0022-538X/81/010431-14$02.00/0
Structures
of Two Spliced
Herpes
Simplex
Virus Type 1
Immediate-Early mRNA's Which
Map at the Junctions of the
Unique and Reiterated Regions
of the Virus DNA S
Component
ROGER J. WATSON,l* MARJORIE SULLIVAN,2 ANDGEORGE F. VANDE WOUDE' Laboratoryof Molecular Virology, National CancerInstitute,'andLaboratory of Molecular Genetics, National Instituteof Child Health and HumanDevelopment,2National Institutes of Health, Bethesda,
Maryland 20205
Wehaveexaminedthe structures of two herpes simplex virus type 1
immediate-early (IE) RNAs (IE mRNA-4 andIE mRNA-5) which map at the junctions of
the unique (Us) and reiterated regions
(TRs/IRs)
of the virus DNA shortcomponent.Hybrids between IE cytoplasmicRNA andherpes simplex virus type
1DNArestriction fragmentsweredigestedwithsingle-strand-specific nucleases
S1 and exonuclease VII, and the productswere analyzedby agarose gel
electro-phoresis. Data obtained with the nuclease digestiontechnique were confirmed by
electron microscopy of R-loopstructures formedwith polyadenylated IE RNA
and virus DNAfragments. Itwasfound thatboth IE mRNA-4 and IE mRNA-5
contained a260-base 5'-terminal cotranscript which mapped at equivalent loci
within
TRs/IRs.
These 5'-terminal sequences were shown to be spliced to3'-terminal cotranscripts of1,450 bases (forIE mRNA-4) and 1,540 bases (for IE
mRNA-5). The 3'-terminal cotranscripts containedsequences encoded by both
TRs/IRs
andopposite ends ofUs,
indicating that the intronscontained by the IEmRNA-4 and IE mRNA-5genes, found to be approximately 150 base pairs in
size,mapped
entirely
within the reiteratedsequences. Thedatasuggest that thesegenes maycontaincommonanduniquecomponents, and theimplicationsof this
modelarediscussed.
The
transcriptional
programofherpessimplexvirus type 1
(HSV-1)
is divided into at leastthree
phases,
namely,
theimmediate-early (IE),
early,
and latestages(9,
11,15,23).
TheIE stageof
transcription
istransient,
and theaccumula-tion of IE
RNAs,
whichmap withinrestrictedregions of the virus genome
(9, 14),
isreadily
apparent
only
in cellsinfected with virus in thepresence of
protein synthesis
inhibitors or incells infected with certain HSV-1
temperature-sensitivemutants atthe
nonpermissive
temper-ature
(26).
Synthesis
ofearly
and lateRNAs,
whichmapatloci
throughout
the virusgenome(9,15),is
continuously dependent
uponthe pres-enceofafunctional
IEproduct (27).
Theswitch fromearly
tolatetranscription
isdependent
alsoupon the onsetofvirus DNA
replication
(2, 9,
11, 15).
The structure of the linear double-stranded DNAgenome ofHSV-1isremarkablein that it
consists oftwo
unique
DNAregions,
UL
(115
kilobase
pairs)
andUs (14
kilobasepairs),
eachflanked
by
different invertedrepeated regions,
TRL/IRL
(9
kilobasepairs)
andTRs/IRs
(6
kil-obase
pairs),
respectively
(20).
Inaddition,
thereis ashort direct repetition (the "a" sequence)
present ateachterminus of the DNA,aswellas one or more
copies
ofthissequence atthe IRL/IRS
joint (25). A consequence of this unusualstructure is that the unique DNA regions, UL
and
Us,
are found inverted relative to onean-other, thus
giving
risetofourgenomearrange-mentswhichareusuallypresentin DNA
prep-arationsin
equimolar
amounts(7, 12).Afurtherconsequenceofthisunusualgenome structure is
thatgenes which map intheinverted reiterated
sequences are present in two molar amounts.
Thisisof
particular
relevancetothegenes which areexpressed
atthe IE stageofvirusreplication,
since many of these map, at least
partially,
within these reiterated
regions
(28). Theapprox-imate
genome locations of the fivepredominantIE mRNA species, designated IE
mRNA-1
through-5 (8), have been described
previously
(8, 28). IE mRNA-1 and IE mRNA-3, which have sizesof3.0kilobases
(kb)
and4.7kb,
mapwholly
within thereiteratedsequencesTRL/IRL
and
TRs/IRs, respectively,
whereas the 2-kbspecieIE mRNA-2 maps in
UL.
Two additional2-kb
species,
IEmRNA-4andIEmRNA-5,
were 431on November 10, 2019 by guest
http://jvi.asm.org/
foundtomap at,or closeto,thejunctionsofUs
andTRs/IRs. From these data,and from
anal-ysesof the orientation of these IE mRNA's (8), it was suggested that IE mRNA-4 and IE mRNA-5 containcommon 5' terminimappingin
TRA/IRs
andunique3' terminimapping at op-posite ends ofUs (28).Wereportthe results of experiments designed to fine-structure map IE mRNA-4 and IE mRNA-5, using the nuclease digestion technique of Berk and Sharp (3) and R-loop analysis (24). We show that these twomRNA's contain a 5'-terminal sequence,
map-ping inTRs/IRs, spliced to non-colinear tran-scripts(cotranscripts)containingboth reiterated anduniquesequences.
MATERIALS AND METHODS Cells andviruses. Vero cellswerepropagated as monolayers in 150-cm2 flasks, usingRPMI 1640 me-diumsupplemented with10%calfserum.HSV-1 strain Pattonstocksweremade byinfectingVero cells at low multiplicities of infection.
DNAfragments. The HSV-1 DNA fragmentsused wereclonedincoliphageXgtWES.Band subcloned in plasmid pBR322 (10; K.Denniston-Thompson,L. W. Enquist, and G.F.VandeWoude,submitted for pub-lication). Deletions of the EcoRI Hfragment, to gen-erateNYS-8 andSRS-4,wereselectedfrom XgtWES-EcoRI-Hclones DEC24 and DEC36(10), as described byK.Umene andL.Enquist (manuscript in prepara-tion).
Restriction endonuclease digestion. The con-ditions used for cleavage of DNA with restriction endonucleases have beenreported previously (Dennis-ton-Thompsonetal., submittedfor publication).
