0022-538X/79/07-0104/08/$02.00/0
Polymorphism
of the
Migration
of
Double-Stranded RNA
Genome
Segments
of Reovirus Isolates
from
Humans, Cattle,
and Mice
DANIEL B. HRDY,12 LEON ROSEN,3ANDBERNARD N. FIELDS1' 4*
Department of Microbiology and MolecularGenetics, Harvard MedicalSchool, Boston,Massachusetts
02115,' Department of Anthropology,HarvardUniversity, Cambridge,Massachusetts02138,2 Pacific ResearchSection,NationalInstituteof AllergyandInfectious Diseases, Honolulu,Hawaii96806,3and
Divisionof Infectious Disease,Peter BentBrigham Hospital, Boston,Massachusetts 021154
Received forpublication20December 1978
A series of 94 isolates of reovirus from humans, cattle, and mice showed
extensivevariabilityin thepatternsofmigrationof the ten double-stranded RNA
genomesegments.This variationwasfound inallthreeserotypes, and involved alltengenomesegments, including the segment responsible for serological
spec-ificity. Although a single pattern was present among several samples isolated
from individuals and collectedatasingletime andplace,therewereoftenmultiple
genetic variants ofa single serotype present ina population. Samples isolated
from widely different geographic origins ordifferent mammalian hosts showed
differentpatterns;samplesfromasingle speciesfromthesamearea over aperiod
oftime showed more limited variations.Among mostisolates, the migration of
the slowest S segment, the segment that encodes the hemagglutinin and is
responsible for serological specificityinlaboratory strains,wassimilar to reference
strains for type 1 and type 3 isolates. However, the type 2 isolates showed
considerable variationinthissegment.
Mammalian reovirusesarewidely distributed
in humansandanumber of animalspecies (21).
Although frequently isolated, their role in
hu-man disease isunclear. In most instancesthey
appear toproduce asymptomatic enteric or
res-piratoryinfections (8). Inspiteof this presumed
minor role inhumandisease, reovirushasbeen
thesubject of intensivestudy primarilybecause
of itssegmenteddouble-stranded RNA(dsRNA)
genome. Asthe list of othersegmented
dsRNA-containing viruses has expanded, ithas taken on additionalsignificanceas amodel system.Other
genera within thefamilyReoviridaeincludethe
insect-borne orbiviruses, phytoreoviruses of plants, cytoplasmic polyhedrosis virus of insects,
andthemedically important rotaviruses, which
arecurrently refractory to detailed study incell
culturesystems (15, 28).
The three serotypes of mammalian reovirus
showconsiderable heterogeneity in
electropho-retic migration among both the dsRNA
seg-ments and viral polypeptides (19). The ten
dsRNA segments migrate at different rates on
polyacrylamidegels and have been classified as
L (large, three segments), M (medium, three
segments), and S (small, four segments) (14, 17). Differences in the migration of these RNA
seg-ments between laboratory strains formed the
basis for identifying viral recombinants derived from different serotypes (19). It was subse-quently foundthat oneof the dsRNAsegments, S1, encodestheviralhemagglutinin (theal outer capsidpolypeptide) (17,27).Thehemagglutinin
reacts with type-specific neutralizing antibody
and is thus the antigen involved in serotype
specificity (26).
Although traditional analysis ofviralstrains
in nature has depended on serological studies,
the availability of electrophoretic analysis of nucleic acidshasbeguntoplayan increasingly important roleinepidemiological analyses. This
method permits precise analysesof segmented
RNAviruses such as reovirus (19), rotavirus (9),
orbivirus (B. M. Gorman, J. Taylor, P. J.
Walker, and P. R. Young, J. Gen. Virol., in
press), and influenza (18), aswellas a
compara-ble analysis for the nonsegmented DNA virus
herpes simplex, using restriction enzyme DNA fragments(13).
Access to alarge number of isolates of reovirus
collected during field investigations in the late
1950sandearly 1960s from severalparts of the
world(L.R.) allowed us to evaluate several
ques-tions. (i) How much variability exists within a
series of isolates of a single serotype from one
mammalianspecies? (ii) How much variability
104
on November 10, 2019 by guest
http://jvi.asm.org/
REOVIRUS RNA SEGMENT
exists withina serotype amongisolatesthat
orig-inate from different species? (iii) Do isolates
within asingle outbreak show variability?(iv) Is
thereevidenceof consistent patterns defined by
geographic areas? (v) Can the dsRNA pattern be used to reliably predict the serotype? The results of the study indicate striking polymor-phisms of migration of dsRNA segments and have provided preliminary answers to these questions.
