0022-538X/79/08-0315/10$02.00/0
DNA
of Epstein-Barr Virus
VI.
Mapping
of the
Internal
Tandem Reiteration
DOUGLASS GIVENANDELLIOTTKIEFF*
SectionofInfectious Disease, Departmentof Medicine,andDepartments of MicrobiologyandVirology,
Kovler
ViralOncology
Laboratories, TheUniversity of Chicago, Chicago,
Illinois60637Received forpublication 15 January 1979
Epstein-Barr virus (B95-8) DNA consists of short (10 x 106) and long (87 x
106) unique DNA sequencesjoinedby10tandem reiterations of a 1.85x 106DNA
segment. The reiterated sequence contains BamI and BglII sites separated by 4
X 105. The 4.5 x 105 and 14.0 x 105 segments generated by cleavage of the
reiterated DNA with BamI andBglIIcontainsequences whichhybridize to each
other,suggestingthat theinternaltandemlyreiterated sequence has a direct or
inverted repeat within it. Theoppositeends of thelinear, nicked, double-stranded
DNAmolecule(R.F.Pritchett,S. D.Hayward,and E. D.Kieff,J.Virol.
15:556-569, 1975) consist of from1to 12direct repeatsof another3 x 105sequence (D.
Given and E. Kieff,J.Virol. 28:524-542, 1978; D. Given, D. Yee,K.Griem, and
E. Kieff,J. Virol.30:852-862, 1979).There isnohomology between the internal
reiterated sequence and either terminus. However, part of theinternal reiteration
(lessthan 5x105)is reiteratedat twoseparate locations in thelong unique region.
The internal reiterations are a source of variation within EBV (B95-8) DNA
preparations. Thus,althoughthemajority of molecules contain 10tandem reit-erations, some molecules have 9, 8, 7, 6, 5, 4, or fewer tandem reiterations. A
consequence of thisvariabilityis that theKpnIAfragmentand theEcoRI/HsuI
Afragmentconsist ofafamilyofsevenor morefragments differinginthe number
of tandem internal reiterations. The EcoRI/HsuI A fragment of EBV (W91)
DNA is approximately 6 x 106 smaller than the largest and dominant
EcoRI/
HsuIAfragmentof EBV(B95-8)DNA. EBV (W91)DNAalso differs from EBV
(B95-8) DNAbyanadditional7x 106to 8x106of DNA in thelongunique DNA
region (D. Given and E. Kieff, J. Virol. 28:524-542, 1978; N. Raab-Traub, R.
Pritchett, and E.
Kieff,
J. Virol. 27:388-398, 1978). These data suggest the possibilitythat thesmaller number of internal reiterations in EBV (W91) DNA may be a consequence of the additional unique DNA and a restriction in the overall size of EBV DNA.Epstein-Barr virus (EBV) DNA is a linear, nicked, double-stranded molecule of 105 x
10'
(15). Thebuoyantdensity (11, 13, 15, 18, 24,25)
andmelting temperature (11, 15) of the DNA
are compatible with a guanine-plus-cytosine
content of57 to 58%. The DNA of the B95-8
isolate of EBV has beenaprototypeforstudies
of theorganizationof EBV DNA(for review,see
reference 10).Both endsof the DNA consist of
avariable number of directrepeatsof thesame
sequence of400to500basepairs (6, 7).The size
(8) and order (6) of theHsuI, Sall, and EcoRI
restriction endonuclease fragments of EBV
(B95-8) DNA areshown in Fig. 1. These data
and the analysis of partially denatured
mole-cules(4) indicatethat,with theexceptionof the
variability inthe number ofcopies ofthe
reit-erationateach endof theDNA,theremainder
ofthe DNAisorganizedin asinglearrangement.
The results of treatment of EBV DNA with
KpnI restrictionendonucleasearediscordant(8)
as KpnIproduces a 20 x 106 fragment in
sub-molar amounts, and the sum of the molecular
weightofallfragments,includingthesubmolar fragment, isapproximately 20 x
106
more than themolecularweight of EBV DNA.Analysis of the fragments produced by
cleav-age of the DNA with BamI andBglII reveals
thepresenceofa2 x 106fragment inexcessive
amounts (8, 17). Complementary RNA made
fromthe 2x 106 BamIfragment, whichis
pres-entin a 10-fold molarexcess, hybridizestothe
HsuIAand EcoRIAfragments, indicatingthat
there areapproximately 10tandemreiterations
of the2 x10' sequence in theHsuIAand EcoRI
Afragments(17).Partof this reiteratedDNA is transcribed in Burkitt tumor tissue (3) and in
restringently infectedBurkitt tumorcellsgrown
315
on November 10, 2019 by guest
http://jvi.asm.org/
EcoRI IKhet I J
HsuI
SalI Hhet
NmH-EcoRI/Hsu I
A
A
A G G F K- B
a JG H E SI
G F
G E C F.H
E H C Dhet
C KL F D FGHhet
B Dhet
A a (H) (F) (C) (E) J (D) (G)(HIhet)
l _ I I I I
[image:2.504.78.408.54.143.2]0 10 20 30 40 50 60 70 80 90 100 110
FIG. 1. EcoRI(6),SailI (6),HsuI(6),andEcoRI/HsuI restriction endonucleasecleavage sites in EB V (B95-8)DNA.Fragmentsaredesignated by capitalletters(6).Acapitalletter enclosedby parenthesesabove the EcoRI/HsuIfragmentsindicatestentativeassignment ofmaplocation based on correspondence between the sizes ofthefragments (Fig. 3)andthe sizes ofthe EcoRI and HsuIoverlap regions. The positions of the EcoRI/HsuIA,B, and Jfragmentsweredetermined byhybridization ofblotsofagarose gelscontaining
EcoRI/HsuIdigest ofEBV(B95-8)DNAtolabeled HsuI AorBorEcoRI Cfragment ofEBV(B95-8) DNA.
in culture (14). The objective of these
experi-ments was todeterminethe location of the 2 x
106reiteration within the EcoRI A andHsuI A
fragments. In thecourseof this work itbecame
apparentthat the discordantfindingwithKpnI isduetothepresence of a minorpopulationof
molecules which contain fewer copies of the
internal reiteration butare otherwise identical
tothe dominantpopulation.