LabelingHSV-1 DNAfragments invitro. DNA fragments were labeled at the 5' termini, using
[y-32P]ATP(-2,000 Ci/mmol)andpolynucleotide kinase
(18). BamHI DNA fragments (-1
Ag)
were labeled at the3'terminiin areactionvolumeof 100pl
containing 60mMTris-hydrochloride (pH7.5), 6 mMMgCl2,100 yCi of[a-32P]GTP (>300 Ci/mmol), and 5 U of the Klenowfragment of Escherichia coli DNA polymer-aseI(Boehringer MannheimCorp.) by incubating for 30min at15°C. Theseprocedures enabled theSaIB5 andB4R1probes tobelabeledtospecific activities of105to 106
cpm/,g.
32P-labelingof DNA fragments by nick translation with DNA polymerase I has been described previously (26).Labeling HSV-1DNAfragmentsinNivo. Plas-midscontainingHSV-1 DNA inserts were32plabeled by growing5ml of E. coli cultures to an optical density at 550nmof 0.7 in a definedmediumcontaining200
,uM K2HPO4 and200,uCi ofcarrier-free 32p;per ml. Chloramphenicol wasthenadded to 100,g/ml, and thecultureswereincubatedovernightat370C. Cells
werelysed by incubationfor 5 min onice in a volume of 1.5ml containing 50 mM Tris-hydrochloride (pH 8.0),60mMEDTA,15% sucrose and 1 mg of lysozyme per ml. Anequalvolumeof10% Triton X-100-50 mM Tris-hydrochloride (pH 8.0)-60 mM EDTA was then added,and cellDNA was pelletedbycentrifugation
for 1hat30,000rpminaBeckmantype 40 rotor. The supernatantwasextracted twice withanequal volume ofphenol-chloroform (1:1) andoncewith chloroform. An equal volume of 0.3 M NaOAc was added, and nucleic acidswereprecipitated by the addition of 2.5 volumes of ethanol and storage at -20°C. Nucleic acidswererecoveredbylow-speedcentrifugation and thendigested with EcoRI. Nucleic acidswereresolved byelectrophoresison anagarosegel, and the linearized plasmid DNA bandswerevisualizedby ethidium bro-midestaining. Theappropriate DNAfragmentswere
electroeluted from the gel and thendigested with a second restrictionendonuclease, and the required frag-ments wereisolatedby gelelectrophoresisasbefore.
Virus infection and RNA extraction.Cell motio-layers, preincubated for 30 min incomplete medium containing200 jagofcycloheximideper ml, were in-fected with HSV-1atamultiplicityof infection of 25 and maintained in this medium forafurther 6 hat
37°C.CytoplasmicRNAwasthenprepared by lysisof the cells with Nonidet P-40, removal of nuclei by centrifugation, and phenol-chloroform extraction of thesupernatant fraction (28). RNAwasstored in a
small volume of distilledwaterat-70°C.Where ap-propriate, polyadenylated RNA was selectedon an
oligodeoxythymidylate-cellulose column,asdescribed previously (28).
RNA/DNA hybridization and nuclease treat-ment. To perform a typical hybridization, approxi-mately0.1,ugof theappropriateDNAfragmentwas
ethanol precipitated with 10,ug of cytoplasmic IE RNA.The nucleic acidswerepelletedinaBeckman microfuge, and the pellet was washed once with ethanol. The pelletwasdried and then dissolved in20
pl
of 90% formamide-400mMNaCl-1 mM EDTA-40 mM PIPES [piperazine-N,N'-bis(2-ethanesulfonic acid)],pH6.4.Afteroverlayingwithparaffinoil, the microfuge tubescontainingthehybridizationmixwereincubated for 10min at 70°C. The tubeswere then transferredto a waterbath heldat56°Cand incubated overnight. Afterincubation,thesampleswerediluted with10volumes of either S1 buffer(250 mMNaCl, 1 mM ZnSO4, 5% glycerol, 30mM NaOAc, pH 4.6) or
exonuclease VII buffer(100mMNaCl,8mMEDTA,
10 mM/3-mercaptoethanol, 10 mM
Tris-hydrochlo-ride, pH7.9). S1digestionswereperformed by adding 2,000U of theenzyme(Boehringer Mannheim Corp.) andincubatingat45°Cfor30min.Exonuclease VII digestionswereperformedbyaddinganappropriate amount of the enzyme, purified from E. coli strain M0676 bythe method of Chase and Richardson (6), andincubatingat45°Cfor 1 h.Digestionswere ter-minatedbyadding 10
,tg
ofyeasttRNA and 2.5 vol-umesof coldethanol.Gelelectrophoresis and autoradiography.The products ofS1 andexonuclease VIIdigestions were collected by centrifugationand analyzed by agarose gelelectrophoresis. Electrophoresisundernative con-ditions wasperformedon1.4% agarosegels cast in 20 mM NaOAc-2 mM EDTA-40 mM Tris-hydrochlo-ride, pH7.8.Electrophoresis underdenaturing condi-tionswasperformed on 1.4% agarose gels cast in 30 mM NaOH-2 mM EDTA. Electrophoresis buffers werethesame asthoseinwhichthegelswere cast. CertainS1-digested sampleswereanalyzedona
on November 10, 2019 by guest
http://jvi.asm.org/
dimensional gel system, as described by Kamen et al. (16).Afterelectrophoresis, gels were either dried un-dervacuum andsubjectedto autoradiography (9) or prepared for DNA transfer to nitrocellulose (22) or diazobenzyloxymethyl-cellulose paper (1) obtained fromSchleicher & Schuell Co.
R-loop analysis. As initial steps, the EcoRI H fragment was excised from hybrid phage, and the plasmid pKL43 was linearized by digestion with re-striction endonuclease EcoRI. Approximately 0.1-pg
amountsof EcoRI-cut DNAwerelyophilized with 0.1 pg ofpolyadenylated cytoplasmic IE RNA. Nucleic acidswereredissolved in4plofR-loop hybridization buffer (75% formamide, 100 mM Tricine [Sigma]-NaOH, pH 8.0, 500 mMNaCl, 10mMEDTA) and incubatedovernightat56°C in sealed glass capillaries. Afterincubation,thesampleswerediluted and spread on ahypophase asdescribedpreviously(19). Visual-ization ofmolecules,measurementof their length, and normalization ofR-loop-containing moleculestogive fractional lengths were also performed as reported previously (19). Generally, it was found that greater than80% of the appropriate DNA fragments contained R-loops.
RESULTS
Mapping IE mRNA-5 bynuclease
diges-tion and alkaline agarose gel
electropho-resis. Theuseofsingle-strand-specificnucleases
S1 and exonuclease VII tomap the regions of
DNAprobes hybridized,under highstringency
conditions (5), to complementary RNA
se-quenceshas beendescribedby BerkandSharp
Map Units
0
0.1,,
IE mRNAs
I
It
(3, 4). The basic principle of this procedure is that nuclease Si digests all of the
single-strandedregions oftheDNAprobe unprotected
by duplex formation with RNA, whereas exo-nuclease VIIdigests single-stranded DNA only
ateither terminus ofaduplexstructure. Hence,
a single-strandedDNAloop boundedoneither
sidebyRNA/DNAduplexes,whichisformedat
the splice point of RNA, serves as a substrate
for nuclease Si but isnotdigested by exonucle-aseVII.
