0022-538X/79/01-0034/09$02.00/0
Isolation
of
a
Nucleocapsid Polypeptide
of
Herpes
Simplex
Virus
Types
1
and 2
Possessing Immunologically
Type-Specific
and
Cross-Reactive
Determinants
CONRAD J. HEILMAN,
JR.,`*
MARTIN ZWEIG,' JOHN R. STEPHENSON,2 ANDBERGE HAMPAR3ViralOncologyProgram,FrederickCancer ResearchCenter,Frederick, Maryland217011;Laboratory of CellularandMolecular Biology,National CancerInstitute,Frederick Cancer ResearchCenter, Frederick, Maryland 217012;andLaboratory ofMolecularVirology,National CancerInstitute,Frederick Cancer
ResearchCenter, Frederick, Maryland217013
Received for publication 21 August 1978
A polypeptide (p40) of approximately 40,000 molecular weight was isolated
from herpes simplex virus type 1 and 2 nucleocapsids by gel filtration and ion
exchangechromatography. This protein appearstobe the same as protein 22a
describedpreviously (Gibson and Roizman, J. Virol. 10:1044-1052, 1972).
Com-petition immunoassayswere developed by using purified p40 and antisera
pre-pared in guinea pigs. The assays indicated that the p40's from herpes simplex
virustypes1and 2possessbothtype-specific and cross-reactive antigenic
deter-minants.Antibodiestothe p40cross-reactive determinant reacted withantigens
in simian herpes virus
SA8-infected
cells, but not with antigens induced bypseudorabies virus. Preliminary results indicated thataradioimmunoprecipitation
test can be used to detect type-specific herpes simplex virus p40 antibodies in
humansera.
Herpes
simplex virus
types 1(HSV-1) and
2(HSV-2)
arelarge
complex
DNA-containing
vi-ruses which have been shown to
induce the
synthesis
of atleast
50 newpolypeptides in
productively
infected cells
(5, 13). Some
ofthe
antigens
inducedby
HSV-1and HSV-2
havebeen shown
to possess cross-reactivedetermi-nants
(which
reactwith antiserumagainst
eitherHSV-1
orHSV-2), whereas other antigens
pos-sesstype-specific
determinants
(which
reactonly
with antiserum
against
thehomologous
virus type) (4, 15).
Several HSV-induced
polypeptides
have
beenpurified
and
immunologically
characterized
by
using
monospecific
antisera and
avariety
of
immunological
techniques (for
areview,
seeref-erence
9). These include the
early
nonstructural
polypeptides
VP 175(molecular weight,
175,000)and VP134, the
envelope glycoprotein
VP 123,and
thenucleocapsid
polypeptide
VP 154.The
nucleocapsids
ofHSV-1 and HSV-2con-tain six
principal
polypeptides ranging
inmolec-ular
weight
fromapproximately
12,000 to154,000 (2; G. H.
Cohen,
W. C.Lawrence,
M.Ponce DeLeon, S.
Vernon,
and H.Diggelman,
Abstr. Annu. Meet. Am. Soc. Microbiol. 1978,
S42,
p. 219; and seebelow).
Thenucleocapsid
polypeptide
VP 154has beenreported
topossesscross-reactive
antigenic
determinants (9, 14).Thestudies described here
concern
theantigenic34
properties of another
nucleocapsid
polypeptide
of HSV-1
and HSV-2 withamolecularweight
ofapproximately
40,000
(p40).
Inpreliminary
stud-ies, p40
wasfound
tobe among the mostsoluble
of the
major
nucleocapsid proteins
and also themost
amenable
to'25I-labeling
athigh specific
activity without
significant degradation
orag-gregation.
Because theseproperties
aredesirable
in
immunoassays,
weselected
p40 for
ourinitial
characterization studies.
Using the
competition
immunoassay
technique
withpurified
p40's
fromHSV-1
andHSV-2
andantisera
prepared
inguinea
pigs,
wefound
that thispolypeptide
pos-sesses
both
type-specific and cross-reactive
an-tigenic
determinants.
MATERIALS AND METHODS
Cellsandviruses.Verocellsweregrown inroller
bottlesorflasks inEagle minimal essential medium supplementedwith10% heat-inactivatedfetal calf
se-rum,penicillin,and streptomycin. MAL (type 1) and
MS (type2) were theprincipal HSV strains employed. Other herpesvirus strains included 14-012 (HSV-1), Miyama (HSV-1), 333 (HSV-2), Savage (HSV-2), pseudorabiesvirus(PRV),and simianherpesvirusSA8 (8).The Verocellswereinfected at amultiplicity of infection of 1 to 5, based on infectious virus titers. Afteradsorptionfor 1 h at 37°C, fresh medium was added, and the cells were maintained at 37°C until maximumcytopathic effectswere observed (18 to 36 hpostinfection). Thecellswere washedwith cold PBS
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 29, 1979
(0.14NaCl,0.02Msodiumphosphatebuffer, pH7.4)
and collectedby centHfugation (1,500xgfor 5 min).
Cellpelletswerestoredat-70°C untilused.