MATERIALS AND METHODS Cellculture, viruspurification, and prepara-tion of viral DNA. Viral DNA was obtained by
sodiumdodecylsulfate-phenolextraction(12)of virus
purifiedfromtheextracellularfluid of B95-8 and W91 cell cultures. InitialculturesofB95-8and W91cells were obtained from G.Miller,Yale University,New
Haven, Conn.Theproceduresused tomaintain cul-turesforviruspurificationand thepurificationof virus havebeen describedpreviously (5).
Separation and determinationof the molecu-larweight ofrestrictionendonucleasefragments
of EBV. EBV DNA was incubated with atwo- to
fivefold excess ofKpnI (NewEnglandBiolabs,
Bev-erly, Mass.), EcoRI, HsuI (8) (or its isoschizomer,
HindIII), Sall, XbaI, BamI,orBglII(BRL, Bethesda, Md.)for 2.5 hat37°C (6). For doubledigestionwith
EcoRI,HsuI, BamI, orBglII,2to 4yig of the DNA wasincubated at37°Cwithafivefoldexcessof enzyme in asolutionconsistingof 50 mMNaCl,7mM
MgC92,
2mM
,8-mercaptoethanol,
and 20 mM Tris-hydrochlo-ride,pH 7.4.EcoRI, HsuI,SalI, XbaI,KpnI,EcoRI/HsuI, BamI, andBglII DNA fragmentswere sepa-ratedbyelectrophoresisat40Cin 0.3 to 0.4%(wt/vol)
agarose gels incylinders (1 by 28 cm) or inslabs(0.5
by23by 27cm) (6, 8). BamI, BglII, BamI/EcoRI,
BamI/HsuI,BglII/HsuI,andBamI/BglIIfragments
were also separated in 0.8% (wt/vol) agarose gels.
DNAfragmentswere stainedwithethidium bromide andphotographed (type57film,PolaroidCorp.,Oak
Brook,Ill.) under UVillumination (6,8). DNA
frag-mentstobecleaved with asecondenzymeorto be labeled in vitro were cut from the gelunder direct visualization.Theagarosewasdissolved in 5 volumes of5 M sodium perchlorate at 450C, the DNA was separated by chromatography on hydroxylapatite
(HTP, Bio-RadLaboratories, Richmond,Calif.),and the ethidium bromidewasremovedbydialysis against
Dowex 50 resin in the H form (6). The molecular
weights of the EcoRI/HsuI fragments were deter-mined from a log linear plot of the electrophoretic
mobility of thefragments in a 0.3% (wt/vol) agarose slab gel relative tolambda DNA (BRL) (22), EcoRI
fragments oflambda DNA (22), HsuI fragments of EBV(B95-8)and (HR-1)DNAs (8), and EcoRI frag-mentsof EBV(B95-8)DNA (8) as reference DNAs in adjacent wells on the same slab. The molecular
weights of BamI or BglII fragmentswere similarly determined from the comparativeelectrophoretic mo-bility of thefragments in 0.3% (wt/vol) agarose gels with the EcoRIfragmentsof EBV(HR-1)DNA(8)as described above or in 0.8% (wt/vol) agarose slab gels relative to EcoRIfragmentsoflambda DNA (22). The
sizesof the BamI orBglIIfragments of theseparated
HsuI A orEcoRI Afragmentsand of theBamI/HsuI,
BamI/EcoRI, BglII/HsuI, BglII/EcoRI, or BamI/
BglIIfragmentsweredeterminedby electrophoresis
in 0.8 to 1% agarosegels relative to theEcoRI frag-ments of lambda DNA(22).Themolecular weight of the smallest EcoRI fragmentof lambda DNA is 2.1 x 108(22).Therefore, determinationof themolecular weightoffragmentssmaller than106from their
elec-trophoreticmobility relativetothe EcoRIfragments
oflambda DNA issubjecttoerror, even in 1% agarose gels.
Labeling ofDNA in vitro. EBV DNA or
frag-ments of EBV DNA were labeled in vitro by nick
translation (6, 9) using DNApolymerase Iof Esche-richiacoli (Boehringer MannheimCorp., New York) and a-3P-labeled dCTP (300 Ci/mmol; Amersham
Corp.,ArlingtonHeights,Ill.). The specific activity of thelabeled DNAwas0.5x l08 to1 x108cpm/,ug.
HybridizationoflabeledDNA toblots of frag-mentsof EBV DNA. Blots (20) ofseparated frag-ments of EBV DNA wereincubatedat680Cin 1mlof asolution consisting of 104 to 105 cpm of denatured, labeled EBV DNA, 1 mg ofdenatured calf thymus DNA,0.01 MEDTA, 1.25 M NaCl, and 0.05 M
Tris-hydrochloride, pH7.4.After18hthe filterwaswashed for4h in4xSSC (lx SSC is0.15MNaCl plus 0.015 Msodiumcitrate) at550Canddried at600Cfor1h. ThefilterwasthenexposedtoX-rayfilm(SB5,Kodak
Corp., Rochester, N.Y.), with an intensifying screen
(Cronex Lightning Plus, Du Pont Co., Wilmington,
Del.)at-70°C.
RESULTS
KpnIfragmentsofEBV(B95-8) DNA.The
orderofarrangement of theKpnIfragmentsin
J.
on November 10, 2019 by guest
http://jvi.asm.org/
EPSTEIN-BARR 317
EBV(B95-8) DNA wasdeterminedby
hybridi-zation of blots ofKpnI
fragmentsof EBV DNA to labeled EcoRI,HsuI, SalI,
or BamIfrag-ments whose map positions have been
previ-ously determined (6). The results (Fig. 2)
indi-cate that each ofthe KpnI fragments of EBV DNA, withthe exception of the minor fragments slightly smaller than A and the B fragment, mapped to a specific and unique location. The B (previously C [8]) fragment was 2 M relative to
A EBV (B95-8)DNAKpn
fragment
linkage
Kpn profile: EcoRIfragrrents
A B F G H I
B-o
C-s; E
-OF - 69 B-10'5X2 -;C
84
26 M
6
__ 2I2
H-24
26
K 24
0H
Hsu fragments Sc
E-4
GK I G-if .,
net a*
.*
I
O
p
t
I~~~~
I~~~~~
Ca fragments
B C D A B C E
B- B
0K-F
:# ~~~~~N-
M-N
EcoRl/Hsul Bam
A v
t
A A f
B
Linkage ofKpnl fragmentsof EBV(B95-8) DNA
EBV(B95-8)DNA probe
IJ A G FK B
Atzt/>_\\-t---\r ErEcoRi fragment
GKi
DE:KhetG A O B J C WMI)1B D E &_N$i)F her
AEC C' B
r--- F- -C--I-I -- Sc! fragment
0 10 20 30 4 5r 6c 7 40Cc0cOC
FIG. 2. Mappingof KpnIfragmentsofEBV(B95-8)DNAby hybridizationtolabeledfragmentsofknown mapposition(Fig.1). (A) Photographofanethidiumbromide-stained agarosegelofanelectrophoretically separatedKpnI digest ofEBV(B95-8) DNA is shown in the upperleft. Thesizes ofthefragments were determined from their electrophoretic mobility in 0.3% (wt/vol) and 0.4% (wt/vol) agarose gels (6, 8).