Comparison,
then, of the sizes of RNA/DNAhybridsprotected againstthesenucleases
enables thepresence ofsplicedtranscripts to be
recognized.
Initially,weusedamodification oftheabove
technique toanalyze the colinearand
non-coli-nearregions ofIEmRNA-5. In thismodification,
the DNA probes whichare hybridizedto
unla-beled RNA are labeled at a unique 3' or 5'
terminus
generated
by
restriction endonucleasedigestion.Consequent requirementsof this
tech-iiique
arethat theuniquely labeledDNAstrandis of the opposite polarity to the RNA to be
analyzed and that the labeled terminus of this
DNAprobemaps withinsequencesrepresented
by this RNA.
Previousanalyses ofIEmRNA-5 (8, 28)
indi-cated thatthe 3' terminus of this
transcript
mapsin the BamHI 6-BamHI 5 DNA
fragment
and the5'terminusmapsinBamHI4-BamHI3(Fig. 1). The probes selected foruse in thisanalysis
0.8
0,9
1 3
*1
2
1.0
5
3
4
TRL
U
LIR,
IRS
r
Ii/
[
EctR
Eco
RIFIG. 1. Approximate physicalmaplocations anddirectionsofsynthesis ofthefive predominantIEmRNA's, IE mRNA-1 through-5(8,28).Also shown arethe restriction mapsofthe Sregion ofHSV-1 DNAforthe
enzymesEcoRI and BamHI. TheBamHI sitesaredesignated BlthroughB8. ThemaplocationoftheEcoRI Hfragmentis indicated.
us
TRS
B5 4
B7E3oRB,B1
EcoRl=
I
Bi 3 as
lr,L-I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.496.49.441.396.620.2]were,therefore, theSall/BamHI5(SalB5)
frag-ment 3'terminally labeledatthe BamHI 5site
and the BamHI 4-EcoRI (B4R1) fragment 5'
terminally labeled atthe BamHI 4 site (Fig. 1
and 12).
Togeneratethe aboveprobes,wetook
advan-tageoftwo
plasmid
clones:pkL63,
which carriesthe permuted B1B5 fragment containing the
SaIB5sequences, and
pkL43,
which carries theB4R1 fragment. These DNA fragments were
subcloned inpBR322 fromanHSV-1-defective
DNA EcoRI
fragment
as described previously(Denniston-Thompsonet
al.,
submitted forpub-lication).DNAsofpkL63 andpkL43wereeach
cleaved withBamHI and then labeled
respec-tively
at the 3' and 5' termini as described inMaterials and Methods.
Subsequent cleavage
ofpkL63 with Sal I and
pkL43
with EcoRIyielded
the
uniquely
labeled SalB5 and B4R1frag-ments, and these were isolated
by preparative
gelelectrophoresis. These
probes
werethenhy-bridizedtototal
cytoplasmic
IERNA, using high
stringency conditions. After
hybridization,
thesampleswerehalved; one-halfwasdigested with
nuclease S1 and the remainderwasdigested with
exonuclease VII. Theproductsofnuclease
diges-tion were
analyzed by
electrophoresis
on analkalineagarose
gel.
Thegel
wasthen dried and [image:4.496.63.252.378.565.2]subjectedto
autoradiography
(Fig. 2).FIG. 2. CytoplasmicIE RNAwashybridizedtothe BamHI4-EcoRI(B4R1)andSalI-BamHI5(SalB5) DNAfragments 32p labeledatthe5' and3'BamHI termini, respectively. The labeled DNA sequences
protectedagainst nuclease Sl and exonuclease VII
(Exo7) digestion were analyzed by electrophoresis
through analkalineagarosegel and autoradiogra-phy. Tracks marked M contain DNA molecular-weight markers, and the sizes (in bases) of these markersaregiventotheleftandright of the figure.
Hybridization of the 5'-labeled B4R1 DNA
probetocytoplasmic IE RNA resulted in
pro-tection ofapredominant 785-base (b) DNA
frag-mentagainstS1 digestion andtwofragmentsof
1,220and1,600bagainst exonuclease VII
diges-tion (Fig. 2). Hybridization of the 3' terminally
labeled SaIB5 probe to cytoplasmic IE RNA
resultedinprotection ofapredominant band of
570 b against S1 digestion and a predominant
band of590bagainst exonuclease VII digestion
(Fig. 2). Minor bands of approximately 900 and
1,250b werealso detectable in the SaIB5
exo-nucleaseVII-digested sample. Before
unambig-uousinterpretation of these data could be
exer-cised, further control experiments were
per-formedto ensure thatprotection of these
frag-mentsagainst nucleasedigestionwasdependent
upontheaddition of
virus-specific
RNA.5'-labeled B4R1 and3'-labeled SalB5 probes
were
incubated,
asbefore,
with eithercytoplas-mic IE RNAor anequivalentamount ofyeast
tRNA. After nuclease S1 or exonuclease VII
digestion,
the products were analyzed asde-scribed above. The results(Fig. 3) indicated that
protection of the 785-b B4R1 and 570-b SaIB5
fragments against S1 digestionand protection of
the 1220-b B4R1 and 590-b SalB5 fragments
against exonuclease VII digestionwere
depend-ent upon the addition of virus-specific RNA.
However, protection of the 1,600-b B4R1 and
1,250-b SaIB5 fragments against exonuclease
VIIdigestionwasindependent of the addition of
virus-specific
RNA. As thesizes of these latterfragments
were equivalent to the sizes of theB4R1 andSaIB5 probes, failuretobe digested
by
exonuclease VIImayhave been the resultoflimited reassociation or folding ofthese DNA
fragments
atthe termini.The protection of minorSalB5 fragments of
750and850b againstexonuclease VIIdigestion
appeared
tobedependentupon theaddition ofvirus RNA (Fig. 3). If these are the result of
splicing
atthe 3' terminus ofIE mRNA-5,thenthese splices must be present in a very small
proportion of transcripts.
