Copyright © 1984, American Society for Microbiology
Comparative Sequence Analysis
of
Rotavirus
Genomic Segment
6-the Gene
Specifying Viral Subgroups
1and
2
GERALD W. BOTH,1 LINDA J. SIEGMAN,' A. RICHARD BELLAMY,2 3* NOBUKO IKEGAMI,4 AARON
J. SHATKIN2 AND YASUHIRO FURUICHI2
Division of Molecular Biology, Commonwealth ScientificandIndustrial Research Organization, North Ryde, NewSouth
Wales, 2113Australia1; Roche Instituteof Molecular Biology, Roche Research Center, Nutley, New Jersey 071102; Department ofCell Biology, Universityof Auckland, Auckland, 1, NewZealand3;and Clinical ResearchInstitute, Osaka
NationalHospital, Osaka, 540, Japan4
Received2February 1984/Accepted 22 March 1984
Cloned DNA copies of rotavirus genomic segment 6 from simian 11 (subgroup 1) and human strain Wa (subgroup 2) rotaviruses have been usedtodetermine thenucleotidesequencesof thegenethatdetermines viral subgroup specificity. Both genomic segments are 1,356 nucleotides in length andpossess5'- and 3'-terminal untranslatedregions of 23 and142nucleotides, respectively. The inferred amino acidsequencerevealsVP6 to beapolypeptide of 397 amino acids in whichmorethan 90% of the amino acidsequenceisconserved between the two viruses. Thereare34 amino acid changes between the subgroup 1 and 2 polypeptides,mostclustered in threeregions of themoleculeatresidues 39through 62, 80 through 122, and 281 through 315.
Rotaviruses,
members of the family Reoviridae, are im-portant causative agents of severe infantilediarrhea
in children andanimals (10, 12). Therotavirusgenome consistsof eleven segments of double-stranded RNA(dsRNA) that areenclosedin adouble-layeredprotein shell typical ofmost
membersof the family Reoviridae(14).However, unlikethe
closely related reoviruses, rotavirus
preparations
derived from infected cells or from diarrheal stool samples alsoincludeparticles with asingle-layeredproteincoat(10, 12). Thesesingle-layered particles have also beentermed"rough particles," "dense particles," and "cores" (1). They are
readily prepared from whole virions by removal of the
proteins of the outer shell, and like reovirus cores, they contain an active virion-associated transcriptase (9). The
single-layered particles, therefore, are most conveniently referred to as cores. The modified single-shelled rotavirus
particles prepared by treatment with calcium chloride and
termed cores (1) are better described as spikeless cores
since, like alkali-treated reovirus cores (30), they lackboth
themajor peripheral protein anddetectable RNA
transcrip-taseactivity (1).
Thepresenceof bothcoresandwhole virionsinrotavirus preparations used for initial serological studies created
un-certainty concerningtheidentity of the antigenically
impor-tant rotavirus surface proteins. The two sets of
surfTace
proteins on cores and virions are distinct (10, 12) and, perhaps predictably, induce different antibodies when a
mixture ofparticles is used to prime the immune system.
Consequently, the serological classification of rotaviruses
recently has required modification (16, 27): separate sub-group and serotypic antigens havebeendefined (16, 27, 28, 32, 33) which are located on the core and whole virion, respectively.
There are at least four serotypes of human rotaviruses (33), theserotypicdeterminantbeingVP7, a
35,500-molecu-lar-weight glycoprotein that is present among the outer
proteins oftheintact virion(10, 12). VP7 has been identified asthe translation product ofgenomicsegment 9 (4, 15), and theprimaryamino acid sequence of simian 11 VP7 has been
*Correspondingauthor.
97
deduced fromthe nucleic acidsequence ofa clonedcDNA copy ofthe dsRNA segment(4).