Radiofluorogramsofblotsof KpnIrestriction endonucleasefragmentsofEBV(B95-8)DNAwhich had been
hybridizedtotheseparated32P-labeledEcoRI,HsuI, SalI, EcoRI/HsuI,orBamIfragments ofEBV(B95-8)
DNA. (B) Summary of linkagedatafor KpnIfragments. Thearrowsdrawn indicate hybridization ofthe labeledEcoRI,HsuI,orSalIfragmenttoKpnIfragmentsonblots.The orderoftheKpnI0 andH,M andI,
andNandKfragmentswithin the map distances indicatedby parentheseshasnotbeendetermined.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.93.409.128.587.2]318
otherfragments (8), could be resolved intotwo
discrete bands in 0.3%agarosegels, and mapped
to two distinct locations, 37 x 106 to48 x 106 and66 x 106to 77 x 106.The minorfragments
smaller thanAranged in size from 16 x 106to
27 x 106, hybridized to all labeled fragments
whichhybridized totheAfragment, and failed
tohybridizetootherlabeled fragmentsof EBV
DNA. These data suggest the possibility that
theminorfragments resulted from thepresence
ofa minorpopulation ofEBV DNA molecules
which differ from the majorpopulation only in
the length of the KpnI A fragment. In several
instances, a labeled fragmentsuch asEcoRI G
orK orHsuI B hybridized to asmall extent to
a KpnI fragment (E, K, and B, respectively)
which mapped by other dataat a distant site.
Thiswaslikelyto be a consequence of
contam-ination ofthelabeled fragmentwith an adjacent
fragment from the gel in which the fragments
were separated before labeling in vitro. Thus,
the labeled EcoRI G fragment hybridizes
pri-marilytoKpnI fragments B, C, and J,but also
toKpnIfragment E. The hybridization to KpnI
fragmentE is, however, likelyto be an artifact
of contamination ofthe EcoRI G fragmentwith
a smallamount of theEcoRI H fragment since
thelabeled EcoRIfragmentHhybridizes
exclu-sively to the KpnI fragment E. Similarly, the
; .og MoIecula We!gh' (daltc
I. .!.s
!
,J/
v
/
/'
/
/ ..
EcoRI fragment K is contaminated withasmall
amountof EcoRIhet, and the HsuI fragment B
iscontaminated with the HsuIfragment C.
EcoRI/HsuI A fragments of EBV (B95-8)
and(W91) DNA. TheKpnIenzyme is difficult
towork with because of itslability (6). Digestion
of EBV DNA withacombination of EcoRI and
HsuIrestriction endonucleasesproducesalarge
fragment, EcoRI/HsuI A,which containsmost
of the DNA of theKpnIAfragment (Fig. 1and
2).Thefragmentsproduced bydouble digestion
of EBV(B95-8)or(W91)DNA withboth EcoRI
and HsuI are shown in Fig. 3. The molecular
weight of theAfragmentof EBV(B95-8) DNA
is estimated from itselectrophoretic mobility in
0.3%agarose gelstobeapproximately28 x 106. Minorcomponentswhichtogether compriseless
than 10% of theAfragmentareevidentat26x
106, 24 x 106, 22 x 106, and 20 x 106 (Fig. 3A).
The major andminorcomponentsof the
EcoRI/
HsuIAfragmentwereseparated,labeled in
vi-tro,andhybridizedtoblotscontainingallof the
EcoRI/HsuIfragments.The labeledmajor
com-ponenthybridizedtothemajorand minor
com-ponents of the EcoRI/HsuI A fragments and
alsohybridized totheEcoRI/HsuI Jfragment
to a much lesser extent (Fig. 3B; see Fig. 8,
column5). The results obtained with the labeled
minor components were identical to those
ob-ns1 B
W91 8
I-4?z L
_m
[image:4.504.127.408.381.565.2]I
FIG. 3. (A) EBV (B95-8) DNAwasincubated with a combination of EcoRI and HsuI restriction endonu-cleases, subjectedtoelectrophoresis ina0.35%(wt/vol)agarose slab gel,stained with ethidium bromide, and photographed under UV illumination. The sizes of theEcoRI/HsuIfragments were determined from a log linearplotof the electrophoretic mobility of theEcoRI/HsuIfragments relativetoEcoRIorHsuI fragments
ofEBV(B95-8) DNAorHsuI fragments ofEBV(HR-I)DNA (8). The calibration curve shownabovewas
constructedusing HsuI (B through L)fragmentsof EBV (B95-8)DNA and intact lambda DNA. (B)EcoRII
HsuIfragmentsofEBV(W91orB95-8)DNAwereseparated by electrophoresisinadjacentwellsofa0.35% agarose slabgel.Photograph of the ethidium bromide-stained gel with the fragments of W91 (column 2) and B95-8(column 3) DNA. Autoradiograms of blots of theEcoRI/HsuI fragmentsof EBV (W91orB95-8) DNA whichwerehybridizedtothe32P-labeledEcoRI/HsuIAfragmentareshowntotheleft (column 1)and tothe right (column 4), respectively.
on November 10, 2019 by guest
http://jvi.asm.org/
taimedwith themajor component. As many as
10 minor components differing in molecular
weight byapproximately2x 106 could be
distin-guished influorogramsofblotsof EBV (B95-8)
DNA hybridized to thelabeledEcoRI/HsuI A
fragment (Fig. 3B). These data suggested the
possibility that the difference in sizes of the
EcoRI/HsuIA fragmentswasdueto variation
inthe number ofcopiesof the 2x106reiteration
(6, 17) and that there was homology between
theEcoRI/HsuIAfragmentand another region
of the EBV genome (EcoRI/HsuI J fragment;
seebelowandFig.8,column5).The size of the
EcoRI/Hsu I A fragment of EBV (W91) DNA
is 22 x 106. Minor components which together
constitutedapproximately10%of the total DNA were visible at20 x 106, 18 x 106, 16 x 106, 14
X 106, 12 x 106,and 10 X 106influorograms of
blots of EBV (W91) DNA hybridized to the
labeledEcoRI/HsuIAfragment (Fig.3B).
