0022-538X/81/050497-07$02.00/0 Vol.38,No.2
Specialized Transduction
with Xplac5: Dependence
onrecA
and
on
Configuration
of
lac
and
attX
RONALD D. PORTER,l MICHAEL W. LARK,'tAND K.BROOKS LOW2
Departmentof Microbiology, Cell Biology,Biochemistry and Biophysics,The PennsylvaniaState
University, University Park, Pennsylvania16802,1andRadiobiologyLaboratories, Yale University School
ofMedicine, New Haven, Connecticut 065102
Received7November 1980/Accepted13January1981
The constructionof Xplac5 transducingphages carrying various lacZ allelesis
described. Genetically disabled(N N-P) Xplactransducing phageswereused
tostudythe dependenceof specialized transductiononhost RecA functionand
onthe location ofthe lacZgene in the recipientstrain. In the absence of
site-specific recombination at attX, transductionwascompletely dependentonhost RecAfunction. Regardlessofthe configuration ofattX,Xplac transducing phages recombinedata20-to50-foldhigher frequencywithF421acthan withalacgene
located in the cellular chromosome. Deletionmutants of lacZ in the recipient strainwereusedtoshowthattheprobability of lac recombinationresulting from Aplac infection is apparently proportionaltothe amount ofhomology between theparental lacZgenes.
Specialized transduction is a mode of gene transfer(2, 9)whichhas beenusedinfrequently inrecombination studies. We recently described asystemforthestudyofrecombination kinetics thatemploys Aplac5 transduction (13), and here we report a more detailed characterization of the Aplac5 transduction system. The isolation and characterization ofAplac derivatives con-taining specificlacZ alleles isdescribed. Wealso report the construction of
genetically
disabledAplac
transducing phages
and their use to ex-amine thedependence
oftransductionon:(i)
the hostrecAgene, (ii)thelocation of the lacZallele in therecipient
strain,
and(iii)
theconfiguration
ofthe attA site in the genome of the recipient cell. The randomness with which recombination can occur along the lacZ gene in specialized transduction is evaluated.
MATERIALS AND METHODS Bacterial strains. TheEscherichiacoli K-12 de-rivatives used in thisstudyarelisted in Table1.The complete pedigreeofall strains isavailableonrequest. Bacteriophage. The bacteriophage lambda and Aplac5derivatives used in thisstudywere prepared from thelysogenicstrains shown in Table 1.
Media. LB medium(11)wasusedforliquidculture medium andagarplateswheneverarich mediumwas required. Modified minimal medium56and its supple-mentationtoproduceselective media have been de-scribed (8). Half-strengthmodified minimal medium 56 (56/2) was used for the dilution and washing of cells. Dilutions ofphagestocksweremadeinAbuffer tPresent address: Department of Microbiology, Case Westem ReserveUniversity,Cleveland,OH44106.
(0.006 M Tris, pH 7.2, 0.01 M MgSO4, and 0.005% Difcogelatin). Top agar was either0.6%or 0.8%Difco agar indeionized water.
Lysate production. Lysates of bacteriophage carrying theXcI857allelewereprepared byheat in-duction ofanappropriate lysogenicbacterial strain. Lysates of AcI+orAi21phagewerepreparedby induc-tion withUVirradiation ofappropriate lysogenic bac-terialstrains.
Preparationoflysogens. Lysogensforparticular bacteriophage derivatives werepreparedbyspotting alysateontoorstabbingaplaqueintoalawn of the desired host strain and then screeningcolonies ob-tainedfrom theareaofclearingfor the presence of the bacteriophageas aprophage.
Plating ofbacteriophage for plaques. Phage stockstobeplatedforplaqueswerediluted in A buffer andplated on anappropriateindicator strain which had been grown to 109 cells per ml in LB medium supplementedto0.2%maltose and 0.01 MMgSO4.