Lessbiochemicalinformationisavailablefor thesubgroup determinant, whichhasbeenidentified bygenetic studiesas VP6(15), a42,000-molecular-weight non-glycosylated poly-peptide that is coded for by genomic segment 6. Two
rotavirus subgroups havebeenidentified, principallyonthe
basis of the immune adsorption hemagglutination test (16)
butalso in a morerefined fashion byuseoftwomonoclonal antibodies (11) capable ofdistinguishing subgroup 1 and 2
strains. Most rotavirus isolates so far examined with these
monoclonal reagents fall into one of these two subgroups.
However, since somemonoclonal antibodiesreactedwithall
viruses studied, the VP6proteins must also share common
epitopes. This conclusion is supported by the ability of polyclonal antiserum that has been raised against a single
rotavirus type to detect most other rotavirus strains. The
exception is therecently defined pararotaviruses (2), which have different nucleic acid sequences and are
serologically
distinct (22).
The availability of cloned DNA copies of rotavirusgenes nowenables therotavirusgroup andsubgroup determinants
to be further characterized. Here we report the complete nucleotidesequenceofthegenomicsegmentcoding theVP6
of simian 11 rotavirus (subgroup 1) and human strain Wa
rotavirus (subgroup 2). The aim ofthis work has been to
identify thoseregions ofthe VP6molecule whicharesimilar inprimary aminoacid sequence(presumablyresponsible for cross-reactivity) and those regions of the polypeptide that
contain amino acid substitutions which could form the
subgroup epitopic determinants.
MATERIALS ANDMETHODS
Virus and viral RNA. Simian 11
(SAl1)
rotavirus wasobtained from I. H. Holmes, Department of
Microbiology,
University of Melbourne, Australia. Human strain Wa
rota-virus was obtained from H. B.
Greenberg,
Laboratory of InfectiousDiseases, NationalInstitutesofHealth,Bethesda,Md. Both viruseswere
propagated
in MA104 cells(26),
andgenomic dsRNAs were extracted from
purified
virions aspreviously described(13, 26).
on November 10, 2019 by guest
http://jvi.asm.org/
(a)
--.
4
0-b
(b)
=:-4
FIG. 1. Strategy ford cloned DNA copy of dsR Wa rotavirus. The numb
102.
The sequences w4mRNA
(OI)
(4,24),by the the dideoxy method afteBamHI
site of phageml:
Molecular cloning. S were applied to the m derived from each viru N. Ikegami, A. Nom4
publication). A library 13) was screened forrn
copy of genomic segm
from segment 6dsRNY amide gel electrophore in this fashion were cc
by using a modified N Nucleic acidsequenci the two cDNA clones three methods
descrit
strategy used is outline
the SAl clone was strands of the
moleculc
dent methods. The stdetermined, principall region between residi Maxam-Gilbert
sequer
residues commencing
sequence found fort
region of the sequence on the messenger RN from Wa cores
(4,
19).Nucleotide sequence
sents the completeseqi of SAl rotavirus
tol
found for human strain
the correct number of'
._.,.__ _ found in the Wa clone must have resultedfrom an in vitro
mutation which presumably occurred during reverse tran-+-u *-E 4-~-U
scription
of the cDNA. Point mutations of thistypehavealso been found to occur occasionally duringthe in vitro synthe-sis of influenza virus cDNA (7). The two nucleic acid_
812
8 sequences are 78% homologous with base changes locatedthroughout the molecule. However, 91%of the amino acid sequence is conserved because many of the base substitu-tions in the nucleic acid sequence are located in the third position of the altered codons.