From
datadetailed above, thefollowingcon-clusions were drawn. First, the 5' terminus of
the colinear transcript maps 785 b upstream
fromthe BamHI 4 site, and the 3' terminus of this
colinear
transcript maps 570 b downstreamfrom theBamHI 5site. Second, from the
differ-ence in size between the B4R1 fragment pro-tected against S1 digestion and that protected
against exonuclease VII digestion, the presence
of a non-colinear transcript
spliced
to the 5'terminusof the majorIE
mRNA-5
sequencewasapparent.Third, theclosecorrelation in thesizes
ofthe
SaIB5
fragmentsprotected againstS1
andexonuclease VII digestions implied that there
on November 10, 2019 by guest
http://jvi.asm.org/
SPLICED HSV-1 mRNA's 435
A
on
a
lSw
'X
.--a
iz-- ~ ~ ~ ~ ~ ~"'
FIG. 3. Either(A) the B4RI DNAfragment32plabeled at the BamHI 5' terminus or (B) theSalB5DNA fragment32plabeledattheBamHI3'terminuswasincubated under hybridization conditions with (+) or without(-)theadditionof cytoplasmicIE RNA. WherenocytoplasmicIE RNA waspresent, anequivalent amountofyeast tRNA wasadded. After digestion ofthese samples with nuclease Sl or exonuclease VII (Exo7), theproductswereanalyzed by electrophoresis throughanalkalineagarose gel andautoradiography. Tracks marked M containmolecular-weight markers,the sizes(inbases)of which are indicated.
was nospliceatthe 3' terminus ofIEmRNA-5: thesmalldifference in size observedwas
proba-bly
aresult of theprocessive
activity ofexonu-clease VII(6),
thereby
notdigestingawaysingle-stranded DNA
completely.
Fourth, assumingthat the small
(185-b)
BamHI 5-BamHI4frag-mentsequencesarepresentin the
major
colinearregion
ofIEmRNA-5(an
assumption
latercon-firmed), the total size of this colinear transcript
is 1,540b
(i.e.,
785+570 + 185b).
Analyses
ofRNA/DNA hybrids.
Thema-jor product of
Si digestion
ofaduplex formedbetween a
spliced
RNA molecule and a DNAfragment encoding that
transcript
is an RNA/DNA
hybrid
whose sizecorresponds
tothe totallength of the
spliced cotranscripts,
the RNAstrand of thehybridholding togetherthetwo or morenon-colinear exons. Inpractice,inausually
minorprotection of these RNA/DNA hybrids,
nicking of the RNAat the splice point occurs
(4). Thus,nativegel
electrophoretic
analysis
oftheSl-digested hybridsformed between the
5'-labeled B4R1 fragment and
cytoplasmic
IERNAshould revealtwobandsupon
autoradiog-raphy,thelargercorrespondingtothemajorIE
mRNA-5sequence
(the
3'-terminalcotranscript)plus the 5'-terminal cotranscript and thesmaller
correspondingtothe3'-terminalcotranscript
se-quencealone. Such aresult wasobserved (Fig.
4). Native gel analysis of the S1-digested,
3'-labeled SalB5
DNA/IE
mRNA-5hybrid (Fig. 4)revealed,
as expected, only one predominant band.The sizes ofthese RNA/DNA
hybrid bands,
as measured with duplex DNA size
markers,
weregreater than
predicted
(Fig. 4).
Forexam-ple, the apparent molecular
weights
of theB4R1/IE mRNA-5
hybrids
wereapproximately
900 and 1,150
b,
whereas the smaller ofthesetwobands would be
expected
tobe785b. Thisapparently
aberrantelectrophoretic
mobility
ofRNA/DNA hybridswas
consistently
noted forduplexes of<1
kb,
although
notforlarger
hy-brids. However, the difference in molecular
weightbetween theobserved
B4R1/IE
mRNA-5 hybrids
suggested
that thelength
of the5'-terminal
cotranscript
wasapproximately
250b.Two-dimensional
gel
analysis
ofBamHI4-EcoRI
DNA/IE
mRNA-5hybrids.
Tocon-firm that the two B4R1
DNA/IE
mRNA-5on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.451.81.368.61.327.2]436 SULLIVAN, AND
-S
13S3 s
1076
9-0 so
SWr i
FIG. 4. Cytoplasmic IE RNA was hybridized to
either a 3', 32P-labeled SalB5 or a 5', 32P-labeled B4RJ HSV-1 DNAprobe, and RNA/DNA hybrids protected against nuclease Si digestion were
ana-lyzed by electrophoresis throughanativeagarosegel andautoradiography. Thepositions of duplexDNA size markersareindicatedin the tracklabeled M.
bands observed by neutralagarose gel analysis were generated by duplex formation with a
spliced RNA molecule, these RNA/DNA hy-brids were subjected to two-dimensional gel analysisasdescribedby Kamenetal. (16). The
hybridswere runfirst, under native conditions,
downone side ofasquare agarose gel. The gel wasthen soaked in alkalito destroy the RNA strands oftheduplexes and turnedthrough900, andelectrophoresiswasresumed under alkaline
conditions. Unlabeled HaeIII-digested 4X174 replicative-form DNA fragmentswere
coelectro-phoresed for size markers. After electrophoresis, the DNAfragments were transferred to nitro-cellulose. Autoradiography ofthis blot (Fig. 5) revealedtwospots.The electrophoreticmobility of thesespotsunder native and alkaline condi-tionswasdetermined withrespect tothe 4X174 DNA size markers, which were visualized by
hybridization with 32P-labeled 4X174 DNA(not
shown). Under native conditions,the mobility of thesespotscorrespondedtothe sizes of thetwo
bandsobserved withtheB4R1 probeonnative
gel analysis (Fig. 4). In the second dimension, under alkaline conditions, the electrophoretic mobility of both spotswassimilar,
correspond-ingtoasize of785b. These data indicatedthat
undernative conditions the RNAstrand,
repre-sented by the
larger
spot, heldtogether
non-colinearly
transcribed
DNAfragments.
Alkalitreatment of this
RNA/DNA
hybrid
releasedthese DNA fragments,
only
one of which waslabeled.
Size
of
the IE mRNA-55'-terminal
cotran-script.
Use ofend-labeled DNAprobes
doesnotallow direct size determination of each
non-col-inear
transcript.
Todetermine the size of the5'-terminal
cotranscript
ofIEmRNA-5,
aninvivo32P-labeled
B4R1 DNAfragment
wasusedas aprobe for the
Si analysis.
Included in thisex-periment also were
analyses using
the in vivolabeled
SalB5
fragment
and theunseparated
fragments
produced by cleavage
ofB4R1 withXhoI. Sequences
protected
against
Sidigestion
by
hybridization
tocytoplasmic
IE RNA wereresolvedby alkalineagarosegel electrophoresis.
Autoradiography of thisgel revealed the
pres-ence of thecharacteristic 570-b SalB5 and
785-b B4R1
fragments
(Fig.