MATERIALS AND METHODS
Reovirus isolates. Human isolates of reovirus
were collected from two sites: the respiratory tract
(throat swab) and the gastrointestinal tract (anal swab
orstool). Murine and bovine isolates were collected
only from feces. Strains were originally isolated by inoculation in primary rhesus kidneycellcultures fol-lowed by a single passage in the same cell (22). Lysates of these cultureswerestoredat-70°C until use. The
samples analyzedarelistedinTable 1with date and
placeoforigin.Asinglerecentstrain was isolated as
acontaminant from human kidney cell culture and wasprovided by Collaborative Research, Inc., Wal-tham, Mass.
The bovinesampleswerecollected from three Mar-yland herds-Jessup, College Park, and Central Farm-the lattertwo of which exchanged members (23).Samples from Toluca, Mexico; Tahiti; themurine
isolates;human isolate HTlb fromWashington, D.C.;
and thecell culture contaminantweretyped by neu-tralization reduction by D.B.H. as outlined below. Samples typedastypes1or2by neutralization were confirmed by hemagglutination inhibition by the method of Rosen (20). The balance of the samples
weretyped by hemagglutination inhibition by L.R. as
previously reported(22)atthe time of collection.
Since all but one of the samples were collected many years ago,allsamples were titrated before pas-sagetodetermine thecontentof infectious virus. The titers varied from <10' PFU/ml (in approximately
one-third ofsamples)to>107(twosamples). One-half
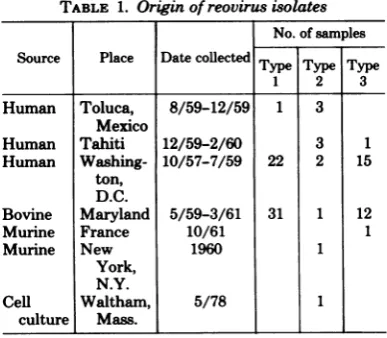
TABLE 1. Originofreovirus isolates
No. ofsamples
Source Place Datecollected Type Type Type
1 2 3
Human Toluca, 8/59-12/59 1 3
Mexico
Human Tahiti 12/59-2/60 3 1
Human Washing- 10/57-7/59 22 2 15
ton, D.C.
Bovine Maryland 5/59-3/61 31 1 12
Murine France 10/61 1
Murine New 1960 1
York, N.Y.
Cell Waltham, 5/78 1
culture Mass.
of the samples had titers of 2104 PFU/ml, attesting to the extraordinarily long-term stability of these viruses when stored at low temperatures.
Cells and preparation of virus stocks. Lysates of original virus isolates were passaged in mouse L cells. The L cells were maintained in suspension
cul-ture in Eagle minimal essential medium (MEM) as
modified by Joklik (Industrial Biological Laboratories) supplemented with 5% fetal calf serum. To prepare stocks, lysateswereinoculated onto confluent mono-layers of mouse L cells inT25flasks (Corning Glass Works, Corning, N.Y.) at 37°C, observed for cyto-pathiceffect(CPE), and harvested by freeze-thawing. With lysates containing little virus, if CPEwas not observedby14days, theflask was harvested. A second passage was made, harvested when CPE appeared, and usedasstock for dsRNAlabeling.
Preparationof labeled cytoplasmic dsRNA. A
modification of the method of Ramig et al. (19) was usedtopreparelabeledcytoplasmic dsRNA. Briefly, Lcellswereinoculated with 1.0ml of passaged viral suspension, and 200 uCi of3P was added in MEM with 0.25pgofactinomycin D per ml. The cells were incubated for2days and harvested by scraping. The cellswerethen treated withTriton X-100 and centri-fugedat 1,200 rpmfor5 min, and the supernatants werediscarded. NaCl and ethanol were added to the
pellets, and the mixtures were stored ovemight at
-20°Ctoprecipitate dsRNA. Thesampleswerethen centrifuged at 12,000 rpm for30minin aSorvall SS-34,dried under vacuum, anddissolved in0.25ml ofgel
samplebuffer (10).Althoughthe viral dsRNA in these
sampleshasnotbeencompletely deproteinized,ithas
previously been reported that RNA extracted from
purifiedvirions andprocessed with extensive
depro-teinization, migratestothe same extentduring
elec-trophoresisasdsRNAprocessedfromcytoplasmic
ex-tracts (19).Thus,the mobilityofdsRNAwhen
pre-pared bythese methods isnotaffected by
contami-nating proteinremaining inthesample.