Testing plaques for the Lac character of the phage.PlatingsofAplaconA(lac-pro)strainsonLB plateswereusedto assessthelacZ+orlacZ character ofindividualplaquesorpopulationsofplaques.Phage platings that had been incubated for10 to 16hat37°C wereflooded with 0.013 M
o-nitrophenol-fi-D-galacto-pyranoside (ONPG) in 0.25 M potassium phosphate buffer(pH7.25).PlaquesfromAplacZ+phagegavea strongyellowcolorinlessthan1minat room temper-ature. Plaques fromAvir or AplacZ gave no yellow color aftermorethan2hof incubationat room tem-perature. Platings containing both
AplacZ'
and AplacZhadto be evaluated within approximately 5 min because theyellowcolor from AplacZ' plaques diffused rapidly across the plate. Stabs of AplacZ plaques from mixedpopulationsintoafresh lawn of cells demonstrated no yellow color after prolonged room-temperature incubation with ONPG. XplacZ497
on November 10, 2019 by guest
http://jvi.asm.org/
498 PORTER, LARK, AND LOW
TABLE 1. E. coli K-12strains
Relevantcharacteristics F-A(lac-pro) supFtrppyrFhisrpsLthi A-RDP100(AcI857Sam7plac5I ZV Y-) RDP100(AcI857Sam7plac5 I Z118Y-) F-A(lac-pro)XlllsupG rpsLthi A-RDP100(Xi21 Pam8O)
C600(AcI857ind Nam7Nam53)
RDP100(XcI857Sam7Nam7Nam53 plac5 I- Z+ Y-) RDP 100(AcI857Sam7Nam7Nam53plac5 I Z118Y-) RDP 100(XcI857Nam7Nam53Pam8Oplac5 I Z+ Y-) RDP 100(XcI857 Nam7Nam53Pam8Oplac5 I Z118Y-)
F-lacZ813 lacI3promettiprpsLthi
A-KL761butrecAl
F-lacZ813 lacI3metA(gal-attA-bio)rpsLthi A-KL763but recAl
F-lacZ813 lacI3promethis trprpsLthi(Xind) KL765 but recAl
F421acZ813lacI3/A(lac-pro)mettrprpsLthi A-KL767 but recAl
F421acZ813lacI3/KL773 KL769 but recAl
F42lacZ813lacI3/A(lac-pro)methistrprpsLthi(Aind) KL771but recAl
KL773 F-A(lac-pro)A(gal-attX-bio) metrpsLthi A-RDP 100 F-A(lac-pro) X1llleuthiacrA(?) supE44 RDP101 F-A(lac-pro)XlllleuthisupE44 RDP 103 F42iacZ118lacI3(KL773)
RDP104 F-lacZ118 lacI3A(gal-attA-bio)metrpsL thi RDP105 F1281acZAH111proA',B I/KL773
RDP 106 F1281acZAH119proA+,B+/KL773 RDP 107 F1281acZAH114proA+,B+/KL773 RDP 108 F1281acZAH145proA+,B+/KL773 RDP 109 F1281acZAH125proA+,B /KL773 RDP110 F1281acZAH138proA+,B+/KL773 RDP111 F1281acZAH220proA+,B /KL773
a Strr,Streptomycin resistance.
Source,reference,comments CSH28(11) cured of F' Phage fromCSH66 (11) Seetext
Derived from BW113 (8)xX7151 (from J. Miller) and
sponta-neousStrr'
Phage from L.Herskowitz
Phage from C. M. Radding Seetext
Seetext Seetext Seetext
All of these strains are lacZ813-ochre (21) derivatives of KL318 (1). In eachcasethe finalRec+/1 recAl pairwasmade bya mat-ing with MA1079, which is a
serAderivative (W. K. Maas) of KL16-99 (7).
From KL320 (1)
C600derivatives with A(lac-pro) Xlllderived from PK191 (8).
FromKL334(1)xKL773 FromCHS14xKL773 FromCSH15xKL773
FromCSH16xKL773 FromCSH17xKL773 FromCSH18xKL773 FromCSH19xKL773 FromCSH20xKL773
phage containingnonsensemutations in the lacZgene could be readily detected by using an appropriate
suppressorstrain for plating and looking for yellow
colorwith ONPG.
IsolationofAplac derivatives containing par-ticular lacZalleles. Strains containingadeletionof thegal-attX-bio region of the chromosome and pos-sessing the desired lacZ alleleonthe chromosomeor
on anFlacwerelysogenized with 'placZ'asdescribed
above.These lysogenswereusedto preparelysates,
andtheresultingphagewereplatedon anappropriate strainfortestingtheLaccharacter oftheplaques with ONPGflooding. Plaques thatwereLac- werestabbed
intofresh lawns forretesting, andlysogenswere pre-paredforthosethattested Lac- twice.
