0022-538X/84/070170-11$02.00/0
Copyright © 1984, AmericanSociety forMicrobiology
Construction and
Functional
Characterization of Polyomavirus
Genomes That Separately Encode the Three Early
Proteins
ZUOYAN ZHU,'t GEERTRUIDAM. VELDMAN,'t ALISON COWIE,1t ANTHONYCARR,' BRIAN
SCHAFFHAUSEN,2 ANDROBERT KAMENlt*
Transcription Laboratory, ImperialCancerResearch Laboratories, London WC2A 3PX, England,' andDepartmentof
Biochemistry
andPharmacology,
Tufts
NewEngland
MedicalCenter, Boston,
Massachusetts
021112
Received 3January 1984/Accepted 4 April 1984Modified polyomavirusgenomesthatindividuallyencode t-helarge-and small T proteinswereconstructed by
exchanging restriction endonuclease fragments between cDNA copies of the respective mRNAs and cloned genomic DNA.The efficacies ofthenewconstructs,and that of the middle T proteingenedescribed previously
(R. Treisman,U. Novak, J.Favaloro,and R. Kamen, Nature [London] 292:595-600, 1981),were
demonstrat-ed with simian virus 40 (SV40)-polyomavirus recombinants in which partorall of theSV40 late regionwas
replaced with the modified polyomavirus early genes. Each of the three recombinant viruses induced the synthesis of only theexpectedpolyomavirus early protein ininfectedCV-1 cells. Theratesof synthesisof large, middle, and small T proteinswere ca. 1.5, 4.0, and 9.0 times the rateof synthesis ofSV40large T protein, respectively. The deletion of introns had no detrimental effect on mRNA biogenesis. Indeed, a further
polyomavirus-SV40 recombinant, containing wild-type polyomavirusearlyregion DNA, expressedanaberrant
58,000-dalton formof the middle Tproteinwhichwebelievetoresultfromutilization ofacryptic splice site.
Immunofluorescence studied withmonkey cells infected by the recombinant viruses allowedustodeterminethe cellular locationsofthe polyomavirusearly proteins.Overproduction ofthe middle T protein didnotresultina
corresponding overproduction of themiddleT protein-associated tyrosinephosphokinase activity.
The oncogenic potential of polyomavirus is contained
within the early region of the viral genome (36, 44). This
2.9-kilobase segment ofDNA encodes three related proteins,
large T, middle T, and small T, with respective molecular
weights of100,000, 48,000, and 22,000 daltons (100k, 48k,
and22k) (8, 17-19, 39, 43). Determination of individualroles
for the three polypeptides in virally induced
oncogenic
transformation would be facilitated ifone could separately
transfer into cells thegenesforthethreeproteins. This isnot
possible
with unmodified genomic DNA because the se-quencesencodingthethreeearlyproteins overlapextensive-ly. The mRNAs for the proteins share common 5' and 3'
regions (see Fig. 1A). They differ only in the size and
position ofa single intron excised by RNA splicing during
theirbiogenesis (45). Differential splicing accomplishes the
changesin translational
reading
frame necessaryto resolvethe overlapping information encoded in the
genomic
DNAsequence (8, 43). The viral genome can be modified to
separately encode the
proteins
byspecific
deletion of thecorresponding
intervening
sequence. Inthis manner,Treis-man etal. (46)constructed a
plasmid
containing
apolyoma-virusgenome encoding onlythemiddleTprotein (pPyMT1)
and used it to demonstrate that expression of the middleT
protein alonewas sufficienttotransform established cells of
the F2408 ratfibroblast lineto an oncogenic state. We now describe the derivation ofanalogous constructs
lacking
the intronsof
the large Tand small T mRNAs. Introduction of each of the three intron-minus polyomavirus earlyregion
variants into the late region for simian virus 40(SV40) has allowedus tocharacterize the geneproductsof the modified genesand thustoconfirm thattheyeach
uniquely
encode theanticipated polypeptide. A preliminary account of these
* Correspondingauthor.
t Present address: Institute of Hydrobiology ofthe Academia Sinica, Wuhan, People'sRepublic of China.
tPresent address:Genetics Institute, Boston, MA02115.
experiments has already appeared (31). The constructs we
describehere havebeen usedtostudytheindividualrolesof
polyomavirus early proteins in oncogenic transformation
(30-32) and have proved important in the definition oftwo
steps involved in transformation mediated by cellular and viral oncogenes (23, 35).
MATERIALS AND METHODS
Cells and viruses. All cells were grown in Dulbecco
modified Eagle medium supplemented with 5% fetal calf
serum. Mouse 3T6cells were infected with wild-type
poly-omavirus strainA2at a
multiplicity
of50 PFU percell. CV-1monkey cells were infected with a 1:10 dilution of the
recombinant SV40-polyomavirus stocksand grownfor40to
48 h at 37°C.
Construction of modified viral genomes expressing the individual early proteins. The specific procedure used to constructthe mutantlackingthe middle T
protein
intronhasalready been described (46). pPyLT1, a mutant
lacking
thelarge T protein intron, was made by
introducing
theAccI-AvaI DNA
fragment
from plasmid pE2.1 (45), whichcon-tainsa
partial
cDNA copyofthelargeT mRNAspanning
thesplice
junction,
intoapartial
AccIplusAvaIdigest
ofafull-lengthgenome clonedattheBamHI site
(plasmid
p53.A6.6)(Fig. 1A). Construction ofpPyST1, a mutant lacking the
small T protein intron, was more difficult because the
only
available partial cDNA clone (pE1.1) spanning the small T
splicejunctionlacked useful restriction sitesonthe 3' side.
Therefore,arestrictionfragment (PstItoHaeIII)from
pE1.1
was used to prime the synthesis of double-stranded DNA
with the genomic PstI fragment inserted into M13mp7
(M13PyPstI-1) as the single-stranded template. An M13
recombinant(M13PyPstI-1ST)that
lacked
the 48nucleotidescorrespondingtothe small Tproteinintron wasisolated,and
the AvaI fragment of replicative form DNA
spanning
thedeletionwasintroduced into
plasmid
p53.A6.6,
replacing
thegenomicAvaI
fragment.
170
on November 10, 2019 by guest
http://jvi.asm.org/
POLYOMAVIRUS GENOMES THAT ENCODE THREE EARLY PROTEINS 171
A-
BamnHIdBBmH
BBamHI Barm HIC.~~~~~C
t~~~~ H inc 11 t / ~~~~~~~~~~~~>~HincII
Ct
pP
3.A6.6v\
pPyLTI
0~~~~~~~0
BamHIEoR 3m 9 c
BgBI AvaI7
competeolyoavirsgeomeineaizedattAdH Bdnt HI Eco RI
ST
MT~~~~~~~~~~
~~Bgl
1 9LT
~~~~~~~~~~~~~~410
410 747
Barn HI Bar H
Hinc11 0 HincI
p
PyMT IT
pPySTI
0S()eaasp36wclkenvuirsxefmr.3eae,dm.
EcoRI Eco RI
BarnHIAIBrHI 5
747~~~~~~~~~~9
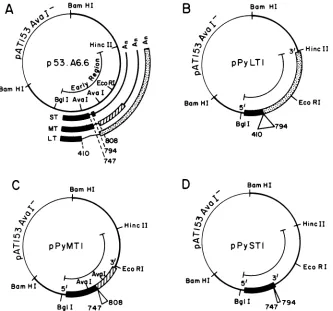
FIG. 1. Mapsof theplasmids containing polyomavirus early regionmutantslackingthe individual earlyregionintrons. (A)p53.A6.6isa
completepolyomavirusgenome linearized atthe BamHl site and cloned intopAT153Aval7 (46). The early regionmRNAs andsplice sites
(numbersrefertothenucleotidenumberingsystem of Soedaetal.,seereference43)areindicated. The threereadingframes whichareusedto
encode small T(ST),middle T(MT),andlargeT(LT)proteinsareshownby black,hatched,ordottedregions.pPyLT1 (B), pPyMT1 (C),and
pPyST1(D)arevariants ofp53.A6.6whichlack the individual introns excised from thelarge T, middleT,orsmall TmRNA,respectively.
