JOURNAL OF VIROLOGY, July1979, p. 42-52 Vol.31, No. 1 0022-538X/79/07-0042/1 1$02.00/0
Separation
and Characterization of
Herpes
Simplex
Virus
Type 1
Immediate-Early
mRNA's
ROGER J.WATSON,t CHRISTOPHER M. PRESTON,ANDJ. BARKLIE CLEMENTS* Departmentof Virologyand Medical Research CouncilVirology Unit, University of Glasgow,Church
Street, Glasgow Gll 5JR, Scotland
Polyadenylated immediate-early transcripts of herpes simplex virus type 1,
made in BHK cells infected and maintained in the presence ofcycloheximide,
have been separated on denaturing agarose gels containing methyl mercuric
hydroxide. Threevirus-specific mRNA bands of estimated sizes 4.7, 3.0, and 2.0
kilobases (kb) were detected, and these mRNA's were mapped on the virus genome and also used to direct protein synthesis in vitro. The 4.7- and 3.0-kb mRNA'shybridizedpredominantly tocertain DNAfragmentswhicharelocated in the shortandlong repetitive regionsof the genome,respectively,whereas the 2.0-kb mRNA's mapped to three discrete regions of the virus DNA. In vitro
translationof theseseparatedmRNA sizeclasses indicated that the 3.0-kb mRNA
specifiedthesynthesisofviruspolypeptideVmw 110,whereas the 2.0-kb mRNA's
specified Vmw 68, 63, and 12. The synthesis of small amounts ofVmw 175 was
specified bythe 4.7-kb mRNA. Incontrastwiththe mRNA's whichspecify these
otherimmediate-early polypeptides, that specifyingVmw 12 is much larger than required for itscodingsequences.
The locations on the genome of transcripts
which accumulate in cells infected with herpes
simplex virus type 1 (HSV-1) at various times
postinfection,in the presenceorabsence of DNA
or protein synthesis inhibitors, have been
ana-lyzedbyhybridizationtothefragmentsof
HSV-1DNAgenerated by digestion withanumber of restrictionendonucleases (3, 10). These, and pre-vious studies (6, 22), have indicated that
tran-scription of the HSV-1 genome is subject to
temporalregulation.
Inhibition of virus protein synthesis with cy-cloheximidewasfoundtoresult inaccumulation ofRNA sequences which hybridized to certain
DNAfragmentsonly (3, 10), and these restricted
RNA sequences, transcribedbyapreexisting
a-amanitin-sensitive host RNA polymerase (4),
have been defined as immediate-early (IE) RNA. IE RNAsaccumulate also in cells infected at the nonpermissive temperature with certain HSV-1 temperature-sensitive (ts) mutants, and aretheonlyvirus transcripts detectable in ts K-infected cells (24).Release of the cycloheximide block (3), orshift-down ofcells infected with ts Ktothepermissive temperature (24), was found toallowtranscription from additional regions of HSV-1 DNA which map throughout the virus genome. These latter regions were also tran-scribed during the normal replicative cycle at early and late times postinfection (3), that is,
t Present address: Laboratory of Molecular Virology, Na-tionalCancerInstitute, Bethesda,Md. 20205.
before and after the onset ofvirus DNA
repli-cation.
The polypeptides specified by IE RNA, the
IEpolypeptides, formasubset of the virus
pro-teins made during productive infection (8, 19).
In ourlaboratory,theIEpolypeptidesidentified have been designated by their apparent
molec-ularweightand include Vmw 175, 136, 110, 87, 68,
63, and 12. The IE polypeptidesinclude the
a-polypeptides class as defined by Honess and
Roizman (8). These authors have shown that
functional a-polypeptides synthesis is required
for the subsequent appearance oflater
classes,
,BandX-polypeptidesin theirterminology,which
are synthesized in the absence of virus DNA
replication (8).
Theobservations detailed above suggest that IE RNA codes for one or more virus
polypep-tides whichregulateamajoreventin virus tran-scription; thatis,the transition from IEtoearly
RNA synthesis. It is as yet unknown how IE
polypeptides functioninthisregulation,butit is knownthatthesepolypeptidesaremodified and
preferentially accumulated in the nucleus (17)
and have DNA binding properties in vitro (R. T. Hay and J. Hay, manuscript submitted for publication).
We have previously mappedthe locationson the HSV-1 genome ofcytoplasmic and nuclear HSV-1 IE RNAs (3) and found that these tran-scripts hybridized predominantlyto DNA frag-ments containing, or adjacent to, the inverted
42
on November 10, 2019 by guest
http://jvi.asm.org/
repeat regions of the virus genome. We report here the results ofexperiments designedto char-acterize the IEmRNA's. IE mRNA's havebeen resolved byelectrophoresisthrough denaturing
gels, and the separated size classes have both
been mapped against HSV-1 DNA fragments
andusedtodirect invitroprotein synthesis in a
cell-free translationsystem.
MATERIALS AND METHODS
Cells andvirus.BHK-21 (C13)cells were grown asmonolayers in 80-oz. (ca 2.37-liter)rollerbottles and infected with HSV-1 (Glasgow strain 17) at a multi-plicity of infection of50PFU/cellas described previ-ously (3).
Labeling RNA.Cellswere pretreated with 200ug ofcycloheximide per ml for1hand either infectedor mock infected (MI) with HSV-1. Cells were labeled with[nP]orthophosphatefrom0 to 6hpostinfection in the presence ofcycloheximide as previously de-scribed (3).