Polyacrylamide gelelectrophoresis of dsRNA.
Slab gels were prepared to a final concentration of
10% acrylamide-0.267%bisacrylamideasdescribedby
Laemmli (10).Electrophoresisofsampleswhich had been heated at 100°C for 1 m wascarried out at a
constant currentof 20 mA for 18 h. Unfixedgelswere
driedontofilter paper inaHoefferScientific
Instru-mentsgel dryer,andautoradiographywasperformed
byexposingNo-ScreenX-rayfilm(Kodak) for1 to2
days.
Neutralizationassay.Antiseraagainstthe three
reovirus serotypeswere obtained from the National Institute ofAllergy and Infectious Diseases (catalog no.V-701-511-570;V-702-501-570-,andV-703-501-570).
Approximately 100 PFU of the unknown virus was
incubated with twofold dilutions of antisera for45min at37°C, thenplatedat37°ConLcellmonolayersby
the method of Fields andJoklik(4).Greater than 80% reduction ofplaqueswasconsidered neutralization of virus.Serological classificationwasdeterninedby
he-magglutination inhibition (20) or neutralization.
Al-thoughnot everysamplewasanalyzed byboth
sero-logical tests,samplesdisplayingrepresentative dsRNA
patterns were tested with freshly grown stocks to
verifythe initialserologicalclassifications.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.508.55.249.489.658.2]106 HRDY,
RESULTS
Nomenclature of dsRNA segments. Pre-vious studies have correlated the dsRNA seg-ments between laboratory strains of the three serotypesofthe mammalian reoviruses (25). Al-though in general thesegmentsof the serotypes correspond in order ofincreasingelectrophoretic mobility, thiswas notalwaysthecase(seefig.6 in ref. 25). Because of this problem, we have
arbitrarily labeled dsRNA segmentsofthe iso-lates reported here by sequentially numbering
from large to small segments in a size class. These segments are referred to in the text in parentheses. Thesegmentsfrom thelaboratory
strainsarelabeledwithoutparenthesesby prior
nomenclature asshown in Fig. 1A, 2A, and 3A. Type1reovirus.(i)Humanisolates. Type
J. VIROL.
1 reovirus isolates, designated TM and WA,
respectively,wereisolatedfromhuman
materi-als at Toluca, Mexico, and Washington, D.C.
(Table 1).There was asingleTMisolateand 22 WAisolates. Oftheseisolates, therewerethree RNApatterns: (i)TM(HTla)(Fig. 1B), (ii)one
WA isolate (HTlb) (Fig. 1C), and (iii) the re-maining 21 WA isolates (HTlc) (Fig. 1D). Al-thoughthe overallRNA patternsweresimilar, certain segments showed considerable variabil-ity. Although there were minor variations in
migrationsof the Lsegments[for example,note L(2) and L(3) of HTla (Fig. 1B)] migrating slower than thecorrespondingsegmentsoftype 1 (Lang) (Fig. 1A), themajorheterogeneitywas
noted in the M segments.Themigrations ofthe
M(1) segments,forexample,weremarkedly
dif-ferentfrom each other(Fig.1B, C, and D).
i
q
t: 3
t:i
(1:
[::)e
-.U.P. r
L.1 L.2
13
o. M2
aAI. .0
MS
S2
54 S3
FIG. 1. 32P-labeleddsRNA from typeIreovirusisolates.Electrophoresis wasfromtop to bottom in a single
10% acrylamide gel containing the Tris-glycine buffer system as described in the text. (A) TypeI(Lang); (B) human type HTla, TM; (C) human typeHTlb, WA; (D) human type THlc, WA; (E) bovine typeBTla, CP; (F) bovinetypeBT1b,CP;(G) bovine typeBT1c,JE; (H) type1(Lang). RNA genome segments of type1(Lang)
areidentified (17).
