV-1 ts mutants. Positive indices
ranged from 2.0to9,090; 39%wereless than 10.
Intestswhichyielded indices between2and5,
resultswereconsistent fromtest totest,i.e.,they
werealwayslowbutpositive.Althoughmutants
which exhibited high levels of leak were
ex-cluded from the study, HSV-2mutants
consist-ently yielded higherlevels of leak than HSV-1
mutants.Leakinesswasthusasignificantfactor
inthe generation oflowcomplementation
indi-ceswith thesemutants.Clearly,such inefficient
complementation created difficulties in
assign-ingmutantstocistrons.
The failureofts2tocomplement 10mutants
also contributedtodifficulties inassignment of mutants to cistrons. Failure of ts2 to
comple-mentwasnotduetoexcessive leak. It ispossible
that ts2may possesssimilarinterfering
proper-tiestothose of tsP23 of HSV-1 describedabove,
i.e.,whetherts2is abletosuppressreplicationof
wild-type virus atthe nonpermissive
tempera-ture is not known. To determine the relative
distance ofts2fromother mutants,allmutants
in Table 7 except tsl (strain UW268) were
screened for theirabilitytorecombine withts2
at the permissive temperature. Recombination
frequencieswereadditive for themostpart and
ranged from3to 18%, indicatingthatts2could
recombineefficiently, demonstratingthat itwas
notcloselylinkedtomutantswith which itfailed
tocomplement.
Of the 11 HSV-2tsmutantstestedagainstthe standard set, andexcluding theresults oftests
with ts2, all but two, tsll and tsl2 (strain
UW268),failed tocomplement one ortwo
mu-tantsin the standardset.In mixedinfection,tsll
+ tsl2 yieldedaCI of 35 and werethus shown
tobeunique.Thesetwomutantswere,therefore,
placedinnewcomplementationgroups2-19 and
2-20. Four of the 11mutantsagainstwhichthe standard set was tested exhibited overlapping
patterns ofcomplementation (exclusiveoftests
withts2).Because ofthisoverlapand theoverall
low indicesobtained,thesemutantscouldnotbe assigned unequivocally to complementation
groups. Inall,20complementationgroupswere
identified by quantitative complementation
testswithtsmutantsof HSV-2.
Qualitative complementation tests with
tsmutantsofHSV-2. Of198qualitativetests,
9falsenegativetests(4.5%) and12falsepositive
tests(6.1%)wereobserved. Eight of the 12 false
positive tests occurred with the
noncomple-menting mutantts2.Thus,although false
nega-tive andfalse positive tests were obtained with
bothHSV-1 andHSV-2tsmutants,the
propor-tion ofanomalousqualitativeresults was higher
in theanalysisofHSV-2ts mutants.
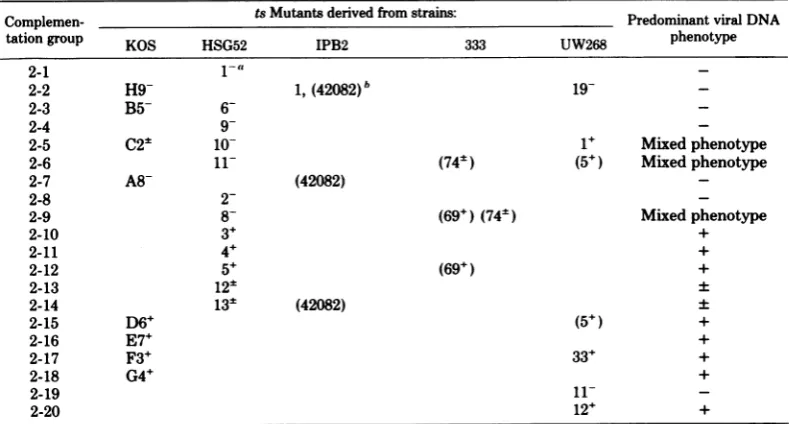
Viral DNA phenotypes of members of complementation groups. Summaries of the complementation groups and the viral DNA
phenotypes ofmutantsin each group are shown inTables8and 9.
Excluding mutants which exhibited
overlap-ping patterns of complementation, the viral
DNA phenotypes of mutants in the 10 HSV-1 complementation groups containing two or more
members are inexcellentagreement (Table 8).