6). Protection ofaB4R1sequence of260 b was alsoapparent, andthis thenmayrepresentthe5'-terminal
cotranscript.
Itcouldnotbeexcluded that
protection
of this260-b fragment resulted from
hybridization
ofthe B4R1
probe
to IEmRNA-4,
which waspreviously
reported to share 5'-terminal se-quenceswith IE mRNA-5(28).
It is shown belowthat IE mRNA-4 and IE mRNA-5 contain a
similar
5'-te:rminal
sequence.Cleavage
of B4R1withXhoI before
hybridization
resulted inpro-tection of bands of
approximately
250and450b(Fig. 6), as
predicted
from the restriction mapfor thisenzyme
(Fig. 12).
R-loop
analysis
of IE mRNA-5 andIE
mRNA-4.Tosubstantiate data obtained
by
thenuclease
analyses,
the 5' terminusofIEmRNA-5 was mapped by
using
theR-loop technique
(24). Cytoplasmic polyadenylated IE RNA was
incubated with EcoRI-cutpkL43 (containingthe
B4R1 fragment insert) under conditions
favora-ble for
R-loop
formation. Thepositions
ofR-loops in the DNA molecule were mapped by
electron microscopy: orientation of the
mole-culeswasfacilitatedby the 4.0-kbpBR322 DNA
sequence of EcoRI-cut
pkL43,
in which noR-loopswere observed.
The two typesofR-loopstructures observed areshown inFig. 7.Some moleculescontained
asingle R-loop,witha smallspur near one end
of the loop. Other moleculescontained two
R-loops, with the
small
spurstill
present. Thisindicated that the spur, which then corresponds tothe intron of the IE mRNA-5 gene, issingle
strandedand may thus form a duplexwith the
complementaryDNAsequences of the displaced
strand. Obviously, the duplex thus formed is
relativelyunstable
thernodynamically.
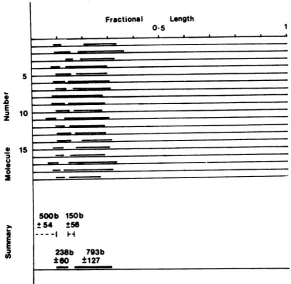
TheR-loops present in 19 representative
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.496.104.195.78.308.2]ecules are shown schematically in Fig. 8. Also
shown is a summary ofthese data, which
indi-cates that the 5'-terminal sequence of IE
mRNA-5 isapproximately500 b from the EcoRI site ofB4R1. This summary also indicates that the sizemeasurementsofthe IE mRNA-5 com-ponents obtained by this analysis are in close agreement with those obtained by the nuclease
digestiontechniques.
To investigate the relationship between the
structureofIE5 and that of IE
mRNA-4, afurther R-loopanalysis wasperformed,this
time using the HSV-1EcoRI Hfragment cloned
inXgt.WES.B. TheEcoRI H fragment contains
thewholeof the Usregion flanked on either side
by
TRs/IRs
sequences (Fig. 12), and thus thecompletetemplates for both IE mRNA-5 andIE
mRNA-4 are presentin this DNA. Asummary
of dataobtained in this experiment is shown in
Fig.9. Themajority of unnicked EcoRIH
frag-ments contained R-loops arranged
symmetri-cally at either end of the molecule. In many
instances,introns were noted atboth ends of the
fragments, andthe positions of these were the
same relativetoeach end. This madeit
impos-sible to orient the DNA molecules, indicating
that the structures of IE mRNA-5 and IE mRNA-4 were very similar. A number of
ran-domly occurring and ranran-domly sized R-loops
were noted in some molecules in addition to
those R-loops notedatthe ends.In no instance
weretheserandom R-loops observedtocontain
splice points.
Sequences of IE 4 and IE
mRNA-1353
w-078
-872 '
0
603
'W310 >
~~~~~~
4~~~~~~~~~~~~~~~A~~~ ~~~ ~~~ ~~~ ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~FIG. 5. Nuclease SI-resistant RNA/DNA hybrids formed between cytoplasmic IE RNA and a 5',
32P-labeled B4RI HSV-1DNAfragmentwereanalyzedbyelectrophoresisintwodimensionsthroughanagarose
gel,followed by autoradiography. Thefirstdimensionofelectrophoresiswasunder nativeconditions. Thegel
wasthen soaked inalkali,turnedthrough90',andrununder alkaline conditions. Thepositions ofunlabeled DNAfragmentsize markersareindicated.
37,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.496.109.397.262.615.2]438 WATSON, SULLIVAN, AND VANDE WOUDE
FIG. 6. CytoplasmicIE RNAwashybridizedtothe in vivo 32P-labeled HSV-1 DNA fragments SalB5, B4R1, and B4RIfurthercleavedwith XhoI.Hybrids
so formed were treated with nuclease Sl, and the productswereanalyzedby alkaline agarose gel elec-trophoresis andautoradiography. Thepositions and sizes(inbases)ofmolecular-weight markersare in-dicated in the track labeled M.
5.Tofurther
investigate
thestructuralrelated-ness of IE mRNA-4 and IE
mRNA-5,
it was necessary to determine which sequences thesetranscripts share in common. The strategy
adopted was togenerate unlabeledSl-digested
RNA/DNA hybrids between cytoplasmic IE
RNAand DNAfragments comprisingopposite ends of the EcoRI Hfragment. These subfrag-ments weregenerated by selectingsemispecific
deletions of the EcoRI H
fragment
cloned inXgt.WES.B,
asdescribedby
Umene andEnquist
(inpreparation), andwere
given
theidentifica-tion codes SRS-4 and NYS-8.
SRS-4, then,
con-tained the entire geneof IE
mRNA-5,
whereasNYS-8 contained that of IE mRNA-4
(see
Fig.12). The
Sl-digested RNA/DNA
hybrids
soformed were separated on native or
alkaline
agarose
gels,
and DNAs were transferred byblotting
to eithernitrocellulose ordiazobenzyl-oxymethyl-cellulose
paper. To these blotsspe-cific nick-translatedDNAfragment probeswere
thenhybridized.
Autoradiographs obtained
by hybridization
ofB4R1 andEcoRI-Hprobestoprotected NYS-8
and SRS-4 DNAsequences,resolvedonanative
agarose gel, are shown in
Fig.