Construction of
SV40-polyomavirus
recombinants. TheSV40-polyomavirus recombinantsused in this
study
(Fig. 2)
represent asubsetofmanyconstructs
developed during
the courseof thisproject. We present heredescriptionsofeachrecombinant plasmid mentioned in the present manuscript.
All structures wereconfirmed
by
restrictionmapping
andby
sequencingthe
junctions
between viral DNAs. Thenomen-clature we have used is either the SV40 DNA numbering
system (indicated by the
prefix "SV")
(44)orthepolyoma-virus DNAnumberingsystem(indicated bythe
prefix "Py")
(43). Allplasmidsarerepresentedin
Fig.
2andaredescribed in the counterclockwise direction starting from the top of each circle.p85.3.SP. p85.3.SP comprises a fragment containing the
complete earlyregion of SV40plus the late promoter region
fromtheBamHIsite(SV-nucleotide
[nt]
2,534) totheHpaII siteat SV-nt 348 (the HpaII site was blunted and ligated toan XhoI linker), a genomic fragment of the polyomavirus
early regionfrom Py-nt 134 (theXhoI site in pdI3035) (20) to
theEcoRI site (Py-nt 1,565), a fragment containing part of the SV40 lateregion from theEcoRI site (SV-nt 1,782) to the BamHI site (SV-nt 2,534), inserted in theBamHIsite of the vectorpAT153 (47).
pspLT5. pspLT5 consists of a fragment containing the early region of SV40 from the BamHI site (SV-nt 2,534) to
theHpaIIsiteatSV-nt 348 (convertedinto anXhoIsite with
a linker). This early region DNA derives from the SV40 deletion mutant d12005 (41) which lacks about 250
nucleo-tides located in the SV40 large T protein intron. This
construct, therefore, expresses a normal SV40 large T
protein but is deficient in expression of SV40 small T
protein. The plasmid further comprises afragment
contain-ingthepolyomavirusearly region fromtheHphlsite(Py-nt
154) to the HincII (Py-nt 2,964) but lacking the large T
protein intron(Py-nt410-794). TheHphIsite andthe HincII
site were converted into an XhoI site and a BamHI site,
respectively, by blunt end ligation with the appropriate
linkers. The vector in this construct ispXf3 (24).
pspMTI. The recombinantpspMT1comprises afragment
containing the early region ofthe SV40 mutant d12005 (41)
from theBamHI site(SV-nt, 2,534) totheHpaII site at
SV-nt348 (converted into an XhoI site), a fragment containing
part of the polyomavirus early region from Py-nt 105 (an
XhoIsitewasinsertedatthisposition)totheEcoRI site
(Py-nt 1,565)but lackingthemiddle Tprotein intron(Py-nt
747-808), afragment containingpartoftheSV40lateregion from
the EcoRIsite(SV-nt 1,782)totheBamHIsite(SV-nt2,534), and the vectorpA6 (46),which is an AvaI-negative deriva-tive ofpAT153.
pspST18. pspST18 comprises a fragment containing the
complete SV40 early region from the BamHI site (SV-nt
2,534)toSV-nt 282; the SV40major late cap site (SV-nt 325) (33) is not present in this construct. The plasmid further consists ofafragment containing part of the polyomavirus early region from Py-nt 146 to theEcoRI site (Py-nt 1,565)
but lacking the small T protein intron (Py-nt 747-794), a
VOL.51,1984
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.140.473.72.383.2][Hpa ir] -[Hph
,HI
BamHI
pspST18 a
SV-282 I
Py-146
BornHI
EcoRI ST Intron
B
280 290 300 310 320 330 340Major
Cap Site
PvuII KpnI
SV40 16S
Late mRNAE
4 I i I I r I
AGCTGGTTCTTTCCGCCTCAGAAGGTACCTAACCAAGTTCCTCTTTCAGAGGTTATTTCAGGCCATGGTGCTGC(
SV40
I .. - r . 1350
Hpa II
360
;TCACGCCA
pspMTl-t2
Xho I Linkers] p85.3.SP-{XhoI Linker ppspST18 pspLT5
fXho
I Linkers100 110 120 130 140 150 160 170 180
POLYOMA
|
l
l
TGGAGGCGCTGTGGGGCCACCCAAATTG,|ATAATAlC CCC AACCGCCTCTTCCCGCCTCAsTTTCAGCCTCACCA
CCATCE3gGATAGA
Bgl I Major Cap Sites Hph I
Py
Early
mRNA
FIG. 2. Maps of plasmidscontaining SV40-polyomavirus recombinants. (A)Adetaileddescriptionof eachplasmidisgivenin thetext.In brief, p85.3.SPcontains the 5' half of the polyomavirus early region inserted between the HpaIIandEcoRI sites ofSV40DNA. Stippled regions indicate SV40 DNA,openregionsindicatepolyomavirus DNA,and thin linesindicate the vector.TheexpectedmRNAsencoding smallT, middle T, anda truncated largeTproteinareindicated;thecoding regionsareboxed and indicate three differentreadingframes (black, hatched,ordotted).spMT1andspST18arevariants ofp85.3.SPlackingthe introns of the middle T and small TmRNAs, respectively. spLT5contains thevirtually complete polyomavirus early region lacking onlythelargeTproteinintron.(B)Junctions between theSV40late promoterregion and the polyomavirus earlygenesin the recombinantsp85.3.SP, pspLT5,pspMT1,andpspST18.Nucleotide numbers refer tothe SV system(44)forSV40andtoSoedaetal.(43)forpolyomavirus.Thelocations ofprincipalcapsites and thepolyomavirusTATAbox,
aswellas theATGcodoninitiating early proteintranslation, areindicated.
fragment containing part of the SV40 late region from the
EcoRI site (SV-nt 1,782) to the BamHI site (SV-nt 2,534), inserted into the vector pA6 (46). All of the plasmids described,and the details of the strategiesusedtoconstruct
them,are available upon request.
Growth of recombinant virus stocks. The SV40 polyoma-virus recombinant DNAs and theSV40early regiondeletion mutantpdl1055 (28)weredigestedwithBamHI and
recircu-larizedby ligationatlow DNA concentration(10 ,ug/ml).The
[image:3.612.89.541.72.583.2]ligation mixtures were extracted with phenol mixture (6),
[Hpa II]
-o1XhoIon November 10, 2019 by guest
http://jvi.asm.org/
POLYOMAVIRUS GENOMES THAT ENCODE THREE EARLY PROTEINS 173 lyophilized, redissolved in 100 pd of 1% NaOAc (pH
5.3)-0.1% sodium dodecyl sulfate (SDS), precipitated with
etha-nol, and run on a 1% low-melting-point agarose gel (Bethesda Research Laboratories, Gaithersburg, Md.). The super-coiled DNA bands were excised from the gel, heated for 5
minat 70°C, and cooledto 37°C. The DNA, without further
purification, was transfected into CV-1 cells with DEAE-dextran (25). For each transfection, 75 ng of supercoiled recombinantDNA plus 75ngof SV40 helper DNAwasused per
106
cells.After 10 days at37°C, the cellswere harvested by freezing and thawing them three times. These extracts (1 ml) were used undiluted to infect fresh monolayers(3 x 106 cells) of CV-1 cells. This cycle was repeated once more toobtain the third-passage virus stocksusedin the experiments reported here.
Analysis of polyomavirus early proteins. The procedures for labeling cells with
[35SJmethionine,
protein extraction, and immunoprecipitation have been described previously (19). The proteins were separated on 10% SDS-polyacryl-amide gels (22). Mouse monoclonal antibodytoSV40 large T protein L16 (15) was generously provided by Ed Harlow.Inmunofluorescence. Cellson cover slips were fixed witha
10% Formalin inphosphate-buffered saline solution at 42 to
48 h postinfection and stained for polyomavirus early
pro-teins with a rat antipolyomavirus tumor serum followed by incubation withafluorescein-conjugated rabbit anti-rat anti-body.
DNA binding assays. The large T protein-DNA binding assay was initiallydescribed by McKay (26). Herewe useda
procedure with modifications described in Haydayetal. (16) and a mixture of two monoclonal antibodies (aLT1 and xLT4) (4)specific forthe polyomavirus largeTprotein. The SV40 antibody was L16 (15).