Lysatesfromthenewly obtained
~placZ
lysogens weretested with ONPGafter platingonappropriatesuppressorandnonsuppressorstrainswhen lacZ
non-sense mutationswere involved. Presumptive XplacZ
phagewerealso tested by transducingstrains contain-ing thesamelacZ alleleaswell asstrainscontaining other lacZalleles(see below).
Phagecrosses.Phagecrosses weredoneby infect-ingasuitablehost strainat2x 10i cellsperml in LB mediumsupplemented to0.2% maltose and 0.01 M MgSO4withbothparental phagesatamultiplicity of Strain
KL528 KL550 KL551 KL754
KL755 KL756 KL757 KL758 KL759 KL760
KL761
KL762 KL763 KL764 KL765 KL766 KL767 KL768 KL769 KL770 KL771 KL772
on November 10, 2019 by guest
http://jvi.asm.org/
infection (MOI) of5.The plaques from these lysates weretestedasdescribed above, and the desired recom-binantphageswererecovered fromstabsaslysogens. Production of disabled AplaZ transducing
agents. The original \placZisolatesarecapable of lysing cellsthatthey infect,andthiscomplicates trans-ductionstudieswhenpotential transductantsarelost. To avoid thelossof potentialtransductants by phage-mediatedcelldeath,disabledversionsof Aplac5were made. TheNam7Nam53 markerswereintroduced by crossing
AplacZ+
with AcI857 indNam7Nam53 and scoring for lacZ -containing phage that required anambersuppressorin the host strainforgrowth.The resulting AplacZ+ Nam7Nam53 phage was then crossed with XplacZ118 to give a lacZ phage that requiredan ambersuppressorin the host strainfor growth.XplacZ+Nam7Nam53wasthen crossed with Ai2M Pam8OtoyieldalacZ+-containingphage which specificallyrequired supEforgrowth (Nam7Nam53is suppressed by either supEorsupF, whereas Pam8Ois
suppressed by supE butnotsupF)and whichfailedto complementeitheraA NaNamam53 phageortheAi21
Pam8O phage when the two types were spotted together on a sup+ strain. The resulting AplacZ+ Nam7Nam53Pam80 phage was then crossed with AplacZ118 Nam7Nam53 toobtainAplacZ118 Nam7-Nam53Pam80. Thesetransducing phages containing both Nam7Nam53 and Pam8O alleles produced no reductioninviable cell number whentheywereused to infect sup+ strains, andthey were used for the transductionexperimentstobedescribed below.
Transductionprocedure.Therecipientcells were
grownto2x10i cells perml in LB medium supple-mentedto0.2%maltoseand0.01MMgSO4at370C. Thetransducingphage preparationwasaddedatthe desired MOIasindicatedforindividual experiments,
and incubation was continued at 37°C for 30 min. Dilutionsweremade with56/2medium,andplatings were doneonminimal lactoseselectionplates which
were incubated at the desired temperature for the developmentofcolonies.
Chemicals and media. Tryptone, yeast extract, andagarwereobtainedfromDifco. OPNG andsugars were obtained from Sigma Chemical Co. All other
chemicalswerereagentgrade. RESULTS
Isolation and characterization ofAplc5 derivativescarryinglacZalleles. Our efforts toconstruct Xplac5derivativescontaininglacZ
alleleshave been concentrated on lacZ118and
lacZ813-ochre because of their suitability for recombination studies (13). The methodology, which is similartothetechniquesusedbyothers (5, 10, 14),is also suitable for other lacZalleles, however,andwehave alsoemployedit to obtain
aderivative of Xplac5carryingthe lacZ36allele. Strains carrying a deletion of thegal-attA-bio
regionof the chromosomeandoneof these lacZ
alleleseitheronthe chromosome oron F421ac werelysogenizedwithAplac5 containing lacZ+.
Such lysogens were characterized as
Lac'
andAplac5 TRANSDUCTION 499
temperature sensitive at420C since the phage carries the AcI857 marker. The lysogens ob-tained segregated Lac-, temperature-resistant colonies at a variable rate (1 to 50%). Phage lysateswereprepared and platedonKL528, and
the plaques were scored for lacZ+ or lacZ by
flooding with ONPG as described in Materials
and Methods. Theplaquescontainingthe
origi-nal lacZ+ version of
?plac5
developed yellowcolor very rapidly, whereas those that had
pickedupthe lacZallele showednoyellow color.