Six of the 23 groups contain DNA- mutants, 3
contain mutants which synthesize20% or less of
wild-type levels of viral DNA, and 11 contain
DNA'mutants.
Ten of the20HSV-2 complementationgroups
maycontainmorethan one member; 4 ofthese
10 groups(2-2, 2-3, 2-5, and 2-15)clearlycontain
two or more mutants. The viral DNA
pheno-typesof mutants inmultimemberHSV-2 groups
were not ingoodagreement. Thebasis for this
disparity maylie in the inavailabilityof defini-tivecomplementationdata orperhapsto
differ-ences in the conditions used for determining
viral DNA phenotypes of mutants in different
laboratories. In any event,availabledataclearly
indicate that 7ofthe 20groups contain
DNA-mutants(groups 1, 2, 3, 4, 7, 8, and
2-19), 2contain DNA'mutants(groups 2-13 and
2-14), and8contain DNA+ mutants (groups
2-10, 2-11,2-12, 2-15, 2-16, 2-17, 2-18, 2-20).
Despitethe fact that theresultsof some
com-plementationtests wereequivocal and that the
viral DNA phenotypes of mutants in certain
multimember groups were dissimilar, 6 of 23
HSV-1 groups and 7 of 20 HSV-2 groups were
shown to contain DNA- mutants. Thus, these
investigationsconfirmthe previous observation
thatalargenumber of cistronsareessential for
viralDNAsynthesis (21, 27).
DISCUSSION
Basedprimarilyonthe results of quantitative
complementation tests,23cistrons of HSV-1and
20 cistrons ofHSV-2 have been identified.
Al-though overlapping patterns of complementa-tion complicated the assignment of some
mu-tants tocistrons,they didnotdiminish the total
number of cistrons identified. Onthe contrary,
overlappingpatterns ofcomplementation have facilitated the identification of potential double
mutants.
The results of quantitative complementation
on November 10, 2019 by guest
http://jvi.asm.org/
R ET AL.
m
J. VIROL.
ko
^e
6
CD
_C4
l^~~
eq -Meq e Xtwo___~__I"- LOeq eq eq eq eq eq eq eq eq eq eq eq ce eq eq eq
+
L3+O
+N;+ cs+
+D+ _+ co+ l£+
+ + L + + +24
+ lq++s
~~~|
+0e>t+
>
~
e~~q~
++t.+o+.+.+.+e,
* e+.+3
+I+`e+ +++g + + ++ m+ X+ + +a
- ' + -++Lo
-++ + + C>+& +c + +D++ + ++ +cQ +ll +ao I 'O + +e
+g +0 +s
+oI
+LO++uVt*+E
+°,l+"q
N+10 so+t-+t+e+eq+
-"' N +.+ +
`@
'tI
+o
+'- + -! +S.+ +.|
+In + +LO m -4mm e cq w w -w m
eq O -4 m e
+ +Lq+)en + + + + L +-!+ Iq+
m V-4_ eq X Ur3e cq
SCHAFFE]
0
.2
eq
500
.4
a
P.
.s
.S
r11
0
.5
ct
O
2
t1
8
P. 4 laE
on November 10, 2019 by guest
http://jvi.asm.org/
tests were considered to be definitive in these
o £ studies. A CI of 2 was selected as the value
x8
AQsignifying
complementation whencomplemen-tation studies with HSV ts mutantswere
initi-2 l, | ated (22). The levelsofcomplementationwhich
representinter- and intracistronic
complemen-v-4 cq tation were notknown at that time, and are not
known now, due to the absence of fine structural
mapping data. Thus, the number of cistrons
AA
S c; X o gSidentified
in this study by the quantitative test7c.s R , , , @ may be somewhat greaterorless than thetrue
numberofcistrons
represented by
themutantsstudied. On theotherhand, problems alsoexist
+88 o with the
qualitative
test.By
using
tsmutantsofHSV-2,
thequalitative
testyielded
resultswhich weresimilartothose obtained ininfectiouscen-ters tests
(30).