10. Asexpected,
B4R1 DNAhybridized
strongly
to theRNA-protected sequences of
SRS-4;
the two bandsapparent
correspond
to thespliced
5'- and3'-terminal cotranscript sequences (upper band)
and thesequencesof the3'-terminalcotranscript
alone (lower band). This DNA fragmentprobe
hybridized also to the equivalent IE mRNA-4
sequences generated with NYS-8 DNA. This
indicated thatnotonly does the 5'-terminal
co-transcriptsequenceofIEmRNA-4map in
TRs/
IRs,
but so also does part of the 3'-terminalcotranscript (corresponding
tothe lower of thetwo
bands). By
extrapolation,
the intron of theIE mRNA-4gene, and thus of the IE mRNA-5
gene, maps
wholly
within TRs/IRs. From thelow autoradiographic intensity of the lower
NYS-8 band obtained upon
hybridization
withthe B4R1
probe,
we inferthat only limitedse-quences of the IE mRNA-4 3'-terminal
cotran-script map in reiterated DNA: the control
ex-periment,
hybridization
to this blot ofnick-translated EcoRI-H DNA (Fig. 10), indicated
that the DNA sequence mass in each ofthese
hybridswas
approximately
equivalent, thusval-idating this inference.It wasalsonoted that the
3'-terminal cotranscript of IE mRNA-4 is
smaller than that ofIEmRNA-5.
Theresults of hybridizationof nick-translated
B4R1 DNA tothe diazobenzyloxymethyl-cellu-lose blot of these RNA/DNA hybrid species, separated on an
alkaline
agarose gel,confirmed
some of the above observations (Fig. 11). Inaddition, the probe hybridized notonly to the
3'-terminal cotranscript sequences of both IE
mRNA-4 and IE mRNA-5, but also to a 260-b
fragmentobtained with both NYS-8 and SRS-4
FIG. 7. Polyadenylated cytoplasmic IE RNAwashybridized to EcoRI-cut plasmid pkL43 (containing a B4R1 HSV-1 DNAfragmentinsert),andR-loopssoformedwereanalyzed by electron microscopy. Represented abovearethetwotypesof R-loopstructures seen.Spurs, corresponding to the IE mRNA-5 gene intron, are indicatedbyarrows.Double-stranded(ds)regions of the hybridsarerepresented by thethickerline,andthe displaced single-stranded(ss)DNA strands arerepresented by the thinnerline.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.496.79.220.85.422.2]439
on November 10, 2019 by guest
http://jvi.asm.org/
5 .0 E z z
0
.3
U
.5
2a
10
15
h.I
Fractional Length
0-5 1
P-5OOb iSOb
1-~~~~~~~~~~~~~~~
±54 ±56 238b 793b *60 *127
FIG. 8. Diagrammaticrepresentation ofa number ofpKL43moleculescontaining R-loops producedas described in thelegend toFig. 7. The measured length of pkL43DNA has been standardizedfor each molecule. The relativepositions of R-loopsare shown above the continuous linerepresenting pkL43DNA. The summary indicates the mean average size (in bases) ofthe two R-loops observed, made with the assumption that the length of pkL43DNAis 5.7kb. Also indicated are the distancefrom the EcoRI-cut terminusof pkL43DNA to thestartofthefirstR-loopandthe sizeofthe intron. Standarddeviationsare
given foreachmeansize estimate in bases.
.0
a
E
z
U
0 2
5 10
[image:10.496.103.395.75.359.2]Fractional Length 0-5
FIG. 9. Diagrammatic representation of a number of R-loop structures formed between the EcoRI H fragmentandpolyadenylated cytoplasmic IE RNA. The measured lengths of the EcoRI H fragment, which is 15.4kb in size, werestandardized. The relative positions of R-loops are shown above the continuous line representing the DNA fragment. No attempt was made to orient these molecules.
DNA. This sequence, then, corresponds to the 5'-terminal cotranscript. Hybridization to this sequence was not observed with the blot of the nativegel,since DNA fragments of this size did
nottransfertonitrocellulose.Thesizeestimates
of the 3'-terminal cotranscripts obtained with
the denaturing gelwere 1,450 b for IE mRNA
and 1,540 bfor IE mRNA-5. The sizes and map
locationof these mRNA species are summarized
inFig. 12.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.496.73.433.441.571.2]-~~~~~~~.~~
,qt
a"so d
FIG. 10. Cytoplasmic IE RNA washybridized to DNAs of coliphage A (isolates NYS-8 and SRS-4) containingdeletions ofthe HSV-1 EcoRI H fragment. TheHSV-1DNAsequencescarriedin these deleted
phageDNAsareshown inFig. 12.After nuclease S1 digestion, theprotected RNA/DNA duplexes were
coelectrophoresed with
0X174
and pBR322 DNA fragment markers (tracks marked M) on a nativeagarose gel. The gelwas then treated with alkali,
andDNAfragmentsweretransferredbyblottingto
anitrocellulosemembrane.Amixtureof32P-labeled nick-translatedpkL43 DNA (containing the B4R1 insert)and
OX174
DNA washybridizedtothe blot, andhybridizationwas visualizedbyautoradiogra-phy. Subsequently, nick-translated EcoRI (Ri H) DNAwashybridizedtotheblot.The sizes(in bases) of DNA marker bandsareindicated.
DISCUSSION
Wehavedescribedthe structures of two HSV-1 IE mRNA's and have provided the first evi-dence that these virus transcripts are spliced.
These datawereobtained with three
independ-ently cloned HSV-1 DNA fragmentsas probes
inthe hybridization reactions, thus miIniizing thepossibilitythat theputativeintrons observed reflectedspecific additionsorrearrangementsof the virus DNA fragments on cloning. Specific
deletions ofthe clonedDNAfragmentsarenot consistent with the experimental data, for
ex-ample, the observation of introns by electron microscopy of R-loopstructures.
The datawe obtained indicate that both IE
mRNA-4 and IE mRNA-5 contain a 260-b
5'-terminal cotranscript. Thesecotranscripts map
atequivalent locationswithin
TRs/IRs,
suggest-ing that the 5'-terminal sequencesof IE
mRNA-4 and IE mRNA-5 are
similar.
More sequence data are required to establish whether thesecotranscripts are identical or, indeed, whether
microheterogeneityexists within the5'-terminal
sequence of each IE mRNA. The 260-b se-quences werefoundtobesplicedto
3'-terminal
cotranscripts of 1,450 b (for IE mRNA-4) and
1,540 b (for IE mRNA-5). These 3'-terminal
cotranscripts share some, butlimited, sequence
homology, indicating that they are encoded in
partby
TRs/IRs
and in partbyopposite endsofUs. It
follows
that the introns of the IEmRNA-4andIEmRNA-5geneslieentirely within
TRs/
IRs.