Mapping within theEcoRI/HsuI A
frag-ment. BamI and
BglfI
are known to cleavewithin the tandem reiteration in the
EcoRI/
HsuIAfragment (6, 17). Therefore, aseriesof
experiments was undertaken using BamI and
BglIItoestablish the arrangement of sequences withinthe EcoRI/HsuI Afragment. One objec-tiveofthis approachwastodemonstrate directly
that the difference inthe sizesof themajorand
minor EcoRI/HsuI A fragments was due to a
difference in the number of copies of the
reiter-ated DNA and not to adifferenceinthe sizesof
theuniquesequence componentsof theEcoRI/
HsuI Afragment. A loglinear plot of the size
and electrophoretic mobility of the BamI and
BglIl fragments of EBV (B95-8) DNA in 0.8%
(wt/vol) agarose
gels
is shown in Fig. 4. The molecular weight ofthe reiterated DNAfrag-ment,BamI V,previously termedBamI S (17)
or
BglIl
R, was determined to be 1.85 x10"
relative to the molecularweightofEcoRI
frag-mentsof lambdaDNA.
TheBglIIAfragmentelectrophoresedin 0.8%
gelsasabroad band and in 0.3%gelsas
multiple
discrete bands differing by approximately 3 x
105 in molecular weight, indicating that the
BglIIA
fragment
extendstoincludeoneend ofthe DNA(6, 7).Threeapproacheswereusedto
map the BamI and
BglII
cut sites within theEcoRI/HsuIAfragment. Inthefirstapproach,
the BamI and BglII fragments which contain
sequences homologous to the EcoRI/HsuI A
fragmentwereidentifiedbyhybridizationof the
labeledEcoRI/HsuI AfragmentDNA toblots
ofseparatedBamI andBglIIfragmentsof EBV
DNA(Fig. 5A andB).The secondapproachwas
totentativelyidentifythe BamI andBglII
frag-ments which contain EcoRI or HsuI sites
by
hybridization ofthelabeledEcoRI/HsuIA
frag-Log Moleculor Weight (doilons)
6 65 7
C.D05.5¢.
m E49
_ 45
L 34
N.l2 5:c5
P2z20
: -x rv
[image:5.504.259.452.68.313.2]Y.42 Z 4&
FIG. 4. Log linearplotofsize andelectrophoretic
mobility ofBamI(photographofethidium
bromide-stainedgel on the left) and BglII (photograph of
ethidiumbromide-stainedgelontheright) fragments
ofEBV(B95-8)DNA in 0.8% agarosegelsrelativeto
EcoRIfragmentsoflambda DNA(22).Themolecular weights of the tuo largest fragments (BamI and
BglII)weredetermined in 0.4% agarosegelsrelative to HsuI (MP) DNA (8). The broken line indicates regionsoftheloglinearplotdrawnby extrapolation.
TheestimateofthemolecularweightsofBamIand BglII fragmentsfromtheseregions istherefore sub-jecttogreater error.
ment to blots of EBV DNA doubly cut with
BglIIandHsuI(Fig. 5A), with BamI and EcoRI (Fig. 5B),orwithBamI and HsuI (Fig. 5B). The
thirdapproachwas to recuttheisolated EcoRI
AorHsuIAfragmentwithBamIorBglIIand
todetermine the size of the resultantfragments
(Fig. 5C). The results are summarized in Fig. 6
andwereasfollows.
(i) The EcoRI/HsuI A fragment hybridizes primarily tothe
Bglll
A, C, or D and Rfrag-ments(Fig. 5A,column2)andtotheBamI Bor
C, G, V, and Xfragments (Fig. 5B, column 2). (ii)TheBglIIAfragmentextendstotheend
ofthe DNA and is 10 x 106 to 13 x 106. This
fragment must therefore contain the short,
unique region. TheBglIICDfragment mustlie
totheright of the reiterated BglIIRfragment
and include the first HsuI cut site, since it is
reduced in size from5.8 x 106 to 4.0X 106when
theDNAiscutwith both HsuI and
Bglfl
(Fig. 5A, column 4) and contains sequences homolo-goustotheEcoRI/HsuIN Bfragment (Fig. 5A,on November 10, 2019 by guest
http://jvi.asm.org/
a ~
4
*
zi.
:I~~~~
%S0
I
I
AA
I
i
,::t:g .I..
af. .
7..-n
1.3 I.F. a .1 1. 'rG :.
:4w, I
el
x. 4:
01
4.
3!v
A
.il,
[image:6.504.64.450.71.318.2]Br! ,
FIG. 5. Radiofluorogramsofblotsof BglIIorBglII/HsuIfragments (A)orBamI,BamI/HsuI,orBamIl
EcoRIfragments (B) ofEBV(B95-8)DNA orofblotsofisolated EcoRI AorHsuI Afragmentrecutwith BamI orBglII (C),which had beenhybridizedtolabeled EBV(B95-8)DNA(designatedT.P.)ortolabeled
restriction enzymefragments ofEBV(B95-8) DNA (designatedbythe letters at the topofeach blot). The molecularsizeofeachfragmentwasdeterminedbycomparison ofelectrophoretic mobilitytoEcoRIfragments ofphagelambda DNAcoelectrophoresedinadjacentwells.
Eco RI IKhlt II A S.I ICV V VV V V V VIVV G IXI
Ugh A R R R R R R R R RR C,0
HsuI A |
o .0 20 30 40
FIG. 6. Mapofthe BamI sitesfrom4.2x 106to 33 x10'and oftheBglIIcut sites from 0 to 34.3 x10'in EBV(B95-8)DNA.
column 3). Thearrangement of theBglII
frag-mentsand of the EcoRI and HsuI
cleavage
sites(6) from 0 to 36 x 106 is shown in Fig. 6. The
arrangement was confirmed by the results of
recleavage of the purified HsuIA or EcoRI A fragment with BglII (Fig. 5C). As expected,
cleavage of the HsuI A fragment with BglII
resulted in three fragments: the BglII A
frag-ment, the reiteratedBglII R fragment, and a
third fragment, BglII CD, whichis reducedin
size because it extendsbeyondtheHsuI
cleavage
site (Fig. 5C, column 1, andFig. 6).