Both thequalitative
and infec-o + tious centers tests involvemultiple
rounds ofreplication,and both arecomplicated bythe fact .2 that false positive results couldbe produced
if
recombinants can be
generated
in the absenceofcomplementation. The classical definition of
the cistron is basedupon virusyields produced
X
~
during one round of replication (2, 8) and3_ + E _ g whether recombinationcanoccurinthe absence
of
complementation
isnotpresently known;
con-.! ; sequently, the quantitativetestwas consideredto be more definitive.
I CD + <
.<
Because the limitsofeithertestarenotpres-ently known, it is essential that the cistron
as-signments ofall mutants beconfirmedby
recom-+
*t g bination and markerrescue tests. Genetic and
ba<+e- physical mapping data would be particularly
2E useful in the case of mutants which appeared to
bedoublemutantsand those which
yielded
mul-- +tiple
low andnegative
results(tsP23
ofHSV-1and ts2 of HSV-2) in complementation tests.
Thefeasibilityofmappingviral genes and
poly-o+ .~ peptidesbyanalysisofHSV-1 x HSV-2 recom-binants has recently been demonstrated (18, 19;
N. M.
Wilkie,
N. D.Stow,
H. S.Marsden,
V.Brown, R. Cortini, M. C.
Timbury,
and J. H.8 + @
.i
Subak-Sharpe, Abstracts of the ThirdInterna-tionalSymposium On Oncogenesisand
Herpes-viruses,
Cambridge,
Mass., 1977). It isantici-> o pated that
complementation,
recombination and31+*_ | markerrescue testscombined with analysesof
HSV-1 x HSV-2 recombinants
will
ultimatelylead to the identification of most essential viral
_ M = f cistrons and to thepositioningofthese cistrons
- + _ EZ a onthe physical maps of HSV DNA.
The identification ofadditional viralcistrons
, X
~
!.
5 should continuetoconstituteaprimary goal
ofgenetic studies of HSV. Brown et al. (3)
con-g8eQ E g @
~~cluded
that the HSV genome encodesamini-La
]$R¢ <^, == mum of 30 essential functions. Given the large
-nX>e theoretical coding capacity of the genome, this
value isprobablyaconsiderableunderestimate.
27,
on November 10, 2019 by guest
http://jvi.asm.org/
J. VIROL. 502 SCHAFFER ET AL.
TABLE 8. HSV-I complementation groups
tsMutants derived from strains: Predominant
Complemen- viralDNA
tationgroup KOS 17 KOS 1.1 KOSpp HFEM" HFEMh 13 phenotype
601
1-1 A1' 656- N103-
-1-2 B2- D- 901- LB2:
-1-3 C4 (661j)d,833- (3NI))e
1-4 D9- 84- (3ND))
1-5 E6+ (LS2+) +
1-6 F18+ LS1+, (LS2+) +
1-7 G3: LB1+ C4+ +
1-8 I11+ +
1-9 (J12+) I+ 822+ B5+ G5+ +
1-10 K135 i
1-11 (L145k) F+ LB3+ +
1-12 M195 +
1-13 N20+ G+ +
1-14 022± (661 ) (3N) ±
1-15
P23-1-16 A+ +
1-17 478+ +
1-18 7ND) ND"
1-19
Br-1-20
B7-1-21 LB4+ +
1-22 LB5+ +
1-23 D10-
-"Mutants
derivedfromstrain HFEMbyA.Buchan.bMutants derived from strain HFEMbyI.
Halliburton.
' +, ±, -,and ND are viral DNAphenotypesfromTable 1.
dMutantsinparenthesesareputativedoubleortriplemutants.
'ND, Notdone.
TABLE 9. HSV-2complementation groups
Complemen- ts Mutants derived fromstrains: Predominant viralDNA
tationgroup KOS HSG52 IPB2 333 UW268 phenotype
2-1 1-a
2-2 H9- 1,(42082)b 19-
-2-3 B5- 6-
-2-4 9
-2-5 C25 10- 1+ Mixedphenotype
2-6 11- (74+) (5+) Mixedphenotype
2-7 A8- (42082)
2-8
2-2-9 8- (69+)(74k) Mixedphenotype
2-10 3+ +
2-11 4+ +
2-12 5+ (69+) +
2-13 12: ±
2-14 13- (42082) ±
2-15 D6+ (5+) +
2-16 E7+ +
2-17 F3+ 33+ +
2-18 G4+ +
2-19 11-
-2-20 12+ +
"+,±, and-areviral DNAphenotypes fromTable 1. If phenotype isnotgiven, it hasnotbeendetermined. bMutants inparentheses are putative multiple mutants.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.501.60.455.436.648.2]HSV 503
The results of the present study demonstrate
thatmostinvestigatorshave isolatedtsmutants
inthe same genesand that the numberof
essen-tial cistrons which remain to be identified by
techniquesalready employedisprobablysmall.