A;h ':'D"M
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.508.72.452.267.600.2]The 21 WA isolates showing identical HTlc patterns (Fig. 1D) were isolated in early 1957, whereas thesingleHTlbpatternwas isolated in
1959. Thus, a virus isolatedfrom one geographic
areafromalargenumber ofindividuals retained a distinct pattern. However, within this same geographicarea a secondclearly distinctpattern
was seen at alater date (HTlb) (Fig. 10).
Fur-thermore, the single isolate from Mexico (Fig. 1B)wasquite distinct fromall the WAisolates. (ii) Bovine isolates. Type 1 reovirus was isolated fromthreeherdsinMaryland over the 6-month period from November 1959 through May 1960(Table1).TheCollegePark and Cen-tral Farm herds, which were separated by 25 miles, frequently exchanged members and are therefore treated as a single herd (CP). The Jessup herd (JE) did not exchange members with the other herds, and was about 15 miles distant from each of them. Ofthe 31 isolates,all samples displayed oneof threepatterns;BTla, 15isolates; BTlb,11isolates; BTlc, five isolates (Fig. 1E, F, and G). Note thecomigration of the L(2), L(3) and M(2), M(1) dsRNA segments in the BTlb pattem (Fig. 1F). The comigrationof
segments results in the appearance of a more
intenseradioautograph bandaswellas the
ap-pearance ofa "missing" segment(s)
[L(3)
andM(2) in Fig. 1F]. The 26viral isolates showing BTla and BTlbpatternswereisolated from the CPherds; the BTlcpatternwas seenexclusively
amongthe JEisolates.Asin thehumantype 1
isolates, there were marked variations in the migration of RNA segments. These included
major variations inmigration among segments ofall size classes: Lsegments (Fig. 1F); M
seg-ments,especiallyM(1) and M(2) (Fig. 1E, F, and
G); and S segments (compare Fig. 1F and G).
Although the overallpatternresembled that of the humantype 1isolates,noconsistentpattem thatcouldidentifyanisolateasbovineinorigin wasdetected. Thesmallest variationswere pres-ent in the M(3), S(1), and S(2) segments.
Al-though all of these 31 isolates were classified
serologically as type 1, the RNA analysis pro-videdadditional data. Viruses classifiedastype 1butcontainingtwodifferent patterns ofRNA
were present in the CP herds (Table 2). The
BTlapattem waspredominant during
Novem-ber1959, but by March 1960 there was a shift to
the BTlb pattern (8 of 10 samples). By April
1960, six of sevensampleswere BTla. The last
isolate(May1960) wasBTlb. Althoughthere is
no way to study the precise origin of these
distinctisolates from availabledata,it ispossible
that thetransfer ofanimalsbetween theCollege
Park andCentralFarmherds wasimportantin
exchanging thesetwodistinct viruses. Studiesof
TABLE 2. Place and date of origin of bovine type 1 patterns
Pattern
Date Herd'
BTla BTIb BTIc
11/59 CP 7 1
JE
3/60 CP 2 8
JE 2
4/60 CP 6 1
JE 3
5/60 CP 1
JE
aSee text.
thistypewill beimportant in futureanalysesof suchoutbreaks andmayprovide importantfacts in theanalysisof virusspread.
Type
2reovirus.(i)Humanisolates. There were eight human isolates of type 2 reovirus obtained from Toluca, Mexico, Tahiti (desig-nated TA), and Washington, D.C. (Table 1).Twopatterns werenotedamongthe threeTM
isolates (Fig. 2B andC),asinglepattem among the three TA isolates (Fig. 2D), and a single
pattem amongthetwoWAisolates(Fig. 2E).As
previously noted with the type 1 isolates,there was marked heterogeneity in the migration of several RNA segments. This was especially
prominentamongtheMand Ssegments.
(ii) Nonhumanisolates.There were single
isolates of bovine(CP herd, Fig.2F),andmurine
origin (Fig. 2G) as well as a contaminant of unknown species origin from cell culture (Fig.
2H). Each of these isolates was distinct, with variations inmigrationseeninseveralsegments. Thus, althoughthe number oftype 2 isolates isquitesmall,it isapparentthatthere isa
large
number of distinct patterns with major
differ-ences in migration noted in each of the RNA
size classes.
Type
3reovirus.(i)Human isolates.Type
3reoviruswasisolated from human materialsat
Tahiti and Washington, D.C. (Table 1). There
were asingle TA isolate (Fig. 3B) and 15 WA
isolates. The 15 WA isolates showed two pat-terns:HT3b in2isolates(Fig.
