Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Spatial and Temporal Organization of Adeno-Associated Virus DNA
Replication in Live Cells
Cornel Fraefel,
1* Anne Greet Bittermann,
2Hansruedi Bu¨eler,
3Irma Heid,
1Thomas Ba¨chi,
2and Mathias Ackermann
1Institute of Virology,1Center for Microscopy,2and Institute of Molecular Biology,3University of Zurich,
Zurich, Switzerland
Received 30 July 2003/Accepted 16 September 2003
Upon cell entry, the genomes of herpes simplex virus type 1 (HSV-1) and adenovirus (Ad) associate with distinct nuclear structures termed ND10 or promyelocytic leukemia (PML) nuclear bodies (NBs). PML NB morphology is altered or disrupted by specific viral proteins as replication proceeds. We examined whether adeno-associated virus (AAV) replication compartments also associate with PML NBs, and whether modifi-cation or disruption of these by HSV-1 or Ad, both of which are helper viruses for AAV, is necessary at all. Furthermore, to add a fourth dimension to our present view of AAV replication, we established an assay that
allows visualization of AAV replication in live cells. A recombinant AAV containing 40lacrepressor binding
sites between the AAV inverted terminal repeats was constructed. AAV Rep protein and helper virus-mediated replication of this recombinant AAV genome was visualized by binding of enhanced yellow fluorescent
protein-lacrepressor fusion protein to double-stranded AAV replication intermediates. We demonstrate in live cells
that AAV DNA replication occurs in compartments which colocalize with AAV Rep. Early after infection, the replication compartments were small and varied in numbers from 2 to more than 40 per cell nucleus. Within 4 to 8 h, individual small replication compartments expanded and fused to larger structures which filled out much of the cell nucleus. We also show that AAV replication compartments can associate with modified PML NBs in Ad-infected cells. In wild-type HSV-1-infected cells, AAV replication compartments and PML NBs did not coexist, presumably because PML was completely disrupted by the HSV-1 ICP0 protein. However, alter-ation or disruption of PML appears not to be a prerequisite for AAV replicalter-ation, as the formalter-ation of replicalter-ation compartments was normal when the ICP0 mutants HSV-1 dl1403 and HSV-1 FXE, which do not affect PML NBs, were used as the helper viruses; under these conditions, AAV replication compartments did not associate with PML NBs.
Adeno-associated virus 2 (AAV) is a nonpathogenic human parvovirus with a genome of linear, single-stranded DNA (2). The DNA is 4.7 kb long and includes 145-base inverted termi-nal repeats (ITRs) at both ends flanking two clusters of genes,
rep and cap (25, 41). The rep genes encode four proteins, Rep78, 68, 52, and 40, from two different promoters, p5 and p19. Thecapgene is transcribed from the p40 promoter and encodes three proteins that form the icosahedral virus capsid. AAV can enter both productive or latent infections, depending on the presence or absence of a helper virus, such as adeno-virus (Ad) or herpes simplex adeno-virus type 1 (HSV-1). In the presence of helper virus, ITRs and either Rep78 or Rep68 are sufficient for replication of the AAV genome. In particular, Rep78 and Rep68 bind to a specific sequence within the ITRs, the Rep binding site (RBS) (26, 34), and cleave in a site- and strand-specific manner at the terminal resolution site located 13 nucleotides upstream of the RBS (3, 17, 39). The RBS and terminal resolution site act as a minimum origin of Rep-me-diated DNA replication (48, 49). In the absence of helper virus, ITRs and either Rep78 or Rep68 are also sufficient to mediate the integration of the AAV genome into the host cell genome,
preferentially into a site termed AAVS1 on chromosome 19 of human cells (19, 20, 22, 44).
Many of its biological properties make AAV an attractive platform for the development of gene therapy vectors, recom-binant AAV (rAAV) vectors (27, 37), or HSV-1–AAV (12, 46) and Ad–AAV hybrid vectors (31). rAAV vectors are derived from bacterial plasmids that contain the AAV ITRs flanking a transgene cassette. For rescue/replication of the ITR-flanked transgene cassette from the plasmid backbone (36) and pack-aging of single-stranded replication products into AAV virions, the AAV replicative and structural genes (rep and cap) are provided intransfrom a separate plasmid, and the helper virus functions are provided from either Ad or HSV-1 (27).
AAV DNA synthesis originates from the palindromic ITR sequence which forms a hairpin and acts as both the origin of DNA replication and the primer (2). A replicative intermedi-ate is formed as a linear duplex molecule covalently linked at one end by the hairpin primer. If the hairpin structure created by the initial priming is not resolved, continued synthesis will lead to double-stranded, dimeric intermediates of the AAV DNA (15). If the covalent link is resolved by the Rep proteins, the hairpin is transferred to the progeny strand and the result-ing 3⬘-terminal gap in the parental strand is repaired by using the transferred sequence as a template (47). Mature single-stranded DNA genomes are removed from the replicative complex and packaged into empty capsids.
* Corresponding author. Mailing address: Institute of Virology, Uni-versity of Zurich, Winterthurerstrasse 266a, CH-8057 Zurich, Switzer-land. Phone: 41 1 635 8713. Fax: 41 1 635 8911. E-mail: cornelf@vetvir .unizh.ch.
389
on November 8, 2019 by guest
http://jvi.asm.org/
Replication of the genomes of many different viruses, in-cluding herpes viruses and Ad, occurs in nuclear compart-ments. Often, these replication compartments initially associ-ate with distinct nuclear structures termed ND10 or promyelocytic leukemia (PML) nuclear bodies (NBs) (8, 18, 40). Overexpression of PML affects replication of vesicular stomatitis virus and influenza virus (33), and the number and intensities of PML NBs increases in response to interferon (32). Moreover, many viruses, including herpes viruses and Ad encode proteins that bind and reorganize PML NBs (1, 6, 9, 21, 28, 30, 53). For example, the Ad E4-ORF3 protein binds and redistributes PML NBs to filamentous structures (4, 6, 21, 28), whereas HSV-1 ICP0 and the cytomegalovirus immediate-early 1 protein completely disrupt PML NBs (1, 9). Many of these results strongly suggest that PML NBs may have an antiviral effect. In particular, data by Chee et al. indicate that PML mediates an interferon-induced anti-HSV-1 state (5), although dispersal of PML NBs is not essential for HSV-1 replication (23). Therefore, it is not clear whether PML NBs in general have, if any, positive or negative effects on virus rep-lication (8).