Purification ofnucleocapsid. Cells grown in
roller bottles were infected with either HSV-1 MAL orHSV-2 MS(multiplicityofinfection,5) asdescribed above andharvested at 24 h postinfection. Thecells
weresuspended in 0.01 MNaCl-0.01 M Tris-hydro-chloride-1.5 mMMgCl2, pH 7.5(bufferA) and incu-bated at0°Cfor 10min. The cells weredisruptedin a Douncehomogenizer in the presence of 1%Nonidet P-40 at0WC,and the nuclei were sedimentedby
cen-trifugation (600 xgfor 5 min) and resuspended in
bufferA.Nuclearlysateswereprepared byaddition of
sodiumdeoxycholateto afinal concentration of0.5%,
followedby sonic treatment. After incubation of
ly-satesfor 30min at37°Cin the presence of 50
Ag
of DNase perml,urea wasadded to afinal concentration of 0.5 M, and debriswasremovedbycentrifugationat8,000 rpm for 5 min in a Sorvall SS34 rotor. The
clarified lysatewaslayeredonto a6-mlcushion of 35%
(wt/vol)sucroseinPBScontaining1mMEDTA and
was centrifuged at 23,000 rpm for 1 h at4°C in a
Spinco SW27rotor(12). Theresulting pelletwas
re-suspendedinPBS, subjectedtosonictreatment,and
layeredontoa33-ml10 to40% (wt/wt)sucrose
gra-dientpreparedinPBSoverlayinga3-ml60%(wt/wt)
sucrose shelf. Centrifugation was performed in a
Spinco SW27rotorfor 60minat20,000rpmat
8°C
(2). Two prominent
nucleocapsid-containing
bands were formed near the middle of the tube andwere collected from the bottom of the tube. Thecontents of the bandswerenegatively
stained withphospho-tungsticacid, examined by electron
microscopy,
andfoundtobefreeof
contaminating
material. Both bandswerepooled,diluted withPBS,sedimentedat23,000
rpm for 1 h at
4°C
in aSpinco
SW27 rotor, andsuspendedinwater.
SDS-PAGE.
Sodiumdodecyl
sulfate-polyacryl-amide gelelectrophoresis
(SDS-PAGE)
wascon-ducted inaBio-Rad model220slab
gel
apparatusfor 16h at 30 V byusingadiscontinuousbuffer systemcontainingalinear 5to20%
(wt/vol)
polyacrylamide
gradient (7). Theprotein bands werefixed in 12.5%
(wt/vol) trichloroacetic acid for1handstained with
0.05%Coomassie brilliant blue R250 in 12.5%
(wt/vol)
trichloroacetic acid for4to16h,
andexcessdye
was removed by washing in5% methanol and 7% acetic acid. Foranalysis
ofl"I-labeled
proteins,
thegels
weredriedontofilter paper withaBio-Rad model224
gel
slab dryer, and
autoradiographs
wereprepared
onKodakX-Omat R film. Absorbance of bands in stained
gelsand
autoradiographs
wasmeasuredby
asoftlazer scanning densitometer(Biomed
Instruments,
Inc.,
Chicago,Ill.).
Isolation ofviralproteins.Purified
nucleocapsids
(totalprotein,10mg)were
suspended
in 0.5ml of0.01M dithiothreitol (DTT)-2 mM EDTA-0.05 M
Tris-hydrochloride (pH 8.5) buffer
containing
8Mguani-dinehydrochlorideandappliedto anagaroseA-5
100-to200-mesh(Bio-RadProducts)
gel
filtration column (0.5by90cm)in 0.02 M sodiumphosphatebuffer, pH
6.5,containing6Mguanidine
hydrochloride
and0.01MDTT(1).Column fractions
containing
amajor peak
at amolecularweightof about40,000asdetermined
NUCLEOCAPSID p40 FROM HSV 35
bySDS-PAGEwerepooled and
exhaustively
dialyzedagainst 200 volumes of a solution containing 0.01
M N,N-bis-(2-hydroxyethyl)-2-aminoethanesulfonic
acid, pH 6.5, 1.0 mM EDTA, and 0.1% Triton X-100 (bufferB) andappliedto aphosphocellulose column (1.0by 5.0 cm;Whatman pll;H.Reeve Angel & Co., Inc., Clifton, N.J.) previously equilibrated with the same buffer. The column was washed with 50 ml of bufferB,and bound proteins were eluted with a 100-mllineargradient (0to 1.0 MNaCl).Fractions eluting at 0.64 to 0.82 MNaCland containing a single protein of about 40,000 molecular weight were pooled, divided
intoaliquots,andstored at -700C.
Antiserapreparation. HSV-1 MAL and HSV-2
MSnucleocapsidproteins (approximately 200ltg)were
separatedbySDS-PAGEon slab gelsand stained with
Coomassie brilliantblue. The 40,000-molecular-weight
bands were cut out,minced in 2 ml of PBS with a7-ml
Dounce homogenizer, and subsequently emulsified
with an equal volume of Freund adjuvant. The antigen
preparations (2ml)were injected into Hartley guinea
pigs intramuscularly. The animals were immunied
threetimesatweekly intervals, and sera were obtained 10 days after the last injection. Complete adjuvant wasused in the first injection, and incomplete adju-vant was used in the two subsequent immunizations.
Immunoassays. Viral proteinswere labeled with
'25Iathigh specificactivity(100,uCi/,ug) by the
chlor-amine T procedure (3) andseparatedfromfreeiodine
byplO (Bio-Rad Products) column chromatography.
