JOURNAL OF VIROLOGY, Sept. 1988,p. 3109-3119 0022-538X/88/093109-11$02.00/0
Copyright ©1988, AmericanSocietyforMicrobiology
Expression and Complex Formation of Simian Virus
40 Large T
Antigen and Mouse p53 in Insect Cells
DAVID,R. O'REILLY ANDLOIS K. MILLER*Departments ofEntomology and Genetics, UniversityofGeorgia, Athens, Georgia30602 Received1March1988/Accepted 16May 1988
Recombinantbaculoviruses wereconstructedwhich expresssimian virus40 largeT antigen (SVT-Ag)or
murinep53tohigh levels in infected insect cells. Characterization of the expressed proteins revealed that they displaymanyproperties of the corresponding mammalian-derived proteins. Both proteinsareof wild-type size,
localize to the nucleus, are recognized by several SVT-Ag- orp53-specific monoclonal antibodies, and are
phosphorylated in this system. Complexes are formed between baculovirus-derived SVT-Ag and p53 after
coinfection of insect cells with both recombinant viruses. After infection of insect cells with either virus individually, each proteincanself-associatetoforma varietyof oligomeric species. Pulse-chase experiments
indicated that both SVT-Ag and p53arehighly stableininsectcells, evenin the absenceof complex formation.
A variety ofgene expression systems have been devel-oped recently withaviewtoachieving enhancedexpression of eucaryotic proteins normally present at extremely low levels(44). A recently developedvectorsystemwith consid-erable potential employs the baculovirus Autographa cali-fornica nuclear polyhedrosis virus (AcMNPV) to mediate expression of the clonedgenein insectcell cultures(25, 31, 32). In this system, the gene is inserted in place of the AcMNPV polyhedrin gene, which is nonessential for viral replication in cell culture. Expression of the cloned geneis then underthe control of thepolyhedrinpromoter,resulting inhigh levels of expression late in infection.
To date, a wide range ofgenes has been expressed by using the baculovirussystemand inmanycases,biologically
activp
proteins have been obtained (32a; reviewedin refer-ence 25). Characterization of the expressed proteins has shown that insect cells can carry out at least some of the posttranslational modifications which occur in mammalian cells. Signal sequence cleavage has been demonstrated for human alpha- and beta-interferon and interleukin-2 and -3 (27, 33, 54, 55), while the appropriateproteolytic cleavage of human immunodeficiency virus env and gag proteins andinfluenza virus hemagglutinin is observed (16, 22, 26, 40). Polyomavirus large T antigen and the Drosophila Kruppel protein display DNA-binding activity after synthesis in in-sect cells (36, 42). Oligomerization, complex formation, or both has beenobserved forbaculovirus-expressed rotavirus majorcapsid antigen (7) andtwoinfluenzaviruspolymerase complex proteins (56).
It isnot yet clear whether allposttranslational modifica-tions will be comparable in mammalian and insect cells. Glycosylation is one modification which appears to differ betweenmammalian andinsectcells, but the significance of this difference is not yet known and several biologically active glycoproteins have already been produced by using the baculovirus system. One of the posttranslational modi-ficationsofkey importance isphosphorylation, because this modification has been found to play a critical role in the
regulation of protein function in a variety ofsystems. It is known that the Drosophila Kruppel protein, c-myc, and
humanT-cell leukemia virustypeIp40" arephosphorylated
after expression in insect cells, but noinformation is avail-*Correspondingauthor.
ableconcerning the natureorsite(s) of the phosphorylation
events involved (17, 34, 36). Baculovirus-derived human
epidermal growth factor receptor displays an autophos-phorylation activity like that of the wild-type protein (12).
We chose to study the expression of simian virus 40 (SV40) large T antigen(SVT-Ag) and murine p53, because prior characterization of the numerousfunctions and post-translational modifications associated with these proteins has been extensive. SVT-Ag displays ATPase activity, DNA-binding activity, and helicase activity (4, 5, 43, 57). Theprotein is essential for viral DNA replication (59) and mayassociate with thehost cell DNApolymerase alpha (53). It can immortalize and transform primary and established celllines, and these propertiesmaybe relatedtoitsabilityto bind and stabilize the host cellprotein p53 (30, 38,reviewed in reference37). p53 itself is animmortalizingoncogene (6, 20, 39) and isimplicatedinthepassageof cellsfrom
Go
toG1 (21). Both SVT-Ag and p53 are phosphoproteins, and thenature and sites ofphosphorylation are largely determined
(29, 45-49, 60). SVT-Ag isaparticularly good modeltostudy phosphorylation since there is muchrecentevidence linking phosphorylationtothe regulation ofanumberof its biolog-ical activities, especially viral DNA replication and DNA binding (11, 35, 48).
Inthisstudy, we describe theexpression of SVT-Ag and
p53 by using the baculovirus expression system.The abun-dantly expressed proteins are localized in the insect cell
nucleus and adopt astructuralconformation similarto that oftheir mammalian counterparts. Both proteins are phos-phorylated and retain the ability to associate together after expression in insectcells.
MATERIALS AND METHODS
Cells and viruses. Spodoptera frugiperda(fall armyworm) IPLB-SF21 cells (SF21 cells)(61)weremaintained in TC-100 medium(GIBCO Laboratories) supplemented with10%fetal calf serum and 0.25% tryptose broth. Stocks ofwild-type (wt) virus, AcMNPV L-1 (24),wereprepared and assayedas
describedpreviously (31). Extractsof clone 6ratcells (30), which express elevated levels of mouse p53, and AdS
SVR111-infected human 293 cells (10) containing SVT-Ag were provided by C. Prives.
Construction ofrecombinant viruses. To generate
recom-binant AcMNPVs expressing SVT-Ag and mouse p53, 3109
Vol. 62, No. 9
on November 10, 2019 by guest
http://jvi.asm.org/
3110 O'REILLY AND MILLER
A
Bam HI EcoRI
1781 CIan B
3 ~~KpnI
ST-Ag
5190 pEV55SVT
(BgIU/StuI) 9.9 kb
p
Polyhedrin5'leoder SVT-Ag5leader
*(Bgl/StuD) I0
AAACCTATAAATAG ATC C CTAG G CTTTT
1t
(BgIll/Kpn
Pst I
Eco RI
XhoI pEV55p53 to
BgIll3 7.8kb
-'p~~~~~~~~~~~~~~~~~~~~~
Polyhedrin5'leoder p53 5 leader
-0 BgIll Eco RI -AAACCTATAAATAGATCTCGAGAATTCCATCCTGG
XhoI
_-29y -105
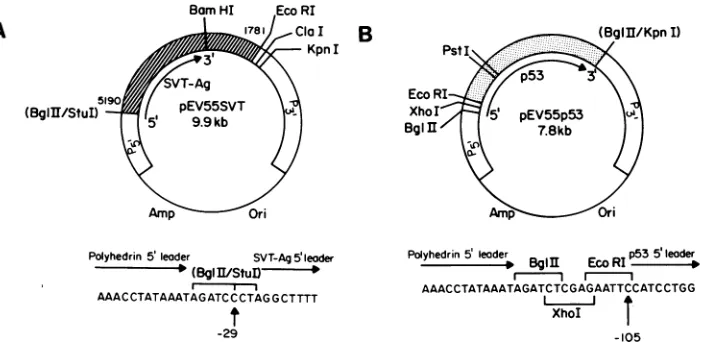
FIG. 1. Structure of the transplacement plasmids pEV55SVTand pEV55p53 (notto scale). (A) InpEV55SVT, SV40 sequences from pSVT#5, including the cDNA copyof the SVT-Ag gene, areindicated by the hatched box. The arrow indicates the expected SVT-Ag transcript. SV40 nucleotide numbersaregiven. (B) ForpEV55p53,thestippled box represents thep53cDNAfrompSV53c.Again, thep53 transcript is indicated. In both plasmids, AcMNPV sequences which flank the polyhedrin genearepresented as openboxes(P5'and P3'). The pUC8 sequences are indicated byathin line. Selected restriction endonuclease sitesareshown.Sitesinparenthesesarethose present in the originalfragments whichweredestroyedduring the construction. ThesequenceofthepEV55-cDNAjunction is given below eachplasmid. Ineach case,the position of the fusion site relative to theAUG of the cloned gene is indicated.