Cleavage
ofthe isolated EcoRI A fragment withBglII
re-sultedinfull-sizeBglIIR andCDfragmentsand
aBglII Afragment reducedin size because of
cleavare
by EcoRIatthe EcoRI siteslocatedat1x 10,3.8x
106,
and6x106
from theleft endoftheBglIIAfragment(Fig.5C,column2,and
Fig. 6).
(iii) Similarly, the BamI BC fragment
con-tains sequences homologous to the EcoRI J
fragment (Fig. 5B, column 3) and is reduced in
sizefrom5.8 x 106to 4.1 x 106when the DNA
is cut with EcoRI andBamI (Fig. 5B and Fig.
5C, column 4), indicating that this fragment
mustlietotheleft of thereiteration.The BamI
XfragmentmustcrosstheHsuIcutsite since it
is reduced from1 x 106 (Fig. 5B,columns2and
4)to0.4x 106 (Fig. 5B,column 5,and Fig. 5C,
column3) when the DNA iscleavedwith BamI
and HsuI.Furthermore,asexpected,thelabeled
EcoRI/HsuIBfragment hybridizestothe BamI
X fragment (data not shown). The BamI G
fragment must extend from the reiteration to the BamI Xfragment since it ishomologousto
the EcoRI/HsuI A fragment and is not
de-creased in sizeby cleavagewith HsuI (Fig. 5B,
column 5) or EcoRI (Fig. 5B, column 4). The
mapof the BamIsites from4.2x 106to33x 106
is shown inFig.6.Theseresultswereconfirmed
by analysis ofthefragments producedwhenthe
purified HsuI A orEcoRI Afragment was cut
with BamI and identifiedonblots with the
la-beledEcoRI/HsuIAfragment (Fig. 5C, columns
3and4). Asexpected, the BamIBC, G, andV
fragmentswere cleaved from theHsuI A
frag-mentandwereidenticalin sizetothefragments
cutfromintact EBV(B95-8)DNA, whereasthe
J. VIROL.
I
& 1.:4
.., 1-5.-..:
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.60.251.387.462.2]X fragment was reduced to approximately 4 x
105, confirming that the size of the component
of the X fragment to the left of the first HsuI
cut site isapproximately 4 x 105.Furthermore,
the BamI BC fragment is smaller when cleaved
from the EcoRI Afragment since the EcoRI site
at6 x 106isin the BamI BC fragment (Fig. 5C,
column 4, and Fig. 6).
(iv) An unexpected finding when blots of
BamIfragmentswerehybridizedtothe labeled
BglIIRorBamI Vfragmentwasthat the
radi-ofluorogramappearedidenticaltothat shown in
Fig. 5B, column 2. All four BamI fragments,
including X, hybridized tothe labeled Bgll R
orBamI Vfragment.These data indicatedthat
the BamI Xfragmentwhich isseparated from
the tandem reiterations by the length of the
BamI G fragment,contains sequences
homolo-gous to BglIl R and BamI V fragments. The
sequences homologoustoBamI V and
BglII
Rfragments mustlie to the left of the HsuI cut
site in the BamI X fragment shown in Fig. 6,
since the labeled EcoRI/HsuI Afragment
hy-bridized to the0.4x 106fragmentof X andnot
to the0.6 x 106fragmentof BamI X (Fig. 5B,
column 5). Thehomologybetween theinternal
reiteration and the BamI Xfragmentisfurther
defined bystudies with labeled components of
thereiterated DNA below.
Toconfirm that themajorand minor
compo-nents of the EcoRI/HsuI A fragment do not
differin thecontentofuniquesequences,a mi-nor band was separated from a
gel,
incubated withBamI,andre-electrophoresed. The sizesof thefragmentsproduced bycleavage
of themajorand minorEcoRI/HsuIAfragmentcomponents
withBamIwereidentical (datanotshown).
Arrangement of BamI and BglH sites
within the internal tandem reiterations.
The sizes of the components of BglII A and
BamIBCtotherightof the EcoRI J-A siteare
4.5x
10"
(Fig.5C, column2) and4.1 x 106 (Fig.5C, column 4), respectively. The sizes of the
componentsofBglII CD and BamI G and Xto
the left of the HsuI A-B siteare 4 x 106 (Fig.
5A, column 4) and4.1 x 106and0.4x 106 (Fig.
5B, column 5),respectively. As summarized in
Fig. 6, theseobservations,that thedistance from
the EcoRI J-Acutsitetothe BgIIIsite within
the first intemal reiteration is 4 x i05 longer
than the distance to the first BamI site and that
the distance from the BglII site in the last
reiterationtotheHsuIA-Bsiteis 5x 105shorter
thanthecorrespondingBamIdistance,both
sug-gest thatthe BamI site is 4 x 105to5 x 105to
the leftof theBglII sitein the reiteration. To
confirm the distance between the BamI and
BglIIsites within the reiteratedDNA,thesizes of the BamI/BglII double-digest fragments
weredetermined by electrophoresis in a 1%
aga-rose gel with EcoRI fragments of lambda DNA
in adjacent wells. As expected, the reiterated
DNA is cleaved into two components (Fig. 7,
column 2). The electrophoretic mobility of the
large component of the reiterated DNA, BglII
RI,
is slightly faster than the smallest EcoRI fragment of lambda, indicating a size of 1.4 x 106. The rapid mobility of the smallercompo-nent,BglIIRK, placesit in the nonlinear part of
thegelbut suggests a size of approximately 4 x
105to 5 x i05.
The data, which are summarized in Fig. 6, indicate that theBamI site in the internal
reit-eration is approximately 4 x 105 to the left of
theBglIIsite and therefore in the leftward 1.4
x
10'
of the reiterated DNA. If the BamI site isnear the left end of the reiteration, BamI BC
wouldcontainonly the left end of the reiterated
sequence and BamI G would contain most of
one complete reiteration, i.e., all of the DNA
betweentheBamIandBglII sites,BglIIR and
most ofthe rest of the reiteration, BglII
RI.