Labeling DNA. HSV-1 DNA was 32P-labeled in vitro bynick translation as previously described (3). Cell fractionation and RNAextraction. Cyto-plasmicRNAwaspreparedbylysisof cells with Non-idetP-40,removal of nucleiby low-speed centrifuga-tion, and phenol-chloroform extraction of the cyto-plasmic fractionaspreviouslydescribed(18).
SelectionofpolyadenylatedRNA.Cytoplasmic RNAwasrecoveredbylow-speedcentrifugation and dissolved in2ml ofbindingbuffer(0.5 MKCI,25mM Tris-hydrochloride, pH 7.9). The RNAwasapplied to acolumn ofoligo(dT)-cellulose (obtainedfrom Collab-orativeResearchInc.)and washed with3volumes of binding buffer. Polyadenylated [poly(A)] RNA was eluted with distilled water, made0.4M with respect tosodium acetate, andprecipitated with 3 volumes of ethanol by overnight storage at -20°C. RNA was recoveredbylow-speed centrifugation.
CH3HgOHagarosegelelectrophoresis. Prepar-ative and analytical RNA electrophoresis was per-formed through 1.5% agarose gels containing 5 mM
methyl mercuric hydroxide (CH3HgOH) basically by the method of Bailey and Davidson (1). Pelleted poly(A) RNAwasredissolved in50plof Ebuffer (50 mM boric acid, 5 mM Na2B407-1OH20, 10 mM Na2SO4,and1mMEDTA,pH 8.2), containing25mM CH3HgOH, 10%glycerol, and sufficientbromophenol blueto act as a tracking dye. RNA waskeptfor 10 minat roomtemperatureimmediately preceding elec-trophoresis.
Preparative RNAelectrophoresis (approximately5
iLgof RNA pergel) wasperformedontubegels(12by 0.6 cm)at 6mA/gel for5h at roomtemperature. Gels werethen soaked for1 hin 10mM ,B-mercaptoetha-nol-20 mM Tris-hydrochloride (pH 7.9) to remove
CH3HgOHboundtoRNA.Gelswerescanned witha
Joyce-Loebel UVscanner to locate the endogenous, contaminating 288 and 188 rRNA's. Gels were cut into 2-mm slices with a Mickle gel slicer, andeach slicewasmacerated in 1 ml ofelutionbuffer(1 mM EDTA,10mMTris, pH 7.9,containing10lOg of yeast tRNA perml), usingaDouncehomogenizer.RNAwas
eluted by shaking at 4°C overnight. The agarose was pelleted by low-speed centrifugation, and the super-natantfractions were retained. A10-plsample of each fraction wascounted in Aquasol to estimate the dis-tribution oflabeled RNA in the gel.
AnalyticalRNA electrophoresis was performed on slab gels (20 cm long). Electrophoresis was at 30 V overnight, and upon completion, gels were soaked in 0.5Mammonium acetate containing 1 ug of ethidium bromide per ml for 1 h. RNA bands were visualized by UVfluorescence,and thegels were dried and subjected toautoradiography by using Kodirex film.
Hybridization of RNA to total HSV-1 DNA. Hybridizationof[nP]RNAto HSV-1 DNA bound to nitrocellulosefilterswasperformed as described pre-viously (9).Individual gel fractions were hybridized to onefiltercontaining2,ugofHSV-1 DNA and one filter containing 2 utg of bacteriophage A DNA. Counts bound to A DNAfilterswere subtracted from those boundtoHSV-1 DNA filterstoestimate HSV-1-spe-cific RNA.
Quantitative estimates of the percentage of virus-specific labeled RNA in the total cytoplasmic poly(A) RNA and in each ofthe pooled fractions were made asabove, except that filters contained 5,g of DNA and reactionswereallowedtoproceedfor40h. The efficiency of these reactionswasestimated inaparallel reaction, using HSV-1 cRNA, made with Escherichia coli RNApolymeraseasdescribedpreviously(3). Un-dertheseconditions,itwasfound thatapproximately 45% of theHSV-1 cRNAinput hybridizedtothe HSV-1DNA filters.
Preselection of RNAagainstHSV-1 DNA
frag-ments.Poly(A)RNAin 70% formamide-2xSSC(lx SSC is 0.15 M NaClplus0.015M sodiumcitrate)was incubatedfor20hat37°C, with10-,Agequivalents of the individualHSV-1 DNAfragments boundto nitro-cellulose disks. Filterswereextensivelywashed in2x
SSC, either withorwithout the addition of25 tLg of RNase A per ml. The filters werethen incubated in iodoacetate solution (0.15 M sodium iodoacetate,0.1 Msodiumacetate in2x SSC, pH5.2)at54°Cfor40 mintodestroy any residual RNaseactivity (23),and further washed in 2x SSC. Hybridized RNA was elutedfrom the filtersby boilingin0.lx SSC for10 imm in the presence of10,ugof yeasttRNA carrier per ml.
Hybridizationtoblots.[ P]RNAwashybridized toHSV-1 DNAfragmentsand immobilized to nitro-cellulosemembraneby theSouthern blot technique
(21),asdescribedpreviously (3).Hybridizationto
frag-ments wasdetectedbyfluorography, using Kodak X-Omat H film and DuPont CronexLightningPlus in-tensifyingscreens.Filmswereallowedtoexposefor up to 3weeksat-70°C.