cDNA copies of these genes were cloned first into the transplacementplasmidpEV55. Thestructureofthis vector hasbeen described previously(31,32).The cDNAencoding SVT-Agwasexcised from the plasmidpSVT#5(constructed byY.Gluzman) by digestion withStuI(nucleotide5191) and EcoRI (nucleotide 1780), yieldingafragment whichextends from 29 base pairs upstream of the ATG of SVT-Ag to approximately a kilobase downstream ofthe translational termination codon. This fragment was cloned into pEV55 whichhadbeen previously digestedwithBglII (filledinwith T4DNApolymerase) andEcoRI. Therecombinant plasmid obtained,pEV55SVT, is illustrated in Fig. 1A.
To construct pEV55p53, a cDNA encoding murine p53 wasexcisedfromtheplasmidpSV53c(19)by digestionwith EcoRI andBglII (filled inwith T4 DNApolymerase). The resultant 1.33-kilobasefragment includes105basepairs of5' flanking sequence and extends 20 base pairs beyond the termination codon. Itwasinserted into pEV55thathad been previously digested with KpnI (bluntendedwithin T4 DNA polymerase) and EcoRI. The structure of the resultant plasmid, pEV55p53, is depicted inFig. 1B.
Toconfirm the structureofthepEV55-cDNAjunctionin both pEV55SVT andpEV55p53, the DNA sequence span-ning the junction was determined by double-stranded se-quencing of plasmid DNA, essentiallyasdescribedbyChen andSeeburg(3). The primerusedwas a17-mer correspond-ing to residues -26 to -42 in the polyhedrin leader se-quence.TheplasmidspEV55SVT andpEV55p53 containthe SVT-AG and p53 genes downstream from the AcMNPV polyhedrin promoter and flanked by polyhedrin 5' and 3' flanking sequences (Fig. 1).
Recombinant virusesexpressing these genes were gener-atedby replacement of thewtpolyhedringenein AcMNPV L-1 with the promoter-gene fusions from
pEV55SVT
and pEV55p53. To this end, 2 x106
SF21 cells were cotrans-fected with 2 ,ug of viral DNA (isolated as described in reference 31) and 18 ,ug of eitherpEVS5SVTorpEV55p53, according to the procedure of Potter and Miller (41). At 5 days later, progeny virus were harvested and then re-plaqued. Plaquesgenerated byrecombinantviruswere iden-tified by visual screening for an occlusion-negativepheno-type (31). The recombinant viruseswere subjectedtothree roundsofplaquepurification before large-scale virus stocks wereprepared. Viral DNA wasisolated, and the structures of the resultant viruses, vEV55SVT and vEV55p53, were verified by restriction enzyme analysis and Southern blot-ting.
Analysisof proteins synthesizedininfected cells.SF21 cells (106/35-mmPetridish)wereinfected withwtorrecombinant viruses at amultiplicity ofinfection (MOI) offrom 10 to50 (see figure legends). At the appropriate times postinfection (p.i.), the mediumwas removed and replaced with TC-100 lacking either methionine or phosphate. The cells were labeled 1 h later with 50,uCi of[35S]methionineor100,uCiof 32p; (New England Nuclear) in 0.5 ml of methionine- or phosphate-deficient medium. The lengths of the labeling periods are indicated in the individual figure legends. In certainexperiments,the [35S]methionine pulse labelingwas chased by incubation of the cells in TC-100 containing an excessofunlabeledmethionine. Thecellswererinsedthree times in cold phosphate-buffered saline (PBS; 8 mM Na2HPO4, 137 mMNaCl,0.5 mMMgCl2, 1.6 mMKH2PO4, 2.7 mMKCl [pH 8.0])andincubated in50 ,ul oflysisbuffer (1%NonidetP-40,150 mMNaCl,50 mMTrishydrochloride [pH 8.0]) containing 1 mM phenylmethylsulfonyl fluoride (Fluka), 40 ,uM pepstatin (Fluka), and 20 ,uM leupeptin (Fluka) for 30 min at 4°C. Lysates were stored at -80°C. Totalproteinspresentwerevisualizedbyelectrophoresisof portionsofthelysates through 10% sodiumdodecyl sulfate-polyacrylamide gels (SDS-PAGE; 23). Alternatively, sam-ples were immunoprecipitated with various antibodies (in tissue culturefluid)directedagainstSVT-Agorp53 (detailed in the text). Trial immunoprecipitation experiments were conducted to ensure the presence ofexcess
antibody.
Im-munoprecipitation experiments were carried out in NET buffer(140mMNaCl,5mMEDTA,0.05%NonidetP-40,50 mMTrishydrochloride [pH 8.0])containing 1mgofbovine serumalbumin perml,at4°Cfor 3to4h. Antigen-antibody complexes were collected by adsorptionto fixed Staphylo-coccus aureus (SigmaChemicalCo.) for 1h at4°C. Immu-noprecipitates were washed three times in NET buffer,J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.136.488.71.242.2]BACULOVIRUS-DERIVED SVT-Ag AND p53 3111 eluted by being boiled for5min inthegel-loadingbuffer (23),
andanalyzed by SDS-PAGE.
Immunofluorescence studies. SF21 cells
(105)
were seeded onto glass coverslips (22 by 22 mm) and infected with the recombinantviruses at an MOI of 10. At the selected times p.i.,the cells were washed three times in cold PBS and fixed in 70% acetone-30% methanol at -20°C for 10 min. The fixativewas removed, and thecoverslips wereair driedand stored at -20°C. Before being stained, coverslips were incubated inPBS at room temperaturefor15min. ThePBS was removed by aspiration, and 50 ,ul of the appropriate dilution (determined empirically) of monoclonal antibody was placed on the coverslip. After incubation at 37°Cfor1 h in a moist environment, the coverslips were washed twice for 15 mineach in PBS at room temperature. The second antibody used was a fluorescein isothiocyanate-conjugated rabbit anti-mouse immunoglobulin antiserum (Sigma), and incubation conditions were as described above. After two further washes withPBS,thecoverslipsweremountedonto microscope slides and examined by using Nomarski orUV optics.Sucrose gradient centrifugation. SF21 cells were infected with the recombinant viruses individually or together and lysedasdescribed above. Lysates(2 x 106 infected cellsper gradient) wereloadedonto5-ml lineargradients of5 to 20% sucroseinPBS andcentrifugedat55,000 x gfor16 hat
4°C.
Equal volume fractions (250 ,ul) were collected and immu-noprecipitated with PAb 419 orPAb 421.
RESULTS
Construction of recombinant viruses and analysisof SVT-Ag and mouse p53expression. cDNAsencoding SVT-Agand murine
p53
were cloned into thetransplacement
plasmid
pEV55asdescribed inMaterials andMethods.The resultant plasmidsaredepictedinFig. 1. In
pEV55SVT
(Fig. 1A),
the junction ofpolyhedrinandSVT-Agleader sequencesoccurs at position -29 relative to theSVT-Ag
ATG,
while inpEVSSpS3
(Fig. 1B),thepolyhedrin-p53
fusion isatposition
-105 relative tothe p53 ATG. The transplacement
plasmid
pEV55 provides the entire
polyhedrin promoter-leader
re-gion to drive geneexpression.