30),
andHT3c in13 isolates (Fig. 3D). HT3cdiffers from type 3
(Dearing) mainly in the M(1) segment. The
HT3bpattern isvirtuallyidenticaltothatof the
Dearingstrain, except foraslightdifference in L(1) migration. The TA sample
(HT3a)
(Fig.
3B) differs from these strains in several
seg-ments,mostnotablytheL(3), M(1),and
S(3,4).
Thustheheterogeneityamong type 3
samples
on November 10, 2019 by guest
http://jvi.asm.org/
108 HRDY, ROSEN, AND FIELDS
A
B
C
D
E
L 1 e`s _."
L23 L 3 "
F
G
H
U
M2
_M3
-00i~
._....;
3S2 :::
:43 :
FIG. 2. 32P-labeleddsRNAfrom type 2 reovirus isolates. Electrophoresis was from top to bottom in a single
10% acrylamide gel containing the Tris-glycine buffer system as described in the text. Lanes have been rearranged for comparison. (A) Type 2 (Jones); (B) human typeHT2a, TM; (C) human type HT2b, TM; (D) human typeHT2c, TA; (E) human typeHT2d, WA; (F) bovinetypeBT2a,CP; (G) murine type MT2a, New York, N.Y.; (H)cellculturecontaminant, type2, Waltham, Mass.; (I) type 2 (Jones). RNA genome segments
oftype2(Jones)areidentified(17). Notethat the type 2 (Jones)strain usedasamarker was slightlydefective (containing reducedamountsof Ll). The mobility of this and the other segments was, however, identical to thatofnondefectivepreparations (19, 25).
appears somewhat more restricted than that
amongthetype 1 or 2samples,but this may be
duetosmallsample size.
(ii) Bovine isolates. There were 12 bovine isolates oftype 3reovirus: 6 from theCPherd
and 6 fromthe JE herd. Three patterns of RNA
were seen: BT3a (two isolates) (Fig. 3E) and
BT3b(four isolates) (Fig. 3F) from the CP herd, and BT3c (six isolates) (Fig. 3I) from JE. Al-though there were minor variations in RNA migrationsbetween the BT3aand BT3b isolates, such as in L(1,2), M(2), and S(1,4), the BT3c patternwasmarkedlydifferent.TheL(1,2) and M(1) RNA segment species among the latter isolateswereespeciallydifferent. Thus, the iso-lates fromtheCPherd weredistinctbutshowed
only slight variations, whereas those isolated from the second herdwereconsiderablyaltered. (iii) Murine isolates.Asinglemurine isolate
oftype 3 (Fig. 3G) was distinct from the other
isolates, with minor variationsin migration of
numeroussegments.
Comparison of human respiratory and
gastrointestinal isolates. Of the 47 human isolates,42 werefromgastrointestinaland 5 were from respiratory sources. Of these five, there
were one type 1 and four type 3 isolates from
WA. The patterns of these isolates were the
sameasvirusesisolatedatthe sametime from
gastrointestinalsources.The type 1 isolate had the HTlc pattern (Fig. 1D), which was also
found in all concurrently isolated type 1
gas-
.1410wai"-qMP: -AMM,.(
1-Almosiak.
'o
..41 ANN
"OOka-.mwmw
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.508.58.467.69.402.2]RNA SEGMENT MIGRATION POLYMORPHISM
A
B
C
D
E
F
G
H
I
Ll _
L2
L3
ml -,
M3
Si-_
S2 .
s
-e
-ac_e
- C_I
U_
a
._1
--__ _
MAW~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
*1 ...*l
*SO-FIG. 3. 32P-labeleddsRNAfrom type 3 reovirus isolates. Electrophoresis was from top to bottom in a single
10%acrylamidegel containing the7ris-glycine buffer system as described in thetext. (A) Type 3 (Dearing);
(B)human type HT3a, TA; (C) human type HT3b, WA; (D) human type HT3c, WA; (E) bovine BT3a, CP; (F) bovine typeBT3b,CP; (G) murinetypeMT3a, France; (H)type 3 (Dearing);(I) bovinetypeBT3c, JE. RNA genomesegmentsoftype3(Dearing)are identified (17).
trointestinal viruses. Similarly, all four type 3 respiratory isolates were HT3a (Fig. 3B). This
wasthe pattern most commonly isolated from gastrointestinal type 3samples from the same time and place. Thus, the site from which the viruses wereisolated did not appear to play an important role.