Ifthere are no reiterations within the reiterated
1.85 x 106sequence, the extent ofhybridization
oflabeledBglII
R,
toBamI BC as opposed toBamI G should be proportional to the amount
A orobe T. BgiR8: R BgIRIBgIRs
BrmnC58 U * i
8rnG
36)-Bgi Rl
9)-BaiRI (i40) I*
SamTX ii0)- 0
Sg;R, .45)-
I
I6B p Bg RIBgi
A- I I I L- I i
S S
S
R-I
9
Diot BiOrn 1/g egSv
FIG. 7. Radiofluorogramsofblotsoffragmentsof
EBV(B95-8)DNAproduced by cleavagewitha com-binationofBamI andBglII(BamI/BglII) (A)orby cleavage withBglII (B)whichwerehybridizedtothe
32P-labeledEBV(B95-8)DNA (T.P., column1)orto
labeledBglIIRfragmentortothe large (BglRd or
small(Bgl-RJfragmentobtained whenBglIIR iscut into two fragments byBamI. Thefragments which hybridizeareidentified byname,whereknown,and by sizeontheleft.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.262.457.358.575.2]322 GIVEN AND KIEFF
ofBglII
Ri
inBamI BCasopposedtoBamIGand to the distance from the left end of the
reiteratedsequence tothe BamI siteasopposed
tothe distance from theBglllsitetotheright
end of the reiteration.
Furthermore, Bglll
R.should hybridizeto BglII A andBamI G, but
not toBamI BC(Fig. 6).To estimate the relative
amountofBglII
RI
inBamIBCversusBamI Gand to further check the data shown inFig. 6,
BglIIR,, BglIIR., andBglIIR were isolated
from gels, labeled in vitro, and hybridized to
blots of BglII and BamI/BglIH fragments of
EBV DNA. The resultsare as follows. (i)
La-beled BglII R1 hybridizes extensively to the
BamI BC
fragment (Fig.
7A, column 2) andhardlyatall totheBamI Gfragment (Fig. 7A, column 2) from which BglII R. had been
re-movedby digestion withBglII and BamI. An
overlong exposureof the blot (Fig. 7A,column
3)demonstrated that thehyrbridizationtoBamI
G was barely above background. These data
indicate that the BamI site must be near the
right endof the left 1.40x106 component of the
reiterated sequence. (ii) Asexpected, BgIII
R.
hybridized extensivelytoBglIIA (Fig. 7B,
col-umn3) andhardlyatalltoBamI BC (Fig. 7A,
column 5). (iii) A surprising finding was that BglII
R,
hybridizedslightlytoBglIIR.
(Fig.7A, column4) and thatBglIIR.
hybridizedtoBglIIR,
(Fig. 7A,column5).These data suggest thatthere ispartialhomologybetween the short and
longcomponentsof the reiteration suchaswould
occuriftherewere adirectorinvertedrepeatof
partof the short segment within thelong
seg-ment. (iv)Thepartialcopy of thereiteration in
the left 4 x 105 component of BamI X has
extensivehomologyto
BglII
R. (Fig. 7A,column 5)andonlytracehomologytoBglII R, (Fig. 7A, column 4). (v) In long exposure,radiofluoro-grams of blots hybridized to labeled BglII
R,
(Fig. 7A, column 4, and Fig. 7B, column 2),
additionalfragments,BglIIIJ andBglII L,with homology to the labeled BglII
R,
preparationcould beseen.BglII IJ isalso apparent inlong
exposures ofBglII blots hybridized to labeled
BglII R, BamI V, or EcoRI/HsuI Afragment
(see Fig. 8) and probably contains DNA with
homology to BglII
RI. BglII
L, however, wasnot apparent in long exposures ofBglII blots
hybridized to labeled BglII R, BamI V, or
EcoRI/HsuIAandisprobablyidentifiedinFig. 7B, column2, because of contamination of the
labeled
BglII
RH
preparationwithanotherfrag-ment of similarsize.
Homology between the internal
reitera-tion and DNA in the long, unique region.
Long-exposure radiofluorograms of blots of
EcoRI,HsuI, EcoRI/HsuI,orBglII fragments
which had been hybridized to labeled EcoRI/
HsuIA, BamI V, or BgIII R fragments revealed
homology between the internal reiteration and
the HsuI F (Fig. 8, column 2),EcoRIC (Fig. 8,
column 3),EcoRI/HsuI J (Fig. 8, column 5), and
BglII IJ (Fig. 8, column 6) fragments. The BglII IJfragmentis part of EcoRI C (Fig. 8, column
7).Theseresultsindicate that within the region
ofoverlapofEcoRI C and HsuI F, i.e., between
94 x
106
and 97 x 106 from the left end of EBVDNA (Fig. 1), there exists a sequence with
ho-mology to theinternal reiteration. The extent of
hybridization ofthe labeledEcoRI/HsuI A or
BamI V fragment to EcoRI C,HsuIF,BglII IJ,
orEcoRI/HsuI Jfragmentwasdetermined from
scans ofautoradiograms tobe less than 3% of
the extent ofhybridizationtoEcoRIA,HsuI A,
BglIIR, orEcoRI/HsuIAfragment,indicating
thatthere is less than 1 complete copy of the
reiteration in the EcoRI C, HsuI F, BglII IJ,
and EcoRI/HsuI J fragments. The BglII IJ
fragment has homology both to the large, BglII RH (Fig. 7B,colunm 2), andsmall, BglIIRK (Fig. 7B, column 3), component of the reiteration.
DISCUSSION
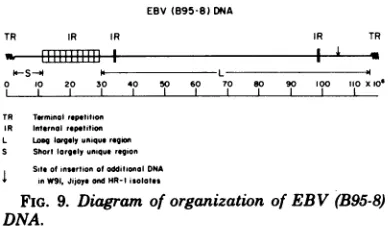
The results of the mapping of the BamI and BglIIrestriction enzymecleavagesites near the internal reiteration whichjoins the short (10 x
106)andlong (87x 106)unique regions of EBV
(B95-8) DNA are summarized in Fig. 6 and 9. The existence of short and long unique DNA
sequences joined by reiterated DNA and
boundedatthe termini by direct repeats is
sim-FIG. 8. Radiofluorograms of blots of the Hsu,
EcoRI,EcoRI/HsuI,orBglIIfr-agmentofEBVDNA whichwashybridizedtothe labeledEcoRI/HsuI A
fr-agment (E/HA),labeled EcoRIfr-agmentC(EcoRI
C),orlabeled EBV DNA(T,P).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.264.455.425.623.2]EPSTEIN-BARR 323
EBV(B95-8) DNA
TR IR IR IR TR
0 10 20 30 40 50 60 70 s0 90 100 110xlol
TR Teminol repetition IR Internol repOeition L Loo lorgelyuniquo region
S Shortlorgelyuniquo region Siteofinsertionofodditionol DNA
inW91,JijoyeondHR-I isolates
FIG. 9. Diagram oforganization ofEBV(B95-8)
DNA.
ilartothe molecularorganization of herpes
sim-plex virus (19, 23), bovine mammillitis virus (2),
and pseudorabies virus (21) DNAs. However,
EBVDNAdiffersfrom these otherherpesviral
DNAs in that there aretandem directinternal
reiterations in EBV DNA and there is no
ho-mology between the internal reiteration and
either terminus. Moreover, the short and long
uniqueregionsof EBV DNA donotinvert
rela-tive to each other during DNA replication as
has been observed with herpes simplex (23),
bovinemammillitis(2),andpseudorabies viruses
(21).