The A of theBglII
site in pEV55 corresponds to the A of thepolyhedrin
ATG in wt AcMNPV. pEV55 is thereforeexpected
toprovide higher
levels of
expression
than vectors such aspAc373
orpEV51
which lack portions of the leader
region
(31, 54). The recombinant viruses vEVSSSVT andvEVSSpS3
were con-structed by cotransfection of wt AcMNPV DNA with pEVSSSVT and pEVSSpS3respectively,
to allow allelic replacement ofthe wtpolyhedrinsequenceswiththecloned gene.Recombinant viruseswereselectedby
screening
foranocclusion-negative phenotype
since thepolyhedrin
gene is nolongerpresent.Theexpression of
SVT-Ag
andp53 by
these viruses wasexaminedby SDS-PAGE
analysis
ofextracts derived from SF21 cells infected with wt or recombinant viruses for various times. The extensive accumulation ofpolyhedrin
after infection with wt virus can be clearly seen in the Coomassie blue-stained
gel
presented inFig.
2A. After infection withvEV55pS3,
highlevelsofa53-kilodalton(kDa)
protein accumulate by 36 and 48 h
p.i.
Thisprotein
is not present in wt-infectedorinmock-infectedcells.Similarly,
a novel 94-kDa protein accumulates by 36 and 48 h after infectionwithvEVSSSVT,
although the steady-state levels of thisproteinaresomewhat lower than those of the 53-kDaprotein.
The kinetics of
synthesis
of theseproteins
wereexaminedby
SDS-PAGE of[35S]methionine pulse-labeled
proteins
of infected cells(Fig.
2B).
Synthesis
of boththe 94- and53-kDaproteins
is detectable at24hp.i.
andincreasesthrough
48 h after infection. In both cases, the kinetics ofsynthesis
arelikethose of
polyhedrin,
andasimilarinhibition ofhost cellprotein
synthesis
is observedatlate timesp.i.
Toconfirmthatthe 94- and 53-kDa
proteins correspond
toSVT-Ag
andp53, respectively, immunoprecipitation
exper-imentswereperformed
withmonoclonal antibodiesspecific
forthese
proteins.
Itcanbe seeninFig.
2C and D that the monoclonalantibody
PAb419,
whichisspecific
forSVT-Ag
(15),
recognizes
the 94-kDapolypeptide expressed
in vEVSSSVT-infected SF21 cells.Similarly,
PAb421,
ananti-p53 antibody
(15),
specifically
immunoprecipitates
the 53-kDaprotein
fromvEV55-infected
cells(Fig.
2C andD).
Again,
nosuchproteins
areimmunoprecipitated
fromwt-ormock-infected
lysates.
Note thatpolyhedrin
precipitates
spontaneously
in theseexperiments
because it isquite
insoluble in the
lysis
buffer used. Asbefore,
synthesis
of bothproteins
is detectable from 24 h after infection and lower levels ofSVT-Ag
areobserved than ofp53.
In these and certainsubsequent
experiments,
the amount ofsample
usedfrom
vEVSSpS3-infected
cellswasreduced becauseof the more efficientexpression
of this vector(see
figure
legends).
Trial titrationexperiments
werepreviously
carriedoutto ensure the presence ofexcess
antibody
inallimmu-noprecipitations.
Wenext carriedout
immunoprecipitations
ofhumancells infected withAd5SVR111,
arecombinant adenovirus which expressesSVT-Ag,
or clone 6 ratcells,
which express elevated levels of mousep53,
in order to compare the mammalian-derivedproteins
withthesameproteins
synthe-sized in infected insect cells.
Figure
2E demonstratesthatSVT-Ag
synthesized
in insect cells isprecisely
thesamesizeas
SVT-Ag
produced
inAdSSVR111 infected humancells.Similarly,
theinsect-derivedp53
displays
anidenticalmobil-ity
to murinep53
made in the cloned 6 cells. Note that in certainexperiments
theinsect-derivedSVT-Ag
is observedas a doublet. This is also seen with
Ad5SVR111-infected
human cellsand mayreflect
postlysis degradation
which has beenfrequently
observed for variousSVT-Ag
preparations
(58).
From the Coomassie blue-stainedgel
ofthesamples
shownin
Fig. 2E,
we estimated thatthe recombinant bacu-lovirusesproduced
from2-to10-fold moreSVT-Ag
andp53
percellthantheir mammaliancounterparts. Onthebasis of
a
comparison
with Coomassie blue-stainedprotein
stan-dards,
we estimate thatSVT-Ag
accumulated toapproxi-mately
25 to50,ug/107 cells,
whereasp53
levelsof60to150 ,ug/107cells are obtained(datanotshown).Subcellular localization of
SVT-Ag
andp53
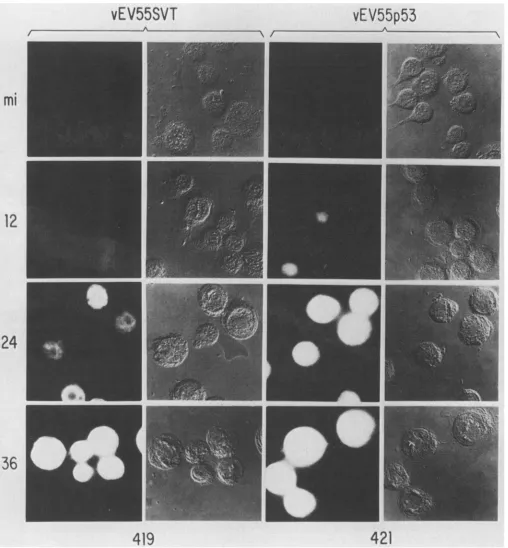
ininsect cells. A series of immunofluorescenceexperiments
were carriedout to examinethesubcellular localizationofSVT-Ag
orp53
in insectcellsatvarious times afterinfection withvEV55SVT
or
vEV55p53,
respectively.
Inthe datapresented
inFig.
3,
SVT-Ag
expression
isinitially
observedat24 hp.i.,
whereas low levels ofp53
are detectableat12 hafter infection. The moreextensiveaccumulation ofp53
is inagreementwith theexpression
studies described above. At 12 hp.i.
whenexpression
levelsarelower,
bothproteins
arepredominantly
found in the nucleus. Later in
infection,
duetothehigh
levelexpression
of theseproteins
andtothefact that the nucleus swellsconsiderably
afterbaculovirusinfection,
it is difficultto establish whether there are
significant
amounts of theseproteins
in thecytoplasm
as well. Thepatterns of accumu-VOL.62, 1988on November 10, 2019 by guest
http://jvi.asm.org/
3112 O'REILLY AND MILLER
I:4b?S- 4e'3EfI.-RICQUI. i4
t-,
j.J ,...
Ir-,
w-., %,:
*1
_,,,ii_W.:8!-s.th;'
& ,; $z..,."..._....:...
i #. " t
.,,.|u.
_ 1 ""_ "
_
*.
Z
_
S - _ S
#s _ _
X
F- _
F
*
F
4 ' F
-*:: * !
S -E
1t*-i.;
.*::*gg.:.g....g<:.
*.:;0p..::
::: st
^
w F :
> .,i
..>wds
.. ^
..:.""
< -. b ...l
..
E
... %, E
Z
.:...
S
.F
.... ,=\X s
ffi
|
Si i
.. > o }
.-- £.i.
.,s: ,., ... *....::
_W,*WS..,. -.