Invitro translation. Thepooled RNAfractions weremade0.5M withrespecttoKCI and reselected onoligo(dT)-cellulose columns,asdescribedabove,to remove residual agarose. The RNAeluted from the columnswasethanolprecipitatedin thepresenceof 5 ytg ofE. coli rRNA carrier. RNAwasrecovered by low-speedcentrifugation, dissolvedindistilled water, and used to direct cell-free translation inthe rabbit reticulocyte lysate system describedpreviously (18).
on November 10, 2019 by guest
http://jvi.asm.org/
44 WATSON, PRESTON, AND CLEMENTS Endogenous mRNAwasdestroyed by preincubation with micrococcal nuclease (16). The products of in vitrotranslationwereseparated bypolyacrylamidegel electrophoresis (PAGE) as described by Marsdenet al. (11), and were detected by autoradiography by usingKodirex film.
Preparation ofin vivo-labeled IE polypeptides. Cellswereinfected with 50 PFU of HSV-1 per cell in medium containing 200 ,ug ofcycloheximide per ml and incubated for 5 h in the presence of the drug. Cultures were then washed three times with pre-warmed mediumcontaining5jigofactinomycinD per ml and incubated for 1 h with phosphate-buffered saline containing 100,uCiof[3S]methionineper ml. Cellswere harvested and disrupted asdescribed by Marsden etal. (11).
RESULTS
Analytical RNA electrophoresis.
Cyto-plasmic RNAswereisolated from infected (IE)
and uninfected (MI) cells labeled for 6 h with
[32P]orthophosphate in the presence of
cyclo-heximide, and thepoly(A) RNAswereselected
by chromatography on
oligo(dT)-cellulose
col-umns.Thisprocedure didnotresult in the
com-plete removal of ribosomal RNA, but thetrace
amounts of endogenous 28S and 18S rRNA's
provided useful size markersonsubsequent
anal-yses.
IE and MI poly(A) RNA's were denatured
with25 mM CH3HgOHandelectrophoresed in
parallelon anagaroseslabgel containing5mM
CH3HgOH. After electrophoresis, the gel was
stained with ethidium bromide, and the RNA
was visualized by UV fluorescence (Fig. 1). In
addition tothe 28S and 18S rRNA bandsseen
in the MI RNA track, there were three
induced bands in the IE RNA track. These virus-inducedbandswerealso detectedby
autoradiog-raphy of the gel (Fig. 2). By using the rRNA
species asmarkers assumedtobe5.3 kilobases
(kb) and 2.1 kb inlength (12), thesizes of the
virus-induced bands were estimated to be 4.7,
3.0,and2.0kb.Henceforth,these RNAs will be
referredtobytheir sizedesignation.
PreparativeRNAelectrophoresis.IE and
MI poly(A) RNA preparations (approximately
5 ,ug each) were electrophoresed in parallel
throughagarosetubegels containing
CH3HgOH.
Afterelectrophoresis,the gels weresoakedin 10
mM 8-mercaptoethanol to remove CH3HgOH
(1) andUV scannedto detect the rRNAmarkers
andthe threevirus-induced bands.
Thegelswerethencut into 2-mm slices,the
slices were homogenized, and the RNA was
elutedovernightinlow-salt buffer. Mostof the
agarose was removed by low-speed
centrifuga-tion, and a sample of each supernatant was
counted to determine the total distribution of
labeled RNA in the gel. This is shown for IE
[image:3.508.294.437.63.283.2]RNA inFig.3.
FIG. 1. Poly(A) cytoplasmic RNAs isolated from HSV-l-infected (HSV)andMI BHK cells were sepa-ratedby electrophoresisthroughCH3HgOHagarose gels anddetected byUV illuminationafterethidium bromide staining. The positions of the endogenous 28Sand18SrRNA's,the4.7-, 3.0-,and2.0-kb virus-induced RNA species, and the bromophenol blue (BPB) dye frontareindicated.
Part of each elutedgel fractionwasthen
hy-bridized to total filter-bound HSV-1 DNA to
determine the distribution ofvirus-specific RNA
(Fig. 3). The distribution ofvirus-specific RNA
closely paralleled that of total RNA, and the
peaks exactly coincided. This experiment did
notallow accurateestimation of the percentage
of totalcountsthatwerevirusspecific, sincethe
eluate volumes varied somewhatand the
hybrid-izationreactions didnot go tocompletion. More
quantitative estimates were made by using, in
parallel, the hybridization of HSV-1 cRNA
(madewithE. coliRNApolymerase)tomeasure
theefficiency of the hybridization reactions; this
showed that total labeledIERNAwas
approx-imately 70% virus specific. These proportions for
the virus-specific peak fractions were greater
than 80%.
Theportions of the IE RNAfractions
remain-ingwerepooled as indicatedin Fig. 3 (fraction
1 contained the 4.7-kb species). The MI RNA
fractionsofequivalentsizewerelikewisepooled,
using theendogenousrRNAspeciestoalignthe
two gels. The pooled IE RNA fractions were
divided;onehalfwashybridizedtoHSV-1DNA
fragments and theother wasusedtodirect
pro-tein synthesis in vitro. The pooled MI RNA
fractionsweresimilarly translatedinvitro.
Hybridization ofgel fractions to HSV-1
DNAfragments. ThepooledIERNA fractions
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HSV
Ml
18S---ated by digestion with theenzymes used in these
analyses have beenobtained (25; A. J. Davison
and N. M. Wilkie, personal communication).
Thesemaps areshownforone orientation of the
unique regions (Fig. 4). Fluorographs of the
pooled IE RNA fractions hybridized to the
BamHI, HindIll, and HpaI digests of HSV-1
DNAareshown inFig.5,6,and7,respectively,
andarecomparedtothe total fragmentpatterns
(revealed by hybridization of 32P-labeled HSV-1 DNA to the blotstrips) and the hybridization
patterns of unresolvedpolyadenylated
cytoplas-mic IE RNAs.