The minor KpnI (8) and EcoRI/HsuI
frag-mentsarise throughthepresencein each EBV
(B95-8) DNA preparation of molecules with a
smaller number of internal reiterations in the DNA.Thus,althoughthemajorityofmolecules
of EBV (B95-8) DNA contain 10 to 11 tandem
reiterations,somemoleculeshave 9,8,7,6, 5, or
4 tandem reiterations. We do not asyet know
whether such variantmoleculesarealways
gen-eratedduring thecourse of viral DNA
replica-tion. The findings that cells infected with the
B95-8 isolate and the cell line from which the
B95-8viruswasisolated contained circular DNA
molecules 6 x 106 smaller than EBV (B95-8)
DNA (1) support the hypothesis that shorter
DNAmolecules containing fewer copies of the
internal reiteration maybe infectious andmay
yieldprogenywithalargerorsmaller number of
internal tandem reiterations. Furthermore, the
finding that theEcoRI/HsuIAfragment of EBV
(W91) and (AG876) (N. Raab-Traub and E.
Kieff, manuscript in preparation) DNAs is
smaller than the EcoRI/HsuI A fragment of
EBV (B95-8) DNA suggeststhat these Burkitt
tumor isolates ofEBV, which have 7 x 106 of
additional DNAinsertednearthe distal end of
the long, unique region (6, 16), may have a
smallernumber ofinternaltandem
reiterations,
possibly as a consequence of the additional
uniqueDNA.
Previous estimates of the size of the HsuI A
andEcoRIAfragmentswerebasedonthe
elec-trophoretic mobilityof thesefragmentsin 0.4%
agarose gels relative to the HsuI fragments of
herpes simplex virus (MPstrain) DNA, the
larg-estfragment of which is 27.5 x
106,
orthe HsuIfragments of T5blDNA, thelargestfragment of
whichis34 x106(8). Thelog linear relationship
between molecular weight and electrophoretic
mobility in 0.4% agarose gels is parabolic for
fragments inexcessof23 x 106 to 25 x 106(8).
The HsuI A and EcoRIAfragmentsareslightly
larger than thelargest standards, and previous
estimates of the sizes of these fragments are
therefore subject to error. Three lines of
evi-dence indicate that the size of theEcoRI/HsuI
Afragment is28 x 106 and, therefore, that the
truesizes ofthe HsuIAandEcoRIAfragments
are 34 x 106 and42 x 106daltons, respectively.
First, theelectrophoreticmobility of theEcoRI/
HsuI A fragment relative to the HsuI B
frag-mentof EBV (B95-8) DNA (20 x 106 [8]), the
HsuI Bfragmentof EBV(HR-1)DNA(27x106
[8]), and intact lambda DNA (32 x 106 [22]) is
compatible withasize of28 x 106. Second, six
minorbands, eachdifferingby approximately2
x 10 in molecular weight, are clearly
distin-guishable above the EcoRI/HsuI B fragment in
blots of theEcoRI/HsuI fragments whichwere
hybridized to the labeled EcoRI/HsuI A
frag-ment.The molecularweight of theEcoRI/HsuI
B fragment is14 x 106.The size ofthe
EcoRII
HsuIAfragment,which islargerthan thelargest
minorEcoRI/HsuI Afragment, musttherefore
be28x 106. Third, the molecular weight of the
unique-sequencecomponentoftheEcoRI/HsuI Afragmentis8.5x106.The size of the reiterated
component, BamI VorBglIIRfragment,is1.85
x
101;
to 1.9 x 106. TheBamI Vfragment com-ponent is present in at least 10-fold excess ofother BamIfragments(17).
The revisedestimates of the sizes of the HsuI
A and EcoRI Afragments suggested by these
databringthesum of themolecularweightsof
the EcoRI or HsuI fragmentsof EBV (B95-8)
DNAto 116x 106, which isapproximately 10%
largerthan estimates basedon ameasurement
of thelengthofEBV DNA (1,7, 15). Although
the latter estimates of the molecularweight of
EBV (B95-8) DNAarebasedultimatelyonthe
length of 4X174 DNA, a molecule of known
molecular weight, there are several possible
sources of error in the determination of the
molecular weight of EBV DNA from its length,
the most significant of which is the possibility that thehigherguanine-plus-cytosinecontentof EBV DNA results in a higher mass per unit
lengththan thatof4X174 replicative-formIIor
PM2 DNA.
The significance of theinternaltandem
reit-eration, of the homology between the
compo-nents of the reiteration which is likely to be a consequence of a reiteration within the
reitera-VOL. 31,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.504.56.250.70.183.2]tion,and of thehomologousDNAsequences in
theBamIX andHsulFfragmentsof thelong,
unique region is uncertain. From the relative
hybridization of labeled BamI V orBglII R to
theHsuIFand HsuI Afragments,thelengthof
the regionofhomology between the reiterated
DNA and HsuI F fragmentis estimated to be
less than 5x 105,whereas the component of the
BamI Xfragment homologous to the internal reiterationiscontained in the4 x 105 region of
BamIX fragment to the left of the HsuI A-B
site. TheHsuI F sequences homologous to the
internalreiteration havemorehomology tothe BglII
RI
componentthantotheBglIIR.
com-ponent,whereas thereverseistruefor BamI X.
Itis not known whether the DNA in BamI X
and HsuIF,which ishomologoustotheinternal
reiteration, is a continuous sequence or is
sepa-ratedby regionsofnonhomologyorwhetherthe
homologoussequenceswithin the reiterationor
in BamI X and HsuI F are direct or inverted
repeats.Regionsofhomologyin the DNA could
play aroleinDNAor RNA
synthesis
or couldbeaconsequenceof gene
duplication.