Protein kinase assays.Theinvitro kinase reactions (37,42) were done as follows. Infected cells werelysed inabuffer (1
ml per 3x 106 cells) containing 0.02 M Tris-hydrochloride
(pH 8.0)-1% Nonidet P-40-0.15 M NaCl. Polyomavirus early proteins were immunoprecipitated from5 to
50-RI
samples(madeup to 150pul with NETbuffer containing 0.15 M NaCl, 0.005 M EDTA, 0.05 M Tris-hydrochloride [pH 7.4], 0.25%gelatin, 0.05%Nonidet P-40, and0.02% sodium azide) by incubationwith 6pul ofratantipolyomavirus tumor
serum and 150pul of 20% protein A-Sepharose (Pharmacia Fine Chemicals, Piscataway, N.J.) for 45 min at4°C. The immunoprecipitates werewashed oncewith phosphate-buff-ered saline, twicewith 0.5 M LiCl-0.1 M Tris-hydrochloride (pH 8.0), and resuspended into 80 pAl of 0.02 M Tris-hydrochloride (pH
7.5)-0.005
MMgCl2. [ry-32P]ATP (10to30pCi) wasadded, and the mixturewasincubated for15 min at
30°C.
Theimmunoprecipitates were washed threetimeswith NET buffer, resuspended into 30 [L of dissociation buffer (0.02 M Tris-hydrochloride [pH 6.8],2% SDS, 10%glycerol, 0.1 M dithiothreitol, 0.03% [wt/vol] bromophenol blue),boiled for 3 min, and analyzed on a 10%
SDS-polyacryl-amide gel. Partial proteolysis with chymotrypsin and
two-dimensional gel electrophoresis were done as described by
Schaffhausen et al. (39).
RESULTS
Construction of polyomavirus deletion mutants precisely
lacking introns of early region mRNAs. Our goal was to
separatelydelete from polyomavirusDNA thesingle introns
of the threedifferentearly region mRNAs (Fig.1A).
Remov-al of the middle T protein intron has been described
previ-ously (46). This was done by exchanging a unique AvaI
fragment spanning the 63-nucleotide intron with the
corre-sponding fragmentderived from a cloned partial cDNA copy
ofthemiddle Tprotein mRNA to generateplasmidpPyMT1.
(Fig.
1C)
(46). Exchange betweengenomic and cDNA clones was used in analogous but somewhat more complex con-structions (see above) to produce plasmids pPyLT1 andpPyST1
(Fig.1B
and D) which lack the 385 or the 49 nucleotides, respectively,corresponding
to the large T and small T protein introns. Removal of the intervening se-quences resolves the overlapping coding information of the polyomavirus early region at the DNA level. The DNA sequences of the viral early regionin pPyLT1, pPyMT1, and pPyST1 are such that their primary transcripts should be identical to the mature mRNAs, and therefore, if there were no interference with efficient mRNA biogenesis caused by intron deletion, they should uniquely encode large T, middle T, and small Tproteins.SV40-polyomavirus
recombinants expressing individual polyomavirus early proteins. We have already established that pPyMT1 encodes only the middle T protein (46) and published evidence suggesting thatpPyLT1 contains a func-tional large T protein gene (48). Further analysis of the coding capacities of the three modified early genes was facilitated by use ofSV40
viral vectors (10, 11, 14, 27) to express the proteins at elevated levels. To this end, we inserted the early genes of polyomavirus into the lateregion ofSV40.
Adetailed descriptionof the recombinants isgiven above. The initial construction was p85.3.SP (Fig. 2A), comprising a genomic polyomavirus DNA fragment encod-ing small T, middle T, and a truncated large T protein, inserted intoSV40
and replacing the 5' portion of the SV40 late region DNA; the recombinant was cloned into pAT153 (47).pspMT1
andpspST18
(Fig. 2A) are similar constructs. The polyomavirus genes, however, lack the middle T or small T protein introns and therefore encode only the respective T proteins. pspLT5 (Fig. 2A) contains acomplete early region of the polyomavirus genome lacking only the large T protein intron. The SV40 DNA fragment encoding part of the VP1 gene, which is present in the constructs p85.3.SP,pspMT1,
andpspST18,
is missing in pspLT5. The mRNA derived from the polyomavirus DNA in pspLT5 should use the polyadenylation site at the end of the large T protein gene, whereas transcription of p85.3.SP, pspMTI, andpspST18
should generate recombinant mRNAs compris-ing the 5' part of the polyomavirus early mRNAs and the 3' part of theSV40
VP1 mRNA. Polyadenylation in these constructs should occur at the signal normally used for theSV40
late messengers.Recombinant virus stocks were produced by excision of the viral genomes from recombinant plasmids (Fig. 2A) and cotransfection of CV-1 cells with these DNAs and the SV40 helper
dllO55
(28),a deletion mutant defective in earlyregion function (see above). After two subsequent undiluted pas-sages onto CV-1 monolayers, high-titer virus stocks were obtained which produced total cytopathic effect on CV-1 cells within 60 to 80 h after infection. Characterization ofthe viral DNA extracted from the infected cells demonstrated that 10 to 50% of the molecules were the expected SV40-polyomavirus recombinants (the rest being helper DNA) and that, in the virus stocks used for subsequent studies, rear-ranged DNAs were only minor components.We made virus stocks from a number of SV40-polyoma-virus recombinants with different junctions between the
SV40
late promoter and the beginning of the polyomavirus early region. The presence or absence of the majorinitiation site ofSV40
late transcription (nt 325) (33) did not influence the expression level of the polyomavirus early proteins.VOL.51,1984
on November 10, 2019 by guest
http://jvi.asm.org/
Furthermore, the presence ofthe polyomavirus early cap
sites (1, 27) was also irrelevant to the expression level
obtained. The particular junctions in p85.3.SP, pspLT5,
pspMT1, andpspST18 areshown inFig. 2B. The SV40 late promoterregion is linkedtopolyomavirus earlyDNAeither
preciselyatthe majorcapsite (pspLT5)or atpointsat most
50base pairs upstream (pspMT1). The translation initiation
codon(43)for theearlyproteins of
polyomavirus (Fig.
2B) isin all cases located at least 18base pairs downstream from
the junctions between the SV40 and polyomavirus DNA
sequences.Although analysis of transcripts derived from the
SV40-polyomavirus recombinants (datanotshown) revealed
no quantitative differences in mRNA levels among
con-structs with differentjunctions, the positions of5' termini
varied. One observation in particular is worthy ofnote. All
constructs including the polyomavirus major early
region
cap sites
(polyomavirus
nt148-153)
synthesized
mRNAsprincipally initiatedatthesepositions, irrespective of
wheth-er the polyomavirus TATAbox was present and also
irre-spective of the presence of the SV40 major late cap site
(SV40nt 325).
Expression of polyomavirus earlyproteins in CV-1 cells. If
theintron-minus variants ofthe
polyomavirus
early regionindeedfunctionally encodethe threeearlyproteins,then the
SV40-polyomavirus recombinants containing them should
individuallyexpress theproteinsinrelatively largeamounts.
CV-1 cells infected with the different viruses were labeled
for2 h with
[35S]methionine.
The celllysates
were divided among three samples which were thenimmunoprecipitated
(Fig. 3) with rat
antipolyomavirus
tumor serum (lanes P),normal rat serum (lanes N),or as areference standard with
mousemonoclonalantibody (15) directed
against
SV40 largeTprotein. spLT5-infected cells containedthe90kSV40
large
Tprotein, which wasexpressed fromtheSV40 early region
of the recombinant viralgenome,anda100k
protein
precip-itated by antipolyomavirus tumorserumwith the same
gel
mobilityasthepolyomavirus largeTprotein.
spMTl-infect-ed cells expressed, besides SV40 large T
protein,
the 56kspLT5 spMTI
polyomavirus middle T protein. Immunoprecipitation of the
spST18-infected cell lysate with antipolyomatumor virus
serumshowedthe 22k small T protein. Each of the polyoma-virus proteins synthesized in the monkey cells comigrated with the corresponding polypeptides immunoprecipitated from mouse cells infected with wild-type polyomavirus (data not shown). In no case was a polyomavirus protein other than the anticipated one detectable.
Thestability of the polyomavirus proteins expressed in the CV-1cells was determined in the same experiment. After the 2-hlabeling period, the cultures were chased for 2 and 5 h
(Fig. 3). The protein bands were excised from the gel and
quantitatedbyscintillation counting. The middle T and small
Tproteins had half-lives of 4 to 5 h, whereas polyomavirus large T protein was more stable, with a half-life of about 11 h.