It is apparent from hybridization to the
BamHI digest (Fig. 5) thatgood separation of
RNAsequenceshas been obtained.Hence,with
peak fraction 5 there was abundant
hybridiza-tiontoBamHI-n, -x, and-z, ascomparedtolittle
or nohybridizationtothesefragments with
frac-li
'o
nw E
rz
z _
(A ._ o
FIG. 2. Autoradiograph ofthegel shown inFig.1.
were each hybridized to blot strips containing
the DNA fragments generated by digestions
with theBamHI,HindIII,andHpaIrestriction
endonucleases,andhybridizationwasvisualized by fluorography. Interpretation of these RNA
mapping datarequiresaknowledge of the
HSV-1 genome structure.
Asfirst describedby SheldrickandBerthelot
(20), thegenomehasanunusualstructure,
con-sistingoftwouniqueDNAregions (ULandUs),
each flanked byinvertedrepeat regions (TRL/
IRL,TRs/IRs) andjoined atIRL-IRs. Four ge-nomearrangements,resultingfrom thepossible
combinations of inversions of the two unique
regions, are present in DNA preparations in
approximately equal amounts (2, 7). A further
consequence of thisstructure is that a number
offragments produced by digestionwith a
re-striction endonucleasemay contain sequences in
common (25). Hence, hybridization ofRNA to a fragment containing any part of the repeat
regionsdoesnotnecessarilyimplythat this RNA
was transcribed from that particular fragment.
Physicalmaps for the DNAfragments
gener-18S 28S
51
3 1A
4L,,
2Fraction Number
FIG. 3. Distribution of32P-labeled total and virus-specific RNAafterresolutionof IE mRNA's on pre-parativegels. The positions of endogenous 28S and 18S rRNA'sareindicated,as arethe positions of the eluted RNAfractions(underlined numbers1through 9), which werepooledand used insubsequent anal-yses.
Map Units
0 01 02 0.3 04 0-5 06 07 0.8 0.9 1
Hind III
o h j kI d imn g
Hpa
mo pkuj n t b h f e qsvrml c g
I
Im
i 1 IBam HI
s e c o m t pu gv rw d h o f b sq nJzxyq
aifi'debj g c'
a=cm
d=g.m
k=s-.q
FIG. 4. Physical mapsforthefragmentsofHSV-1 DNA generated by restriction endonucleases HindIII, HpaI,and BamHI.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.508.79.224.72.398.2] [image:4.508.258.449.276.415.2] [image:4.508.256.450.508.640.2]IE 1 2 3 4 5 6 7 8 9 DNA *e
41.KI ..
k ;
4-nvi
0
n ... .
S
FIG. 5. Fluorographs ofthepooledIERNAfractionsindicated inFig.3(fractionsl1through9)hybridized
totheBamHIfragments ofHSV-1I DNAarecomparedtothehybridizationpatternsofunresolvedcytoplasmic poly(A)IEG RNAs(IEG)and HSV-1DNAlabeledbynick translation(DNA).Fractions1,3, and5containedthe 4.7-,the3.0-, and the 2.0-kb RNA sizeclasses,respectively.
IE 1 2 3 4 5 6 7 8 9 DNA
[image:5.508.107.405.54.383.2]his ... .
FIG. 6.
Fluorographs
ofthepooledIERNAfractions(i through9)hybridizedtotheHindrII
fragments of HSV-1 DNA arecompared
to thehybridizationpatternsofunresolvedcytoplasmic poly(A) RNAs(IE)and HSV-1DNA (DNA).46
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.508.122.405.441.636.2]FIG. 7. Fluorographs ofthepooledIERNAfractions (1 through 9) hybridized to the HpaI fragments of HSV-I DNA arecomparedtothehybridization patterns of unresolved cytoplasmic poly(A) RNAs (IE) and HSV-IDNA(DNA).
tions 3 and 1. A smallamount of material
hy-bridizing toBamHI-j, -k, and -qappears to be
distributedthroughout the gel, and this
presum-ably reflects considerable sizeheterogeneity of
RNAspecies containing thesesequences. Little
RNAcomplementarytoBamHI-owasdetected
afterresolutionof IE RNAsonthesegels, and it
is possible that these sequences were resolved
outside thegel fractions analyzed.
The RNAmapping data for fractions1,3,and
5 (corresponding to the 4.7-, 3.0-, and 2.0-kb RNA species) have beensummarized in Fig.8.
Inmaking this summary,onlyabundant
hybrid-ization tofragments has been considered. This
analysis may locate only the major sequences
present in the separated RNAs. Hence, small noncontiguous orleader sequences may fail to
bedetected.
Fraction 1 hybridized predominantly to
BamHI-yand alsoto-kand-q.Fragmentsyand
q are adjacent and map wholly within TRs/
IRs (Fig. 4),whereas k is a fusionfragmentof q
and s. Because there was no hybridization to
fragments, the 4.7-kb RNAappears tohybridize
exclusivelytopart ofthe TRs/IRs region.
Fraction 3 (the 3.0-kb RNA) hybridized
pre-dominantly toBamHI-b and -e, and also to -s.
BamHI-slies entirelywithinTRL/IRL,whereas
fragmentsb and e contain the same sequences
fromTRL/IRLbut differ incontainingsequences
from opposite ends of UL. That hybridization
was to the repetitive rather than the unique
DNA sequences of BamHI-b and -e may be
deduced from the HpaI digest (Fig. 7), where
there was hybridization to HpaI-m, but not to
HpaI-sor toanyotherfragmentwhichdoesnot
containsequences presentinHpaI-m. The
3.0-kb RNA, therefore, hybridized only to part of TRL/IRL.