9 ,
ti-FIG. 2. Proteinsynthesis in recombinantand wtAcMNPV-infected insect cells. SF21 cells were infected with vEV55SVT,vEV55p53, or wtAcMNPV at an MOIof 20. At 12, 24, 36, and 48 h p.i., cells were pulse labeled with[35S]methioninefor1h and thenlysed (50,ul/35-mm dish). (A and B)A20-and 3-,ul sample, respectively, of each lysate was analyzed directly by SDS-PAGE.ACoomassieblue-stained gel is shown inpanel A, andanautoradiograph is shown inpanelB.(C)A20-,ulsample of eachvEV55SVTlysate and 7
RI1
of eachvEV55p53lysate were immunoprecipitated with PAb 419 and PAb 421, respectively, before SDS-PAGE. The same respective volumes of wt-infected and mock-infected (mi) cells were analyzedascontrols. TheCoomassie blue-stained gel is shown. (D) The same as panel C except that 10,u1of thevEV55SVT lysates and 3,u1
of eachvEV55p53 lysate were used for theimmunoprecipitation experiments. The gel was visualized by autoradiography. (E) Immunoprecipitates of vEV55SVT- or vEV55p53-infected lysates (48 h p.i.) were analyzed in parallel with immunoprecipitates of lysates ofAd5SVR111-infected293 cells or clone6 ratcells.Ineach case, the volume oflysate used for the mammalian samples representsthreetimes more cellsthan thequantity used for the insect cell samples. Mock-infected (mi) insect cells were included asnegative controls.Anautoradiographisshown. Themolecular sizemarkers (M)aregiven in kilodaltons, and the positions of SVT-Ag (T), p53,andpolyhedrin (PH) areindicated.lation observed were unchanged after coinfection of SF21 cells with both viruses (data not shown).
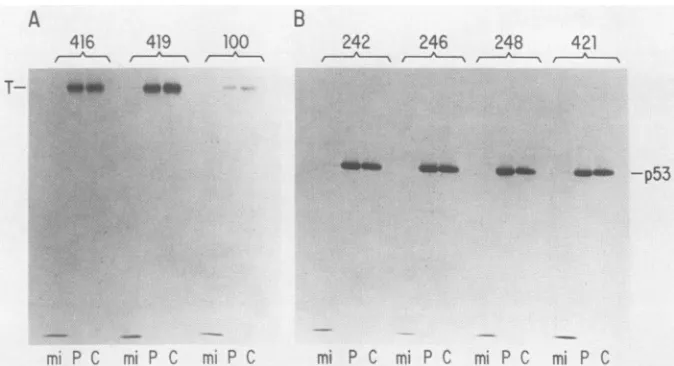
Epitope analysis of insect-derived SVT-Ag and p53. The data obtained in the experiments described above indicate that theinsect-derived SVT-Ag andp53eachdisplay at least one epitope known to be found on the corresponding wt proteins.Toinvestigate further thesimilarity of these recom-binantproteinstotheirmammalian counterparts, we exam-ined the ability of several other monoclonal antibodies to
recognizethem. In thisexperiment, infected SF21 cellswere pulselabeled with [35S]methionine48 hp.i.andlysedeither immediately or after a 3-h chase. The lysates were then immunoprecipitated with the appropriate antibodies and analyzed bySDS-PAGE. Theantibodies tested forSVT-Ag were PAb 416 and PAb 419, which recognize distinct epi-topestoward the N terminus ofSVT-Ag (15), andPAb 100, which recognizes a determinant present in the center ofa subsetofSVT-Agmolecules(13, 14, 50).The latterepitope, J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
BACULOVIRUS-DERIVED SVT-Ag AND p53 3113
vEV55SVT
vEV55p53
I
[image:5.612.56.564.71.619.2]419
421
FIG. 3. Immunofluorescence localization of SVT-Ag and p53 in insect cells. SF21 cellsonglass coverslipswereinfected with vEV55SVT
or vEV55p53 at an MOI of 10. Infections were allowed to proceed for 12, 24, or 36 h before the cells were fixed and processed for
immunofluorescenceanalysis. The primary antibodies usedwerePAb 419orPAb 421.Mock-infected cells (mi)wereprocessed in parallelas
controls. Thefixed and stained cellswere visualized by either UV (columns 1 and 3)orNomarski(columns 2 and 4) illumination.
which has been associated with theDNA-binding properties of SVT-Ag, is dependent on the molecule assuming an
appropriate tertiary structure,becauseantibody recognition isdestroyed by denaturation. The baculovirus-derived SVT-Ag displays all three epitopes, as shown inFig. 4A.
The antibodies used to analyze p53 were PAb 242, PAb 246, PAb 248, and PAb 421 (15, 63). The former three are
apparently specificformousep53while the PAb 421epitope is morehighlyconserved and is found onp53 molecules of several species (62). The PAb 242, PAb 246, and PAb 248
mi
12
24
36
VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
3114 O'REILLY AND MILLER
A
4}6
479
1QO
--Ps
.:.::.
Y
i...
...
...
*::::::::: ..
.:...
...:.:....:...
.:... .:
....
*.::.::::.:
:. :: [image:6.612.136.473.75.258.2]...
...::
.. :::.. :.:::.*...;0..B!iS.:S-''...
..Si...
.:::
B
242 246 248 421
__
__b
_ __ -p53mS P C MiC m Dr mi PC mi C mi P mi P C
FIG. 4. Epitope analysis ofinsect-derivedSVT-Ag(T)andp53.SF21cellswereinfectedwithvEVSSSVT(A) orvEVSSpS3(B) at anMOI of 20. At36 hp.i., the cellswere labeled with [35S]methioninefor 1 h.The cells wereeitherlysedimmediately (lanes P) orincubatedin completeTC-100 containinganexcessof cold methionine for3 hbeforelysis(lanesC). Then,10 Il of eachlysatewasimmunoprecipitated
withPAb 416, PAb 419, or PAb 100 (panel A) orwithPAb242, PAb 246, PAb 248, or PAb 421 (panel B).Mock-infected(mi)cell lysateswere
immunoprecipitated with each antibody as a control (lanes mi). Immunoprecipitated proteins were analyzed by SDS-PAGE, and the autoradiographs obtainedareshown.
epitopesarelocalizedtoward the N terminusofthemolecule (exons 1, 3, and 4, respectively), whereas the PAb 421 epitope isatthe extremeCterminus ofp53 (62). PAb 246 is of furtherinterest since it has been reported that the deter-minant recognizedbythisantibodyisstabilizedbycomplex formation with SVT-Ag. However, itcanbe seenfrom Fig. 4B thatthebaculovirus-produced p53 displays all four epi-topes, regardless of the presence of SVT-Ag. For both SVT-Agand p53, no significant difference was observed in theability ofanyantibodytorecognize these proteinsafter eitherapulseor apulse-chaselabeling (compare lanes P and C). This result indicates that all epitopes examined are present onboth recentlysynthesized andoldermolecules.
Complex formation between insect-derived SVT-Ag and p53. One of the most characteristic properties of SVT-Ag and p53 in mammalian systems is the ability to associate togethertoformatightcomplex(reviewedinreference37). Todetermine whetherthis associationtakes place in insect cells, we examined the sedimentation profiles of SVT-Ag andp53 through linear sucrosegradients. 35S-labeled lysates were prepared 48 h after infection of SF21 cells with vEV55SVT and vEV55p53 either individually or together, and thelysates werecentrifuged through sucrosegradients. After centrifugation, the gradients were fractionated and immunoprecipitated withPAb 419(anti-SVT-Ag)orPAb 421 (anti-p53). Figure 5A shows the sedimentation profile of SVT-Agextracted from SF21 cells infected withvEVSSSVT alone. While the protein sediments ratherheterogeneously, twomajor species can be distinguished (fractions 3 to 5 and fractions8 to10).This is similar to the sedimentation profile observedforSVT-Agextractedfrom SV40-infectedmonkey cells in which it has been proposed that the slower-sedi-menting form represents monomers and dimers, while the faster-sedimentingform corresponds to tetramers and higher oligomericforms (8). Thus, these data suggest that SVT-Ag synthesized in insect cells is also capable of forming a varietyofoligomeric forms.