Duringiodination, however, it was necessary to reduce
the chloramine T concentration to 3.6 ug/ml to
pre-ventdegradationof the protein. Immunoprecipitation
assays were done byincubating
l"I-labeled
p40(10,000 cpm) with serialtwofolddilutions of antisera.Com-petitionimmunoassayswereperformedby testing
un-labeled viral antigens at serial twofold dilutions for
ability to compete with
l"I-labeled
p4O for bindinglimited amountsof antiserum. Antisera in the
com-petitionimmunoassays were employed at adilution
(1:400 to 1:5,000) which precipitated about 35% of the
"SI-labeled
p40.Antiserum andunlabeled competingantigenwereincubatedfor 1 h at37°C, followedby
addition of 10,000 cpm of
'25I-labeled
p40. Reaction mixtures contained 0.01 MTris-hydrochloride,pH 7.8, 1.0 mMEDTA,0.4% TritonX-100, 1% bovine serumalbumin, 0.05 M
NaCL
and0.05% NaN3 in a totalvolume of 0.2 ml. The antigen-antiserum mixtures wereincubated for 3 h at37°Cand 18 h at4°C;20Ml
of a second antibody (goat anti-guinea pig
immuno-globulin G;Office of Resources andLogistics, National
CancerInstitute,Bethesda, Md.)wasadded,and in-cubationwascontinued for1h at37°Cand 3 h at4°C.
After the addition of 0.04 ml of 10 mM
Tris-hydro-chloride (pH 7.8)-10 mM NaCl-0.1% Triton X-100, samples were centrifuged for 15 min at 2,500 rpm, supernatants wereaspirated, and radioactivity in the
precipitates was measured in a Searle model 1285
gammacounter.
RESULTS
Isolation and
immunoprecipitation
of
HSV-1 and HSV-2
nucleocapsid
p40's.
The
p40's
ofHSV-1
and HSV-2nucleocapsids
wereon November 10, 2019 by guest
http://jvi.asm.org/
36
HEILMAN ET AL.isolated
by
gel filtration and ion exchange
chro-matography.
After
labeling
athigh specific
ac-tivity
with
1"I,
their purity was
determined
by
SDS-PAGE
analysis.
The
nucleocapsids
of
HSV-2
(Fig.
1)
and
HSV-1
(data not shown)
consist of
six well-defined major proteins and
about six minor
proteins, with
molecular
weights
ranging
from 155,000
(p155)
to12,000
(p12). For
clarity,
only
the
major
components are
identified
in
Fig. 1. The
12'I-labeled
p40
of
HSV-2 migrated
as a
single
homogenous
band of
radioactivity
ata
molecular
weight of about 40,000, which
cor-responded to a
Coomassie
brfilliant blue-stained
protein
band
observed
after
SDS-PAGE
analy-sis of
unlabeled
nucleocapsid preparations.
The
corresponding
protein of
HSV-1
was
slightly
lower in
molecular weight (data
not
shown).
Immunoprecipitation
tests were
performed
by
using
the
'25I-labeled
p40
proteins of HSV-1 and
HSV-2 and antisera
prepared in guinea pigs. As
shown in
Fig.
2, the
homologous
p40
was
precip-itated with a
higher
dilution of antiserum than
was the
heterologous
p40.
The
difference
in
an-tiserum dilution for
precipitation
of
homologous
versus
heterologous
p40
was
greater with the
anti-HSV-2
serumthan with the
anti-HSV-1
serum.
One
possible
model accounting
for
this
phenomenon is that both antisera contained
twoclasses of
antibodies, one which reacted with
type-specific determinants and
asecond which
reacted with
cross-reactive determinants. The
ratio of each class of
antibody
in any
oneserumpreparation could vary.
Support
for the
proposal
that
these sera
contained two classes of
antibod-ies
is provided in the
immunoassays
described
below.
Homologous
competition
immunoassays
for p40
in
fast- and
slow-sedimenting
nu-cleocapsids.
It has been
reported
that
HSV-infected cell
nuclei
contain two
species of
nu-cleocapsids, designated
A
and B
(2).
These
stud-ies showed that the B
nucleocapsids
contain
10times the amount of DNA and sediment faster
compared with the A
nucleocapsids.
In
addition,
B
nucleocapsids
contain
amajor protein
com-ponent
(protein 22a) not found in A
nucleocap-sids. We have
also observed two different classes
of
nucleocapsids which sediment at
different
rates and appear to
correspond to
the A and B
groups.
SDS-PAGE
analysis
of
preparations of
the
slow-sedimenting
class of
nucleocapsids
clearly revealed
that
they possessed
relatively
lower amounts of
p40
than did preparations
containing the
fast-sedimenting
class
(Fig. 1 and
Hsw4ris
Nluc§eocap.S4t-.;;f',,;
r'rx,tst
-e,i:,r.:s.-
; r,c: p _*a:
--er:--S
ses!J
F4 t_^
FIG. 1. SDS-PAGE analysis ofpolypeptidesand
'"I-labeledp40
fromdisruptedHSV-2 MSnucleocapsids. (A)Scanning densitometer tracing of Coomassie brilliant blue-stainedgel. (B)Autoradiogram ofgelcontain-ing
125I-labeledp40.
(C)Scanningdensitometertracing ofautoradiogram. Molecular-weightstandards usedincluded
,8-glactosidase
(130,000),phosphorylase a(92,000),ovalbumin (43,000), DNase I(32,000),andmyoglo-bin(17,000).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
NUCLEOCAPSID p40 FROM HSV 37
RECIPROCAL ANTISERADILUTON(bilo)
FIG. 2. Immunoprecipitation ofHSV-1and HSV-2 '251I-abeled p40 by homologous and heterologous guineapigantisera. 1251-labeledp40 (10,000cpm)from
HSV-1 (A) orHSV-2 (0) wasincubatedwith serial twofolddilutions ofantisera elicited againsteither HSV-1 (A)orHSV-2(B) p40. The reactionmixtures andconditionswere asdescribed in thetext.