Fraction 5 (the 2.0-kb RNA) hybridized
pre-dominantlytothree distinctregions of thevirus
genome. There washybridization toBamHI-b,
-n, -x, and -z (Fig. 5). Fragments x and z are
adjacent and lie at the rightend of Us (inthe
orientationgiven inFig. 4). Fragmentzcontains
only Ussequences, whereas x contains part of
TRs/IRsinaddition to Ussequences. A priori,
hybridization to BamHI-n could, therefore, be duetosequencesshared withx.However,since
there was hybridization to HindIII-n (Fig. 6),
which maps entirely within the left end ofUs,
fraction 5 must contain sequences
homologous
to each end of Us. Evidence that these
tran-scriptsarein partcomplementarytoTRs/IRsis
presented below.
Thethird regionto which fraction 5
hybrid-izedmaps inBamHI-b.The
HpaI digest
(Fig.
7)indicates thatthis
hybridization
wastothe ULsequencesof
BamHI-b,
sincethere washybrid-izationtoHpaI-s,butnot toHpaI-mor-r.
a c.d.o.
9.h
5 ... ....:
on November 10, 2019 by guest
http://jvi.asm.org/
48 WATSON, PRESTON, AND CLEMENTS
Map Units
0 01 02 03 04 0-5 06 0-7
Total IE RNA
Ci
Fraction 1
addition, RNA sequences homologous to BamHI-z. These sequencesweresusceptible to 0:8 0:9 1 RNase treatment and arethus nonhomologous to fragment n. The RNase-resistant sequences whichhybridizedtofragment x,therefore,must be complementary to part of TRs/IRs, since these sequences alone are common to both -n
<=:2 and -x.
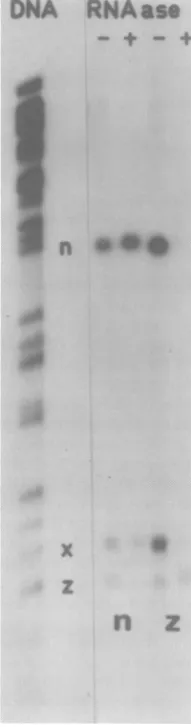
The RNase-resistant sequences selected against BamHI-z hybridized only back to BamHI-z (Fig. 9). Undigested RNA selected against thisfragment hybridized toboth-xand -n, in addition to-z. Thisimplies that there isa
DNA
RNA
ase
L
TRL U
J
be6 S 0
IRL IRSUs TRs
{
1--FIG. 8. Summary oftranscriptmappingdata.Map locations ofthe IE mRNA's contained in resolved fractions 1, 3,and 5(the4.7-, 3.0-,and 2.0-kbRNAs, respectively) are compared to those ofunresolved cytoplasmicpoly(A)IE mRNA'sbythefollowing cri-teria: (i) regions to which an abundance ofRNA hybridizedareindicatedbyadouble continuousline;
(ii) clearly detectable hybridization to a region is representedbyasinglecontinuousline;and(iii) low
levels ofhybridizationareindicatedbya
discontin-uousline.
Selection of transcripts which overlap thejunction between Us and TRs/IRs. To provide evidence that the 2.0-kb transcripts whichmap at eachend ofUsare also
homolo-gous in part to TRs/lRs, total cytoplasmic poly(A) IE RNAs werehybridized atlow tem-perature to duplicate filters containing either BamHI-n or -z. After hybridization, the filters
werewashedextensively. One of each duplicate
was then treated withRNase under conditions
inwhichRNA in an RNA/DNA hybrid is
pro-tected, but single-stranded RNA is susceptible
to digestion. This treatment, therefore, elim-inates tails ofnonhomologous RNA sequences
covalently linked to the hybridized RNA
se-quences. RNA sequences from the
RNase-treated anduntreatedfilterswerehybridizedto thetotalBamHI DNAfragments (Fig.9).RNA selected against BamHI-n, both from treated and untreated filters, was homologous to
BamHI-n and-x.Nondigested filters selected,in
.!..
~ ~
no:.-:'
x
z
n
zFIG. 9. IE mRNA's selected by hybridization to filters containing HSV-1 DNAfragments BamHI-n
or-zwereeither treated withRNase(+)oruntreated
(-), andthe hybridizedRNAs wereeluted. Fluoro-graphs of the elutedRNAsampleshybridizedtoblot stripscontainingtheBamHI DNAfragments(right)
are compared to the total fragment pattern (left) revealedbyhybridization of32P-labeled HSV- I DNA tothefragments.
Froction 3
E::-Froction 5
J. VIROL.
JL
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.508.67.258.75.346.2] [image:7.508.317.413.216.581.2]transcript whichspansthe BamHIcleavagesite
between-zand-xand which containssequences
presentinTRs/IRs.
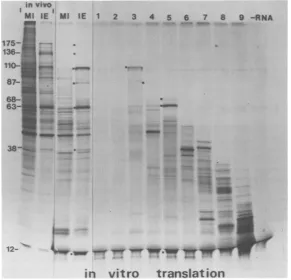
In vitrotranslation ofpooledRNA
frac-tions. The pooled IE and MI RNA fractions
wereusedtodirectprotein synthesisinarabbit
reticulocyte cell-free translation system. For
comparison, unfractionated IE and MI RNAs
were translated in parallel. The products of in
vitro translation were analyzed by PAGE, and
theresultantautoradiographsareshown inFig.
10and 11. Also shown areIE and MI
polypep-tideslabeledinvivo.