Correlation of S(1) segment and
sero-type.The
Si
dsRNAsegmentof type 3(Dear-ing) reovirus migratessignificantlyslower than
that of type1 (Lang) or type 2 (Jones). The Si
segmentsfromthe lattertwo strains aredifficult
todistinguishfrom each other. This finding is of
some potentially predictive importance, since
the
Si
segmentencodes the viral hemagglutinin,thetype-specific polypeptide(27). A preliminary determination of serotype could be made by analysis of the migration of S(1) segments in
certain instances. The S(1) segment oftype 1 strainsmigrates faster than thatof type 3.Thus,
in every instance, the S(1) segment of type 1
migrated closetotheS(2)segment(Fig. 1),while that oftype3migrated considerably slower (Fig. 3). Instrikingcontrast totherelatively
consist-ent pattern seen with type 1 or 3, the S(1)
segmentoftype2migrateswith enormous
var-iability. For example, the S(1) segment ofone isolate of TM migrates very slowly (Fig. 2B),
whereas theS(1) of the TA isolatemigratesas
fastastheSiof type3(Fig. 2D). Thus, although
the migration of the S(1) segment has some predictive value for type 1 or 3 strains, the variability of migration of S(1) among type 2 isolates indicates the limitations of this ap-proach.Thesignificanceof the marked variabil-ityinS(1) migrationof type2strains is
poten-:.
..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.508.66.443.69.411.2]tially of considerable interest but is currently
notunderstood.
DISCUSSION
The analysis of the pattems of the rate of migration ofdaRNAgenome segments among a number of isolates of reovirus indicates that there are majorvariations in the
migration
of each segment. The analysis of these data has provided insights into features of the intrinsicgenetic variability of the reovirus genome and therelationshipof thisvariabilitytothe molec-ularepidemiologyof the m mlianreoviruses. There arethree serotypes of themammalian reovirus, and serological variability within the
type 2serotype has been previously described (6).We havereported differencesinthepatterns ofmigrationof dsRNA segments between pro-totypemembers ofeach of thethreeserotypes (19). It is apparent from the data described in this study that thereis nosingleRNApattern
thatdistinguishes aparticularserotype. All ge-nomesegments appear to possess some variabil-ity.Infact, the actualextentof
polymorphism
isprobably even higher than that indicated by variation in the migration ofthe dsRNA seg-ments, sincedifferencesdue to base pair substi-tution or limited deletions might not be de-tected. Itshouldbe stressed that thevariability
in mobilityof the RNA segments between iso-lates is notsimplydue tochangesinviral RNA
occurringas aresultofselectionofviralvariants
duringpassageandgrowthofviralstocks in the
laboratory. The pattern of migration of RNA segmentsremains stablethroughoutseveral pas-sages(R.
Ahmed,
personal communication),and thusthe differences betweenisolatesseeninthis study could not have been generated after theone or two passages required to develop
high-titered stocks.
Thisvariability in migration of dsRNA
seg-ments isseen among several isolates within a
serotype, amongthe three serotypes, and among
isolates derived from different species or
geo-graphic areas. Thus, in comparing the RNA
pattern of isolates of reoviruses of different origins, the RNA pattem does not identify the serotype, host of
origin,
orgeographic locale.In spite of theextraordinary polymorphisms inthe overall rates ofmigration of most of the dsRNA segments, two points ofinterest have
clearly emerged. Although it had previously been noted that more than one serotype can exist in apopulation during a single outbreak (21), it is now apparent that multiple genetic
variants of a single serotype may be present
during a single outbreak. This was best
illus-tratedbythetwobovinetype1isolates obtained
from the CP herds. The identification of these two discrete isolates and their emergence and
disappearance over the 2-year period provide importantepidemiologicaldata notsuspectedat the time that thesesampleswereinitially
ana-lyzed.This type of detailedanalysis will clearly
provide additional data in future studies. Recent
analyses of rotavirus outbreaks (3,9) have
re-portedevidence formultiplevariants inasingle humanpopulation basedonRNA patterns. Se-rological data on these outbreaks have not yet beencorrelatedwiththese RNA patterns.