Data from the experiment
presented
inFig.
3 furtherallowed the estimation ofsynthetic rates relativeto that of
theSV40largeT
protein
forthepolyomavirus early proteins
produced in themonkey cells. We chosetocompareratesof
synthesis to the SV40 large T reference standard because
this protein was encoded
by
the same DNA molecules aswere the polyomavirus proteins (Fig. 2) rather thanby the
helperDNA. Wefoundthat the
polyomavirus large, middle,
andsmall Tproteinswere
produced
at ca.1.5, 4.0,
and9.0times the molarrateoftheSV40
large
Tspecies,
respective-ly. These proportions varied somewhat among different
experimentsinwhichanumberof
independent
virus stockswere used, but the trend was the same: production rates
werelow with largeTand moderate withmiddle
T,
andonly
with the small T protein were they at the
anticipated high
level for
synthesis
directedby
theSV40late promoter.To assure that the polyomavirus
proteins
were indeedexpressed under control of the SV40 late promoter, we
demonstratedthattheir
synthesis
was blockedby
the DNAsynthesis inhibitorcytosine
arabinoside,
whereas the SV40large T expression was not affected, as shown for
spMT1-infected cells in
Fig.
4.Cytosine
arabinoside blocks viralspSTI8
0 2 5 0 2 5
S N P S N P S N P S N P S N P S N P
Q 2 5
0
25I
PP P
Py-Py-Lt~~~~~LT _ ~~** *~~~*
3-V-LT
SLSV-LT X
MT- i
-MT-4w'
0 2 5
S N P S N P S N P
6w-SV-LT- T..
I.
-*-ST- , *
FIG. 3. Immunoprecipitationof polyomavirus early proteins fromCV-1cells infected with the recombinant virus stock spLT5,spMT1,or
spST18.The cells were labeled 40 h postinfection with[35S]methioninefor 2 h and subsequently chased in Dulbecco modified Eagle medium
containing methionine for2 or 5 h.Polyomavirus early proteins as well asSV40large T protein were immunoprecipitated from cell lysates and
analyzed on 10% SDS-polyacrylamide gels. Symbols: S, SV40 large T monoclonal antibody (L16) (15); N, normal rat serum; P, rat
antipolyomavirustumorserum. In thespMT15-hchase, the amount of material loaded on the gel was lower due to a technical error. The
im-munoprecipitations with antipolyomatumor serum from spMT1 lysates chased for 0, 2, or 5 h were repeated and are shown in the adjacent
three lanes. These datawereusedtodeterminedthehalf-life ofpolyomavirus middle T protein expressed in CV-1 cells.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.86.541.474.665.2]POLYOMAVIRUS GENOMES THAT ENCODE THREE EARLY PROTEINS
39h
26h
26h
18h
18h
_ _
~+
- +S N P S N P S N P S N P S N P
...,,j
_~~,,
A_zw SE i -1'~~
spMTI
85.3.SP
S N P N P S N P N P
:
:Zw=
.
. .1
.~~~~~~~~~I.,
s,
X
2 -.- =_58 K -MT(56 K)
-Truncated LT (40K)
FIG. 4. CV-1 cells infected with spMT1 were grown in the presence(+)orabsence (-)of 40,g of cytosinearabinosideperml for18to39 hasindicated. The cellswerelabeled with
[35S]methion-ineand lysed, and samples oflysateswereimmunoprecipitated and
analyzed on SDS-polyacrylamide gel (see legend to Fig. 3 for details).
DNA replication and consequently late SV40 transcription. Our results, therefore, imply that the expression of the polyomavirus early proteins is under control of the late
promoterofSV40.
Individual expression of the polyomavirus early proteins was achieved bydeletion of the introns. In the case of the
rabbit beta-globin(13, 14) and SV40 VP1genes(12), removal
of intervening sequences drastically reduced the level of functional gene expression. Our previous experience (46)
suggested that RNAsplicingwas,bycontrast, notimportant for the efficient production of polyomavirus early region mRNAs. To address directly the question of whether intron deletion accounted for the rather low rate ofpolyomavirus proteinproduction measured in themonkey cells, we
com-pared theamount ofpolyomavirus middle Tprotein synthe-sized by spMT1 (intron-minus) with that made by the corresponding virus stock 85.3.SP (intron-plus) (Fig. 2A). The intron-minus polyomavirus DNA in spMT1-expresses the56k middle Tproteineven moreefficiently than doesthe unmodified genomic DNA in 85.3.SP (Fig. 5). Direct analy-sisof the mRNAsproducedby spMT1 showed thatthevast
majority initiatedatthepolyomavirus early regioncap sites (Fig. 2B), and thus they could not have been spliced by removal of introns within 5'-untranslated regions derived from SV40sequences in the vector.
The immunoprecipitationanalysis oftheproteins
synthe-sized in CV-1 cellsinfected by 85.3.SP (Fig. 5) revealed, in addition to the expected 56k middle T and 40k truncated largeT proteins, a prominent band at 58k. The 58k protein wasshowntobe relatedtomiddle Tprotein by immunopre-cipitation in which a middle T-specific antipeptide serum wasused(38) andby two-dimensional gel electrophoresis of
partial chymotryptic digests. Itdidnot reactwithSV40VP1 antiserum and therefore is unlikely to represent a fusion protein. Forreasons explainedbelow,wetentatively identi-fy this58kspecies as theproduct ofan erroneously spliced
mRNA resulting from utilization of a cryptic splice site withinthe middle T mRNAintron.
We conclude that the modified polyomavirus DNAs
in-FIG. 5. Analysis ofpolyomavirus early proteins expressed by
CV-1 cells infected with spMT1 or 85.3.SP. Infected cells were labeledwith[35S]methionine, and samplesoflysateswere
immuno-precipitated with either SV40 large T monoclonal antibody (S),
normal rat serum (N), or rat antipolyomavirus tumor serum (P).
85.3.SP-infectedcells expressed three polyomavirus proteinswith molecularweightsof58k, 56k (middle T),and 40k(truncatedlarge T), respectively; the22k small T protein, alsoexpressed in these
cells, was run off the gel shown here. spMTl-infected cells
ex-pressed only the 56k middleTprotein.
deedindividuallyexpresstheexpectedpolypeptides, but the expression levels obtained in SV40vectors were in general lower than expected. Splicing isnotanimportantstepinthe production of recombinant SV40-polyomavirus mRNAs. Moreover, introduction of polyomavirus genomicDNAinto recombinantgenomescanleadtotheactivation ofacryptic splice site which results in the synthesis of at least one
aberrantprotein.
Intracellular localization of polyomavirus early proteins. The individual expression of polyomavirus large, middle, and small Tproteins in CV-1 cells infected with the
appropri-ate SV40-polyomavirus recombinant viruses enabled us to
examine the intracellular localization of these proteins by simple indirect immunofluorescence with polyclonal
antitu-morserum(Fig. 6); thisserumdidnot reactwithCV-1 cells infectedby SV40 alone. Cells expressing polyomavirus large Tprotein showed the classicpatternof nuclear
[image:6.612.317.535.65.264.2]immunofluo-MT ST
FIG. 6. Immunofluorescence ofpolyomavirus earlyproteins ex-pressed in CV-1 cells infected by spLT5 (LT), spMT1 (MT), or
spST18 (ST). - SV-LT
- Py-MT
..,:.-. .:-,... ow-'%.V
'111w. .-,
..:V-domb-ow- --
d--- --. . lw
VOL. 51, 1984 175
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.49.285.66.250.2] [image:6.612.310.548.576.693.2]rescence with dark nucleoli which characterizes polyoma-virus-infected mouse cells, and thus this pattern must be predominantly due to the large T protein. Expression of the middle T protein resulted in an entirely different result. We found marked perinuclear immunofluorescence, possibly indicating localization in the Golgi apparatus, plus a struc-tured distribution throughout the cytoplasm without obvious concentration in the plasma membrane (Fig. 6B). Our visual-ization of middle T protein in CV-1 cells is virtually indistin-guishable from the pictures obtained with mouse cells infect-ed by wild-type polyomavirus and middle T-specific monoclonal antibodies (S. Dilworth, personal communica-tion). The results obtained with CV-1 cells overexpressing the small T protein, however, were novel. Monoclonal antibodies specific for this protein do not exist, and thus it hasneverbeenvisualized in cells before. The small T protein is present in the nuclei and in thecytoplasm oftheinfected CV-1 cells (Fig. 6C). Most notable, however, was the coral-likeappearanceof CV-1cells expressing the protein. Phase-contrast observation of the cells revealed the same drastic change in morphology relative to uninfected CV-1 cells or CV-1 cells infected by other SV40-polyomavirus recombi-nant viruses. We suspect from these observations that
overexpression ofpolyomavirus small T protein is toxic to
cells and causes dramatic changes in the cytoskeleton. We shall return to this point below, when we compare our findings with other recent results obtained with both the
polyomavirus (M. Rassoulzadegan and F. Cuzin, personal
communication) andSV40 (D. Livingston, personal
commu-nication) small Tproteins.