Anumberofvirus-specific polypeptideswere
made when the cell-freetranslationsystemwas
programmed with total IERNA, and attention
is drawntopolypeptides Vmw 175, 136, 110, 87,
63,and 12(Fig. 10).Thesynthesis,both in vivo
and in vitro, ofa virus-induced polypeptide of
estimated molecular weight 38,000 was als6
noted(Fig. 10and11).Thispolypeptidehasnot
been classifiedanIEpolypeptide, since its
syn-thesis isnotinvariablyobserved.
The fractionated IE RNApreparations,
like-wise,directed thesynthesisofanumber of virus
in vivo
MlIGE Ml IE 1 175- .._.
136-110-r
87-68-_ 63-_
38::::0s_
polypeptides. Fraction 3 (the 3.0-kb mRNA
band) specified primarily Vmw 110, although
smallamountsof Vmw 87were also synthesized
(Fig. 10). Synthesis of this latter polypeptide
was also directed by fraction 4 RNA, which indicates that the major IE mRNA contained by
fraction3specified Vmw110.
Fraction5 (the 2.0-kb mRNA's) specifiedthe
invitro synthesis ofVmw68, 63,and12(Fig. 10).
Thesynthesisof small amounts ofVmw 63 and
reducedamountsof Vmw12 wasalso directed by
fraction4.Thevirus-inducedpolypeptide of
es-timated molecular weight38,000 wasprimarily specified byfraction7.
Few products of translation of fraction 1
mRNA (the 4.7-kb size class) were apparent
(Fig. 10),and thispresumably reflectsthe diffi-cultyofisolatinglargemRNA's intact. Alonger
exposure ofthegel shown in Fig. 10 (datanot
shown) indicated thatsmallamountsofVmw175 weredirectedbyfraction1.Becausesynthesisof
other high-molecular-weight polypeptides was
also apparent at such an exposure,
demonstra-tionthat the 4.7-kb virus mRNAspecified Vmw
175alone isequivocal.
2 3 4 5 6 7 8 9 -RNA
:':.:!'.: :,::Zi,B;
. .,
__
12-
w-_
VW v *in
vitro
translation
FIG. 10. In vitro translation ofresolved IE mRNA's. Comparison by PAGE oftheproducts of in vitro translationoftheresolved IE mRNA's(fractions I through9)with those directed bytotal IE mRNA's and total MI mRNA's translated both in vivo and in vitro. Also shown are the products oftranslation of
endogenous mRNA whenthecell-freesystem wasincubatedinthe absenceofadded RNA(-RNA). Virus-specificpolypeptidesareindicatedtotheleftoftheappropriatetrack(0).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.508.106.398.335.615.2]50 WATSON, PRESTON, AND CLEMENTS V.iVQ1YvIII vitro
zE Ml IE MI 1 2 3 4 5 6 7 8 q -PNA
FIG. 11. In vitro translation ofresolved MImRNA's. Comparison by PAGE oftheproducts of in vitro translationoftheresolved MI mRNA's(fractions 1 through 9)with thosedirectedbytotal MImRNA'sand totalIEmRNA's translated both in vivo and in vitro. Virus-specificpolypeptidesareindicatedtotheright of theappropriate track (0).
DISCUSSION
Three distinctvirus-specificRNAbandswere
observed when IE RNAwasseparated by
elec-trophoresis through CH:,HgOH agarose gels.
Thesehadestimated sizes of 4.7, 3.0, and2.0kb. Thetwolarger specieswererelatively
homoge-neous asdetermined by in vitro translationand genomelocation,whereasthe2.0-kbcomponent
washeterogeneous. Thus, the 4.7-kb RNA
hy-bridizedtopartof theTRs/IRsregion(between
map coordinates0.830and0.866,and0.964and 1.000) andspecified small amounts of Vmw 175, whereasthe 3.0-kb RNAhybridized to part of
TRL/IRL (between map coordinates 0.000 and
0.038, and 0.792and0.830)and specified predom-inantlyVmw 110.The2.0-kbRNA hybridizedto threeregions(defined bymapcoordinates0.741
and 0.754, 0.865and 0.898,and0.938and 0.964) andspecifiedthesynthesisofthreepolypeptides (Vmw68, 63,and12).
Thesevirus RNAspeciesweresized by using
28S and 18S rRNA'sasmarkers, andestimates
depend on theaccuracy with which the molec-ularweightsof these markers have been deter-mined. The rRNA molecular weight estimates used herewereobtainedby using denaturinggel systems (12), but similar studies have given somewhat different sizes (1).Nonetheless, with the notable exceptionofthe mRNAcodingfor
Vmw 12,the mRNA'scodingfor IEpolypeptides
are consistent with the sizes required for their
protein codingsequences.
Figure 8 shows that the three virus-specific RNA bandshybridized to restricted regions of the virusgenome,and theseweremostlylocated
within or adjacent to the repetitive DNA
se-quences. Unfractionated IE mRNA's were
shown (Fig. 8) tohybridize to a number of ad-ditional loci mapping in the unique DNA
re-gions,and these RNAsweremostlyof low abun-dance (Fig. 8).IERNAs of low abundancewere
not considered whenmaking the summaries of the DNAregionsrepresented bythethree virus mRNA size classes. Furthermore, certain
tran-scripts, i.e., those hybridizing to BamHI-j (Fig.
.1 ".
;36--%810 7-
87-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.508.116.401.73.385.2]5), were heterodisperse in size and were not
resolved by using this gel system. Therefore,
these analyses allow characterization of the
abundantIEmRNA'sonly.
HSV-1 IEpolypeptideshavepreviously been
mapped by analyses of the virus genomes and
thepolypeptidesexpressed by intertypic
recom-binants between HSV types 1 and 2 (13, 19).