3). This finding, together with similarities in
molecularweight, suggeststhatp40is thesame
asprotein22a and thatourslower-sedimenting
nucleocapsid preparations probably representA
nucleocapsids. To establish thatour
immunolog-icalprocedureswere assaying aprotein having
thepropertiesofprotein22a andp40,a
homol-ogous HSV-2 competition immunoassay was
performed by usingunlabeledpurified p40 and
preparationsofdisruptedfast(B-enriched)-and
slow(A-enriched)-sedimenting nucleocapsidsas
competing antigens (Fig. 4). Precipitationof
la-beledantigenwas50% inhibitedbythepresence
of about 1.5 ng of purified p40, 10 ng of
fast-sedimenting nucleocapsids, or 170 ng of
slow-sedimenting nucleocapsids.Thisassayindicates
thatp40accounts for about 15%of theprotein
found in
fast-sedimnenting
nucleocapsids andabout 1% of the
protein inslow-sediinenting
nucleocapsids.
Verysimilar
results wereob-tained
inhomologous HSV-1
assays (data notshown). Preparations of disrupted
HSV-1nu-cleocapsids
did not compete inthe homologousHSV-2 immunoassay, further
suggesting theex-istence of type-specific
antibodies top40.
Theseassays
also show that
ourtotal
nucleocapsidpreparation contained predominantly
fast-sedi-menting (B) nucleocapsids, because
the totalnucleocapsid
preparation contained about thesame amount
of
p40
asdid
the Bnucleocapsid-enriched preparation.
Homologous
competition
HSV-1
andHSV-2 immunoassays for p40 in cell
ex-tracts.
In
the
homologous HSV-1 immunoassay
(Fig. 5A), unlabeled
extractsof
HSV-l-infected
celLs inhibited
precipitation ofHSV-1
'WI-la-beled
p40, whereas
extractsof cells infected with
HSV-2
orSA8 virus
competed toonly
asmall
extent.
No
competition
wasobserved with
ex-tracts
of
uninfected cells
orPRV-infected
cells.
In the
homologous HSV-2 immunoassay (Fig.
5B),
extractsof HSV-2-infected
cells inhibited
i
FIG. 3. SDS-PAGEanalysis ofthepolypeptides of theslow-sedimenting nucleocapsids. PurifiedHSV-2 MS slower-sedimenting nucleocapsids (30ug) were
subjectedtoelectrophoresis, andascanning densi-tometertracingwasmadeoftheresultingCoomassie brilliant blue-stained polyacrylamide gel. Two classesof nucleocapsidswerepurified byseparately
collecting the slow (A nucleocapsid-enriched)- and fast (B nucleocapsid-enriched)-sedimenting bands
formed afterthesucrosegradient centrifugation
pro-cedure described in thetext. Thenucleocapsidswere
diluted with PBS, sedimented by centrifugation at
23,000rpmfor 1 h at4°Cin a SpincoSW27rotor,
andsuspendedin 0.01 MTris-hydrochloride-0.001M EDTA, pH 7.5. After sonic treatment at 0°C, the nucleocapsids were layeredonto a 16-ml 45to 65% (vol/vol) Renografin 76gradientpreparedin0.01 M
Tris-hydrochloride-.001MEDTA, pH7.5, and
cen-trifugedtoequilibriumat25,000rpmfor3 hat4°Cin
aSpincoSW27.1 rotor(12). The slow- and fast-sedi-menting nucleocapsids formed lighter and heavier
bands, respectively, aftercentrifugationin the Reno-grafin 76gradient (2, 11). Theslow-sedimenting
nu-cleocapsidswerediluted with 0.01 M Tris-hydrochlo-ride-0.001 MEDTA, pH 7.5,pelleted by centrifuga-tionat23,000rpmforIh at4°CinanSW27.1rotor,
andfinally suspendedin water.
a
z
0
z0
C
F. z
a
5,
A Nucloocapsids
p156 p50 p32 p25 p12
VOL. 29, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.58.214.53.410.2]38
HEILMAN ET AL.0
20
0~~~~ ~ ~ ~~
0 2 3 4
[image:5.501.69.267.54.276.2]Competing Nucleocapsid Protein(loglong) FIG. 4. Homologous competition immunoassay for p40in whichpurified p40anddisrupted nucleocap-sidswereusedascompetingantigens. Total
nucleo-capsids, purified asdescribedin thetext, and
slow-andfast-sedimenting nucleocapsids, purifiedas
de-scribed in thelegendtoFig. 3,weredisruptedin 0.01
M Tris-hydrochloride (pH 7.8)-0.01 M NaCI-0.1% Triton X-100containing1%SDS and 5 mM DTTby heating at100°C for5min. Thepreparationswere
then dilutedfourfoldwith the same buffer without
SDS orDTT and tested atserial twofolddilutions
for abilitytocompete with '25I-labeledHSV-2 MSp40 (10,000 cpm) for binding limiting concentrations of guinea pigantiserum(diluted1:5,000)elicitedagainst HSV-2 MSp40. Thecompeting antigenswereHSV-2
MSp40 (0; purified asdescribed in the text),
dis-ruptedtotalnucleocapsids ofHSV-1 MAL (Q) and
HSV-2 MS (0), and disrupted slow (V)- and fast-sedimenting(A) nucleocapsids ofHSV-2 MS.
precipitation, whereas uninfected cell extracts
and extracts of HSV-1-, PRV-, or SA8
virus-infected cells didnotdetectably compete. The
homologous assays showed that about 5 x
104
HSV-1-orHSV-2-infectedcellsextracted late in
infectioncontainedapproximately0.2jigofp40.