While the molecular mechanisms of AAV DNA replication are well investigated (2), relatively few studies report on the spatial organization of AAV DNA and proteins within the nucleus of the infected cell (13, 16, 52, 54). Using immunoflu-orescence microscopy, Hunter and Samulski (16) demon-strated that all four Rep proteins occupy the same intranuclear compartments and that Rep and capsid proteins colocalized. Immunofluorescence and in situ hybridization experiments in-dicated that AAV DNA or rAAV DNA colocalize with Ad replication compartments (52) and that early during AAV and Ad coinfection, AAV Rep forms punctate structures in the cell nucleus which colocalize with AAV DNA (54).
In this study, we have monitored the spatial and temporal organization of AAV DNA replication in live cells. The method is based on lac operator (lacO)-lac repressor (LacI) interactions and includes an rAAV that contains 40 LacI bind-ing sites between the ITRs (rAAVlacO) and a reporter mole-cule consisting of enhanced yellow fluorescent protein (EYFP) linked to LacI (EYFP-LacI). AAV Rep protein and helper virus-mediated replication of the rAAV genome was visualized by binding of the EYFP-lacrepressor fusion protein to double-stranded replication intermediates of the AAV ITR-flanked
lacO cassette. Furthermore, we constructed a plasmid that expresses the first 522 amino acids of Rep78/68 linked to a red fluorescent protein (DsRed2) from the AAV p5 promoter to simultaneously visualize AAV DNA replication and Rep pro-tein in live cells. In addition, a plasmid that expressed en-hanced cyan fluorescent protein (ECFP) fused to PML also allowed the visualization of PML NBs in live cells (10, 29). We demonstrate that (i) AAV DNA replication occurs in compart-ments and the formation of these compartcompart-ments was depen-dent on AAV Rep, AAV ITRs, and helper virus and (ii) AAV replication compartments colocalize with AAV Rep foci. Fur-thermore, we show that in Ad-infected cells, AAV replication compartments can associate with modified PML NBs. How-ever, neither absence of PML NBs (wild-type HSV-1-infected cells) nor overexpression and presence of unmodified PML (HSV-1 ICP0 deletion mutant-infected cells) had an effect on
the formation and development of AAV DNA replication compartments.
MATERIALS AND METHODS
Cells and viruses.VERO 2-2 cells (38) and HeLa cells were maintained in Dulbecco’s modified Eagle medium supplemented with 10% fetal bovine serum. Stocks of Ad type 2 were provided by U. Greber (University of Zurich). Wild-type HSV-1 strain F was grown and its titers were determined in VERO 2-2 cells. Recombinant HSV-1 dl1403, which contains a deletion in both copies of the ICP0 gene (42), and recombinant HSV-1 FXE, which contains a deletion in the Ring-finger domain of ICP0 (7), were obtained from P. Lomonte (University of Lyon). Because of the absence of ICP0, HSV-1 dl1403 does not mediate the redistribution of PML; because of the mutation in the Ring-finger domain, HSV-1 FXE-encoded ICP0 can bind but not disrupt PML NBs (9).
Plasmids.Plasmids pSV2-EYFP/lacI, which expresses EYFP linked to thelac
repressor (LacI), and p16PC(B⫺/⫹), which contains 10 kb of 292-bplac
oper-ator (lacO) repeats, were kindly provided by D. L. Spector (Cold Spring Harbor
Laboratory, Cold Spring Harbor, N.Y.) (45). p16PC(B⫺/⫹) was partially
di-gested withEcoRI, and a fragment of⬃1.5 kb, which includes 5 copies of the
292-bplacOsequence comprising 40 LacI binding sites, was inserted into the
EcoRI site of pBluescript SK(⫺); the resulting plasmid was named pBslacO. Plasmid pAAVlacO contains the AAV ITRs flanking 40 LacI binding sites and the bovine growth hormone polyadenylation signal (bGHpA). This plasmid was
generated by inserting the 1.5-kbSpeI-EcoRV fragment from pBslacO into the
SpeI andPmeI sites located between the AAV ITRs of pAV2GFP (12). The AAVrep-expressing plasmid, p-rep, has been described previously (12). Plasmid
p-rep-red was constructed by inserting the ⬃1.7-kbBglII-HindIII fragment,
which includes the AAV p5 promoter and the first 522 codons ofrep78/68, from
p5R78 (kindly provided by M. Urabe, Jichi Medical School, Tochigi, Japan) (44)
between theBglII andHindIII sites of pDsRed2-3ad (kindly provided by B.
Vogt). Bacterial artificial chromosome fHSV⌬pac⌬27⌬Kn and plasmid
pEB-HICP27 together represent a replication-competent, packaging-defective HSV-1 genome (35) and were used in some experiments to provide HSV-1 helper functions for rAAV replication (12). Plasmid pECFP-PML, which expresses ECFP fused to PML (isoform IV) was a gift from R. D. Everett (MRC Virology Unit, Glasgow, United Kingdom). This fusion protein is known to associate with PML NBs and respond to HSV-1 ICP0 (10, 29).