Theseanalysesdependeduponsmall differences
inmobilityonPAGE between equivalentHSV
types 1 and2polypeptides which allowed
iden-tificationof the type-specific proteins expressed byrecombinants.Strictly, this methodmapsthe
genome regions affecting polypeptide mobility
rather than delineating the polypeptide coding
regions. Thetechniqueused heredirectlymaps
themRNA's coding for thesepolypeptides. The
combination of both sets of data has allowed
someof the IE polypeptides tobe mapped on
theHSV-1 genomemoreprecisely.
Intertypic recombinant analyses have mapped
Vmw 175, which is equivalent to ICP 4 (8), in
TRs/IRs (13, 19) andVmw 110,which is
equiva-lent to ICP 0 (8), in TRL/IRL. Recombinants
whichareheterotypicfor TRL/IRLexpressboth the HSVtype 1Vmw110and its Vmw118type 2
equivalent (19).Because bothTRL and IRLare
expressedeveninsuch recombinants which have
afixed,noninvertingorientation of theLregion
(19), this indicates that Vmw 110-coding
se-quences lieentirely within repetitive DNA.This
location is consistent with our data.
Recombi-nants heterotypic for TRs/IRs made both the
HSV-1ICP4polypeptideand itsHSV-2
equiv-alent (13). This suggests that these coding
se-quencesmapentirelywithinTRs/IRs.Our data
are consistent with this location and indicate
thatthe mRNA is homologous toonly partof
TRs/IRs(therewas nohybridization of the
4.7-kb mRNAtoBamHI-nor-x).
Previous analyses ofintertypic recombinants
in ourlaboratory havemappedVmw63between
coordinates0.562and0.790, andVmw68andVmw
12in theS component(19). Correlation of these
analyseswiththe data shown heregivesa more
precise maplocation for Vw63 (betweenmap
coordinates0.741 and0.754) and placesVmw68
and 12,respectively, betweencoordinates0.865
and 0.898, and0.938and0.964.Similaranalyses
of intertypic recombinants reported by other
workers(13)mapped ICP27(anIEpolypeptide probably equivalent to Vmw 63) between map
coordinates 0.790 and 0.830, which is part of
TRL/IRL. The reason for the discrepancy be-tween these data and our map location is un-clear.However,ouranalyseswereperformed by
labelingtheIEproteinssynthesizeduponrelease
of a cycloheximide block, whereas those of Morseetal. (13)wereperformedby
pulse-label-ing thepolypeptidesmadeatvarioustimes
post-infection. Since ICP 27 waslabeled when
cells
werepulsed from9 to 10hpostinfection,a
time
atwhich IE polypeptide synthesisis negligible
(8), it is
unlikely
that the protein mappedwasequivalent to the IE polypeptide Vmw 63. It is
also
pertinentthatrecombinants heterotypicforTRL/IRL made only one type-specific ICP 27
(13).
The results presented here indicate that the
two2.0-kbtranscripts whichmap in theS region
overlapthejunctionbetween Usand both TRs
andIRs.The reasons forthis conclusionare that
only the 2.0-kbRNA (fraction 5) hybridized to
fragments (BamHI-n and -x) containing these
junctions, and that IE transcripts homologous
to BamHI-n are also, in part, homologous to
BamHI-x. Data recently obtained in our
labo-ratory (J. B. Clements and D. J. McGeoch,
manuscript submitted for publication),by
map-ping
cDNA made tothe 3' end ofIE mRNA'sagainst HSV-1 DNAfragments, have indicated
thatthe 3' ends of both these mRNA's liein Us.
This
implies
thatthe 5' ends of these mRNA'smapwithinTRs/IRs,and itseemsreasonableto
presume that thetwotranscriptsaretranscribed
from
locationally
distinct promoters of identicalsequence. Two such mRNA's would contain
identical5'ends, and distinct 3' ends.
Vmw 12 is synthesized from a much larger
mRNA
than isrequired for its coding sequences.Suchasituation has been described for the small
T-antigen of simian virus40 (14), which
shares
N-terminal
peptides
with the largeT-antigen
(15)and isterminatedatacodon which appears
tobe excised from the large T-antigen
rnRNA
(5). Polypeptides with identical N-terminal
re-gionscould
arise
withHSV-1 as aconsequenceof the genome structure. Hence, the mRNA's
which code for Vmw 68 and
Vmw
12 and whichmay map at either end of Us could code for proteins with identical N-terminals (codedfor byTRs/IRssequences)butdistinctC-terminals (coded for by Us
sequences).
These twopoly-peptides chains would be terminated by noni-denticalterminator codons
(encoded
byuniqueDNA sequences from
opposite
ends ofUs),andthe
maxinux
length encoded bytheTRs/IRsDNAwould be that
required
tospecify Vmw
12.ACKNOWLEDGMENTS
Wethank J.H.Subak-Sharpefor hisadvice andinterest in thiswork.
R. J. Watsonwas the recipient ofa Medical Research Council grant fortraininginresearchtechniques.
LITERATURE CITED
1. Bailey,J.M.,and N. Davidson.1976.Methylmercury as areversibledenaturingagent for agarosegel electro-phoresis.Anal. Biochem.70:75-85.
VOL. 31, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
52 WATSON, PRESTON, AND CLEMENTS 2. Clements,J.B.,R. Cortini,and N. M. Wilkie. 1976. Analysis ofherpesvirus DNA substructure bymeansof restrictionendonucleases. J. Gen. Virol. 30:243-256. 3. Clements, J. B.,R. J.Watson, andN.M.Wilkie.1977.