Theanalysisofmigrationof theS(1) dsRNA segments of the three serotypes has revealed anotherinteresting finding. Since this segment
encodes the viral hemagglutinin in laboratory
strainsand is thetype-specific dsRNAgenome segment(27), theanalysisof the rate of
migra-tion of this segmentwasofparticular interest.
The finding that there are relatively constant
migrationsof theS(1)segments amongdifferent
type 1and3isolates is consistent withthe fact
that this segment is responsible for serotype
specificity.
Thestrikingvariabilityin the rate ofmigrationof the S(1) segment seen among the
limitednumber of type 2isolates suggests that the hemagglutinin of these isolates is highly variable,consistent withthe previousfinding of
serological subtypes among type 2 strains (6).
The chemicalnatureof thisvariabilityand ge-netic basis isnotunderstood.It ispossiblethat type 2,the leastfrequentlyisolatedofthethree
reovirus serotypes(23),representsan evolution-arily unstable serotype. Studies underway on the RNA sequence of these isolates as wellas
biological properties of the hemagglutinin of type 2 (such as cell and tissue tropism) may provide important insights into the biological significanceofsuchvariation.
Itshould be pointed out thatvariabilityinthe genetic structure of the mammalian reoviruses is notunique. Otherviruses whose genome
struc-tures have recently been analyzed have also
shownconsiderablevariability.Forexample,the DNAs ofherpesvirusesafter endonuclease treat-ment have recently been reported to vary
be-tweenisolates(13).Influenza virus has also been
shown tovary in themobilityofgenome RNA
segmentsinvarious strainsisolatedindiffering
hosts andgeographic areas (18, 24). Even labo-ratory-adapted viruses possess genetic variation; RNAphage QB hasrecentlybeen showntohave large nucleotide sequence heterogeneity in a sin-gle population (2). The genetic variability of reovirusisolates and other viruses isthe coun-terpart ofprotein variation reported for other
organisms,including Escherichia coli (16),
dro-sophila (12), and humans (5). Detailed studyof
on November 10, 2019 by guest
http://jvi.asm.org/
the genetic variability of viruses in a natural population should elucidate the amount of var-iability necessary to allow rapid evolutionary change while maintaining a stable virus popu-lation.
Finally, isolates from different sources do not reveal consistent differences in RNA patterns betweenspecies, although this does not prove that species-specific patterns might not exist. Rather, reovirus isolates seem to represent a
genetically heterogeneous group in all species, ashasbeen reported for influenza (7). Of more significance, we have not been able to determine whether genome segment exchange takes place betweenhuman and nonhuman species, as has beenpostulated tooccur ininfluenza (11). Such geneexchangeclearlycouldhelp rapid evolution by exchange of genetic material and assist in the maintenance ofanonpersistent virus in small
host populations (1). In addition, the possibility that mammalian reoviruses infect multiple
mamnmalian species under natural conditions could play asimilarrole in their maintenance in apopulation.
ACKNOWLEDGMEN1S
This work was supportedby research grant R01 AI 13178 from theNational Institute ofAllergyandInfectious Diseases. D.B.H.wasthe recipient ofPublic Health Servicefellowship
1 F32 AI06673from the National Institute of Allergy and
InfectiousDiseases.
Wethank Susan LPevear,Linda Powers, R. Frank Ramig, DonaldRubin, Howard LWeiner,and Roz White for
inval-uableaasistance.
LITERATURE CITED
1. Black,F.L 1975.Infectious diseasesinprimitivesociety.
Science 187:515-518
2. Domingo, E.,D.Sabo, T. Taniguchi,andC.
Weis-mann. 1978.Nucleotidesequenceheterogeneity of an
RNAphage population. Cell 13:735-744.
3. Espejo, R.T., E.Calderon, and N. Gonzalez. 1977.
Distinctreovirus-likeagentsassociated with acute
in-fantilegastroenteritia.J.Clin.Microbiol. 6:502-506.
4. Fields,B.N., and W. K.Joklik. 1969. Isolationand
preliminary geneticand biochemicalcharacterization of
temperature-sensitivemutantsof reovirus.Virology37:
335-342.
5. Harris,H. Enzymepolymorphismsin man. 1966.Proc.
RSoc. Br. 164:298-310.
6. Hartley,J.W.,W. P.Rowe,and J. B.Austin.1962.
Subtypedifferentiation of reovirus type 2 strains by
hemagglutination-inhibition with mouse antisera.