Biochemical activities of the polyomavirus early proteins
expressedin CV-1cells. We tested the specificDNA binding
activity of the polyomavirus large T protein expressed in
CV-1 cells infectedwith spLT5 by the immunoprecipitation
method (26). Radiolabeled fragments of viral DNA were
incubated withcrude cell extract. The large Tprotein-DNA
complexes were precipitated with two monoclonal
antibod-ies(4) specificfor the polyomavirus large T protein, and the
precipitated DNA fragmentswere identifiedby agarose gel
electrophoresis. Cell lysates from polyomavirus-infected
3T6 cells and SV40-infected CV-1 cells served as controls. The amounts of lysate used in the binding assays were
adjusted so that approximately equal quantities of total
proteinwere used. Polyomavirus large Tprotein expressed
in monkey cells binds specifically to the same regions of
polyomavirus DNA asdoes authenticprotein from infected
mouse cells (Fig. 7A). As described elsewhere (S. M.
Dilworth, A. Cowie, R. I. Kamen, and B. E. Griffin, Proc.
Natl. Acad. Sci. U.S.A., in press; and A. Cowie and R. Kamen, manuscript in preparation), the majorbinding site
withinthe DNAfragment designated "a" inFig. 7A occurs
adjacentto theviral originofDNAreplication onthe early
region side. The minor binding site located in DNA fragment
b(Fig. 7A) maps from the TATA box to the cap sites of the
principal early region promoter. The amount of specific
DNAbinding activity recovered from the monkey cells was very similar to that extracted from the mouse cells. Howev-er, because CV-1 monkey cells do not reach the same cell densities as 3T6 mouse cells, the level of active polyoma-virus large T protein produced perculture infected by the recombinant virus spLT5 is lower than that of wild-type
polyomavirusinfections of 3T6 mouse cells. The SV40large
T protein extracted from SV40-infected CV-1 cells binds
efficientlyto the same polyomavirus DNAfragments as the
polyomavirus protein (for this measurement, we used a
monoclonalantibody specific forSV40largeTprotein)(Fig.
3T6 CVI CVI
x ~~x x
A,
rnPy
wt sp LT5SV40
mA 0020--0
15 10 25 50 20 40 100 200 20 40100200
.. . ..
.w.
-S-
-
.4:
- a
-b
CVI CVI 3T6
x x x
B -- s
spLT5
SV40
Pywt
m5 10 25 501 10 25
5df1t.25.5
6 12 "*_i.
S
-c0
FIG. 7. Immunoprecipitationassaysof thespecificDNAbinding activities of polyomavirus and SV40 large T proteins. Extracts
prepared from either mouse ormonkey cells infected with various virus stocks were incubated with 32P-labeled restrictiondigests of either cloned polyomavirusDNA (A) orSV40 DNA (B). Specific restrictionfragments boundtothelargeTproteinswere immunopre-cipitated withmonoclonal antibodies directedagainstthe polyoma-virus or SV40 large T proteins and identified by agarose gel electrophoresis followed by autoradiography. (A) The 32P-labeled
DNA wasderivedfrom plasmidp37.3.A2 (a recombinant substan-tiallyidentical top53.A6.6shown inFig. 1)by combineddigestion withrestriction endonucleasesBamHI, Bgll, andSacl to generate
the ninefragments run asmarkers in tracks m. Thetwofragments (designated aandb) knowntocontainhigh-affinity bindingregions (9, 29; Dilworthetal.,inpress;Cowie andKamen, inpreparation) for polyomavirus large T protein were immunoprecipitated by monoclonal antibodies to polyomavirus large T protein from ex-tractsofpolyomavirusstrain A2-infectedmouse3T6 cells(3T6xPy wild type) andfrom extracts of recombinant virusspLT5-infected CV-1 cells (CV-1 x spLT5). The numbers above each gel lane indicate the volumes ofextract used, whichwere adjusted among
differentlysatestoobtainca.30, 60,150, and300
F±g
of totalprotein for the fourassaysdoneoneachextracttested.PolyomavirusDNAfragments aand b werealso immunoprecipitated fromextracts of SV40-infectedCV-1 cellsby reactionwithamonoclonalantibodyto
SV40largeTprotein(CV-1x SV40).(B)The32P-labeledDNAused was aHindIII digest of SV40DNA (track m) in whichfragmentc containsthe knownSV40largeT DNAbindingregions. Fragmentc was immunoprecipitatedfrom extractsofspLT5-orSV40-infected
CV-1 cellsbyantibodytoSV40largeT(CV-1x spLT5and CV-1 x SV40)butnotfromextractsofpolyomavirus-infected3T6cells(3T6
x Py wild type) by monoclonal antibodies to the polyomavirus protein.
7A). By contrast, the results in
Fig.
7B show that thepolyomavirus
protein
doesnotinteract with SV40DNA ina manner detectableby
theassay weused.Polyomavirus
middle Tprotein
can bephosphorylated
aton November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.323.559.76.385.2]POLYOMAVIRUS GENOMES THAT ENCODE THREE EARLY PROTEINS 177
tyrosine residues by usinganin vitro protein kinaseassay(3,
5, 40, 42). Although it is not definitely known whether the kinase involved isthemiddleTprotein itselforanassociated cellular enzyme, the recent work of Courtneidge and Smith (2) argued for the latter alternative and suggested that the kinase is pp60csr", complexed to middle T protein. We thought that it wouldbeof interesttodetermine whetherthe middle T protein synthesized in CV-1 monkey cells could be phosphorylated at tyrosine residues in vitro. Were this the case, we could exploit the relative overproduction of the protein in monkey cells to determine whether the kinase activity increased proportionally. Middle T protein was
immunoprecipitated from a lysate of spMT1-infected CV-1 cells with rat antitumor serum. The immune complex was
incubated with [_y-32P]ATP and analyzed on an
SDS-poly-acrylamide gel (see Fig. 9B). Two32P-labeled proteins were
observed, one of which comigrated with the 56k middle T species immunoprecipitated from a [35S]methionine-labeled cell extract (data not shown); the other protein represents
the inimunoglobulin G of which the heavy chain was phos-phorylated in this assay.
The 32P-labeled middle T protein was characterized in
more detail by partial chymotryptic digestion and two-dimensional gel electrophoresis (Fig. 8). Qualitatively, the patterns from the CV-1 cells and the 3T6 cells are identical. This result, as well as V8 protease and cyanogen bromide mapping results (datanotshown),indicatesthat the middle T protein expressed in CV-1 cells is phosphorylated at the same residues as the middle T protein from 3T6 cells. The major phosphorylation site is tyrosine 315, but tyrosine 322 is also significantly labeled (39; B. Schaffhausen, manuscript inpreparation). Quantitatively, there is a striking difference between the middle Tproteins expressed in3T6 cellsand in CV-1 cells. SchaffhausenandBenjamin (37) have shownthat
[35S]methionine-labeled
3T6 cells infected with polyoma-virus expresstwo forms ofmiddle T protein, the 56k and58k species, in a 10 to 1 ratio. The 58k species, which differs from the 56k species in its invivo phosphorylation pattern,has a 10-foldhigher specific activity in the in vitro phosphor-ylation reaction, becausethe ratio of32P-labeled 56k and58k middle T proteinsfound afterin vitro phosphorylationisone
to one. Figure 8 compares the chymotrypticfragmentsofthe 56k and 58k middleTspecies afterinvitrophosphorylation. Mouse 3T6 cells or BMK cells infected with wild-type polyomavirus showed equal amounts of the doublet pairs 39k-33k (derived from 58k) and 37k-31k (derived from 56k). CV-1 cells infectedwith spMT1, however, showed clearlya much higher level ofthe 56k deriveddoublet compared with the 58k derived fragments after in vitro phosphorylation. Whether thisdifference wascausedbyalowerinitial ratioof 58k to 56k middleTproteinorbyareducedspecific activity of the 58k species in the in vitro phosphorylation reaction was furtherexaminedby analysis ofchymotrypticfragments of in vivo-phosphorylated middle T protein derived from spMT1-infected CV-1 cells (data not shown). No in vivo-phosphorylated 58k middle T protein could be detected, indicating indeed a reduced ratio of 58k to 56k protein in spMT1-infected cells compared with wild-type
polyoma-virus-infected mouse cells.