Temporal regulation ofherpes simplex virus type 1 transcription: location oftranscripts on the viral ge-nome. Cell 12:275-285.
4. Costanzo,F.,G.Campadelli-Fiume,L.Foa-Tomasi, and E. Cassai. 1977. Evidence that herpes simplex virusDNAistranscribed bycellularRNApolymerase B. J. Virol. 21:996-1001.
5. Crawford, L.V., C. N. Cole, A. E.Smith,E.Paucha, P.Tegtmeyer, K. Rundell, and P.Berg. 1978. Or-ganization andexpressionofearlygenesof simian virus 40.Proc. Natl.Acad. Sci. U.S.A.75:117-121. 6. Frenkel, N., and B.Roizman. 1972. Ribonucleic acid
synthesis in cells infected with herpes simplex virus: controlsoftranscriptionand of RNAabundance.Proc. Natl. Acad.Sci.U.S.A. 69:2654-2658.
7. Hayward, G.S.,R. J.Jacob,S. C.Wadsworth,and B. Roizman. 1975.Anatomy ofherpes simplex virus DNA:evidenceforfourpopulationsofmoleculesthat differ in the relativeorientations of theirlong and short components. Proc. Natl. Acad. Sci. U.S.A. 72:4243-4247.
8. Honess, R.W., and B.Roizman. 1975. Regulation of herpesvirusmacromolecular synthesis: sequential tran-sition ofpolypeptidesynthesis requires functional viral polypeptides. Proc. Natl. Acad. Sci. U.S.A. 72:1276-1280.
9. Jamieson, A. T.,J.C. M.Macnab, B. Perbal, andJ. B.Clements.1976.Virusspecifiedenzymeactivityand RNAspeciesinherpes simplexvirus type 1transformed mousecells. J. Gen.Virol.32:493-508.
10. Jones,P.C.,G. S.Hayward,andB.Roizman. 1977. Anatomyofherpes simplexvirus DNA. VII. a RNA is homologoustononcontiguous sitesin boththeL and S components of viral DNA. J. Virol. 21:268-276. 11. Marsden, H. S., I. K. Crombie, andJ. H.
Subak-Sharpe. 1976.Controlof proteinsynthesisin herpes-virusinfected cells-analysisof thepolypeptides induced by wildtypeand sixteen temperature-sensitive mutants ofHSV strain17.J.Gen. Virol. 31:347-372.
12. McMaster, G. K., and G. G. Carmichael.1977.Analysis ofsingle-and double-stranded nucleic acidson poly-acrylamide and agarose gelsbyusingglyoxaland acri-dineorange. Proc.Natl.Acad. Sci.U.S.A. 74:4835-4838.
13. Morse, L. S., L.Pereira, B.Roizman, and P. A. Schaf-fer. 1978. Anatomy of herpes simplex virus (HSV) DNA. X.Mappingof viral genesbyanalysisof polypep-tidesand functions specified by HSV-1 xHSV-2 recom-binants.J. Virol. 26:389-410.
14. Paucha, E.,R.Harvey,and A. E.Smith.1978.Cell-free synthesis of simian virus 40 Tantigen.J.Virol. 28:154-170.
15. Paucha, R., A.Mellor, R.Harvey, A. E. Smith, R. M. Hewick, and M. D. Waterfield.1978.Large and small tumour antigensfromsimian virus40 have identical aminoterminimappingat0.65mapunits.Proc. Natl. Acad. Sci.U.S.A. 75:2165-2169.
16. Pelham, H. R. B., and R. J. Jackson. 1976.Anefficient mRNA-dependenttranslationsystemfromreticulocyte lysates.Eur.J.Biochem. 67:247-256.
17. Pereira, L., M.H.Wolff,M.Fenwick,and B. Roizman. 1977. Regulation ofherpesvirus macromolecular syn-thesis V.Propertiesofpolypeptides madeinHSV-1and HSV-2 infected cells.Virology 77:733-749.
18. Preston, C.M. 1977. Thecell-freesynthesisof herpesvi-rusinduced polypeptides.Virology 78:349-353. 19. Preston,V.G.,A. J.Davidson,H. S.Marsden,M. C.
Timbury, J. H.Subak-Sharpe, and N.M.Wilkie. 1978.Recombinantsbetweenherpessimplexvirustypes 1and 2:analysesofgenome structuresandexpression ofimmediateearlypolypeptides. J. Virol. 28:499-517. 20. Sheldrick, P., and N. Berthelot. 1974.Inverted repeti-tionsinthe chromosomeofherpes simplexvirus.Cold SpringHarborSymp. Quant. Biol. 39:667-678. 21. Southern, E. M. 1975. Detection ofspecificsequences
amongDNAfragments separated by gel electrophore-sis. J.Mol. Biol. 98:503-533.
22. Swanstrom,R.I.,and E. K.Wagner.1974.Regulation of synthesis of herpes simplex type 1 virus mRNA during productiveinfection.Virology60:522-533. 23. Taylor, K.,Z. Hradecna, and W.Szybalski. 1967. A
symmetric distribution of thetranscribing regionson thecomplementary strandsofcoliphageADNA.Proc. Natl. Acad.Sci.U.S.A.57:1618-1625.
24. Watson,R. J., andJ. B.Clements. 1978. Characteri-zation oftranscription-deficient temperature-sensitive mutantsofherpessimplexvirus type 1.Virology 91: 364-379.
25. Wilkie, N. M. 1976. Physicalmaps forherpes simplex virustype 1DNA for restrictionendonucleasesHindIII, Hpa-1, and X. bad. J. Virol. 20:222-233.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/