ITR rescue/replication assay.ITR rescue/replication assays were performed
essentially as described by Heister et al. (12). Briefly, 106VERO 2-2 cells were
plated on 6-cm-diameter tissue culture plates. The following day, the cultures
were cotransfected with 0.5g of pAAVlacO, 2g of fHSV⌬pac⌬27⌬Kn, 0.2g
of pEBHICP27, and 0.2g of either p-rep or p-rep-red by using the
Lipo-fectamine procedure as described by the manufacturer (Life Technologies, Basel, Switzerland). Control transfections in the absence of pAAVlacO, AAV rep, or helper virus functions were also performed. After 2 days, extrachromo-somal DNA was isolated by the procedure described by Hirt (14). The DNA was
digested extensively withDpnI, separated on a 0.7% agarose gel, and transferred
to a nylon membrane (Hybond N⫹; Amersham). Hybridization with a
digoxi-genin-labeled bGHpA probe and immunological detection with an alkaline phos-phatase-conjugated anti-digoxigenin antibody and chemiluminescence substrate (CDP Star) were performed as described by the supplier (Roche Diagnostics, Rotkreuz, Switzerland). The bGHpA probe was a 264-bp fragment derived from
plasmid pHyRaNGFPa (12) by PCR amplification with primers pGHpA1 (5⬘-G
ATCAGCCTCGACTGTGCCTTC-3⬘) and pGHpA2 (5⬘-CTCCATCACTAGG
GGTTCCTTG-3⬘) and the PCR digoxigenin probe synthesis kit (Roche
Diag-nostics). PCR conditions were as follows. An initial denaturation step at 94°C for 2 min was followed by 35 cycles of amplification for 1 min at 94°C, 30 s at 56°C, and 1 min at 72°C. For final extension, the reaction mixture was incubated at 75°C for 10 min.
Production of rAAV stocks.rAAVlacO was produced in 293T cells by trans-fection of pAAVlacO and pDG, which provides AAV Rep and Cap proteins as well as Ad helper functions (11). The virus preparation was purified by iodixanol gradient ultracentrifugation and heparin-Sepharose high-performance liquid chromatography, and virus titers (genome-containing particles per milliliter) were determined by slot blot hybridization with a probe specific for bGHpA.
Visualization of rAAV DNA replication. (i) Transfection-infection procedure. The day before transfection-infection, 20,000 cells (HeLa or VERO 2-2) were plated on Lab-Tek four-well chamber slides (Nalgene Nunc International, Na-perville, Ill.) for standard fluorescence microscopy or on glass-bottom no. 0 tissue culture plates (MaTek Corporation, Ashland, Mass.) for time-lapse and confocal microscopy. The amounts of the individual plasmids used for transfection (Li-pofectamine) were as follows: pAAVlacO or pBslacO, 50 ng; pSV2-EYFP/lacI,
on November 8, 2019 by guest
http://jvi.asm.org/
6.5 ng; pEBHICP27, pECFP-PML, p-rep, or p-rep-red, 25 ng; fHSV⌬pac
⌬27⌬Kn, 125 ng. Variable amounts of pBluescript SK(⫺) were added to adjust
the total amount of DNA to 231.5 ng. In some experiments, helper virus
func-tions from fHSV⌬pac⌬27⌬Kn and pEBHICP27 were replaced by infecting the
cells at 4 h after transfection with either HSV-1 or Ad. HSV-1 was used at a multiplicity of infection (MOI) of 2 PFU per cell, and Ad was used at an MOI of 100 PFU per cell. In some experiments, the cells were transfected only with p-rep (or p-rep-red) and pSV2-EYFP/lacI, and 4 h later, cells were coinfected
with rAAVlacO at an MOI of 5⫻103genome-containing particles per cell and
either HSV-1 or Ad. Control transfections-infections in the absence of
p(r)AAV-lacO, AAVrep, or helper virus functions were also performed. Cultures of live
cells or fixed cells were examined by confocal, time-lapse, or standard fluores-cence microscopy between 12 and 48 h after transfection-infection. For bro-modeoxyuridine (BrdU) incorporation assays, cells were labeled with BrdU (1 mM) from 16 to 24 h after transfection.
(ii) Immunofluorescence.For immunofluorescence, cultures were washed with phosphate-buffered saline (PBS) and fixed with paraformaldehyde (4% in PBS) for 20 min. Fixation was stopped by incubation in 0.1 M glycine (in PBS) for 10 min, and the cells were permeabilized with 0.2% Triton X-100 in PBS for 15 min. For BrdU detection, the cells were treated with 4 M HCl for 10 min. The primary antibodies against PML (sc-966; Santa Cruz Biotechnology, Inc., Santa Cruz, Calif.) or against BrdU (Roche Diagnostics) were diluted 1:100 in PBS contain-ing 0.2% Triton X-100 and 1% bovine serum albumin (PBS-Triton-BSA). After incubation for 1 h at room temperature, the cells were washed 3 times for 5 min with PBS and then incubated for 1 h at room temperature with rhodamine- or fluorescein isothiocyanate-conjugated secondary antibodies diluted 1:100 in PBS-Triton-BSA. Coverslips were mounted in glycerol gelatin containing 2.5% DABCO to retard bleaching.
(iii) Standard fluorescence microscopy.Cultures were observed with a Zeiss Axiovert 100 microscope equipped with filters specific for enhanced green fluo-rescent protein, EYFP, ECFP, and DsRed2 and a Hamamatsu charge-coupled device digital camera.
(iv) Time-lapse microscopy.A wide-field fluorescence microscope (Leica DM-IRBE) equipped with a filter specific for EYFP was housed in a humidified
gas-tight box with temperature-controlled air circulation and a CO2detection
and maintenance device (Ludin; Life Imaging Services, Reinach, Switzerland). Images were captured with a Hamamatsu charge-coupled device digital camera. (v) Confocal microscopy.Live or fixed cultures were analyzed by confocal microscopy with a Leica SP2 laser scanning microscope with settings specific for EYFP, DsRed2, and rhodamine. The different channels were detected sequen-tially, and the laser power and detection windows were adjusted for each channel to exclude overlap between different fluorochromes. The images were processed by using Imaris (Bitplane, Zurich, Switzerland) and Volocity (Improvision, Cov-entry, United Kingdom) software.