The overproduction of middle T protein in monkey cells
enabled us to study the correlation between the amount of
middle T protein and the in vitro kinase activity. We
compared lysates containing middle T protein from CV-1
cells infected with spMT1 with lysates from 3T6 cells
infected with either wild-type polyomavirus or a tempera-ture-sensitive large T protein mutant (7) which was grown
3T6
56
58
3r
3
31
cv-'
BMK
5856
1.58
[image:8.612.315.554.80.284.2]p.0
FIG. 8. Partial chymotryptic digestion of polyomavirus middleT proteins labeled in vitro with [y-32P]ATP. Polyomavirus proteins were isolated from 3T6 cells or BMK cells infected withwild-type polyomavirus or from CV-1 cells infected with spMT1. After immunoprecipitation, the proteins were labeled and run on 10-cm cylindrical 10% SDS-polyacrylamide gels. Digestion with chymo-trypsin was carried out on 12.5% SDS-polyacrylamide gels. Arrow-heads indicate positions of the 39k-33k and 37k-31k doubletsderived from the 58k and 56k middle T species, respectively.
for 36 h at the permissive temperature
(32°C)
and then shifted to the nonpermissive temperature(39°C)
for an additional 6 h. This protocol derepresses early mRNA synthesis and results in the synthesis of elevated amountsof middle T protein. The quantities of middle T protein present in these infected cells were determined in a continuous labeling experiment. Cells were infected with the appropri-ate virus stocks and labeled with[35S]methionine
for 42 h under conditions of methionine excess. Middle T protein was immunoprecipitated from cell lysates containing equal amounts of total protein. CV-1 cells infected with spMT1 contained5 times more middle T protein than the mouse cells infected with the temperature-sensitive mutant and 20 times more than cells infected with wild-type polyomavirus (Fig. 9). The results of in vitro kinase assays are shown in Fig. 9B. Increasing amounts oflysatescontaining 7, 28, and 70,ug
of total protein were immunoprecipitated and incubat-ed with[-y-32P]ATP.
The32P-labeled
proteins were analyzed on an SDS-polyacrylamide gel (Fig. 9B). With each lysate, the amount of32P-labeled
middle
T protein obtained was directly proportional to the quantity of extract protein add-ed. This indicated that neither the ATPconcentrationnorthe antibody concentration was limiting the assay. The level of kinase activity in the activity in the extract of spMT1-infected CV-1 cells was somewhat lower than the levels obtained with the mouse cell extracts (Fig. 9B). It is appar-ent that the level of kinase activity does notcorrelate at all with the amount ofmiddleT protein expressed by theCV-1 cells. This may indicate that a factor which is essential for the in vitro kinase activity is limiting in the monkey cell extract. Two possible explanations for this will be discussed.DISCUSSION
We constructed polyomavirus mutants which lack the introns of the early region mRNAs. The mutants pPyLT1, VOL.51, 1984
on November 10, 2019 by guest
http://jvi.asm.org/
A
In vivo
Continuous Labeling
3 3T6 3T6 CVI
x x x
Py wt Py tsAspMTI 7
'...M
B
In
v/f
ro
Phosphory lat ion
T6 x Py wt 3T6 x Py tsA CVI x spMTI
I I --II I
28 70 7 28 70 7 28 70
_
FIG. 9. Comparison of middle T protein production and in vitro kinase activity in lysates of mouse 3T6 cells infected with either wild type or atemperature-sensitive (tsA) mutant of polyomavirus or of CV-1 cells infected with spMT1. (A) Cells were labeled continuously with
[35S]methionine during the infection period. Cell lysates containing equal amounts of total proteins were immunoprecipitated with rat
antipolyomavirus tumor serum and analyzed on 10% SDS-polyacrylamide gels. The middle T protein bands were quantitated by densitometry. (B) In vitro kinase activity assay onlysates containing increasing amounts (7, 28, or 70
pLg)
of total protein. The32P-labeled middle T protein bands were excised from the gel and counted in a scintillation counter.pPyMT1, (46), and pPyST1 encode thepolyomavirus early
gene products large T, middle T, and small T protein,
respectively. SV40-polyomavirus recombinant DNAs, in
which part of the SV40 late region was replaced by the
modified polyomavirus early genes, were introduced into
CV-1 cells in the presence ofan SV40 helper DNA. The
recombinantvirus stocksobtained inducedtheexpression of
the individual polyomavirusearlyproteins uponinfection of
CV-1 cells as was shown by immunoprecipitation of the
proteins from
[35S]methionine-labeled
cellextracts.The expressionlevelsobtained in CV-1 cells infected with the recombinantvirus stocksspLT5 and spMT1werelower thanexpectedfor synthesis directed bythe late promoterof
SV40. spST18-infected cells, however, expressed small T
protein at the anticipated high level. The mRNA levels
expressedbythe threerecombinantvirus stockswere
indis-tinguishable
by
Northern blot andSi
mappinganalysis (datanotshown), suggestingthat thedifferenceinexpression may
be related to differences in the translation efficiency of the mRNAsor to otherundetermined factors.
The virus stock spLT5 is not veryuseful forthe
produc-tion of large amounts of polyomavirus large T protein because theexpressionlevelisessentiallythesame asthatof
wild-type polyomavirus-infected mouse cells. spMT1
ex-pressesmiddle Tproteinin CV-1 cellsat alevel that ishigher
thanwild-typepolyomavirus-infected mousecells. A
contin-uouslabeling experiment (Fig. 9A) showed thatweobtained
inmonkey cells five timesmoremiddleTproteinthanin 3T6 cells infected with the temperature-sensitive mutant. This system, therefore, could be used to produce middle T
protein forsubsequentpurification. The spST18virus stock
offers anexcellent systemforhigh-levelexpressionof small T protein which could subsequently be purified from cell
lysates
of infected CV-1 cells.Theexpression of the individual polyomavirus early
pro-teins enabled us to study their intracellular localization by
indirect immunofluorescence. Large Tproteinwaslocalized
in the nucleus. Middle T protein was largely perinuclear,
probablyinthe Golgi apparatus, as well as found throughout
thecytoplasm in a structure which could be the endoplasmic
reticulum. Small T protein was found in the cytoplasm as
well as in the nuclei of infected CV-1 cells. Recently, SV40 small T protein was also observed in both the nuclei and
cytoplasm of infected CV-1 cells (D. Livingstone, personal
communication). Therole of small T protein,
especially
as anuclear protein, is still obscure. Another observation was that the cells expressing large amounts of polyomavirus small Tprotein were rounded up and detached easily from
theplasticculturedishes. This isconsistent withthe results
of effortstoestablish mouse or ratcell linesexpressing only
the small T protein by transfection with pPyST1 DNA (J. Griffin and R. Kamen,unpublished data;M.Rassoulzadegan and F. Cuzin, personal communication); the transfected
cells detach fromthesubstrate(butdonotdie)andcannotbe
propagated on unmodified plastic surfaces. It is, therefore,
likely that polyomavirus small T protein disrupts the
cyto-skeleton of the cells, as wasfound for SV40 smallTprotein
(34).