The reactivities ofp40in thesehomologous
as-saysindicatedthat theproteinsfromboth HSV
typespossesstype-specificimmunological
deter-minants. To verify that a single protein was
beingcharacterizedbythe aboveimmunoassays,
unlabeled nucleocapsid polypeptides were
sub-jectedtomolecularsizing analysis by gel
filtra-tion in thepresenceof 6 Mguanidine
hydrochlo-ride, and individualcolumn fractionsweretested
by competition immunoassay. Reactivityinthe
homologous HSV-1 p40 assay (Fig. 6A) and
HSV-2 p40assay (Fig. 6B)chromatographedas
a single homogenous peak at approximately
40,000molecular weight. The specificity of the
p40 antiserum reaction was also tested after
separating nucleocapsid polypeptides by
SDS-PAGE (Fig.
7),
because gel filtration did notprovide good separation of the p50, p40, and p32
components.After thepolyacrylamide gels were
stained and sliced into segments, the
polypep-tides wereeluted, and the eluants were tested by
competition
immunoassay. Reactivity
wasonly
eluted from thegel segments containing the p40
protein.
To characterize the competing
antigen withrespect to
charge,
an HSV-2 p40 preparationwas
analyzed
bycolumn isoelectric focusing. Asshown inFig. 8, awell-defined peak of competing
reactivity was found at pH 6.7, which provides
further
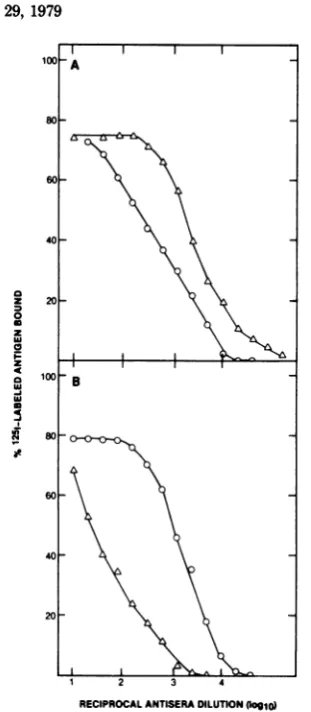
support for the high specificity of our immunoassay.Heterologous
competition immunoassay
for HSV-1 and HSV-2
p40.
Theprecipitation
of
heterologous p40
byguinea
pig antisera atrelatively
high
titers(Fig.
2)suggested that these
nucleocapsid proteins also contain shared
deter-minants. To
testthis
possibility, competitionimmunoassays with HSV-1 p40 (Fig.
9)and
HSV-2 p40 (data
notshown)
wereperformed
with
heterologous antisera.
In both cases, theresults
wereessentially the
same. In an assay inwhich
antibody
to HSV-1p40
was utilized atlimiting dilution for precipitation
of1251-labeled
HSV-2 p40,
extractsofcells
infected with eitherHSV-1
orHSV-2 competed
with comparablehigh efficiencies (Fig.
9).Similar
highefficiencies ofcompetition
wereobserved
whenHSV-1 andHSV-2 nucleocapsids
were used as competingantigen (data
notshown). Uninfected cell
ex-tracts
and
extractsof
cells infected
with PRVwere not
reactive,
whereas extracts ofcells
in-fected
withSA8
viruscompeted to afinal
extentof 50%.
Immunoprecipitation of
p40 with
ab-sorbed
sera. Tofurther
establish that
p40
con-tains both
type-specific
andcross-reactive
deter-minants, immunoprecipitation
tests wereper-formed
with seraabsorbed
withunlabeled
HSV-1 orHSV-2 disrupted nucleocapsids
(Fig. 10).Absorption of anti-HSV-2
p40
with HSV-1nu-cleocapsids
did not measurably reduce these-rum titer against
'25I-labeled
HSV-2p40,
whereas
absorption
withHSV-2 nucleocapsids
caused a substantial reduction in titer.Similarly,
a
slight decrease
intiter against HSV-1p40
wasdetected after
absorption
of HSV-1 antiserumwith HSV-2
nucleocapsids,
whereas a very largereduction in titer was observed afterabsorption
with HSV-1
nucleocapsids
(data not shown).Theseresults suggest that
p40
induces thepro-duction of one
class
of antibodies which reactwith type-specific
determinants
and a secondclass which bindcross-reactive determinants.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
NUCLEOCAPSID p40 FROM HSV
I
2
0
0
i
-I
c31d
I-0
40
20
Fraction Number
FIG. 6. Specificity of competition
radioimmunoas-sayfor HSV-1 (A) and HSV-2(B)p40'sby guinea pig antisera. Nucleocapsids (1 mg) were disruptedand
appliedtoaBio-Gel A-5agarosecolumn(1.5 by90
cm) in thepresenceof6Mguanidine hydrochloride
and 0.01 M DTT. Fractions (1.0ml) were coUected,
dialyzed against 0.01 M Tris-hydrochloride (pH
7.8)-i mMEDTA-0.1% TritonX-100, andtested at
serialtwofolddilutionsfortheirabilitytocompete in thehomologous p40 immunoassays. Results are
ex-pressedasthepercentageoftotalantigenic reactivity
in eachcolumnfractionandarebasedonthe dilution atwhich 50% displacement of the
"*I-labeled
antigenwas achieved. Mokcular-weight markers include blue dextran (>100,000), bovine serum albumin
(68,X000), Rauschermurineleukemiavirusp30(30,X000),
avian leukemiavirusp12(12,000), andbromophenol blue(<1,000).