Anexplanation for the failure toidentify
ad-ditional cistrons isreadilyapparentfrom Table
1. Of the 72 ts mutantstested, 57wereinduced
with BUdR. Amongthe 23HSV-1
complemen-tation groups, 3 containasinglemutantinduced
by a mutagen other than BUdR (1-10,NTG;
1-12,NTG;and 1-15,AP).Specialmentionshould
bemade of LS1 and LS2-theonlytwo mutants
isolated in the absence of mutagen. Although
difficulttoisolate,spontaneousmutantsarefree ofmutationalartifact.Ofthe20HSV-2
comple-mentation groups, each contains one or more
BUdR-inducedmutants,reflectingthefactthat
26 of the 29 HSV-2 ts mutants tested were
BUdR-induced.
Thatmutational hot-spottingmay occur with HSV DNA and BUdR must be considered in
evaluating the number of cistrons identified in thisstudy.Mutational artifact isa common fea-tureofgeneticsystems inwhich mutagens with
highly specificmodes of actionare
employed
(6,16). Because nucleotidesare notrandomly
dis-tributedinHSVDNA,largeT-rich geneswould bepreferentially mutagenized bytheT-specific
mutagen BUdR. In thefutureit willbe of
inter-est to compare the base compositions of the
sequences known to encode individual viral
geneswith the mutagens usedtoinduce
muta-tions in those sequences. For the present, it is
clear that further attemptstoisolate HSV
mu-tantsshould utilize mutagens whosespecificities
differ from that of BUdR. Nitrousacid, which
canproducetransitions and transversions in all fourbases, wouldseem aprimecandidate.
With regard to mutagenesis with NTG, it
should be noted thattwo of the four
NTG-in-ducedmutantsofstrain KOS usedtoestablish
the standard set of HSV-1 mutants may be
doublemutants. Because NTG isnotorious for
inducing multiple closelylinked mutations (10,
26), this mutagen should be used advisedly. In
additiontotheuseof different mutagens, future
effortsmightalso be directed towards the
isola-tion of other types ofconditional lethalmutants.
Cold-sensitivemutants,forexample, have been
shown tobeclustered inregions of the genome
notcontainingtsmutations(24).
Thepresentstudywas undertakenin partto
identify additional essential HSV cistrons from
amongts mutants ofHSV-1and HSV-2 isolated
in different laboratories. New cistrons both of
HSV-1 and HSV-2have,indeed, been identified.
However, having demonstrated that most
inves-tigators have isolated mutants with defects in
the same series ofcistrons, the need to identify
additionalcistronsbynewmethods isclear. Fur-thermore, recombination analysis, marker
res-cuestudies, and phenotypic characterization of
mutantsassignedtothe same cistron are
essen-tial toverify the results of the complementation
testsreported herein.
The second purpose of thisstudywas to
facil-itatefutureinvestigationswith HSV ts mutants
by (i) introducing a uniformsystem of mutant
nomenclature and (ii)
identifying
mutants in different series which are defective in the sameviralgenes toavoid duplicationofeffort in
phe-notypiccharacterization of mutants. In addition,
it is theintent of thecollaboratorsinthis study
tomake available series of ts mutants
represent-ingeach ofthe known cistrons to facilitate the
identification of additional essential viral genes. Auniformsystemofnomenclatureforts
mu-tants of HSV-1 and HSV-2 as suggested by
Timbury et al. (30) has been introduced. Al-thoughthe systempermits individual
laborato-ries to retaintheir own mutant designations, it
ishopedthatpublicationsinvolving studies with
HSVts mutantswillinclude the cistron
assign-ment of mutants by reference to the common
system suggested here. Numbers rather than
letters have been used toreflectthe large coding
capacity of the virus, and the prefix "1, or "2"
has been used todifferentiateHSV-1and
HSV-2 complementation groups, respectively. The
proposedsystem isnotintended to be rigid but
toform the basis for asystemof nomenclature
which is boundtoevolve asknowledge advances
in thefield.