Inthe comparisonofmiddleTproteinexpressionin CV-1 cellsbythevirus stocks spMT1 (intron-minus) and 85.3.SP
(intron-plus), weobserved, besides theexpected56k(middle
T) and 40k (truncated large T)proteins,aprominent protein
of 58k immunoprecipitated by antipolyomavirus tumor
se-rumfrom85.3.SP-infected celllysates (Fig. 5). Thisprotein
could be precipitated with middle T-specific antipeptide serum (38) but notwith antiserum against SV40 VP1 (data not shown); it is therefore probably not a fusion protein
includingthe SV40 sequence. Moreover,peptidepatternsof
58k and56k middleTspeciesobtainedbypartial
chymotryp-tic digestion and two-dimensional gel electrophoresis were
AP
It
'P.,(i
*0
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.146.480.74.305.2]POLYOMAVIRUS GENOMES THAT ENCODE THREE EARLY PROTEINS 179 virtually indistinguishable (i.e., the 37k-31k doublet was
found in both patterns). This distinguishes the
85.3.SP
58kprotein from the 58k middle Tcharacterized bySchaffhausen and Benjamin (37) because the latter yields a 39k-33k dou-blet. The 58k protein from 85.3.SP-infected CV-1 cells was further tested in the in vitro kinase assay and proved not to be a phosphorylatable substrate. This 58k middleTspecies, therefore, does not represent the modified form of 56k middle T identified by Schaffhausen and Benjamin (37), which was an enhanced activity as a substrate in the in vitro kinase assay. The two recombinants 85.3.SP and spMT1are virtually identical (Fig. 2) with no major differences in the regions of transcription initiation, the cap sites, or the 3'-junction between the polyomavirus and
SV40
sequences. They differ,however,
in the presence of the 63-base pair middle T intron in 85.3.SP. We therefore suspect that the58k middleT species is the product of an aberrant splicing event within the intron for the middle T protein. Upon investiga-tion of the intron DNA sequences (nt 747-808) (44) anumber of potential splice sites were identified which do notchange the translation reading frame of the mRNA. Splicing be-tween nt 747 and nt 760 would generate a middle T species containing an extra 16 amino acids, whereas splicing be-tween nt 766 and nt 794 (43) would encode a proteinhaving an additional 11 amino acids; these would cause increasesin molecular weight of 2k and 1.3k, respectively, either of which could account for the observed 58k species. Such aberrant splicing has never been observed in wild-type polyomavirus early mRNAs. If the recombinant transcripts made by 85.3.SP indeed use one of the alternative splice sites, one might argue that a full-length polyomavirus early transcript somehow is required to prevent incorrectsplicing. In an experiment to study the correlation between the amount of middle T protein and in vitro kinase activity, we compared the specific activities of middle T from lysates ofspMT1-infected
CV-1 cells and polyomavirus-infected 3T6 cells (Fig. 9). Although5 to 20 times more middle T protein was present inspMT1-infected
cell lysates, the in vitro kinase activity was very similar to the activity of lysates from polyomavirus-infected mouse cells. The absence of a proportional increase in activity might indicate that afactor which is essential for in vitro kinase activity is limiting in the CV-1 cell extract. This result is consistent with the proposal (2) that the enzyme responsible for in vitro phosphorylation of middle T protein ispp60-src,
a protein present in small amounts in all mammalian cells. Because only a small proportion of the middle T protein present in infected cellsis associated with the kinase activity (3) or occurs as a complex withpp60c-sr
(2), one would not expect, underthis hypothe-sis, to observe increased kinase activity by overproduction of the middle T protein. Another observation, however, must be considered. The ratio of the 58k and 56k middle T species expressed in monkey cells is lower than in wild-type polyomavirus-infected mouse cells. This was found by chy-motryptic mapping of protein phosphorylated in vitro and subsequent two-dimensional electrophoresis (Fig. 8). The relatively lower in vitro kinase activity of monkey cell lysates might reflect the amount of 58k middle T species rather than the total amount of middle T protein. It will be of interest to determine whether the fraction of middle T protein that is associated withpp6O-crc
represents predomi-nantly in vivo phosphorylated middle T species such as the 58k protein.The modified polyomavirus genomes whose construction and characterization are described have already been instru-mental in the elucidation of oncogenic transformation
in-duced
by
both viral and cellular oncogenes (23, 30-32, 35,46).
Separate
roles have been assigned to the middle andlarge T proteins in the alteration of cell growth regulation
(30-32). Future experiments will be directed towards
deter-mination of the function of the small T protein in the
transformation of nonestablished cells and towards an
un-derstandingoflarge Tprotein-mediated establishmentatthe
biochemical level.
ACKNOWLEDGMENTS
We thank Joe Sambrook forthe plasmid d12005, David
Living-stonefor providinguswith SV40 VP1 antiserum, Mary-Jane Geth-ing for useful discussions, and Marybeth Erker for her help in
preparingthe manuscript.
LITERATURE CITED
1. Cowie, A., P. Jat, and R. Kamen. 1982. Determination of
sequences atthe capped 5' ends of polyoma virus early region transcripts synthesized in vivo and in vitro demonstrates an
unusual microheterogeneity. J. Mol. Biol. 159:225-255. 2. Courtneidge, S. A., and A. E. Smith. 1983. Polyoma virus
transforming protein associates with the product of the c-src
cellular gene. Nature (London) 303:435-439.
3. Dilworth, S. M. 1982. Protein kinase activities associated with distinct antigenic forms of polyoma virus middle T antigen. EMBO J. 1:1319-1328.
4. Dilworth, S. M., and B. E. Griffin. 1982. Monoclonal antibodies against polyoma virus tumor antigens. Proc. Natl. Acad. Sci. U.S.A. 79:1059-1063.
5. Eckhardt, W., M. A. Hutchinson, and T. Hunter. 1979. An
activity phosphorylating tyrosine inpolyomaTantigen immuno-precipitates. Cell 18:925-933.
6. Favaloro, J., R. Treisman, and R. Kamen. 1980. Transcription maps of polyoma virus-specific RNA: analysis by two-dimen-sional nuclease S1 gel mapping. Methods Enzymol. 65:718-749. 7. Fried, M. 1965. Cell transforming ability of a temperature-sensitive mutant of polyoma virus. Proc. Natl. Acad. Sci. U.S.A. 53:486-491.
8. Friedmann, T., A. Esty, P.LaPorte, andP.Deininger. 1979. The nucleotide sequence and genome organizationof the polyoma early region: extensive nucleotide and amino acid homology with SV40. Cell 17:715-724.
9. Gaudray, P., C. Tyndall, R. Kamen, and F. Cuzin. 1981. The
high affinity binding site on polyoma virus DNA for the viral large T protein. Nucleic Acids Res. 9:5697-5710.
10. Gething, M.-J., and J. Sambrook. 1981. Cell-surface expression of influenza haemagglutinin from a cloned DNA copy of the RNA gene. Nature (London) 293:620-625.
11. Gruss, P., and G. Khoury. 1982. Gene transfer into mammalian cells: useof viral vectors to investigateregulatory signalsfor the expression of eukaryotic genes. Curr. Top. Microbiol. Im-munol. 96:159-170.
12. Gruss, P., C.-J.Lai, R.Dhar, andG.Khoury. 1979.Splicingas a
requirement forbiogenesis offunctional 16S mRNAs of simian virus 40. Proc. Natl. Acad. Sci. U.S.A. 76:4317-4321. 13. Hamer, D. H., and P. Leder. 1979. Splicing andtheformation of
stable RNA. Cell 18:1299-1302.
14. Hamer, D. H., and P. Leder. 1979. Expression of the chromo-somal mouse,B-globin gene cloned in SV40. Nature (London) 281:35-40.
15. Harlow, E., L. V. Crawford, D. C. Pim, andN. M. Williamson. 1981. Monoclonal antibodies specific for the simian virus 40
tumor antigens. J. Virol. 39:861-869.
16. Hayday, A. C., F. Chaudry, and M. Fried. 1983. Loss of
polyoma virus infectivity as a result of a single amino acid change in a region ofpolyoma virus largeT-antigen which has
extensive amino acid homology with simian virus 40 large
T-antigen. J. Virol. 45:693-699.
17. Hunter, T., M. A.Hutchinson, and W.Eckhardt. 1978. Transla-tion of polyoma virusTantigensinvitro. Proc. Natl. Acad.Sci.
U.S.A. 75:5917-5920.
18. Hutchinson, M. A., T.Hunter, and W.Eckhardt. 1978. Charac-VOL. 51,1984
on November 10, 2019 by guest
http://jvi.asm.org/
terization of T antigens in polyoma-infected and transformed cells. Cell15:65-80.