Thesedimentationprofile of p53 extracted from SF21 cells infectedwith vEV55p53 aloneindicates that this protein also exists inavariety of oligomericforms (Fig. 5B). Again, two
species can be discerned(fractions 4 to 6 and fractions 14 to 17), although p53 appearsto sediment even more heteroge-neously than SVT-Ag.
After coinfection of SF21 cells with vEV55SVT and vEV55p53, immunoprecipitation of the gradient fractions with the anti-SVT-Ag antibody shows that much of the SVT-Agnow exists as heavy oligomeric forms which sedi-ment tothebottom ofthegradient(Fig. 5C, fractions 13to 17). p53 is nowfound tobecoprecipitated by the anti-SVT-Agantibody from the heavier fractions. These data indicate that complex formation has taken place between SVT-Ag and p53, and that the complexed forms of these proteins cosediment through the sucrosegradients,as seen in SV40-infected rodent cells (28).
Conversely, when the same gradient fractions are immu-noprecipitated with the anti-p53 antibody (Fig. 5D), p53 is found to sediment throughout the gradient and complexed SVT-Ag, which is coprecipitated with pS3 by the anti-p53 antibody, cosediments toward the bottom of the gradient.
Stability of baculovirus-produced SVT-Ag and p53. The results described above demonstrate that baculovirus-pro-ducedSVT-Ag and pS3arecapable ofassociatingtogether to formahigh-molecular-weightcomplex in insect cells. One of the consequencesattributed tocomplex formation in mam-malian cells is the stabilization ofp53, which is otherwise turned over very rapidly (38). We undertook a series of pulse-chase experiments to determine the stability of SVT-Ag and p53 in SF21 cells both with and without complex formation. SF21 cells were infected with vEVSSSVT and vEVSSpS3 either separately or together. At 36 h p.i., the cellswerepulselabeled with[35S]methioninefor 30 min and thenchased with an excessof cold methionine for selected times. The autoradiograph presented in Fig. 6 shows that bothSVT-Agand pS3are highly stable ininsectcells, with nosignificantturnoverobservedevenaftera25-h chase. No significant differences were observed when the SF21 cells werecoinfected with both viruses. The fact that coprecipi-tation of SVT-Ag and p53 was observed even after the 30-min pulse suggests that complex formation takes place rapidly in this system.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
BACULOVIRUS-DERIVED SVT-Ag AND p53 3115
A
vEV55SVT
U 171615 14 1312 1110 9 8 7 6 5 4 3 2 l
B
vEV55p53
U 17161514131211 109 8 7 6 5 4 3 2 1
*lo - W -,A
-..w.ww..MPM--o
ES~nhlIPsE-W'
.
419
-
p53
421
vEV55SVT+ vEV55p53
U 171615141312 1 109 8 7 6 5 4 3 2 1
UINI--KIMUe'M
-1--ShAmkm
-D
vEV55SVT+ vEV55p53
U 17 16 15 1453 4112 10 9 8 7 6 5 4 3 2 1
p53- _ .:::::C-w.
419
-p53
421
FIG. 5. Sucrose gradient analysis of insect-derived SVT-Ag (T) and p53. SF21 cells were infected with vEV55SVT (MOI, 50) (A), vEV55p53(MOI, 10) (B),orcoinfected with both viruses(MOIs,50 and10, respectively) (CandD)for 36 h.Theywerethenpulselabeled
with [35S]methionine for 1 h before lysis. Lysates were centrifuged through linear 5 to 20% sucrose gradients, and the fractions were
immunoprecipitatedwith PAb419orPAb 421. ImmunoprecipitateswereanalyzedbySDS-PAGEandautoradiographedasdescribedinthe
legendtoFig.4.Fractionsarenumbered1through17 from thetopof thegradient.LaneU,Analiquotof thelysatenotsubjectedtogradient
centrifugationandthenimmunoprecipitated inparallel.
vEV55SVT P 1 2.5 5 10 25
vEV55SVT vEV55p53
P 1 2.5 5 10 25
vEV55p53 P 1 2.5 5 10 25
vEV55SVT
vEV55p53
P 1 2.5 5 10 25
x
b
-T_
-p53.,1 \.. /
419 421
FIG. 6. StabilityofSVT-Ag(T)and p53 in insect cells. SF21 cellswerepulselabeled with[35S]methioninefor 30minafter infection for 36hwithvEV55SVT, vEVSSpS3,orboth(MOIsasin thelegendtoFig. 5).Labeledcellswereeitherlysedimmediatelyafterthepulse(lanes
P)orincubated inTC-100 withexcesscold methionine for1,2.5, 5, 10,or25 h.Lysateswerethenimmunoprecipitatedandanalyzedasbefore.
C
4mwwdw..l---- .Am.
VOL. 62,1988
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.56.564.75.357.2] [image:7.612.137.476.482.700.2]3116 O'REILLY AND MILLER
A
B
vEV55SVT
vEV55p53
wt
12 24 36 48 12 24 36 4812 24 3648 mi
-T
vEV55SVT vEV55p53 wt 12 6r4- 2
M 12 2436i4812243648}2 2436 48
205-
116- 97.4-66)
-
45-
29- 97.4-
66--p53
e sete: t; i .iP
1 ....'
Z7ii3IF
C
VEVS5SVT
vEV55p53 vEV55SVTt
vEV55p53
ml 12 24 3648mi 12 24 36 481224 36 4812 24 3648
45-
29-D
vEV55SVT 12 24 3648
i
l
.
-_L.
-:
-T
-p53
vEV55p53 vEV55SVT vEV55p53
12 24 36 48 12 2436 4812 2436 48
116-97.4-
:...:S
::: :::#.F.:7..w:**@.."a
i
..K.'.-''...'= '" - T.:...
66-
45--p53
205- 97.4- 66-
45-
29-_."..._ ...--::o
419 421 419 421 419 421 419 421
FIG. 7. Phosphorylation of SVT-Ag and p53 in insect cells. SF21 cellswereinfected withvEV55SVT(MOI, 20),vEV55p53(MOI, 20),
or wtvirus(MOI, 20),orcoinfectedwithvEV55SVT andvEV55p53(MOIs,50and10,respectively). At12, 24, 36,or48hp.i.,thecellswere
labeled with
32p,
for 1 h before lysis. Samples of the lysates were either analyzed directly by SDS-PAGE (panels A and B) or wereimmunoprecipitated with PAb 419 or PAb 421 as described in Materials and Methods(panels CandD). (AandC)Coomassieblue-stained
gels;(B and D)Autoradiographs.Thesizesof the molecular markersaregivenin kilodaltons(lane M),and thepositionsofSVT-Ag(T),p53, and polyhedrin (PH)areindicated. mi,Mock infected.