In thecasesoflacZ118 andlacZ813-ochre,we
examined 1,000 plaques from each ofatleast 10 independent isolates containing the lacZallele
inthe chromosome and 10 independent isolates
containingthelacZalleleonF421ac.The results areshown in Table2.
Characterization of
ApkacZ
isolates. Rep-resentative plaques that appeared Lac- when platedonKL528and flooded with ONPGweretransferredtofresh lawns of KL754 andKL528 withsteriletoothpicks and retested with ONPG after 12 h of incubationat370C. Both lacZ118 and lacZ813-ochrearesuppressible by the supG
allele of KL754, and the stabs of phage carrying lacZ118orlacZ813-ochreproducedyellowcolor within 5min of flooding with ONPG. This result
serves to demonstrate the sensitivity of the ONPGflooding method for detecting fl-galacto-sidase since the efficiency of suppression of lacZ813-ochre bysupGis only about 1% (R. D. Porter,unpublished data).
Lysogens of selected XplacZ118 and AplacZ813-ochre isolates were prepared as de-scribed in Materialsand Methods.Lysateswere thenprepared and platedonboth KL528(supF) and KL754(supG) for testing with ONPG. With both of thesephage isolates, the plaquesgave no
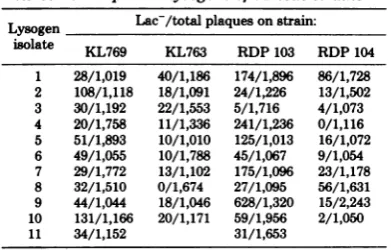
TABLE 2. Lac plaquesobtainedfromlysatesof AcI857Sam7placZ+lysogensofvariousstrains'
Lysogen Lac/totalplaques on strain:
isolate KL769 KL763 RDP 103 RDP 104 1 28/1,019 40/1,186 174/1,896 86/1,728 2 108/1,118 18/1,091 24/1,226 13/1,502 3 30/1,192 22/1,553 5/1,716 4/1,073
4 20/1,758 11/1,336 241/1,236 0/1,116 5 51/1,893 10/1,010 125/1,013 16/1,072 6 49/1,055 10/1,788 45/1,067 9/1,054 7 29/1,772 13/1,102 175/1,096 23/1,178
8 32/1,510 0/1,674 27/1,095 56/1,631
9 44/1,044 18/1,046 628/1,320 15/2,243 10 131/1,166 20/1,171 59/1,956 2/1,050
11 34/1,152 31/1,653
aAcI857Sam7placZ lysogensof the indicatedstrainswere
prepared using phage preparedfrom KL550. Lysates from
each lysogenwereplated onKL528, andtheplatingswere flooded with ONPG afterapproximately12h of incubationat 37°C. The numbers given represent Lac- plaques/total plaques.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.500.258.454.486.611.2]yellow color when KL528platingswereflooded
withONPG,and theplaques yieldedauniform
level of yellow color when KL754 platingswere
tested.Theselysateswerealso usedin transduc-tionexperiments with recipientstrainscarrying the same lacZ allele as the transducing phage
and recipient strains carrying the other lacZ allele. In theselfingexperiments where both the transducingphage and the recipientstrain
car-ried the same lacZ allele,
Lac'
colonies wereobservedatonly thesamelevelascontrol
plat-ings for spontaneous revertants (see Table 3). When the transducing phage and therecipient strain carried thetwodifferent lacZalleles,
Lac'
transductants were observed at levels several orders of magnitude greater than spontaneous reversion. The details of transduction experi-mentsaregiven below.
Dependence of Aplac5 transduction on
recA,attA,andthelocation of therecipient lacZ gene. We have previously reported that Aplac5transducing phage recombine more effi-cientlywithF421acorF1281acthan withalacZ
genelocatedonthechromosome,andthat lacZ
x lacZ recombination is totally dependent on
J. VIROL.
RecAfunctionin thetransduction system(13). Inthosestudies, nondisabled Aplac transducing
phages
wereused andaresidentAindprophage
in the recipient strain was used to control the
lytic
functions of the transducing phage. We have now used thedisabled transducing phagetomakeamoredetailedanalysisofthe
param-etersaffectingrecombinationfrequencyin
Xplac
transduction.