19. Ito, Y., N. Spurr, and R. Dulbecco. 1977. Characterization of polyoma virus T antigen. Proc. Natl. Acad. Sci. U.S.A.
74:1259-1263.
20. Jat, P., U. Novak, A. Cowie, C. Tyndall, and R. Kamen. 1982. DNA sequences required forspecific andefficient initiation of transcription at the polyoma virus early promoter. Mol. Cell.
Biol. 2:737-751.
21. Kamen, R., P. Jat, R. Treisman, J. Favaloro, and W. R. Folk. 1982. 5' Termini of polyoma virus early region transcripts synthesized in vivo by wild type virus and viable deletion mutants. J. Mol. Biol. 159:189-224.
22. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
23. Land, H., L. F. Parada, and R. A. Weinberg. 1983. Tumorigenic conversion of primary embryo fibroblasts requires at least two cooperating oncogenes. Nature (London) 304:596-602.
24. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular
cloning. Cold Spring Harbor Laboratory. Cold Spring Harbor,
N.Y.
25. McCutchan, J. H., and J. S. Pagano. 1968. Enhancementofthe
infectivity of simian virus 40 deoxyribonucleic acid with dieth-ylaminoethyl-dextran.J. Natl. Cancer Inst. 41:351-357. 26. McKay, R. D. G. 1981. Binding ofasimian virus40 Tantigen
related proteinto DNA. J. Mol. Biol. 145:471-488.
27. Mulligan, R. C., B. H. Howard, and P. Berg. 1979. Synthesis of
rabbitbetaglobin in cultured monkey cellsfollowing infection withaSV40 beta globin recombinant genome. Nature (London)
277:108-114.
28. Pipas, J. M., S. P. Adler, K. W. C. Peden, and D. Nathans. 1980.
DeletionmutantsofSV40that affectthestructure of viraltumor
antigens. Cold Spring HarborSymp. Quant. Biol. 44:285-291. 29. Pomerantz, B. J., C. R. Mueller, and J. A. Hassell. 1983.
Polyomavirus largeTantigenbinds independently tomultiple, unique regionson theviralgenome.J. Virol. 47:600-610. 30. Rassoulzadegan, M., A. Cowie, A. Carr, N. Glaichenhaus, R.
Kamen, and F. Cuzin. 1982. The roles of individual polyoma
virus early proteins in oncogenictransformation. Nature
(Lon-don)300:713-718.
31. Rassoulzadegan, M., A. Cowie, A. Carr, N. Glaichenhaus, R. Treisman, J. Favaloro, F. Cuzin, and R. Kamen. 1982.Separate butcomplementary roles for polyoma virusearly proteinsin the
alteration of cellgrowth requirementsleadingtotransformation,
p. 269-293. In D. L. Robberson and G. F. Saunders (ed.), Perspectives on genes and the molecular biology ofcancer. Raven Press,NewYork.
32. Rassoulzadegan, M., Z. Naghashfar, A. Cowie, A. Carr, M.
Grisoni,R.Kamen,and F.Cuzin. 1983. Expressionof thelarge
T protein of polyoma virus promotes the establishment in
culture of "normal" rodent fibroblast cell lines. Proc. Natl. Acad. Sci. U.S.A. 80:4354-4358.
33. Reddy, V. B., P. K. Gosh, P. Lebowitz, and S. M. Weissman. 1978.Gaps and duplicatedsequencesintheleaders of SV4016S RNA. Nucleic AcidsRes. 5:4195-4213.
34. Rubin, H., J. Figge, M. T. Bladon, L. B. Chen, M. Ellman, I. Bikel, M. Farrell, and D. M. Livingston. 1982. Role of small t antigen in theacutetransformingactivityofSV40. Cell 30:469-480.
35. Ruley, E. 1983.Adeno early region1Aenables viralandcellular transforminggenes totransform primary cells inculture.Nature (London) 304:602-606.
36. Schafibausen,B. 1982.Transforminggenesandgeneproducts of polyoma and SV40. Crit. Rev. Biochem. 13:215-286.
37. Schaffhausen, B., and T. L. Benjamin. 1981. Comparison of phosphorylation oftwopolyoma virus middleTantigens in vivo
and invitro. J.Virol. 40:184-196.
38. Schaffhausen, B., T. L. Benjamin, L. Pike, J. Casnellie, and E. Krebs. 1982. Antibody to the nonapeptide Glu-Glu-Glu-Glu-Tyr-Met-Pro-Met-Glu is specific for polyoma middleTantigen
andinhibits in vitro kinase activity. J. Biol. Chem. 257:12467-12470.
39. Schaffhausen, B., J. Silver, and T. Benjamin. 1978. Tumor
antigen(s) incells productively infected bywild type polyoma
virusand mutant NG18. Proc. Natl. Acad. Sci. U.S.A. 75:79-83.
40. Schaffhausen, B. S., and T. L. Benjamin. 1979. Phosphorylation ofpolyomaTantigens. Cell 18:935-946.
41. Sleigh, M. J., W. C. Topp, R. Hanich, and J. F. Sambrook. 1978. MutantsofSV40with an altered small Tproteinarereduced in theirabilitytotransformcells. Cell 14:79-88.
42. Smith, A. E., R. Smith, B. E. Griffin, and M. Fried. 1979.
Protein kinaseactivity associated with polyoma virus middle
T-antigenin vitro. Cell 18:915-924.
43. Soeda, E., J. R. Arrand, N. Smolar, J. E. Walsh, and B. E.
Griffin. 1980. Coding potential and regulatory signals of the polyoma virus genome. Nature(London)283:445-453.
44. Tooze,J. (ed.).1980. The molecularbiology oftumorviruses.II.
DNA tumor viruses. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
45. Treisman, R., A. Cowie, J. Favaloro, P. Jat, and R. Kamen. 1981. The structuresofthe spliced mRNAsencoding polyoma
virusearly region proteins. J. Mol. Appl. Genet. 1:83-92. 46. Treisman, R., U. Novak, J. Favaloro, and R. Kamen. 1981.
Transformation ofratcellsbyanalteredpolyoma virusgenome
expressing only the middle-T protein. Nature (London) 292: 595-600.
47. Twigg, A. J., and D. Sherratt. 1980. Trans-complementable copy-numbermutantsofplasmid ColEl. Nature(London)382: 216-218.
48. Tyndall, C., G. LaMantia, C. M. Thacker, J. Favaloro, and R. Kamen. 1981. A region of the polyoma virusgenomebetween the replication origin and late protein coding sequences is required in cis forboth early gene expression and viral DNA
replication. Nucleic Acids Res. 9:6231-6250.

![FIG. 3.containinganalyzedantipolyomavirusmunoprecipitationsthreespST18. Immunoprecipitation of polyomavirus early proteins from CV-1 cells infected with the recombinant virus stock spLT5, spMT1, or The cells were labeled 40 h postinfection with [35S]methio](https://thumb-us.123doks.com/thumbv2/123dok_us/1426588.95304/5.612.86.541.474.665/containinganalyzedantipolyomavirusmunoprecipitationsthreespst-immunoprecipitation-polyomavirus-proteins-infected-recombinant-labeled-postinfection.webp)


![FIG. 8.trypsinwerepolyomavirusimmunoprecipitation,cylindricalproteinsfromheads Partial chymotryptic digestion of polyomavirus middle T labeled in vitro with [y-32P]ATP](https://thumb-us.123doks.com/thumbv2/123dok_us/1426588.95304/8.612.315.554.80.284/trypsinwerepolyomavirusimmunoprecipitation-cylindricalproteinsfromheads-partial-chymotryptic-digestion-polyomavirus-middle-labeled.webp)
![FIG. 9.ordensitometry.antipolyomavirusmiddle[35S]methionine a Comparison of middle T protein production and in vitro kinase activity in lysates of mouse 3T6 cells infected with either wild type temperature-sensitive (tsA) mutant of polyomavirus or of CV-1](https://thumb-us.123doks.com/thumbv2/123dok_us/1426588.95304/9.612.146.480.74.305/ordensitometry-antipolyomavirusmiddle-methionine-comparison-production-temperature-sensitive-polyomavirus.webp)