-T
-p53
PhosphorylationofSVT-Agandp53ininsect cells.Another notable characteristic ofSVT-Ag and p53 is that they are both phosphorylated in mammalian cells. We investigated whether the proteins produced by our expression system were phosphorylated by labeling SF21 cells with
32Pi
at selected times after infection withvEV55SVT, vEV55p53,or wt virus. The labeled lysates were then analyzed by SDS-PAGE; autoradiography (Fig. 7B) shows clearly that bothSVT-Ag and p53 are phosphorylated in this system. In vEV55p53-infected cells, themajorphosphoprotein present in the infected cell late in infection is p53. Immunoprecipi-tation before SDS-PAGE confirmed the identities of the phosphoproteins (Fig. 7D). Comparison of the autoradio-graphs(Fig. 7B andD) with theCoomassieblue-stainedgels (Fig. 7A andC) suggests that thedegreeofphosphorylation is maximalat24hp.i.,droppingtwo- tofivefoldby48h after J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
BACULOVIRUS-DERIVED SVT-Ag AND p53 3117
infection.
Analysis
ofSVT-Ag
andp53
extracted from SF21 cellscoinfected with bothvEV55SVT
andvEV55p53
reveals nomajor
differences in thedegree
ofphosphorylation
of either oftheseproteins
aftercomplex
formation(Fig.7C andD).
DISCUSSION
Inthis
study,
wedescribetheconstructionandanalysis
of recombinant baculovirus vectorsexpressing SVT-Ag
andmouse
p53.
The vectorsproduce
levels of theseproteins
ranging
from 50to 150,ug/107
cells. These levelsare excel-lentcompared
with those ofthepresently
available mamma-liangeneexpression
systems.Recently, Jeang
etal.(18)
describedtheconstructionofa recombinant baculoviruscontaining
thecoding
sequences forSV40
large
T and smalltantigens.
However,
thisvector includedSV40early-gene
splice
sites,
andtheauthorsreport thatonly
small tantigen
issynthesized
insignificant
amounts. There is little or no detectable accumulation of
large
Tantigen.
The fact thatwe havereadily
obtained theexpression
oflarge
Tantigen
from a cDNA clone further demonstrates theimportance
ofusing
intronlessgenes with the baculovirusexpression
system, asdiscussedpreviously
(32).
Several lines of evidence indicate that the baculovirus-derived
proteins
are similar to their mammalian counter-parts. Theproteins
are of normal size(Fig. 2E)
and aretransported
to the nucleus(Fig. 3), indicating
that the nuclear transportsignals
of bothSVT-Ag
andp53
arerecognized
ininsect cells. Bothproteins
also appeartoadopt
a native conformation since
they
bothdisplay
severalepi-topescharacteristic ofthewt
proteins (Fig. 4). Interestingly,
the baculovirus-derived
p53
isclearly recognized by
the monoclonalantibody
PAb246,
even in the absence ofSVT-Ag (Fig.
4B).
Yewdell et al.(63) reported
that thisepitope
isgenerally
unstable in the absence ofSVT-Ag.
However,
those authors do report that the PAb 246epitope
is present on
p53
from at least onespontaneously
trans-formedmousecelllinein theabsence ofSVT-Ag.
Thebasis ofthisphenomenon
isnotknown.Sucrose
gradient centrifugation
analyses
indicated that baculovirus-derivedSVT-Ag
andp53
arecapable
ofself-associating
toformavariety
ofoligomeric
forms(Fig. 5).
At least forSVT-Ag,
thispropertyisimportant
because there is much evidenceshowing
that differentoligomeric
forms of theprotein display
differentposttranslational
modifications andbiological
functions in mammalian cells(reviewed
in reference43).
Our
experiments
also show thatbaculovirus-produced
SVT-Ag
andp53
can associatetogether
in insect cells to formarapidly sedimenting high-molecular-weight
complex.
Theprecise
roleplayed by complex
formationin mammalian cellsisnotyet clear but it may haveprofound
effectsontheability
ofSVT-Ag
to support viral DNAreplication
and immortalize or transform cells(1, 9, 30;
for areview,
see reference37).
It istherefore of interesttoobserve thatthis property is retainedby
theproteins
in insect cells. Our results suggest that someSVT-Ag
andp53
remainuncom-plexed
in this system since the total amount ofSVT-Ag
immunoprecipitated
from these cells(Fig.
SC)
is greater thantheamount
coprecipitated
withp53 (Fig. SD).
The converseis alsotrue.Since
p53
appears tobe inexcessofSVT-Ag
in coinfectedcells,
thereislikely
tobeacertainamountof freep53
inthis system. Inaddition,
thereis evidenceto suggest thatonly
asubpopulation
ofSVT-Ag
iscapable
ofbinding
p53 (63). However, it is also difficult torule out the
possi-bility that there is some dissociation of the complex postlysis.
We notice that the sedimentation profile of p53does not change significantly in the presence orabsence ofSVT-Ag. Since the sedimentation profile of uncomplexed p53 in mammalian cells has not been reported, we do not know whether this is ageneral phenomenon.
Another interesting observationconcerning complex for-mation in insect cells is that a pulse-chase analysis (Fig. 6) revealed thatcomplex formation is very rapid in these cells and that association is complete in less than 30min. This is in contrast to the situation in mammalian cells in which Carroll andGurney (2) reported that although p53 israpidly incorporatedinto thecomplex, SVT-Ag enters the complex moreslowly, requiring from 3 to6 h for maximum incorpo-ration. Those authors proposed that this phenomenon indi-cates that newly made SVT-Ag requires some posttransla-tional modification before it can complex p53. More recently, Schmeig and Simmons (51) have postulated that the kinetics ofcomplex formation depends on the ratio of SVT-Agtop53 in thecell line studied and that competition between newly synthesized and complexed SVT-Ag is the major determinant of the rate of entry ofSVT-Ag into the complex. The rapidrateofcomplex formation observed here would tend to support the latter hypothesis; in our system there isample p53which should allow prompt entry ofnewly synthesizedSVT-Ag into the complex.
Thepulse-chase experimentsalso revealed that both SVT-Agandp53arehighlystable ininsect cells. This is in striking contrasttowhat is observed in mammalian cells inwhich, in theabsence ofSVT-Ag, p53ishighlyunstable(38). Although we areunsureofthebasis for thisenhanced stability ofp53 in insect cells, it should be noted that in this system, p53 expressiontakesplace late ininfection, when the virus has already largely shut down host cell protein synthesis and disrupted hostcell metabolism.
Further evidence that thebaculovirus system can express authentic mammalianproteinswasprovidedby the observa-tion that both SVT-Ag and p53 are phosphorylated in this system.ThedatapresentedinFig. 7 suggestthat phosphor-ylation is maximalat24hp.i.anddeclinesthereafter. It will now beimportanttodetermine thetype(s)andsite(s) ofthe
phosphorylation
eventsinvolved,
since boththe natureand degree of phosphorylation appear to be critical for the correct functioning of SVT-Ag and p53. Recent evidence suggeststhatphosphorylation
of serineresidues down regu-latestheability
ofSVT-Ag
to supportviral replication (11, 35). This may ormaynot be mediatedby decreasedorigin-binding
activity (11, 35, 52). It seems also that the appear-ance of higher oligomeric forms of SVT-Ag is coincident with greater phosphorylation ofthe protein as it ages (8). Furthermore, Samadetal. (45)reportthatatleasta compo-nent ofp53 phosphorylation is dependent on SVT-Ag. We believe that these proteins therefore provide a valuable model systemtoestablish whether insect cellscanphosphor-ylation proteins
in a mannerqualitatively and quantitatively similartomammalian cells.In summary, we have successfully used the baculovirus expressionsystemtodirect theefficientsynthesisofSVT-Ag andmurinep53ininsectcells. Theseproteinsareidenticalto thecorrespondingmammalianproducts byallcriteria exam-ined, and the levels ofexpression obtained compare favor-ablywith those of mammalianexpression systemspresently available. Recentlyobtained evidence indicates that baculo-virus-derived SVT-Ag is functional in an in vitro
SV40-VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
3118 O'REILLY AND MILLER
origin-dependent replication system (C. Prives, personal communication). These facts, coupled with the ease ofuse and inherent safety of the baculovirus system (32), should makethese vectors aconvenient source ofSVT-Agandp53 for in vitro biochemical studies. We anticipate that the furtheranalysisof these recombinantproteins and in partic-ular a more detailed characterization of their state of phos-phorylationwillallow us to better evaluate thepotential and limitations of thebaculovirus expression system.