Aseries of strainswasconstructed to testthe importance of: (i) FlacZ813-ochre/Alac versus
F-lacZ813-ochre as the recipient DNA struc-ture, (ii) recA+ orrecA, and (iii) anormal attA
siteasopposedtoaXindprophageor a
gal-attX-biodeletion.Strainscontainingallcombinations
of these parameters were transduced with
XplacZ118
Nam7Nam53Pam80 andXplacZ'
Nam7Nam53Pam80 atan MOI of2,and plat-ings on miniimal lactose selection plates wereincubated at 37°C. The results of these trans-duction experiments are shown in Table 3. It
canbe seenthat lacZx lacZ recombination is totally dependent on recA+, since only
back-ground reversion levels of
Lac'
colonies werepresentwhen recAstrainswereusedas
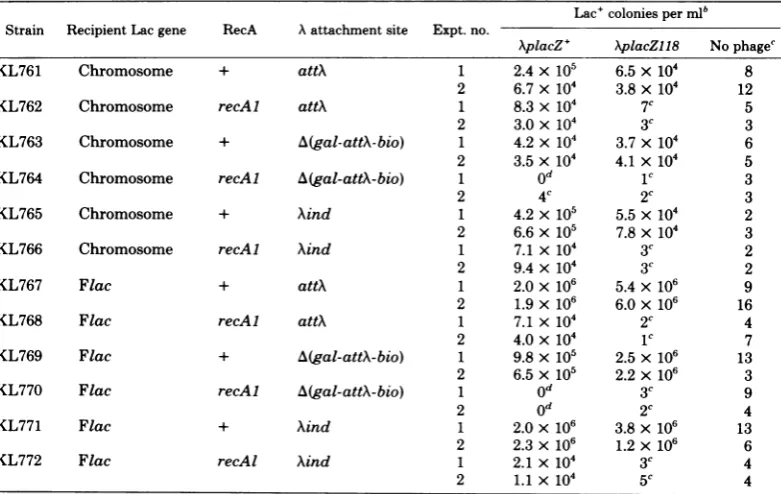
recipi-TABLE 3. Dependence ofXplactransductiononseveralparametersa
Lac'colonies perml' Strain RecipientLacgene RecA Aattachment site Expt.no.
AplacZ+
XplacZ1l8 NophagecKL761 Chromosome + attA 1 2.4x 105 6.5x 104 8
2 6.7x 104 3.8x104 12
KL762 Chromosome recAl attA 1 8.3x 104 7C 5
2 3.0 x 104 3C 3
KL763 Chromosome + A(gal-attA-bio) 1 4.2 x104 3.7x104 6
2 3.5 x104 4.1x 104 5
KL764 Chromosome recAl A(gal-attA-bio) 1 od 1c 3
2 4c 2c 3
KL765 Chromosome + Aind 1 4.2x105 5.5 x 104 2
2 6.6 x105 7.8 x104 3
KL766 Chromosome recAl Aind 1 7.1x 104 3c 2
2 9.4 x104 3c 2
KL767 Flac + attA 1 2.0x106 5.4x 106 9
2 1.9x 106 6.0x 106 16
KL768 Flac recAl attA 1 7.1 x104 2c 4
2 4.0x104 1c 7
KL769 Flac + A(gal-attA-bio) 1 9.8 x 105 2.5x
105
132 6.5x 105 2.2x106 3
KL770 Flac recAl A(gal-attA-bio) 1 od 3c 9
2 o0 2c 4
KL771 Flac + Aind 1 2.0x 106 3.8x106 13
2 2.3x 106 1.2x106 6
KL772 Flac recAl Aind 1 2.1x 104 3C 4
2 1.1x 104
5'
4a Theindicated strainsweretransduced with
XcI857
Nam7Nam53Pam8O
placZ+ from KL759 andAcI857 Nam7Nam53Pam80 placZ118 from KL760 as described in the text at an MOI of 2. The minimal lactose selectionplateswereincubatedat 37°C for 48 to 72 h beforecolonieswerecounted.Allrecipientstrainscarry lacZ813-ochre(21).bExceptasindicated.
cPer0.2ml.
dPer
0.1ml.on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.500.64.455.351.598.2]Aplac5 TRANSDUCTION 501
entsintransduction. When
AplacZ'
transducing phagewere used withrecA recipients thatcon-tainanormalattXoraXindprophage, significant
levels of
Lac'
transductants were seen. TheseLac'
transductants didnotappearinarecAcellcontaining the gal-attX-bio deletion, indicating thatthistypeoftransduction is totally depend-entonsite-specific recombination of the
trans-ducingphageat attX(3, 4).