RESULTS
Visualization of rAAV DNA replication in live cells.lac
op-erator-repressor interactions combined with autofluorescent proteins have been previously employed to visualize funda-mental cellular processes, such as chromatin organization (43), gene activity (45), or movement of replication origins (50). We set out to adapt these principles for visualizing the spatial and temporal organization of AAV DNA replication in live cells. We constructed pAAVlacO, which contains the AAV ITRs flanking 40lacrepressor (LacI) binding sites. Because of the ITRs, pAAVlacO can serve as substrate for AAV Rep protein and helper virus-mediated DNA replication, which was as-sumed to increase the number of LacI binding sites to levels that should support visualization by EYFP linked to the DNA binding protein LacI (Fig. 1A). We proceeded to test the functionality of pAAVlacO by ITR rescue/replication assay (Fig. 1B). In the presence of AAV Rep and HSV-1 helper functions, pAAVlacO produced DpnI-resistant monomers, dimers, and higher-order multimers of the ITR cassette (Fig. 1B, lane 2). No replication intermediates of the ITR cassette were visible in the absence of either AAV Rep (Fig. 1B, lane 1), HSV-1 helper functions (Fig. 1B, lane 4), or pAAVlacO
(Fig. 1B, lane 5). Also, the ITR cassette from pAAVlacO was efficiently rescued and packaged into AAV virions in the pres-ence of Ad helper functions and AAV Rep and Cap, further showing that pAAVlacO was a functional substrate for AAV DNA replication. The resulting rAAVlacO virus stocks had titers of 2⫻1012genome-containing particles per ml (data not
shown).
A first experiment to visualize rAAV DNA replication was carried out in VERO 2-2 cells (Fig. 2A). Cells transfected with pAAVlacO, pSV2-EYFP/lacI, the rep-expressing plasmid p-rep, and HSV-1 helper DNA formed yellow fluorescent, nu-clear compartments by 16 h posttransfection (p.t.), which in-creased in size over time. These compartments colabeled with BrdU, indicating that they are sites of active DNA synthesis (Fig. 2B). The expression of the EYFP-lacIfusion gene did not appear to be rate limiting for the kinetics of the formation of the nuclear dots, as a faint and diffuse nuclear fluorescence was visible throughout the course of infection (Fig. 2A). In the absence of either AAV Rep (Fig. 2Ab) or HSV-1 helper DNA (Fig. 2Ac), no dots were observed but a diffuse nuclear fluo-rescence was observed. The nuclear fluofluo-rescence remained diffuse also when pBslacO, which contains the 40 LacI binding sites but no AAV ITRs, was cotransfected with p-rep and HSV-1 helper DNA (Fig. 2Ad). In summary, the nuclear dots are sites of active DNA synthesis (BrdU incorporation) and formed only under conditions compatible with AAV DNA replication; the absence of ITRs, Rep, or helper virus functions resulted in diffuse nuclear fluorescence.
Next, we carried out visualization experiments in HeLa cells
FIG. 1. (A) The principle of visualizing AAV DNA replication. Plasmid pAAVlacO (or rAAV [rAAVlacO]) contains the AAV ITR-flanking 5 copies of thelacoperator sequence (lacO), which comprise a total of 40lacrepressor (LacI) binding sites, and the bGHpA. Helper virus and AAV Rep-mediated formation of double-stranded, mono-meric (ITRm) and dimono-meric (ITRd) AAV replication intermediates is visualized by EYFP fused to LacI (ⴱ), which is expressed from pSV2-EYFP/lacI. (B) Southern analysis of AAV ITR rescue/replication. VERO 2-2 cells were cotransfected with pAAVlacO and HSV-1 helper DNA (lane 1); pAAVlacO, HSV-1 helper DNA, and either p-rep (lane 2) or p-rep-red (lane 3); pAAVlacO and p-rep (lane 4); or HSV-1 helper DNA and p-rep (lane 5). Hirt DNA prepared 48 h later was digested withDpnI and analyzed by Southern blot with a digoxigenin-labeled bGHpA probe. ITRm and ITRd are indicated (arrows). The fragments of the digoxigenin-labeled molecular size standard (M) have the sizes 23.1, 9.4, 6.6, 4.3, 2.3, and 2.0 kb (Roche Diagnostics).⫹, present;⫺, absent.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:3.603.311.531.71.225.2]on November 8, 2019 by guest
http://jvi.asm.org/
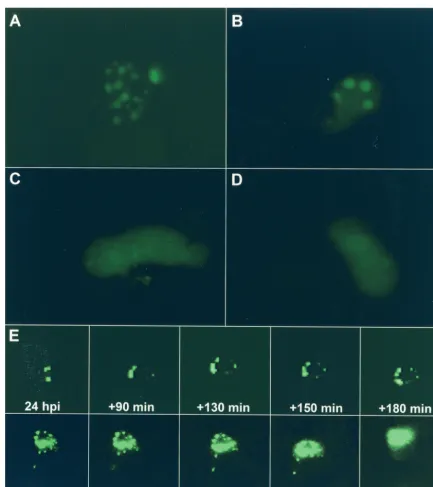
and used infection with rAAV (rAAVlacO), as opposed to transfection of pAAVlacO, to provide the substrate for AAV DNA replication (Fig. 3). Cells cotransfected with p-rep and pSV2-EYFP/lacI and subsequently infected with rAAVlacO and helper virus, either HSV-1 (Fig. 3A) or Ad (Fig. 3B), developed nuclear dots by 14 to 20 h postinfection (p.i.). The numbers of replication compartments were comparable in Ad-and HSV-1-infected cells Ad-and ranged from 2 to more than 40 per cell nucleus. The proportion of cells that showed AAV replication compartments appeared to be slightly higher when Ad was used as the helper virus and was in general between 1 and 5% of all cells. However, these numbers may be strongly influenced by transfection efficiency and dose of both rAAV and helper virus. Nuclear fluorescence remained diffuse in the absence of either helper virus (Fig. 3C) or AAV Rep (Fig. 3D). Figure 3E shows selected time points of a time-lapse series of HeLa cells transfected with p-rep and pSV2-EYFP/lacI and infected with rAAVlacO and Ad. The first frame was taken at 24 h p.i. and shows two relatively large replication compart-ments. Several new replication compartments appear later, expand, and fuse to much larger structures.