ACKNOWLEDGMENTS
We thankCarol Prives for criticalreadingofthemanuscript,for providing lysatesof clone6-andAd5SVR111-infectedcells, and for pSVT#5and monoclonal antibodies PAb 416, PAb419, PAb421,
andPAb 100. We aregratefultoMarcusFechheimer forhelp with
theimmunofluorescenceexperiments. WealsothankEvelyne May and Jean-Claude Erhart for providing pSV53c and monoclonal antibodies PAb242, PAb 246, and PAb 248.
This workwas supported inpartby Public Health Service grant A123719 from the National Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Braithwaite,A.W.,H.-W.Sturzbecher,C.Addison,C.Palmer, K. Rudge, and J. R. Jenkins. 1987. Mouse p53 inhibits SV40 origin-dependent DNA replication. Nature (London) 329:458-460.
2. Carroll, R., and E. G. Gurney. 1982. Time-dependent
matura-tionof the simian virus40largeTantigen-p53 complexstudied by usingmonoclonal antibodies.J. Virol. 44:565-573.
3. Chen,E.Y., andP. H. Seeburg. 1985. Supercoil sequencing:a
fast andsimple method forsequencing plasmidDNA. DNA 4: 165-170.
4. Clark, R.,M.Tevethia,and R.Tjian.1984.TheATPaseactivity ofSV40large Tantigen, p. 363-368. InG.Vande Woude, A. Levine, W. Topp, and J. Watson (ed.), Cancer cells, vol. 2. ColdSpring HarborLaboratory,ColdSpring Harbor, N.Y. 5. Dean, F. B., P. Bullock, Y. Murakami, C. R. Wobbe, L.
Weissbach,andJ.Hurwitz.1987. Simianvirus 40 DNA replica-tion: SV40largeTantigenunwindsDNAcontainingtheSV40 originofreplication. Proc. Natl. Acad. Sci. USA84:16-20. 6. Eliyahu, D., A. Raz, P. Gruss, D. Givol, and M.Oren. 1984.
Participationofp53cellulartumorantigenintransformation of normalembryoniccells. Nature(London)312:646-649. 7. Estes, M. K., S. E. Crawford, M.E. Penaranda, B. L.Petrie,
J. W. Burns,W.-K. Chan,B.Ericson,G. E.Smith, and M. D. Summers. 1987. Synthesisandimmunogenicityof therotavirus major capsid antigen usingabaculovirusexpressionsystem. J. Virol.61:1488-1494.
8. Fanning, E., B. Nowak, and C. Burger. 1981. Detection and characterization ofmultiple forms of simian virus 40 large T
antigen.J. Virol. 37:92-102.
9. Gannon,J.V.,and D. P. Lane.1987.p53and DNApolymerase alphacompeteforbindingtoSV40Tantigen.Nature(London) 329:456-458.
10. Gluzman, Y., H. Reichl, and D. Solnick. 1982. Helper-free
adenovirus type 5 vectors, p. 187-192. In Y. Gluzman, (ed.),
Eucaryoticviralvectors.ColdSpringHarborLaboratory,Cold Spring Harbor,N.Y.
11. Grasser, F. A., K. Mann, and G. Walter. 1987. Removal of serinephosphatesfrom simian virus40largeTantigenincreases itsabilitytostimulateDNAreplicationinvitro buthasnoeffect
onATPaseorDNAbinding.J. Virol. 61:3373-3380.
12. Greenfield, C., G.Patel,S. Clark, N.Jones,andM. D.
Water-field. 1988. Expressionof thehumanEGFreceptor withligand
stimulatable kinase activity in insect cellsusing abaculovirus
vector.EMBO J. 7:139-146.
13. Gurney, E. G., R. Harrison, andJ. Fenno. 1980. Monoclonal antibodies against simian virus 40 T antigens: evidence for distinctsubclasses oflargeTantigenandforsimilaritiesamong non-viralTantigens. J. Virol.34:752-763.
14. Gurney,E.G.,S.Tamowski,and W. Deppert.1986. Antigenic bindingsites ofmonoclonal antibodies specificforsimian virus 40largeTantigen.J. Virol.57:1168-1172.
15. Harlow, E.,L. V.Crawford,D. C.Pim,and N. M. Williamson.
1981. Monoclonal antibodiesspecificfor simian virus 40tumor
antigens.J. Virol. 39:861-869.
16. Hu,S.-L.,S.G.Kosowski, and K.F.Schaaf. 1987.Expression ofenvelopeglycoproteinsof humanimmunodeficiencyvirusby
aninsect virus vector. J. Virol.61:3617-3620.
17. Jeang, K.-T., C.-Z. Giam, M. Nerenberg, and G. Khoury. 1987.
Abundant synthesisof functional human T-cell leukemia virus
type I p40x protein ineucaryotic cells by usingabaculovirus expressionvector.J. Virol. 61:708-713.
18. Jeang, K.-T., M. Holmgren-Konig, and G. Khoury. 1987. A
baculovirusvector can expressintron-containing genes.J.
Vi-rol. 61:1761-1764.
19. Jenkins,J.,K.Rudge,P.Chumakov,andG. Currie. 1985.The
cellularoncogene p53can beactivatedby mutagenesis.Nature
(London) 317:816-818.
20. Jenkins, J., K. Rudge, and G. Currie. 1984. Cellular
immortal-ization by acDNA clone encoding the
transformation-associ-ated phosphoproteinp53. Nature(London)312:651-654.
21. Kaczmarek, L., M. Oren, and R. Baserga. 1986. Cooperation
between the p53 proteintumorantigen and platelet-poorplasma
in theinduction of cellular DNAsynthesis. Exp. Cell Res. 162:
268-272.
22. Kuroda, K., C. Hauser, R. Rott, H.-D. Klenk, and W. Doerfler.
1986.Expressionof the influenza virushaemagglutininin insect
cells byabaculovirusvectors. EMBOJ. 5:1359-1365.
23. Laemmli, U. K. 1970. Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature (London)
227:680-685.
24. Lee, H. H., and L. K. Miller. 1978. Isolation of genotypic
variants ofAutographa californica nuclear polyhedrosisvirus.
J. Virol. 27:754-767.
25. Lucknow, V. A., and M. D. Summers. 1988. Trends in the
development of baculovirus expression vectors. Bio-Tech-nology 6:47-55.
26. Madisen, L., B. Travis, S.-L. Hu, and A. F. Purchio. 1987.
Expression of the humanimmunodeficiency virus gag gene in
insect cells. Virology158:248-250.
27. Maeda,S.,T.Kawai,M.Obinata,H.Fujiwara,T.Horiuchi, Y. Saeki, Y. Sato, and M. Furusawa. 1985. Production of human alpha-interferoninsilkwormusingabaculovirusvector.Nature (London) 315:592-594.
28. McCormick, F., and E. Harlow. 1980. Association of a murine
53,000-dalton phosphoprotein with simian virus 40 large T antigenintransformed cells. J.Virol. 34:213-224.
29. Meek, D. W., and W. Eckhart. 1988.Phosphorylationof p53in
normal and simian virus 40-transformed NIH 3T3 cells. Mol.