Reiprocal Antigen Dilution(logio)
FIG. 5. Homologous competitionimmunoassayfor
HSV-1 and HSV-2 p40. Unlabeled infectedcell
ex-tractsweretestedatserial twofold dilutionsfor their abilitytocompete with(A)
"LI-labeled
HSV-1 MAL p40(10,000 cpm) for binding limiting concentrations of guinea pig antiserum (diluted 1:400) directed against HSV-1p40, and (B)125I-labeled
HSV-2 MS p40(10,000 cpm) for binding limitingconcentrations of guinea pig antiserum (diluted 1:5,000) directed against HSV-2p40. CeUswere infectedatamulti-plicityof infection of Iasdescribedin thetextand
harvestedatthe timewhenmaximumcytopathic ef-fects wereobserved. This wasat18h postinfection with theHSV-1strainsandat36h postinfectionwith HSV-2strains, PRV, and SA8 virus. Cell extracts
werepreparedbysuspending approximately2.5x 106
cells in 0.5mlof PBS,subjectingthemtosonic treat-ment,andaddingSDS andD1T tofinal
concentra-tionsof1% and 5
mM,
respectively.The extractswereheatedat IOOOCfor2min, debriswas removed by centrifugation (1,2(M) xgfor15min),andthe extracts were diluted in 0.01 M This-hydrochloride (pH 7.8)-0.01 MNaCl-0.1% TritonX-100. Undiluted
com-petingantigenin the reactionmixturerepresentsan extractofabout 5xIO ceUs.Thecompetingantigens
wereextractsofcellsinfectedwith HSV-1 MAL(0), HSV-1 14-012 (0), HSV-2 MS (A), HSV-2 Savage (V), HSV-2333 (A), SA8virus (L), PRV(U), and
uninfectedVero cells(0).
C ->
I IIs
F
I I I ID->
30 50 70 90 10
VOL.
29,
197939
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.246.442.50.338.2] [image:6.501.70.214.66.518.2]40
HEILMAN ET AL.FractionNumber
FIG. 7. Specificity of competition radioimmunoas-say for HSV-2 p40 as shown by SDS-PAGE. Dis-ruptedHSV-2 MSnucleocapsids (approximately30
ug) were subjected to electrophoresis in cylindrical
12%polyacrylamide gels (7), stained with Coomassie brilliant blue, and scanned for absorbance (A). The gelwaswashed severaltimes with water, frozen, and sliced into 1.35-mm segments. Protein was eluted overnightat roomtemperaturefrom each segment in 0.5mlof elutionbuffer containing0.1MTris, pH7.8,
0.4MNaCl,and 0.1% SDS. Eachfractionwastested
forcompetition(B)inahomologousassayfor HSV-2
MSp40in serialtwofolddilutionsasdescribed in the
text.Resultsareexpressedasthe percentageof total antigenic reactivity in eachfraction andare based onthedilutionatwhich50%o displacementof the
1"5I
labeledantigenwasachieved.Analysis
of human
serafor antibodies
toHSV-1 and HSV-2
p40's.
In
view of the
sen-sitivity of the
immunoprecipitation procedure
and its
ability
todiscriminate 1 and
HSV-2
p40's,
it
wasof interest
toassessthe
potential
application
of this
test toanalysis
of
human
serafor antibodies directed
against
either
HSV-1
orHSV-2.
For this purpose, aseries ofserafrom
individuals with
suspected
HSV-1
orHSV-2
ex-posure were tested
for
immunoprecipitation
of'25I-labeled
HSV-1 and HSV-2
p40 proteins,
bothdirectly
and
afterabsorption
with
unlabeled
de-tergent-disrupted
nucleocapsids.
We had
anticipated
from the
p40
immunoas-say results with
guinea pig
antisera that wewould be able todivide the humanserum
sam-ples
into fourcategories: (i) HSV-negative
sera,(ii) anti-HSV-1
sera,(iii)
anti-HSV-2
sera, and(iv) anti-HSV-1
and -HSV-2 sera.Our
prelimi-nary
findings
(Table 1)
indicated that each ofthe sera tested
clearly
fall
into one of the fourexpected
categories.
Assignments
ofthese seratothe category of
anti-HSV-1
positive
oranti-HSV-2
positive
wasbasedonthedemonstration
that
absorption
of the serum with one antigencaused a substantial decrease in its ability to
precipitate
1251-labeled
p40,
whereas absorptionwith the other
antigen resulted in
only
a smallreduction in its
precipitating
activity. The
exist-ence of
human antibodies
tothe cross-reactive
determinant(s) of p40
wassuggested
by
the
find-ings that both
theanti-HSV-1 and
anti-HSV-2
sera
precipitated the labeled
heterologous
pro-tein and that these
precipitations
wereblocked
by
absorption with either
homologous
orheter-ologous nucleocapsids.
DISCUSSION
The
approximately
40,000-molecular-weight
(p40)
polypeptides
from HSV-1 and
HSV-2
nu-cleocapsids
werepurified, and antisera
werepre-pared for utilization
indirect and
competition
radioimmunoassays. The
findings
indicated that
the
p40's of HSV-1 and HSV-2
possessboth
cross-reactive and
type-specific antigenic
deter-minants. The various
procedures
carried
out toverify the
purity of the p40 isolates
suggeststhat
both the
cross-reactiveand
type-specific
deter-minants
reside
on the samepolypeptide
mole-cule.