AAV Rep forms nuclear foci which colocalize with AAV DNA
replication compartments. To analyze in live cells whether
AAV DNA replication compartments colocalize with AAV Rep, we constructed p-rep-red, which uses the AAV p5 pro-moter to expresses the first 522 codons ofrep78/68linked to a red fluorescent protein (DsRed2). The functionality of this truncated and modified Rep-Red fusion protein was first con-firmed by ITR rescue/replication assay, which clearly yielded
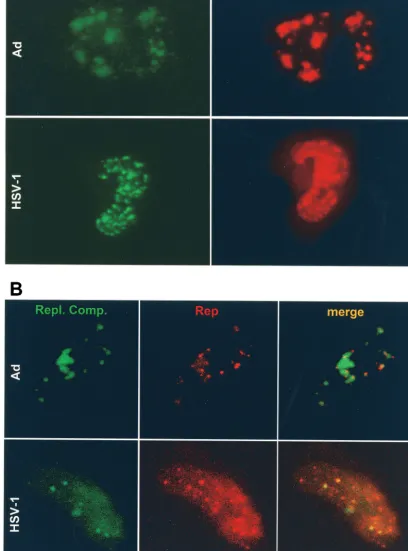
DpnI-resistant monomeric and dimeric replication intermedi-ates of the ITR cassette (Fig. 1B, lane 3), although at lower levels than those produced in the presence of wild-type Rep (p-rep) (Fig. 1B, lane 2). Cells transfected with p-rep-red and superinfected with rAAVlacO and either Ad or HSV-1 formed nuclear red fluorescent foci (Fig. 4). As judged by standard fluorescence microscopy, AAV DNA replication compart-ments (green) and Rep foci (red) appeared to occupy the same areas (Fig. 4A). Confocal microscopy confirmed that AAV replication compartments colocalized with red fluorescent Rep foci (Fig. 4B). Of note, the intensity of Rep (red) was consis-tently much higher when HSV-1 was used as the helper virus.
AAV DNA replication compartments and PML NBs.The
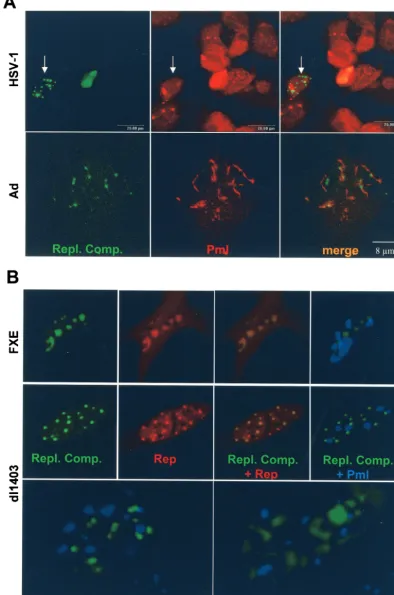
replication compartments of HSV-1 and Ad are associated, at least at some point, with PML NBs. However, both viruses also encode proteins that bind and reorganize PML NBs (for a review, see reference 8). In particular, HSV-1 ICP0 binds and disrupts PML NBs while Ad E4-ORF3 alters PML NB mor-phology to fibrous structures. In the following experiment, we addressed the question of whether AAV DNA replication also associates with PML NBs. HeLa cells were transfected with p-rep and pSV2-EYFP/lacI and infected with rAAVlacO and either HSV-1 or Ad. When AAV DNA replication compart-ments were well visible under the fluorescence microscope (14 to 18 h p.i.), the cells were fixed and stained with a PML-specific antibody and a rhodamine-conjugated secondary anti-body (Fig. 5A). Coexistence of AAV DNA replication com-partments and PML NBs was not observed when wild-type HSV-1 was used as the helper virus (Fig. 5A, top panels). Therefore, the formation of AAV replication compartments appears to not be dependent on the presence of PML NBs.
FIG. 2. Visualization of AAV DNA replication in live VERO cells. (A) Cells were cotransfected with pAAVlacO, pSV2-EYFP/lacI, p-rep, and HSV-1 helper DNA (a). Control experiments include cells cotransfected with pAAVlacO, pSV2-EYFP/lacI, and HSV-1 helper DNA (no Rep) (b); pAAVlacO, pSV2-EYFP/lacI, and p-rep (no helper virus DNA) (c); or pBslacO, pSV2-EYFP/lacI, p-rep, and HSV-1 helper DNA (no ITRs) (d). Photographs in panels a were taken from the same cell as indicated between 16 and 22 h p.t. wit h a standard fluorescence microscope. Photographs b to d were taken at 24 h p.t. (B) Confocal microscopy of VERO cells cotransfected with pAAVlacO, pSV2-EYFP/lacI, p-rep, and H SV-1 helper DNA. After 16 h, the cells were labeled for 8 h with BrdU, fixed, and stained with a BrdU-speci fic antibody and a rhodamine-conjugated secondary antibody. (a) EYFP (AAV replication compartments); (b) rhodamine (BrdU incorporation); (c) merge.
on November 8, 2019 by guest
http://jvi.asm.org/
With Ad as the helper virus, PML NBs were visible but exhib-ited an altered, fibrous morphology; nevertheless, some repli-cation compartments were associated with these modified PML NBs (Fig. 5A, bottom panels). To specifically examine the question of whether disruption or modification of PML NBs is required for AAV replication, we performed the fol-lowing live cell visualization experiment. Cells were transfected
[image:6.603.74.508.65.553.2]with pSV2-EYFP/lacI, prep-red, and pECFP-PML and then superinfected with rAAVlacO and recombinant HSV-1 helper virus, FXE or dl1403 (Fig. 5B). Although HSV-1 FXE-en-coded ICP0 can bind but not disrupt PML NBs, as it contains a mutation in the Ring-finger domain, and HSV-1 dl1403 does not modify PML NBs because it lacks both copies of the ICP0 gene (7, 9), the formation of AAV replication compartments
FIG. 3. Visualization of AAV DNA replication in live HeLa cells. Cells were cotransfected with pSV2-EYFP/lacI and p-rep and subsequently infected with rAAVlacO and helper virus, either HSV-1 (A) or Ad (B). Control experiments include cells cotransfected with pSV2-EYFP/lacI and p-rep and infected with rAAVlacO alone (no helper virus) (C) and cells transfected with pSV2-EYFP/lacI alone and infected with rAAVlacO and HSV-1 (no Rep) (D). The cultures were examined under the fluorescence microscope from 16 to 28 h p.t. (E) Time-lapse microscopy of developing AAV DNA replication compartments in HeLa cells transfected and infected as described for panel B. The first frame was taken at 24 h after coinfection with rAAVlacO and Ad; further frames were taken every 10 min over a 6-h period. Selected images at the indicated times (minutes) after the first frame (24 h) are shown.
on November 8, 2019 by guest
http://jvi.asm.org/
FIG. 4. Visualization of AAV DNA replication compartments (green) and AAV Rep (red) in live HeLa cells. (A) Cells were transfected with pAAVlacO, pSV2-EYFP/lacI, and p-rep-red and infected with Ad (top) or HSV-1 (bottom). Images were taken between 14 and 24 h p.i. with a standard fluorescence microscope and filters specific for enhanced green fluorescent protein-EYFP (AAV replication compartments [Repl. Comp.]) or DsRed2 (AAV Rep). (B) Confocal microscopy of HeLa cells transfected with pSV2-EYFP/lacI and p-rep-red and infected with rAAVlacO and either Ad (top) or HSV-1 (bottom). The cell in the top panels is living; the cell in the bottom panels is fixed. Microscope settings were specific for EYFP (AAV replication compartments) or DsRed2 (AAV Rep).