Therewas astriking effect of location of the
recipient lac region on Lac+ transduction
fre-quency.A 20-to50-fold-higher transduction
fre-quency for Flac-carrying strains was
ob-served,comparedtoF-lac, in thecaseswithattX
orA(gal-attX-bio). In the Xind strains this
dif-ferential holds for transduction withAplacZ118, but the effect appears smaller (only fourfold)
with XplacZ+. However, this difference with
XplacZ'
andAplacZ118 hinges on therate ob-served in recA+ cells containing the residentprophage where
Lac'
colonies can arise fromgeneral recombination between
\placZ'
and the residentprophage but notfromrecombination betweenXplacZ118
and theresidentprophage. Thisenhanced recombination of\placwith Flac isnotdue toanyfunctionssuppliedintransby the F factor since the presence of anon-lac-containing F factor (either in the autonomous stateasF15thyorintegrated into thegenomeas
a Cavalli Hfr) had no effect on the level of
recombination observed between Aplac and a
chromosomal lac allele (13).
Transduction of kacZ deletions with
AXpkcZ+.
Aplac5 carries the entire lacZ geneand part of the lacIgeneaswellaspartof the
lacYgene.ThelacZgenemakesupthe bulk of the lac material carried by Xplac5 since it is about 3,700 base pairsout of about 4,200 base pairs of total Lac DNA (15). To rule out the possibility of recombinational hotspots in our
transduction assay, we wanted to determine whether ornot recombinationcanoccurfairly
randomly across the Lacregion carried by the
transducing phage.To test thispossibility, trans-duction experiments were done withrecipients
thatcarrieddeletionsstartingatapoint beyond
the lacIgene and extendingvarious distances intothelacZgene(11).ThelacZ deletionswere
carried on F1281ac-pro in a A(lac-pro) strain
containingadeletion of thegal-attA-bioregion.
These strains were transduced with
XplacZ'
Nam7Nam53Pam8O at an MOI of 2, and the lactoseselectionplateswereincubatedat370C. The results of these experiments are shown in
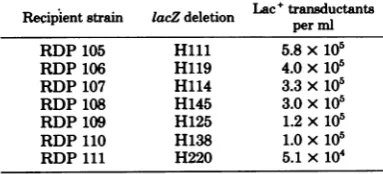
Table 4. The level of
Lac'
transductants ob-tainedisroughlyproportionaltothe amount of lacZ material remaining in the recipient F128 episomes and indicates that recombination isreasonably randomacrossthe Lacgenematerial carried by
Xplac5.
DISCUSSION
We have described indetailour version ofa
technique for transferring and lacZ allele onto
Xplac5
by usingattXdeletionstrains containing the desired lacZ allele. Although the frequency of XplacZ phage obtained varies considerably from isolate to isolate (Table 2), the method works well for thetwolacZalleles testedexten-sively and also works for other lacZalleles (un-published data). For reasons that are not
en-tirelyclear,thechancesofobtaining thedesired lacZ alleleonAplac is three-tofivefold better if the lacZ allele islocated onF421acrather than onthecellulargenome (seeTable 2). Thismay
berelated, however, tothe finding that plac5 initially recombines more readily with F421ac
than withachromosomal lacgene (see Table 3
and reference 13). The mechanism for generat-ing \placZ phage presumably involves
recom-bination between the two tandem lac regions surrounding the prophage inanaddition trans-ductant. TheuseofattXdeletionsgreatly facili-tatesfinding thenecessary
Lac',
prophage-con-taining,addition transductantssinceanyphage
containing transductantsmustnecessarily have theprophage locatedinthe lacregion. It should be noted that when theXplac5 phagewas
inte-gratedat attX,wedidnotfindphage containing
the desired lacZ allele after the examination of
over25,000 plaques fromanappropriate lysate.
Theconstructionofgenetically disabled Aplac transducing phages has facilitated analysis of those factors important in determining trans-duction frequency. XcI857 Nam7Nam53 phage establishlysogenyasplasmidsinnonsuppressing
strains(6, 18) and resultinlittlecelldeath. The
TABLE 4. TransductionoflacZdeletionstrainsa
Lac+transductants
Recipientstrain lacZdeletion
peracn
per mlRDP105 Hlll 5.8 x 105
RDP106 H119 4.0x 105
RDP107 H114 3.3x 105
RDP108 H145 3.0x105
RDP109 H125 1.2x105
RDP110 H138 1.0 x105
RDP111 H220 5.1 x104
aThe indicated strainsweretransducedwith
AcI857
Nam7Nam53Pam80placZ'preparedfromKL759at an MOI of2 as described in the text. The lactose selectionplateswereincubatedat37°C.Thedeletions have beendescribed(11) andinvolve the deletion of increasingamountsof the lacZgene carriedbyF128 startingfrom Hlll up to H220, whichhas the least amountoflacZDNAremaining.