This issimilar
towhat
has beendescribed
for
polypeptides of mammalian oncornaviruses
(10).
50
I-4
z
m
4c
40
30
20
10
o0
6
2
10 20 30
Fraction Number
FIG. 8. Isoelectricfocusing analysis of HSV-2p40. Purified HSV-2 MS nucleocapsids (5 mg)were
dis-rupted and appliedtoan agarose-guanidine
hydro-chloridecolumnasdescribed in the legendtoFig.6.
After dialysis against 0.01 MTris-hydrochloride(pH
7.8)-i mM EDTA-0.01% Triton X-100, fractionswere
analyzed bySDS-PAGEasdescribed in thetext.The
p40-containing fractions werepooled and analyzed
by isoelectric focusing. Focusing wasperformedat
300 Vfor 48 h in the pHrangeof3.5to10 ina
110-ml-capacity column (LKB Instruments, Inc.,
Rock-ville, Md.) (10). Fractions (3 ml) werecollected,
di-alyzed, and analyzed by competition immunoassay asdescribed in thelegendtoFig.6.
-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.70.260.48.215.2] [image:7.501.271.463.321.526.2]NUCLEOCAPSID p40 FROM HSV 41
i
20 lao
02
260
IC
04
-IO
20~~~~
4 3 2 1
ReciprocalAntigenDilution(log91)
FIG. 9. Heterologous competition immunoassay for HSV-2 p40. Cellextracts,preparedasdescribed
in thelegendtoFig. 5, weretestedatserial twofold dilutionsfor abilityto competewith'25I-labeled HSV-2MS p40(10,000 cpm)for binding limitingamounts
ofguinea pig antiserum elicited against HSV-1 MAL p40.The antiserumwasusedatadilution of 1:1,000,
which immunoprecipitated about 30% of the 'MI-la-beled HSV-2 MS p40 antigen (Fig. 2). The competing antigens, which were those used in the experiment
describedinthelegendtoFig.5,wereextractsofcells infected with HSV-1 MAL (0), HSV-1 14-012 (0), HSV-2 MS (A), HSV-2 Savage (V), HSV-2333 (A), SA8 virus(L), PRV (U), anduninfected Vero cells (01).
In agreement with studies
reported by others
(Cohen
etal.,
Abstr. Annu. Meet. Am. Soc.Microbiol.
1978,
S42,
p.219),
thep40's
ofHSV-1 and HSV-2 were localized to the nucleus of
productively
infected cellsby
immunofluores-cent
staining (unpublished
data).
Thecompeti-tion
immunoassays
didnotdetectintratypic
dif-ferences in the
antigenic specificities
ofp40's
from the
different strains of HSV-1 or HSV-2studied.
Antigens produced
in PRV-infectedcelLsfailedtoexhibitdetectable
cross-reactivity
with
the
p40's
of HSV-1 orHSV-2. In contrast,cross-reactivity
between HSV-1 and HSV-2p40's
andantigens produced
incellsproductively
infected
withSA8
viruswasobserved,
which isconsistent with
previous
reports(6, 16)
thatthese
viruses sharecross-reacting
structuralan-tigens.
Thefindings by competition
radioim-munoassay suggest thatSA8
virus and HSVo AAA
REIOAL A 0
10. a
ew
z
40 at
2040
1 2 3 4
[image:8.501.270.429.53.284.2]RECIPROCAL ANTISERADILUTION(Ioglo)
FIG. 10.
Immunoprecipitation
awsay
for
`~I-la-beledHSV-2
p40 by
unabsorbed andnucleocapsid-absorbedantisera.
Limniting
amountsof "MI-abeled
HSV-2p40 (10,0(10
cpm)
were reacted withtwofold
dilutionsof guineapig anti-HSV-2p40 serum that was either unabsorbed (0) orabsorbed witheither HSV-1 (A) orHSV-2(0) nucleocapsids.Adsorptionwasaccomplished by incubatingundiluted antiserum
withdisrupted nucleocapsids
(20
pg) for2h at37°C.The nucleocapsids were disrupted by heating at
100°C for5min in the presenceof 0.1% SDSand 1
mM DTT.
share cross-reactive
antigenic
determinants.
The
development
ofsensitive
radioimmunoas-say
procedures
for
specific HSV proteins
should
allow for
moreextensive
immunological
studies
with this
virus
group.
Utilizing
the
p40
assays
asdescribed here, for
example,
will
allow the
rapid
typing
of
newHSV isolates.
Furthermore,
asobserved in
ourpreliminary
studies with human
sera, the
useof the
p40 immunoassay
should
allow for the
accuratedetermination of
antibody
levels in human
sera toHSV-1 and HSV-2
type-specific
and
cross-reactive
antigens.
Such studies
will facilitate the determination of
antibody
lev-els in human
seraand
fluctuations which may
occur
in
these
antibody
levels
during
various
disease
statesin
which HSV has been
impli-cated. Studies
arepresently underway
toexpand
the
available
immunoassay
procedures
tootherHSV-1- and HSV-2-induced
proteins,
whichshould further enhance the
numberof
variablesamenable
tostudy.
ACKNOWLEDGMENT9
Thisworkwassupported byPublic Health Servicecontract N01-CO-75380 from the Virus CancerProgram,National Can-cerInstitute.
VOL. 29, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.501.63.215.60.322.2]42 HEILMAN ET AL.