Pertinenttothe latter is theobservation that the internal
reiteration encodesstable
polyadenylated
RNAinBurkitttumortissueand themostabundant
RNA in
restringently
infected Burkitt tumorcellsgrowninculture
(3,
14).
ACKNOWLEDGMENTS
NancyRaab-Traub,WalterKing,and MarkHeller collab-oratedon someof theseexperiments. MaryHawke andNancy
Treptowprovidedexcellentassistance.
This research was supported by Public Health Service grantsCA 19264-03andCA 17281-04 from the National Cancer Institute and grant VC 113D from the American Cancer Society. D.G. isa predoctoraltrainee supported byPublic Health Service grant HD 07009-03.
LITERATURE CITED
1. Adams, A., G.Bjursell, C.Kaschka-Dierich,and T. Lindahl.1977.CircularEpstein-Barrvirus genomes of reducedsize inahumanlymphoidcell line of infectious mononucleosisorigin.J.Virol.22:373-380.
2.Buchman, T.G., andB.Roizman. 1978.Anatomyof bovinemammilitisDNA.I.Restriction endonuclease mapsof fourpopulationsofmolecules thatdifferin the relative orientation of their long and short components. J.Virol.25:395-407.
3.Dambaugh, T., E.Kieff,F. K.Nkrumah, and R. J. Biggar.1979.Epstein-Barrvirus RNA. IV. Viral RNA inBurkitt tumor tissue.Cell16:313-322.
4. Delius, J.,and G. W. Bornkamm.1978.Heterogeneity ofEpstein-Barr virus.III.Comparisonofatransforming and anontransformiing virus by partial denaturation mapping of their DNAs. J. Virol.27:81-89.
5. Dolyniuk, M.,R.Pritchett,and E. Kieff. 1976. Proteins ofEpstein-Barrvirus.I.Analysisofthepolypeptides of purifiedenvelopedEpstein-Barr virus. J. Virol. 17:935-949.
6. Given, D., and E. Keiff. 1978. DNA ofEpstein-Barr virus. IV. Linkage map of restriction enzyme fragments of the B95-8 and W91 strains ofEpstein-Barr virus. J. Virol. 28:524-542.
7.Given, D.,D.Yee,K.Griem,and E. Kieff. 1979. DNA of Epstein-Barr virus. V. Direct repeats of the ends of Epstein-Barr virusDNA. J. Virol. 30:852-862. 8. Hayward,S. D.,andE. Kieff. 1977. DNA of
Epstein-Barr virus. II. Comparison of the molecular weights of restrictionendonuclease fragments of the DNA of Ep-stein-Barr virus strains and identification of end frag-mentsof the B95-8 strain. J. Virol. 23:421-429. 9. Kelly,R.B., N. R.Cozzarelli,M. P. Deutscher,L.R.
Lehman, and A. Kornberg. 1970. Enzymatic synthe-sisof deoxyribonucleic acid. J. Biol. Chem. 245:39-45. 10.Kieff,E., D.Given,N. Raab-Traub, A. L T.Powell, W.King,and T.Dambaugh. Nucleicacid of EBV. Biochim. Biophys. Acta Rev. Cancer, in press. 11. Kieff, E.,and J.Levine.1974.Homology between
Burk-ittherpesviral DNA and DNA incontinuous lympho-blastoidcells frompatientswith infectious mononucle-osis. Proc. Natl.Acad.Sci.U.S. A. 71:355-358. 12. Marmur,J. 1963. A procedure for isolation of DNA from
microorganisms.Methods Enzymol. 6:726-738. 13.Nonoyama, J. M., and J.S.Pagano. 1971. Detection of
Epstein-Barrviral genomein nonproductivecells. Na-ture(London)NewBiol. 233:103-106.
14.Powell, A.LT., W. King,and E.Kieff. 1979. Epstein-Barrvirus-specific RNA.III. Mapping of DNA encoding viral RNA in restringent infection. J. Virol. 29:261-274. 15.Pritchett, R. F., S. D. Hayward, and E. D. Kieff. 1975. DNA ofEpstein-Barrvirus. I.Comparative studies of the DNA ofEpstein-Barrvirus fromHR-1 and B95-8 cells:size,structure,and relatedness. J. Virol. 15:556-569.
16.Raab-Traub, N.,R.Pritchett,and E. Kieff.1978.DNA ofEpstein-Barr virus. III.Identificationof restriction enzymes that contain DNA sequences which differ amongstrains ofEpstein-Barrvirus. J. Virol.27:388 398.
17. Rymo,L,and S.Forsblum.1978. Cleavage of Epstein-Barrvirus DNAbyrestriction endonucleases EcoRI, HindIIIand BamI. Nucleic Acid Res. 5:1387-1402. 18. Schulte-Holthausen, H., and H. zur Hausen. 1970.
Partialpurificationof theEpstein-Barr virus and some propertiesof its DNA.Virology40:776.
19.Sheldrick, P.,and N. Berthelot. 1974. Inverted repeti-tions in the chromosome ofherpessimplexvirus. Cold SpringHarborSymp.Quant. Biol.39:667-678. 20. Southern, E. M. 1975. Detectionofspecific sequences
among DNAfragmentsseparatedby gel electrophore-sis. J. Mol. Biol.98:503-517.
21. Stevely, W.S. 1977.Invertedrepetition in the chromo-someofpseudorabiesvirus. J.Virol. 22:232-234. 22. Thomas,M.,and R.Davi8.1975.Studiesonthecleavage
ofbacteriophagelambda DNA with EcoRI restriction endonuclease. J. Mol. Biol.91:315-328.
23. Wadsworth, S., R. J. Jacob, and B. Roizman. 1975. Anatomy ofherpessimplex virus DNA. II. Size, com-position,andarrangement of invertedterminal repeti-tions. J. Virol. 15:1487-1497.
24. Wagner, E. K., B.Roizman, T. Savage, P. G. Spear, M.Mizell,F.Darr,and D.Sypowiez.1970. Charac-terizationof the DNA of the herpesvirus associated with Lucke adenocarcinoma of thefrog and Burkitt lymphoma of man. Virology 42:257-261.
25. Weinberg, A., andY.Becker.1969.StudiesonEB virus ofBurkitt'slymphoblasts. Virology 39:312-321.
on November 10, 2019 by guest
http://jvi.asm.org/