VOL. 38,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.500.257.449.496.583.2]502
PORTER, LARK, LOWadditional inclusion of the X Pam8Oallele pre-ventsthe establishment of the
plasmid
stateand providesanonreplicating
DNAmolecule which cannotcontributetoLac' colony
formation un-less the transducing phage DNA recombines withthe cellularDNAbefore it is lostby
segre-gation (R. D. Porter,unpublished data).
The data in Table3confirm ourprevious
result that Xplactransducing phages
recombinemore read-ily with F421ac than with a chromosomal lac gene (13), and thesenewdata showthat this is the caseregardless
oftheconfiguration
of the attXregion of the chromosome. The transduc-tants seenwithAplacZ'
in a cell that is recA when the cell contains anormal attA site or a Xind prophage wouldappear tobe the result of site-specific recombinationatattAoratthehy-brid attachment sites of the
prophage,
since strains thatarebothrecA andA(gal-attX-bio)
(KL764 andKL770) failto yieldany transduc-tantswith
XplacZV.
Thisclass of transductants probably arises because thetransducing
phage
can
lysogenize
at attA or atthehybrid
attach-mentsitesat alowefficiency
as aresult of Xint gene expressionfrom thep intpromoter(16). It would betheoretically
possible to obtainLac'
transductants by
site-specific
recombination at secondary X attachment sitesevenwhenattX
is deleted (17), but suchintegration
ofthe trans-ducingphageatsecondary attachment sitesap-parently
does notoccur at significant levels in the present experiments. Similar results dem-onstrating a RecA dependence for plus-times-minus allele recombination have also been re-ported for\pro
and Agal (19, 20). One of these reports indicated low but significant levels of transduction mediated byXred functions
in a recA strain withXgal
(19). We conclude from this thatXred
functions are not significantly expressed inourdisabled transducingphages.Withonenotable exception (12), minus-times-minustransductional crosses have notbeen uti-lized for recombination studies in previous re-ports. The data in Table3indicatethatlacZx lacZtransductional crosses aretotally depend-enton host RecAgenefunctioneven when site-specific recombination at
attA
ispermitted. The data in Table 3 also point up the unexpected resultthatlacZ x lacZ crosses yield more Lac' recombinants than lacZ+ x lacZ crosses when the recipient lacZ allele is located on F421ac, but not when therecipientlacZallele
islocated onthechromosome.We have noexplanationfor this observation, but this phenomenon may pointup someaspectof apossible mechanistic differencein the recombination betweenAplac and F421acversus F-lac.We have also used Aplac transduction as a measureof theabilityofrecombinationtotake
J. VIROL.
place fairly randomly along the length of the recombining lacZ genes, or at least in rough proportiontothelengthofhomologypresent. A disabled
AplacZ'
transducing phage lysate pre-paredfrom KL759 was used to transduce a series of strainscontainingdeletions of variouslengths in the resident lacZgene. The datain Table 4 indicate that the level oftransductantsobtained is roughly proportional to the amount of the lacZ generemaining
in therecipient
strain. Hence,therecombinationseenbetween Aplac5 derivatives and lacZgenesin recipientstrains appearsnot to involvespecific sites in thelacZ gene.The Aplac transduction system has several advantages over previously utilized specialized transductionsystems.Numerous well-character-ized lacZallelesareavailable andcanbeplaced on the transducing phages. The separation of the lacregion from attXonthecellulargenome allows theuseof strains deleted for
attX,
which ismoredifficult withAgal
transduction. Theuse ofattA deletions facilitates theanalysis of gen-eralversussite-specific recombination in lacZ+ x lacZtransductions. Theuseof thegenetically disabledplaque-forming
transducing phages
de-scribed herein also allows transduction experi-ments to be done without thekilling
of cells in the recipient culture and without thepresence ofhelperphage. Thissystemshouldproveuseful for further studies of recombination mecha-nisms.ACKNOWLEDGMENTS
We thank TomMcLaughlin,TomWitkowski,and Susan Lambert for their expert technicalassistance, and J. Beckwith forhelpfuldiscussion.