TABLE 1. Precipitation of '25I-labeledHSVp4O by human seraa
%Precipitation of p40 from:
Human'serum HSV-1 MAL HSV-2MS
Humanb serum____________ _____
Absorbed Absorbed Absorbed Absorbed
Unabsorbed HSV-1 HSV-2 nabsorbed HSV-1 HSV-2
HSV negative
101 3.7 3.8 5.1 1.8 2.0 2.3
102 3.2 3.3 3.9 2.1 2.0 1.7
HSV-1positive
1689 74.8 19.3 69.7 49.9 4.2 3.9
1690. 77.2 17.6 66.0 44.6 5.8 5.5
1697 73.3 16.7 70.6 65.3 3.0 3.3
HSV-2positive
1691 36.4 11.5 11.7 77.1 59.1 5.1
1694 66.2 18.7 13.2 87.0 75.5 7.2
1719 39.4 12.0 12.8 92.1 76.4 5.5
HSV-1 and HSV-2positive
1696 61.0 19.0 40.2 70.6 51.8 5.4
1698 69.9 19.8 34.6 83.0 76.5 6.1
1705 38.5 11.1 27.1 40.4 28.8 3.3
a
'25I-labeled
p40
(10,000 cpm) was incubated with sera asdescribed in the text.Goat anti-humaninmmuno-globulin Gwasusedasthe second antibody. SerawereabsorbedbyincubationwithdisruptedHSV-1or HSV-2nucleocapsids (20fg)for1hat37°Cbefore the addition of'251I-labeledp40. The boldfacing indicatessignificant
precipitation of '251-labeled p40 after absorption with the putative heterologous antigen and serves as an
indicator oftype-specific reactivity.
bThe human sera wereassayedat a1:10dilution. The HSV-negative serawerecharacterized assuchby
neutralization and immunofluorescence. The anti-HSV-positive serawere obtained from K. Hsu, Columbia
University.
Wethank E. Ambush and M.Chakrabartyfortheir excel-lenttechnicalassistance.
LITERATURE CITED
1. Fish, W.W.,K.G.Mann,andC. Tanford.1969.The estimation ofpolypeptide chain molecularweights by gel filtration in6Mguanidine hydrochloride.J.Biol. Chem. 244:4989-4994.
2. Gibson,W.,and B. Roizman. 1972.Proteinsspecified
by herpes simplex virus. VIII. Characterization and
compositionofmultiple capsidformsofsubtypes1and 2.J.Virol. 10:1044-1052.
3. Greenwood,F.C.,W. M.Hunter, and J. S. Glover. 1973. The preparation of "3LI-labeled human growth hormone of high specific activity. Biochem. J. 89:
114-123.
4. Honess, R.W., K. LPowell,D.J.Robinson, C. Sim,
andD. H.Watson.1974.Typespecificand type com-monantigens in cells infected with herpes simplex virus type 1 and on the surfaces of naked and enveloped
particlesof the virus. J.Gen. Virol. 22:159-169. 5. Honess, R.W.,andB.Roizman.1973.Proteinsspecified
by herpes simplex virus. XI. Identification and relative molar ratesofsynthesisofstructural andnonstructural herpes viruspolypeptides in the infected cell. J. Virol. 12:1347-1365.
6. Hull,R. N. 1968. Thesimian viruses.Virol. Monogr. 2: 1-66.
7. Laemmli, U. K. 1970. Cleavage ofstructural proteins during theassembly of the head of bacteriophage T4. Nature(London)227:680-684.
8. Malherbe,H.,andR.Harwin. 1958. Neurotropic virus inAfricanmonkeys.Lancetii:530.
9. Melnick,J.L.,R.J.Courtney, K. L Powell, P. A.
Schaffer,M.Benyesh-Melnick,G. R. Dreesman, T.
Anzai,and E. Adam. 1976. Studies on herpessimplex
virus andcancer. Cancer Res. 36:845-856.
10.Oroszlan,S.,R. J.Huebner,and R. V. Gilden.1971.
Species-specific and interspecies antigenic determi-nantsassociated with the structural protein of feline C-type virus. Proc.Natl. Acad.Sci. U.S.A. 68:901-904. 11. Perdue,M.L.,J.C.Cohen,M.C.Kemp, C. C. Randall,
and D. J.O'Callaghan.1975.Characterization ofthree species ofnucleocapsids ofequine herpesvirus type 1 (EHV-1). Virology64:187-204.
12. Perdue,M.L.,M.C.Kemp,C. C.Randall,and D. J.
O'Callaghan.1974.Studies of themolecular anatomy of L-Mcellstrainofequineherpesvirus type1:proteins of the nucleocapsid and intact virion. Virology 69:
201-216.
13. Powell,K.L.,and R. J.Courtney. 1975.Polypeptides
synthesized in herpes simplex virus type 2-infected HEp-2cells.Virology66:217-228.
14. Powell,K.L., andD.H. Watson.1975.Somestructural antigens of herpessimplex virus type 1. J. Gen. Virol.
29:167-178.
15.Schneweis, K. E., andA. J.Nahmias.1971.Antigens ofherpessimplexvirustype1and2-immunodiffusion
and inhibition passive haemagglutination studies. Z. Immunitaetsforsch. Exp.Ther.141:471-487. 16. Stevens,D.A.,T.Pincus,M. A. K.Burroughs, and
B. Hampar. 1968. Serologic relationship of simian herpsevirus (SA8) and herpes simplex virus: heterogen-icityinthe degreeofreciprocalcross-reactivity shown by rabbit 7S and 19S antibodies. J. Immunol. 101: 979-983.