395
on November 8, 2019 by guest
FIG. 5. (A) Confocal microscopy of HeLa cells transfected with pSV2-EYFP/lacI and p-rep and infected with rAAVlacO and either HSV-1 (top) or Ad (bottom). After 14 to 18 h, the cells were fixed and stained with a PML-specific antibody and a rhodamine-conjugated secondary antibody. Microscope settings were specific for EYFP (AAV replication compartments [Repl. Comp.]) or rhodamine (PML NBs). The arrows in the top three panels point to a cell that contains replication compartments but no PML NBs. PML NBs are visible in most surrounding cells. (B) Simultaneous visualization of AAV DNA replication compartments (green), AAV Rep (red), and PML NBs (blue) in live HeLa cells. Cells were transfected with pSV2-EYFP/lacI, p-rep-red, and pECFP-PML and infected with rAAVlacO and either HSV-1 FXE or HSV-1 dl1403 as the helper virus. Images were taken between 14 and 24 h p.i. from individual cells by using different fluorescence filters. The bottom two panels show AAV replication compartments and PML NBs of two individual cells at a higher magnification.
396
on November 8, 2019 by guest
[image:8.603.97.492.56.651.2]was equally efficient with either helper virus tested, ICP0 mu-tants, wild-type HSV-1, or Ad. These results demonstrate that the formation of AAV replication compartments is not af-fected by the presence of unmodified PML NBs. Although AAV replication compartments and PML NBs coexisted in HSV-1 FXE- or dl1403-infected cells, no association was ob-served under these conditions (Fig. 5B, right and bottom pan-els).
DISCUSSION
The molecular mechanisms of AAV DNA replication are well investigated (2), but relatively few studies report on the spatial organization of AAV DNA and proteins within the host cell nucleus (13, 16, 52, 54). Hunter and Samulski (16) showed that in Ad-infected cells, all four Rep proteins colocalized in distinct nuclear foci. The foci increased in size and filled out the entire nucleus as the infection progressed. Similar experi-ments in Ad-infected cells by Wistuba et al. (54) also showed a punctate pattern of Rep. Moreover, Rep was shown to co-localize with AAV DNA, indicating that AAV replication oc-curs in compartments, similar to the replication compartments of other viruses. Using immunofluorescence and in situ hybrid-ization, Weitzman et al. (52) demonstrated that AAV DNA was, in fact, recruited into Ad replication compartments.
We have visualized the spatial and temporal organization of AAV DNA replication and AAV Rep in live cells. Our results confirm previous results on fixed cells that AAV DNA repli-cation occurs in compartments which colocalize with AAV Rep foci. In addition, our assay revealed that several small replication compartments rapidly expand and fuse to larger structures that fill the nucleus within 4 to 8 h.
We also detected some association of AAV DNA replication compartments with modified PML NBs when Ad was used as the helper virus. No such association was observed with HSV-1 as the helper virus, although HSV-1 DNA replication itself initially associates with PML NBs (24, 40). The HSV-1 ICP0 protein is known to disrupt PML NBs (9). As AAV replication requires expression of early HSV-1 genes (51), we hypothesize that ICP0, which is expressed with immediate-early kinetics, disrupts PML NBs before AAV DNA replication starts or is visible in our assay. By contrast, Ad E4-ORF3 does not disin-tegrate but changes the morphology of PML NBs (4, 6, 21, 28) and, therefore, allowed codetection of AAV replication com-partments and modified PML NBs in our assay. Interestingly, the absence of Ad E4 gene products has been shown to reduce the mobilization of AAV DNA into replication centers (52). Thus, we hypothesized that disruption of PML NB morphology may be important for making the host cell competent for AAV replication. However, this was not the case, as AAV replication compartments formed efficiently with recombinant HSV-1 helper viruses that do not affect PML.
The live cell visualization assay established for this study can be applied to examine various aspects of replication of AAV and potentially other viruses. The advantage over experiments with immunofluorescence and in situ hybridization on fixed cells is that it is possible to create a dynamic, four-dimensional picture of events, such as replication, reactivation, and perhaps even genomic integration. Furthermore, formation of replica-tion compartments in response to various treatments,
includ-ing antivirals, mutant helper viruses, and mutant rep or cap
genes can be studied in real time. Sourvinos and Everett (40) have utilized the tetracycline operator-repressor interaction combined with fluorescent proteins to visualize the formation of HSV-1 amplicon replication compartments in live cells. This system can be easily combined with the visualization of AAV replication described in this study to simultaneously analyze the formation and interactions of replication compartments of AAV and its HSV-1 helper virus.
ACKNOWLEDGMENTS
We thank U. Greber, P. Lomonte, D. L. Spector, M. Urabe, B. Vogt, and R. D. Everett for providing reagents and T. Heister for technical assistance.
This work was supported by the Swiss National Science Foundation no. 3100-100195 (to C.F.).
REFERENCES
1. Ahn, J. H., and G. S. Hayward.1997. The major immediate-early proteins IE1 and IE2 of human cytomegalovirus colocalize with and disrupt PML-associated nuclear bodies at very early times in infected permissive cells.
J. Virol.71:4599–4613.
2. Berns, K. I.1996.Parvoviridae: the viruses and their replication, p. 2173–
2197.InB. N. Fields, D. M. Knipe, and P. M. Howley (ed.), Fields virology,
vol. 3. Lippincott-Raven, Philadelphia, Pa.