The size of theseintrons wasdeterminedbyR-loopanalysis to be 150 ± 56 b. Thenuclease
analysesindicated that the intron size was 175
b, that is, the size of the B4R1 fragment
pro-tected againstexonuclease VII digestion (1,220
-0* __724
_* _ -
~~4~s
ts ~ ~ l' *m
FIG. 11. Cytoplasmic IE RNA was hybridized to
the HSV-1 DNAEcoRI Hfragmentortothe DNAs
of coliphageA containingdeletions oftheEcoRI H
fragment (isolatesNYS-8andSRS-4). Afternuclease Si digestion,theprotectedDNA sequenceswere
coe-lectrophoresed withpBR322 and
OX1
74DNAfrag-mentsizemarkers(tracksmarkedM)on analkaline agarosegel.DNAfragmentswerethentransferred by blotting to diazobenzyloxymethyl-cellulosepaper. A mixture of 32P-labeled nick-translatedpkL43 DNA
(carryingtheB4RJDNAinsert)and
OX1
74DNAwashybridized to the blot, and hybridization was visu-alized by autoradiography. The sizes (in bases) of DNAfragmentsizemarkersareindicated.
on November 10, 2019 by guest
http://jvi.asm.org/
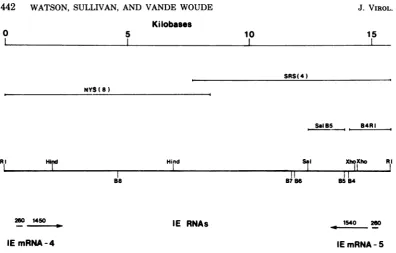
[image:11.496.83.221.73.326.2] [image:11.496.258.450.296.533.2]Kilobases
0 5
SRS(4) NYS(a)
B
Sol S B4R I
RI Hir Hind Sol XhoXho RI
I
~~~III I I88
26 1450
_ - IE RNAs
IEmRNA-4
FIG. 12. Summary ofthemaplocationsofIEmRNA-4 and IE mRNA-5. Apartialrestrictionmapofthe EcoRI Hfragmentisshown, indicatingthecleavagesitesofBamHI(B4-B8), HindIII (Hind),andrelevant SalI(Sal)and XhoI(Xho)sites. Also indicatedarethemaplociof fragmentsused inanalyses reported here, SalI-BamHI 5(SalB5)and BamHI 4-EcoRI(B4R1),and the HSV-1 DNAsequencescontained in deletedA
isolatesNYS-8 and SRS-4. The sizes(in bases) ofthecotranscripts ofIEmRNA-4and IEmRNA-5aregiven,
and their directionsof synthesisareindicatedbyarrows.
b)minus the total of the two B4R1cotranscripts protected against Si digestion (785+260b). As theR-loop analysis indicatedthat the 5' termini of bothIE mRNA-4 andIE mRNA-5 were lo-cated 500 bfromthe EcoRI site,thereiterated DNA sequences of the S region must extend further than 910 b from that restriction site.
Nuclease analyses indicate that the sizes of the transcribed regions ofIE mRNA-4 and IE mRNA-5 are 1,710 and 1,800 b, respectively.
Whenconsiderationistaken ofaddition ofa
3'-terminalpolyadenylate tail of 150to200b(21), it is apparent that these size estimates are in
close agreement tothesize estimates of 2.0 kb for each ofthesemRNA'sdetermined by methyl
mercury agarosegelelectrophoresis (28).
No evidence was found for the presence of
unspliced transcripts partially colinear with IE mRNA-4 and IE mRNA-5, either in polyaden-ylatedorintotalcytoplasmic IE RNA samples.
Presumably,then, the splicingeventswhich
gen-erate these mRNA'stake place in the cell
nu-cleus, and unspliced transcripts are not
trans-portedto,orare veryrapidlyturnedoverin,the
cytoplasm. Furthermore, alternative splicing patterns which generate anumber of mRNA's
from asingle transcriptional unit, as has been
observedwith theearly transcripts of adenovirus 2(4) and simian virus40 (3),werenotobserved
with these HSV-1 IE mRNA's. Consistent with these observations, use of the Northem blot
technique (1) to analyze wild-type HSV-1 IE transcripts hasnotrevealedthepresenceof ad-ditional transcripts partially colinear with IE
mRNA-4and IEmRNA-5 (R. J. Watsonetal., manuscript in preparation). Thepossibility that these additionalcytoplasmic IE transcripts are
presentinveryminoramounts cannot,however, beentirelydisregarded.
From thesequencescontainedbyIE
mRNA-4 and IE mRNA-5, it is implicit that both S
reiterated regions are transcribed, at least in
part. As has beensuggested previously (8, 28), thesemRNA'smaybe transcribedbyusing lo-cationally distinct promoters of identical se-quencemappinginTRsandIRs.DNAsequence
datato identify possible promoter regions and analyses of possible nuclear mRNA precursor
moleculesarerequiredtosubstantiate this
pro-posal. The observation, on R-loop analysis, of
randomtranscripts mappinginUs (Fig. 9) may
indicatethat theIE mRNA'sdescribed indetail abovearegenerated fromlargeprecursors
con-tainingtheseadditional RNAsequences.
Alter-natively, these random RNA speciesmayhave
been transcribed byusingpromoters normally recognizedatearlyorlatetimespostinfection. It
mustbeemphasized that these additional
tran-10 15
I1
87 6 B584II
1540 260
IE mRNA-5
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.496.56.453.55.308.2]scriptswererandomas tosize andlocation,were
found in minoramounts, andwerepresentina
cytoplasmic fraction selected by binding to an
oligodeoxythymidylate-cellulose column.
IE mRNA-4 and IE mRNA-5 specify virus
polypeptides of apparent molecular weights
68,000and 12,000 (28), and these proteins have
been designated Vmw 68 and Vmw 12,
respec-tively. In vitro translation of IE mRNA-4,
se-lected by hybridization toHindIII-n, indicated
that this mRNA
specified
Vmw 68 (2). Thetranscribed region of IE mRNA-4 was shown
heretobe 1,710b, sufficient tocode fora
poly-peptide ofa little more than60,000 molecular
weight. Although phosphorylation of Vmw 68
(17) mayhave ledto an overestimation of the
molecular weight,theclosecorrelation between
thecoding capacity ofIEmRNA-4 and the size
of itsputativeprotein productsuggeststhat the
transcript doesnotcontainextensive noncoding
regions. It is possible, then, that the 260-b
5'-terminal cotranscript contains codingsequences
and isnotmerelyaleader RNA. If this
cotran-script
istranslated,
then itmaybe anticipated thatthepolypeptides specified
by IE mRNA-4and IE mRNA-5, Vmw 68 and Vmw 12, share
commonN-terminalpeptides and that the
ter-minationsignals for those proteinsare encoded
by-'opposite
ends ofUs.