ACKNOWLEDGMENTS
We thank M. L.Hendricks forexcellenttechnical assistance andM.J. Tevethia forhelpfulcommentsandsuggestions.
Thisinvestigationwasconductedin part inthe Department ofVirologyandEpidemiology, Baylor College ofMedicine, Houston, Tex., supported byPublic HealthServiceresearch contractN01 CP53 526 within theVirus CancerProgram of the NationalCancerInstituteand research grant CA 10,893 fromthe NationalCancerInstitute. The workconductedin Boston wassupported by research grant CA 20,260 and Pro-gram Project grant CA 21,082 from the National Cancer Institute. M.C.T. was therecipient ofanEleanor Roosevelt International Cancer Fellowshipfrom theAmericanCancer Society and aBellahoustonTraveling Fellowship from the ListerCentenary Appeal Fund, UniversityofGlasgow.
LITERATURE CITED
1. Aron,G.M.,D. J. M. Purifoy,and P. A. Schaffer. 1975.DNAsynthesis and DNA polymerase activity of herpes simplex virus type 1 temperature-sensitive mu-tants.J.Virol.16:498-507.
2. Benzer, S. 1957. The elementaryunits ofheredity, p. 70-93. In W. D. McElroy and B. Glass (ed.), The chemicalbasis ofheredity. The Johns Hopkins Press, Baltimore.
on November 10, 2019 by guest
http://jvi.asm.org/
504 SCHAFFER ET AL.
3. Brown,S.M.,D. A.Ritchie,and J. H.Subak-Sharpe. 1973.Geneticstudies withherpessimplexvirus type 1: theisolation oftemperature-sensitive mutants, their arrangement intocomplementationgroups and recom-binationanalysisleadingtoalinkagemap. J. Gen. Virol. 18:329-346.
4. Chu,C.-T.,and P.A. Schaffer. 1975.Qualitative com-plementationtestfortemperature-sensitivemutantsof herpessimplexvirus. J. Virol. 16:1131-1136.
5. Delius, H.,and J. B. Clements. 1976. Apartial denatur-ationmapofherpessimplex virustype1DNA:evidence for inversions of the twouniqueDNAregions. J. Gen. Virol. 33:125-133.
6. Drake, J.W. 1963.Propertiesofultraviolet-induced rII mutantsofbacteriophage T4. J. Mol. Biol. 6:268-283. 7. Dreesman, G. R., and M. Benyesh-Melnick. 1967. Spectrum of humancytomegalovirus complementfixing antigens. J. Immunol.99:1106-1114.
8. Edgar,R.S., G.H.Denhardt,andR. H.Epstein.1964. Acomparativegeneticstudyofconditionallethal mu-tations ofbacteriophage T4D.Genetics49:635-648. 9. Esparza, J., D. J. M.Purifoy,P.A.Schaffer,andM.
Benyesh-Melnick. 1974. Isolation, complementation andpreliminaryphenotypic characterization oftemper-ature-sensitivemutantsofherpessimplexvirus type 2. Virology57:554-565.
10.Guerola, N., J. L Ingraham,and E.Cerda-Olmedo. 1971.Introductionofclosely-linkedmultiplemutations bynitrosoguanidine. Nature (London) 230:122-124. 11. Halliburton,L.W., R. E. Randall, R. A.Kfllington,
and D. H. Watson.1977.Some propertiesofintertypic recombinants betweentype 1andtype 2 herpes simplex virus.J.Gen. Virol.36:471-484.
12. Halliburton,L. W., and M. C. Timbury. 1973. Charac-terizationoftemperature-sensitivemutants of herpes simplex virustype 2.GrowthandDNAsynthesis. Vi-rology54:60-68.
13. Halliburton,I.W., andM.C.Timbury.1976. Temper-ature-sensitivemutants of herpes simplex virus type 2: description ofthree newcomplementationgroups and studiesontheinhibitionsofhostcellDNA synthesis. J. Gen. Virol.30:207-221.