3. Brister, J. R., and N. Muzyczka.1999. Rep-mediated nicking of the adeno-associated virus origin requires two biochemical activities, DNA helicase
activity and transesterification. J. Virol.73:9325–9336.
4. Carvalho, T., J. S. Seeler, K. Ohman, P. Jordan, U. Pettersson, G. Akusjarvi, M. Carmo-Fonseca, and A. Dejean.1995. Targeting of adenovirus E1A and E4-ORF3 proteins to nuclear matrix-associated PML bodies. J. Cell Biol. 131:45–56.
5. Chee, A. V., P. Lopez, P. P. Pandolfi, and B. Roizman.2003. Promyelocytic leukemia protein mediates interferon-based anti-herpes simplex virus 1
ef-fects. J. Virol.77:7101–7105.
6. Doucas, V., A. M. Ishov, A. Romo, H. Juguilon, M. D. Weitzman, R. M. Evans, and G. G. Maul.1996. Adenovirus replication is coupled with the
dynamic properties of the PML nuclear structure. Genes Dev.10:196–207.
7. Everett, R. D.1989. Construction and characterization of herpes simplex virus type 1 mutants with defined lesions in immediate early gene 1. J. Gen.
Virol.70:1185–1202.
8. Everett, R. D.2001. DNA viruses and viral proteins that interact with PML
nuclear bodies. Oncogene20:7266–7273.
9. Everett, R. D., and G. G. Maul.1994. HSV-1 IE protein Vmw110 causes
redistribution of PML. EMBO J.13:5062–5069.
10. Everett, R. D., G. Sourvinos, and A. Orr.2003. Recruitment of herpes simplex virus type 1 transcriptional regulatory protein ICP4 into foci
juxta-posed to ND10 in live, infected cells. J. Virol.77:3680–3689.
11. Grimm, D., A. Kern, K. Rittner, and J. A. Kleinschmidt.1998. Novel tools for production and purification of recombinant adenoassociated virus
vec-tors. Hum. Gene Ther.9:2745–2760.
12. Heister, T., I. Heid, M. Ackermann, and C. Fraefel.2002. Herpes simplex virus type 1/adeno-associated virus hybrid vectors mediate site-specific inte-gration at the adeno-associated virus preinteinte-gration site, AAVS1, on human
chromosome 19. J. Virol.76:7163–7173.
13. Henry, C. J., L. P. Merkow, M. Pardo, and C. McCabe.1972. Electron microscope study on the replication of AAV-1 in herpes-infected cells.
Vi-rology49:618–621.
14. Hirt, B.1969. Replicating molecules of polyoma virus DNA. J. Mol. Biol. 40:141–144.
15. Hong, G., P. Ward, and K. I. Berns.1994. Intermediates of adeno-associated
virus DNA replication in vitro. J. Virol.68:2011–2015.
16. Hunter, L. A., and R. J. Samulski.1992. Colocalization of adeno-associated
virus Rep and capsid proteins in the nuclei of infected cells. J. Virol.66:
317–324.
17. Im, D. S., and N. Muzyczka.1989. Factors that bind to adeno-associated
virus terminal repeats. J. Virol.63:3095–3104.
18. Ishov, A. M., and G. G. Maul.1996. Nuclear domain 10 as preexisting potential replication start sites of herpes simplex virus type-1. Virology 217:67–75.
19. Kotin, R. M., R. M. Linden, and K. I. Berns.1992. Characterization of a preferred site on human chromosome 19q for integration of
adeno-associ-ated virus DNA by non-homologous recombination. EMBO J.11:5071–5078.
20. Kotin, R. M., M. Siniscalco, R. J. Samulski, X. D. Zhu, L. Hunter, C. A. Laughlin, S. McLaughlin, N. Muzyczka, M. Rocchi, and K. I. Berns.1990. Site-specific integration by adeno-associated virus. Proc. Natl. Acad. Sci. USA87:2211–2215.
on November 8, 2019 by guest
http://jvi.asm.org/
21. Leppard, K. N., and R. D. Everett.1999. The adenovirus type 5 E1b 55K and E4 Orf3 proteins associate in infected cells and affect ND10 components.
J. Gen. Virol.80:997–1008.
22. Linden, R. M., E. Winocour, and K. I. Berns.1996. The recombination signals for adeno-associated virus site-specific integration. Proc. Natl. Acad.
Sci. USA93:7966–7972.
23. Lopez, P., R. J. Jacob, and B. Roizman.2002. Overexpression of promyelo-cytic leukemia protein precludes the dispersal of ND10 structures and has no effect on accumulation of infectious herpes simplex virus 1 or its proteins.
J. Virol.76:9355–9367.
24. Lukonis, C. J., and S. K. Weller.1997. Formation of herpes simplex virus type 1 replication compartments by transfection: requirements and
localiza-tion to nuclear domain 10. J. Virol.71:2390–2399.
25. Lusby, E., K. H. Fife, and K. I. Berns.1980. Nucleotide sequence of the
inverted terminal repetition in adeno-associated virus DNA. J. Virol.34:
402–409.
26. McCarty, D. M., D. J. Pereira, I. Zolotukhin, X. Zhou, J. H. Ryan, and N. Muzyczka.1994. Identification of linear DNA sequences that specifically
bind the adeno-associated virus Rep protein. J. Virol.68:4988–4997.
27. Monahan, P. E., and R. J. Samulski.2000. AAV vectors: is clinical success
on the horizon? Gene Ther.7:24–30.
28. Muller, S., and A. Dejean.1999. Viral immediate-early proteins abrogate the modification by SUMO-1 of PML and Sp100 proteins, correlating with
nuclear body disruption. J. Virol.73:5137–5143.
29. Parkinson, J., and R. D. Everett.2000. Alphaherpesvirus proteins related to herpes simplex virus type 1 ICP0 affect cellular structures and proteins.
J. Virol.74:10006–10017.
30. Puvion-Dutilleul, F., M. K. Chelbi-Alix, M. Koken, F. Quignon, E. Puvion, and H. de The.1995. Adenovirus infection induces rearrangements in the intranuclear distribution of the nuclear body-associated PML protein. Exp.