This work wassupportedby Public Health Service grants CA-06519 (to K.B.L.) and GM-26422 (to R.D.P.) from the National Institutes of Health andbyNationalScience Foun-dation grant PCM-73-01609(to K.B.L.).
LITERATURE CITED
1. Birge,E.A., and K. B.Low. 1974.Detection of
tran-scribablerecombinationproductsfollowing conjugation inRec+,RecB- and RecC- strains ofEscherichia coli K12. J.Mol. Biol. 83:447-457.
2. Campbell,A. 1964.Transduction, p. 49-85. In I. C. Gun-salasand R. T. Stanier (ed.), The bacteria, vol. 5. AcademicPress, New York.
3. Echols,H.,R.Gingery,and L.Moore.1968.Integrative recombination functions ofbacteriophageA;evidence forasite-specific recombination enzyme. J. Mol. Biol. 34:251-260.
4.Gottesman,M.E., and M. B. Yarmolinsky. 1968.
In-tegrationnegative mutants ofbacteriophage lambda. J. Mol. Biol. 31:487-505.
5. Jordan,E.,H.Saedler,and P.Starlinger.1968.0° and strong-polar mutations in the gal operon are insertions. Mol. Gen. Genet. 102:353-363.
6. Lieb,M. 1970.A mutants whichpersist asplasmids.J. Virol. 6:218-225.
7. Low,B.1968.Formation ofmerodiploidsinmatingswith
aclass of Rec- recipient strains of Escherichia coli
K-12.Proc. Natl. Acad. Sci. U.S.A.60:160-167.
on November 10, 2019 by guest
http://jvi.asm.org/
8. Low, B. 1973. Rapid mappingofconditionaland
auxo-trophic mutations in Escherichia coli K-12. J.Bacteriol. 113:798-812.
9. Low, K. B., and R. D. Porter. 1978. Modes ofgene
transfer and recombination in bacteria. Annu. Rev. Genet. 12:249-287.
10. Malamy,M.,H.Fiandt,and W.Szybalski.1972.
Elec-tronmicroscopy of polarinsertions in the lacoperonof Escherichia coli.Mol.Gen.Genet. 119:207-222.
11. Miller, J. H. 1972.Experiments inmolecular genetics.
Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
12.Morse, M. L., E. M.Lederberg, and J. Lederberg.
1956.TransductionalheterogenotesinEscherichia coli. Genetics41:758-779.
13.Porter, R. D.,T.McLaughlin,and B.Low.1978.
Trans-duction versus"conjuduction": evidence for multiple roles forexonuclease V in genetic recombination in Escherichiacoli. Cold SpringHarbor Symp. Quant. Biol. 43:1043-1047.
14. Shapiro, J. A.1969.Mutations causedbythe insertion of
genetic material into thegalactoseoperon of Esche-richia coli.J. Mol.Biol.40:93-105.
TRANSDUCTION 503
15. Shapiro,J., L MacHattie, L Eron, G. Ihler, K. Ippen, and J.Beckwith. 1969. Isolation of pure lac operon DNA.Nature(London) 224:768-774.
16. Shimada, K., and A.Campbell. 1974.Int-constitutive mutants ofbacteriophage lambda. Proc. Natl. Acad. Sci.U.S.A. 71:237-241.
17. Shimada, K.,R. A. Weisberg, and M. E. Gottesman. 1972.Prophage lambda at unusual chromosomal loca-tions. I.Location of thesecondary attachment sites and theproperties of thelysogens.J.Mol.Biol. 63:483-503. 18. Signer, E. R. 1969. Plasmidformation: a new mode of
lysogeny by phage A.Nature(London) 223:158-160. 19. Wackernagel, W., and C. M. Radding. 1974.
Transfor-mationandtransduction of Escherichia coli: the nature of recombinants formed byRec, RecF,andARed, p. 111-122.In R. F. Grell(ed.), Mechanisms in recombi-nation. PlenumPress,NewYork.
20. Weisberg, R. A., and N.Sternberg. 1974. Transduction ofrecBW hosts ispromotedbyAred function,p. 107-109.InR. F. Grell(ed.),Mechanims in recombination. PlenumPress, New York.
21.Zipser, D. 1967. Orientation of nonsense codons on the genetic map of the Lac operon.Science157:1176-1177.