14. Hughes, R. G., Jr., and W.H.Munyon.1975. Temper-ature-sensitivemutantsofherpes simplex virus type 1 defectiveinlysis but not intransformation. J.Virol. 16:275-283.
15. Koment,R.W.,andF.Rapp. 1975. Temperature-sen-sitive hostrange mutants of herpes simplex virus type 2.J. Virol.15:812-819.
16. Krieg,D.R. 1963.Specificityofchemicalmutagenesis. Prog.NucleicAcidRes.2:125-168.
17. Manservigi,R. 1974.Methodforisolationandselection oftemperature-sensitivemutants of herpes simplex vi-rus.Appl. Microbiol.27:1034-1040.
18.Morse,LS., T. G.Buchman,B.Roizman, andP.A. Schaffer.1977.Anatomyofherpes simplex virus DNA. IX.Apparentexclusionofsome parental DNA
arrange-mentsin the generation ofintertypic (HSV-1 xHSV-2) recombinants. J. Virol.24:231-248.
19.Morse,LS.,LPereira,B.Roixman,and P. A. Schaf-fer. 1978. Anatomy ofherpes simplex virus (HSV) DNA. X.Mappingof viral genes by analysis of polypep-tides and functionsspecifiedbyHSV-1 x HSV-2 recom-binants. J. Virol. 26:389-410.
20.Purifoy, D. J. M., and M. Benyesh-Melnick. 1975. DNA polymerase induction by DNA-negative temper-ature-sensitive mutants ofherpessimplex virus type 2. Virology68:374-386.
21. Schaffer, P. A. 1975.Temperature-sensitive mutants of herpesviruses. Curr. Top. Microbiol. Immunol. 70:51-100.
22.Schaffer, P. A., G. M. Aron, N. Biswal, and M. Ben-yesh-Melnick.1973. Temperature-sensitive mutants of herpes simplex virus type 1:isolation,complementation and partial characterization.Virology 52:57-71. 23. Schaffer, P. A., M. J. Tevethia, and M.
Benyesh-Melnick. 1974.Recombinationbetween temperature-sensitive mutants ofherpessimplexvirustype1. Virol-ogy58:219-228.
24. Scotti, P. D. 1968. A new class of temperature conditional lethal mutants of bacteriophage T4D. Mutat. Res. 6:1-14.
25. Sheldrick, P., and N. Berthelot. 1974. Inverted repeti-tions in the chromosome of herpes simplex virus. Cold SpringHarborSymp. Quant.Biol. 39:667-681. 26. Siccardi, A. G., F. A.Ferrari,G.Mazza,and A.
Gal-izzi.1976.Identificationofcoreplicating chromosomal sectorsin Bacillus subtilisby nitrosoguanidine-induced comutation. J. Bacteriol. 125:755-761.
27. Subak-Sharpe,J.H., and M. C.Timbury. 1977. Ge-netics ofherpesviruses,p. 89-131. In H. Fraenkel-Con-rat and R. R.Wagner(ed.),Comprehensive virology, vol.9.PlenumPublishing Corp.,New York.
28. Takahashi,M., and K. Yamanishi.1974. Transforma-tionof hamsterembryo and human embryo cells by temperature-sensitivemutantsofherpessimplexvirus type 2.Virology61:306-311.
29.Timbury,M. C. 1971.Temperature-sensitivemutantsof herpessimplexvirus type 2. J.Gen. Virol. 13:373-376. 30.Timbury,M.C.,M.L.Hendricks,andP.A.Schaffer. 1976.Collaborativestudyoftemperature-sensitive mu-tants of herpes simplex virus type 2. J. Virol. 18:1139-1142.
31. Wadsworth, S.,R. J.Jacob,and B.Roizman. 1975. AnatomyofherpessimplexvirusDNA.II.Size, com-position andarrangement ofinvertedterminal repeti-tions. J. Virol. 15:1487-1497.
32.Westmoreland, D., and F. Rapp. 1976. Host range temperature-sensitivemutantsofherpessimplexvirus type 2. J. Virol. 18:92-102.
33. Zygraich, N.,and C.Huygelen.1973.Invivobehavior of atemperature-sensitive (ts)mutantsofherpesvirus hominis type 2. Arch.Gesamte Virusforsch.43:103-111. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/