Cell Res.218:9–16.
31. Recchia, A., R. J. Parks, S. Lamartina, C. Toniatti, L. Pieroni, F. Palombo, G. Ciliberto, F. L. Graham, R. Cortese, N. La Monica, and S. Colloca.1999. Site-specific integration mediated by a hybrid adenovirus/adeno-associated
virus vector. Proc. Natl. Acad. Sci. USA96:2615–2620.
32. Regad, T., and M. K. Chelbi-Alix.2001. Role and fate of PML nuclear bodies
in response to interferon and viral infections. Oncogene20:7274–7286.
33. Regad, T., A. Saib, V. Lallemand-Breitenbach, P. P. Pandolfi, H. de The, and M. K. Chelbi-Alix.2001. PML mediates the interferon-induced antiviral state against a complex retrovirus via its association with the viral transactivator.
EMBO J.20:3495–3505.
34. Ryan, J. H., S. Zolotukhin, and N. Muzyczka.1996. Sequence requirements for binding of Rep68 to the adeno-associated virus terminal repeats. J. Virol. 70:1542–1553.
35. Saeki, Y., C. Fraefel, T. Ichikawa, X. O. Breakefield, and E. A. Chiocca.2001. Improved helper virus-free packaging system for HSV amplicon vectors using an ICP27-deleted, oversized HSV-1 DNA in a bacterial artificial
chro-mosome. Mol. Ther.3:591–601.
36. Samulski, R. J., K. I. Berns, M. Tan, and N. Muzyczka.1982. Cloning of adeno-associated virus into pBR322: rescue of intact virus from the
recom-binant plasmid in human cells. Proc. Natl. Acad. Sci. USA79:2077–2081.
37. Samulski, T., T. Shenk, R. J. Samulski, and F. Rolling.1996. AAV as a viral vector for human gene therapy. Generation of recombinant virus. J. Virol. 70:3227–3234.
38. Smith, I. L., M. A. Hardwicke, and R. M. Sandri-Goldin.1992. Evidence that the herpes simplex virus immediate early protein ICP27 acts
post-transcrip-tionally during infection to regulate gene expression. Virology186:74–86.
39. Snyder, R. O., R. J. Samulski, and N. Muzyczka.1990. In vitro resolution of
covalently joined AAV chromosome ends. Cell60:105–113.
40. Sourvinos, G., and R. D. Everett.2002. Visualization of parental HSV-1 genomes and replication compartments in association with ND10 in live
infected cells. EMBO J.21:4989–4997.
41. Srivastava, A., E. W. Lusby, and K. I. Berns.1983. Nucleotide sequence and
organization of the adeno-associated virus 2 genome. J. Virol.45:555–564.
42. Stow, N. D., and E. C. Stow.1986. Isolation and characterization of a herpes simplex virus type 1 mutant containing a deletion within the gene encoding
the immediate early polypeptide Vmw110. J. Gen. Virol.67:2571–2585.
43. Straight, A. F., W. F. Marshall, J. W. Sedat, and A. W. Murray.1997. Mitosis
in living budding yeast: anaphase A but no metaphase plate. Science277:
574–578.
44. Surosky, R. T., M. Urabe, S. G. Godwin, S. A. McQuiston, G. J. Kurtzman, K. Ozawa, and G. Natsoulis.1997. Adeno-associated virus Rep proteins target DNA sequences to a unique locus in the human genome. J. Virol. 71:7951–7959.
45. Tsukamoto, T., N. Hashiguchi, S. M. Janicki, T. Tumbar, A. S. Belmont, and D. L. Spector.2000. Visualization of gene activity in living cells. Nat. Cell
Biol.2:871–878.
46. Wang, Y., S. M. Camp, M. Niwano, X. Shen, J. C. Bakowska, X. O. Breake-field, and P. D. Allen.2002. Herpes simplex virus type 1/adeno-associated
virus rep(⫹) hybrid amplicon vector improves the stability of transgene
expression in human cells by site-specific integration. J. Virol.76:7150–7162.
47. Ward, P., and K. I. Berns.1996. In vitro replication of adeno-associated virus DNA: enhancement by extracts from adenovirus-infected HeLa cells. J. Vi-rol.70:4495–4501.
48. Ward, P., and K. I. Berns.1995. Minimum origin requirements for linear
duplex AAV DNA replication in vitro. Virology209:692–695.
49. Ward, P., M. Falkenberg, P. Elias, M. Weitzman, and R. M. Linden.2001. Rep-dependent initiation of adeno-associated virus type 2 DNA replication by a herpes simplex virus type 1 replication complex in a reconstituted
system. J. Virol.75:10250–10258.
50. Webb, C. D., A. Teleman, S. Gordon, A. Straight, A. Belmont, D. C. Lin, A. D. Grossman, A. Wright, and R. Losick.1997. Bipolar localization of the rep-lication origin regions of chromosomes in vegetative and sporulating cells of
B. subtilis. Cell88:667–674.
51. Weindler, F. W., and R. Heilbronn.1991. A subset of herpes simplex virus replication genes provides helper functions for productive adeno-associated
virus replication. J. Virol.65:2476–2483.
52. Weitzman, M. D., K. J. Fisher, and J. M. Wilson.1996. Recruitment of wild-type and recombinant adeno-associated virus into adenovirus
replica-tion centers. J. Virol.70:1845–1854.
53. Wilkinson, G. W., C. Kelly, J. H. Sinclair, and C. Rickards.1998. Disruption of PML-associated nuclear bodies mediated by the human cytomegalovirus
major immediate early gene product. J. Gen. Virol.79:1233–1245.
54. Wistuba, A., A. Kern, S. Weger, D. Grimm, and J. A. Kleinschmidt.1997. Subcellular compartmentalization of adeno-associated virus type 2 assembly.
J. Virol.71:1341–1352.
![FIG. 1. (A) The principle of visualizing AAV DNA replication.Plasmid pAAVlacO (or rAAV [rAAVlacO]) contains the AAV ITR-](https://thumb-us.123doks.com/thumbv2/123dok_us/246677.58604/3.603.311.531.71.225/fig-principle-visualizing-replication-plasmid-paavlaco-raavlaco-contains.webp)