Vi-rology16:94-96.
7.Hinshaw,V.S.,W. J.Bean,R.G.Webster,and B.C.
Easterday. 1978. The prevalence of influenza viruses inswine and theantigenic and genetic relatedness of influenzavirusesfrom man and swine.Virology 84:51-62.
8. Jackson, G. G., and R. L Muldoon. 1973. Viruses
causing common respiratory infection in man. IV. Reo-viruses and adenoReo-viruses. J. Infect. Dis. 128:811-822. 9. Kalica, A. R., M. M.Sereno, R. G. Wyatt, C. A. Mebus, R.MLChanock, and A. Z. Kapikian. 1978. Compari-son ofhuman and animal rotavirus strains by gel elec-trophoresis. Virology 87:247-255.
10. Laemmli, U. K. 1970. Cleavage of structuralproteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680-5.
11.Laver,W.G.,and R.G.Webster. 1973. Studies onthe
originof pandemic influenza.IH.Evidenceimplicating duck and equine influenzavimses as possible progeni-tors of the Hong Kong strain of human influenza.
Vi-rology51:383-391.
12.Lewontin,R.C., and J. L Hubby. 1966. A molecular
approach to thestudyofgenetic heterozygosityin
nat-uralpopulations.II. Amount ofvariationand degree of
heterozygosity in natural populations of Drosophila
pseudoobscwura.Genetics64:596-609.
13.Linnemann, C. C.,L.J.Light, T. G.Buchman, J. L
Ballard, and B. Roizman. 1978. Transmission of
herpes-simplexvirus type 1 in a nursery forthe
new-born.Identification of viral isolatesof DNA
"finger-pinting."Lancet i:964-966.
14.McCrae,M.A.,and W.KI Joklil. 1978.The nature of the polypeptide encoded by each of the 10 dsRNA segmentsofreovirustype 3.Virology89:578-593. 15.McNulty,M.S.1978.Rotavirusme. J. Gen.Virol. 40:1-18.
16.Mlkman, R. 1973.Electrophoretic variationin E.coli
fromnatural sources.Science182:1024-1026.
17.Mustoe,T.A.,R. F.Ramig,A H.Sharpe,and B. N.
Fields. 1978. Genetics of reovirus: identification of the daRNA segmentsencoding thepolypeptidesof theit andasizeclasses.Virology89:594-604.
18.Palem,P. 1977. Genes ofinfluenza. Cell 10:1-10.
19.Ramig, R. F.,R. KI Cross, and B. N. Fields. 1977.
Genome RNAs andpolypeptidesofreovirusserotypes 1, 2,and 3. J.Virol. 22:726-733.
20.Rosen, L 1960.Serologicgrouping ofreovirusesby
he-magglutination-inhibition.Am.J.Hyg.71:242-249.
21.Rosen, L 1960. Reoviruses,p. 73-107. In S. Gard, C.
Hallaver,and K. F.Meyer, (ed),Virology monographs.
Springer-Verlag,NewYork.
22.Rosen,L, and F. R. Abinanti.1960.Natural and
exper-imental infectionofcattlewithhumantypes ofreovirus.
Am.J.Hyg.71:424-429.
23.Rosen, L., F. R. nanti, and J. F. Hovin. 1963.
Further observations on thenaturalinfection of cattle
withreovinruses.Am.J.Hyg. 77:3848.
24. Scholtimk, C.,W.Rhode,V. VonHoyningen,and R.Rott. 1978.Ontheoriginof the human influenza virussubtypesH2N2andH3N2.Virology87:13-20. 25. Sharpe,A.H.,R.F.Ramig,T. A.Mustoe,and B. N.
Fields.1978.Ageneticmapofreovirus.L. Correlation
of genome RNA's between serotypes1,2and3.Virology
84:63-74.
26.Weiner,H.L,and B.N. Fields.1977.Neutralization of reovirus: the gene responsible for the neutmlization
antigen.J.Exp.Med. 146:1305-1310.
27. Weinmer,H.L.,R. F.Ramig,T. A.Mustoe,and B. N.
Fields.1978.Identification of the genecodingfor the
hemagglutinin ofreovirus.Virology86:581-584.
28. Wood,H. A. 1973.Viruses withdouble-stranded RNA
genomes. J. Gen.Virol. 20:61-85.
on November 10, 2019 by guest
http://jvi.asm.org/