Whereas further dataarerequiredtoprovethishypothesis, it remains
aninterestingwayin whichtogenerate
polypep-tidescontaining bothcommonandunique
pep-tides.
ACKNOWLEDGMENTS
We thankKatherine Denniston-Thompson and Lynn W. Enquist forcritical evaluation of this manuscript and for helpful suggestionsduringthecourseof this work.Also,we thank Kenichi Umeneforprovidinguswith SRS-4 and NYS-8DNA.
LITERATURE CiTED
1.Alwine, J. C., D. J.Kemp, andG. R. Stark. 1977. Method for detection of specific RNAsinagarosegels bytransfertodiazobenzyloxymethyl-paperand
hybrid-izationwithDNAprobes. Proc. Natl. Acad. Sci.U.S.A. 74:5350-5354.
2. Anderson, K. P., R. H.Costa, L. E. Holland, and E. K.Wagner.1980.Characterizationofherpessimplex virus type 1RNApresentinthe absence of de novo proteinsynthesis.J.Virol. 34:9-27.
3. Berk,A.J., andP.A.Sharp.1978.SplicedearlymRNAs of simian virus 40. Proc. Natl.Acad. Sci. U.S.A.75: 1274-1278.
4. Berk, A. J., and P. A.Sharp. 1978.Structure of the adenovirus2earlymRNAs.Cell14:695-711. 5. Casey, J.,andN.Davidson.1977. Ratesof formation
and thermalstabilities of RNA:DNA and DNA:DNA duplexesathighconcentrationsofformamide.Nucleic AcidsRes. 4:1539-1552.
6. Chase,J.W.,and C. C. Richardson.1974.Exonuclease VII ofEscherichiacoli:purification andproperties.J. Biol. Chem. 249:4545-4552.
7. Clements,J.B., R.Cortini,andN. M. Wilkie. 1976. Analysis of herpesvirus DNA substructure by means of restriction endonucleases. J. Gen. Virol. 30:243-256. 8. Clements, J. B., J. McLauchlan, and D. J. McGeoch.
1979.Orientation of herpes simplex virus type 1 imme-diateearly mRNAs. Nucleic Acids Res. 7:77-91. 9. Clements,J.B., R. J. Watson, and N. M. Wilkie. 1977.
Temporal regulation of herpes simplex virus type 1 transcription: location oftranscripts on the viral ge-nome.Cell12:275-285.
10.Enquist, L. W., M. J. Madden, P. Schiop-Stansly, and G.F. Vande Woude. 1979. Cloning of herpes simplex type1DNA fragments inabacteriophage lambda vec-tor.Science203:541-544.
11. Frenkel, R., and B.Roiznan. 1972. Ribonucleic acid synthesis in cells infected with herpes simplex virus: controls of transcription and of RNA abundance. Proc. Natl.Acad. Sci. U.S.A.69:2654-2658.
12. Hayward,G.S.,R.J.Jacob,S.C.Wadsworth, and B. Roizman. 1975. Anatomy of herpes simplex virus DNA: evidence for four populations of molecules that differ in the relative orientations of their long and short components. Proc. Natl. Acad. Sci. U.S.A. 72:4243-4247.
13. Holland,L.E.,K. P.Anderson,C.Shipman,Jr., and E. K.Wagner. 1980. Viral DNA synthesis is required for the efficient expression of specific herpessimplex virus type 1mRNA species. Virology 101:10-24. 14. Jones,P.C.,G. S. Hayward, andB.Roizman. 1977.
AnatomyofherpessimplexvirusDNA.VII.aRNA is homologous to noncontiguous sites in both the L and S componentsofviral DNA. J. Virol. 21:268-276. 15. Jones, P. C., and B. Roizman. 1979. Regulation of
herpesvirus macromolecularsynthesis.VIII.The tran-scriptionprogramconsists of threephase duringwhich bothextentoftranscriptionand accumulation of RNA in thecytoplasm are regulated. J. Virol. 31:299-314. 16. Kamen,R.,J.Favaloro,and J. Parker. 1980.
Topog-raphy of the three late mRNAs ofpolyomavirus which encode the virionproteins.J.Virol.33:637-642. 17. Marsden,H. S., N. D. Stow,V. G. Preston, M. C.
Timbury,and N. M. Wilkie. 1978.Physical mapping ofherpessimplexvirus-inducedpolypeptides.J. Virol. 28:624-642.
18. Maxam,A.M.,and W.Gilbert.1977. A newmethod for sequencingDNA. Proc.Natl. Acad. Sci.U.S.A. 74:560-564.
19. Meissner,H.C.C.,J.Meyer,J. V.Maizel, Jr.,and H. Westphal. 1977. Visualization and mapping of late nuclear adenovirus RNA. Cell10:225-235.
20. Sheldrick, P.,and N. Berthelot.1974.Inverted
repeti-tions in the chromosome ofherpessimplexvirus. Cold SpringHarborSymp. Quant. Biol.39:667-678. 21. Silverstein, S.,R.Millette,P.Jones,and B. Roizman.
1976.RNAsynthesisincells infected withherpes sim-plexvirus.XII.Sequencecomplexityandpropertiesof RNA differinginextentofadenylation.J. Virol. 18: 977-991.
22. Southern,E. M. 1975. Detection ofspecific sequences amongDNAfragmentsseparatedbygel
electrophore-sis. J.Mol.Biol.98:503-518.
23.Swanstrom,R.I.,and E. K.Wagner.1974.Regulation
ofsynthesis of herpes simplex type 1 virus mRNA duringproductiveinfection.Virology60:522-533. 24. Thomas, M., R. L. White, and R. W. Davis. 1976.
Hybridization of RNAtodouble-stranded DNA: for-mation ofR-loops. Proc. Natl. Acad. Sci. U.S.A. 73: 2294-2298.
25. Wagner,M.J.,andW.C. Summers.1978.Structure of thejointregionandthetermini ofthe DNAofherpes simplexvirustype1.J. Virol.27:374-387.
26. Watson,R.J., andJ. B. Clements. 1978. Characteri-zationoftranscription-deficienttemperature-sensitive
on November 10, 2019 by guest
http://jvi.asm.org/
mutantsofherpes simplex virus type 1.Virology 91: 364-379.
27. Watson,R.J.,and J. B. Clements. 1980. Identification ofaherpes simplexvirus type 1 functioncontinuously required forsynthesis ofearlyand late virus RNAs.
Nature (London) 285:329-330.
28. Watson, R. J., C. M. Preston, and J. B. Clements. 1979.Separation andcharacterization of herpes simplex virustype1immediate-earlymRNA's. J. Virol. 31:42-52.
J. VIROL.