Cell. Biol. 8:461-465.
30. Michalovitz, D.,D.Eliyahu,and M.Oren. 1986. Overproduction
ofprotein p53 contributes to simian virus 40-mediated
transfor-mation. Mol. Cell. Biol. 6:3531-3536.
31. Miller, D. W., P. Safer, and L. K. Miller. 1986. An insect
baculovirus host-vector system for high level expression of
foreigngenes, p. 277-298. In J. K. Setlow and A. Hollander, (ed.), Genetic engineering, vol. 8. Plenum Publishing Corp.,
NewYork.
32. Miller,L. K.1987.Baculoviruses forforeigngeneexpressionin
insectcells,p.457-465.InR.Rodriguez,and D.Denhardt(ed.),
Vectors:asurveyofmolecularcloningvectorsandtheiruses.
Butterworth's, Stoneham, Mass.
32a.Miller, L. K. 1988. Baculovirusesas geneexpression vectors.
Annu. Rev. Microbiol.42:177-179.
33. Miyajima,A., J.Schreurs,K.Otsu,A.Kondo, K.-I.Arai,andS.
Maeda.1987. Useof thesilkworm,Bombyxmori, andaninsect
baculovirus vectorfor high-level expression and secretion of
biologicallyactivemouse interleukin-3.Gene 58:273-281. 34. Miyamoto, C., G. E. Smith, J. Farrell-Towt, R. Chizzonite,
M.D.Summers,and G.Ju. 1985. Productionof human c-myc protein in insect cells infected with abaculovirus expression
vector. Mol. Cell. Biol.5:2860-2865.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
BACULOVIRUS-DERIVED SVT-Ag AND p53 3119
35. Mohr, I. J., B. Stillman,and Y. Gluzman. 1987. Regulation of
SV40DNAreplication by phosphorylation ofTantigen.EMBO J.6:153-160.
36. Ollo, R., and T. Maniatis. 1987. Drosophila Kruppel gene
product produced in a baculovirus expression system is a nuclearphosphoprotein that binds DNA. Proc.Natl.Acad. Sci.
USA84:5700-5704.
37. O'Reilly,D. R.1986.p53andtransformationbySV40. Biol.Cell 57:187-196.
38. Oren, M., W. Maltzman, and A. J. Levine. 1981.
Post-transla-tional regulation of the54Kcellulartumorantigen innormaland transformed cells. Mol. Cell. Biol. 1:101-110.
39. Parada, L.,H.Land, R. Weinberg, D.Wolf, and V. Rotter. 1984.
Cooperation betweengeneencoding p53tumourantigen andras in cellulartransformation. Nature (London) 312:649-651.
40. Possee, R. D. 1986. Cell-surface expression of influenza virus
haemagglutinin in insectcellsusing abaculovirusvector.Virus Res. 5:43-59.
41. Potter, K. N., and L. K. Miller. 1980. Transfection of two
inverterbrate cell lines with DNA of Autographa californica nuclearpolyhedrosis virus. J. Invertebr. Pathol.36:431-432.
42. Rice, W. C., H.E. Lorimer, C. Prives, and L. K. Miller. 1987.
Expression of polyomaviruslarge Tantigenbyusinga baculo-virusvector. J.Virol.61:1712-1716.
43. Rigby, P., and D. Lane. 1983. The structure and function of
simian virus40largeTantigen.Adv. ViralOncol.3:31-57. 44. Rodriguez,R., and D. Denhardt (ed.). 1987.Vectors:asurveyof
molecularcloningvectorsand theiruses.Butterworth's,
Stone-ham,Mass.
45. Samad, A., C. Anderson, and R. Carroll. 1986. Mapping of phosphomonoester and apparent phosphodiester bonds ofthe
oncogene product p53 from simian virus 40-transformed 3T3
cells. Proc. Natl. Acad.Sci. USA 83:897-901.
46. Scheidtmann, K. H. 1986. Phosphorylation of SV40 large T
antigen: cytoplasmic and nuclear phosphorylation sites differin their metabolic stability. Virology 150:85-95.
47. Scheidtmann,K.H., B. Echle, and G. Walter.1982.Simian virus 40largeTantigen is phosphorylatedatmultiple sites clustered
in two separateregions. J. Virol.44:116-133.
48. Scheidtmann, K. H., M. Hardung, B. Echle, and G. Walter. 1984. DNA-bindingactivity of simian virus 40 largeTantigen correlates withadistinctphosphorylation state.J. Virol. 50:1-12.
49. Scheidtmann, K. H., A. Kaiser, A. Carbone, and G. Walter. 1981. Phosphorylation of threonine in theproline-rich
carboxy-terminalregionof simianvirus40 Tantigen.J.Virol.38:59-69.
50. Scheller, A.,L.Covey,B.Barnet, and C. Prives. 1982. A small
subclass of SV40Tantigen binds to theorigin ofreplication.
Cell 29:375-383.
51. Schmeig, F.,andD. T.Simmons. 1984.Intracellular location and
kinetics ofcomplexformation between simian virus 40 Tantigen
andcellular proteinp53. J. Virol. 52:350-355.
52. Simmons,D.T.,W.Chou,and K.Rodgers. 1986. Phosphoryla-tiondownregulatestheDNA-binding activityof simianvirus40 Tantigen.J. Virol.60:888-894.
53. Smale, S. T., and R. Tjian. 1986. T-antigen-DNA polymerase
alpha complex implicatedin simian virus 40 DNAreplication.
Mol.Cell. Biol. 6:4077-4087
54. Smith,G. E.,G.Ju,B. L.Ericson, J. Moschera,H.-W. Lahm,
R. Chizzonite, and M. D. Summers. 1985. Modification and
secretion of human interleukin-2 produced ininsect cells by a
baculovirusexpression vector. Proc. Natl. Acad. Sci. USA82:
8404-8408.
55. Smith,G. E., M. D. Summers,andM.J. Fraser. 1983. Produc-tion of human beta-interferon in insect cells infected with a
baculovirusexpressionvector. Mol. Cell. Biol. 3:2156-2165.
56. St.Angelo, C., G. E. Smith, M. D. Summers,and R. M.Krug.
1987. Two of the three influenza viral polymerase proteins
expressed using baculovirus vectorsformacomplex ininsect cells. J. Virol. 61:361-365.
57. Stahl, H., P. Droge, and R. Knippers. 1986. DNA helicase
activity ofSV40 largetumorantigen. EMBOJ. 5:1939-1944.
58. Tegtmeyer, P., K. Rundell,and J.K.Collins. 1977.Modification
ofsimian virus40proteinA. J. Virol.21:647-657.
59. Tooze, J. 1981. DNA tumor viruses, p. 125-296. InJ. Tooze
(ed.), Molecularbiology oftumorviruses, 2nd ed. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
60. VanRoy, F., L. Fransen, and W. Fiers. 1981. Phosphorylation
patterns oftumorantigens in cells lytically infected or
trans-formedby simian virus40. J. Virol. 40:28 44.
61. Vaughn, J. L., R. H. Goodwin, G. L. Thompkins, and P. McCawley. 1977. The establishment of two cell lines from the insectSpodopterafrugiperda (Lepidoptera; Noctuidae). In Vi-tro13:213-217.
62. Wade-Evans,A., andJ. Jenkins.1985. Preciseepitope mapping of themurine transformation associated proteinp53. EMBO J. 4:699-706.
63. Yewdell,J. W., J. V.Gannon,andD. P. Lane.1986.Monoclonal antibody analysis of p53 expression in normal and transformed cells. J. Virol. 59:444 452.
VOL. 62, 1988