Both genomic segments possess a 5'-terminaluntranslated
region of 23 nucleotides that precedes an initiator codon at residues 24 through 26, resulting in a single open reading frame that codes for a polypeptide of397 amino acids. The other two reading frames contain multiple termination co-dons, the longest product (28 aminoacids) being initiated at
_
, . residue 28,the site ofanATG,
which would beonly
aweak initiator (17). The single termination codon at nucleotides1215
through
1217 is followed by a3'-terminal
untranslated8 12 region of 142 nucleotides. The termini of both genomic segments exhibit conserved nucleotide sequences at the 5'-etermining the nucleotide sequence of the (5' GGCTTT 3') and 3'-(5' ATGTGACC 3') ends of the NA genome segment 6 of (a)SAl and (b) molecules. These sequences have also been found for other ers refer to the distance in nucleotides x
ere determined by copying cDNA from
rotavirus
genomic segments (3, 4, 6, 14), indicating thatfor Maxam and Gilbert method (U) (20), or by genomic segment 6, the cloningprotocols used have yielded r subcloningSaou3Al
fragments into thefull-length
copies
ofboth viral RNAs.3mp8
(0) (21, 24). The information presented in Fig. 2 also enables the dsRNA segments to be searched for the consensus sequence that surrounds the translational initiation site on themessen-tandard molecularcloning techniques ger RNAs of eucaryotes (17). For the ATG thatprecedes the ixture of eleven segments of dsRNA open reading frame, a purine at position -3 has been
Is(3, 5, 13; Y. Okada, M. Richardson, conserved for both rotavirus mRNAswith an A-- G change
oto, and Y. Furuichi, submitted for found between the simian and human sequences. Both of rotavirus genomiccDNAclones (3, sequences contain aG residue at position +4 downstream ecombinant plasmids thatcontained a from the AUG. Neither sequence contains the 3'-terminal
lent
6 by using a cDNA probe copied AAUAAA polyadenylation signal (23) as anticipated from that had been purified bypolyacryl-
the absence of polyadenylate on the messengerRNA of the sis (13, 26). Selected clones obtained Reoviridae (14).nfirmed as being copies of segment 6 Comparison of the amino acid sequences of the subgroup
orthern hybridization procedure (26). specific genes. Since SAl and Wa rotaviruses are
repre-ing.
Complete nucleotide sequences of sentative members of the two major rotavirus subgroupswere determined by a combination of (11), it is of interest to compare the amino acid sequences
)ed previously (20, 21, 24, 25). The inferred from the
two
nucleic acid sequences. The 34 amino d in Fig. 1. The complete sequence of acid substitutions found for the Wa strain are presented in established first, either from both Fig. 3 in conjunction with the hydrophobicity profile for the or by a combination of two indepen-SAll
protein that has beencomputed by themethodofKyte equence of the Wa clone was then and Doolittle (18). The amino acidsubstitutionsfound for the y on one strand except for a small inferred Wa protein sequence do not generate substantial ues 750 through 850 in which the changes in the overallhydrophobicity of the molecule, since ice on both strands indicated four T the hydrophobicity plot for the Wa strain is similar to that at position 810 rather than the(T)5
shown in Fig. 3 for SAl (data not shown). Many of the -he SAl sequence. Therefore, this amino acid changes found between the two subgroup pro-was also checked by primer extension teins are conservative: forexample,the Asp - GluchangesA prepared by in vitro transcription (at positions 2, 45, 86, 114 and369) and Ile= Valchanges (at positions 39, 56, 89, 92, 109, 281, 385). Thebasic amino acids Arg, Lys, and His are completely conserved between the RESULTS two sequences. With regard to the location of the amino acid for genomic segment 6. Figure 2 pre-
changes,
ca. 70% of the changes are clustered in three uence of the cloned copy of segment 6regions
ofthemolecule at residues 39through62, 80 throughgether with the nucleotide changes
122,
and 281through
315. The significance of the locationWa rotavirus. Ambiguity concerning and nature of the amino acid changes is of interest as
T residues in the Wa clone at residues discussed below.
810 through 815 was resolved by results obtained whenthis region was also sequenced by primer extension on the
messenger RNA. The sequence determined in this manner yielded the sequence (T)5, indicating that the (T)4 sequence
DISCUSSION
Definitive information on the structural location of the subgroup protein VP6 is not available, but its removal by
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.65.300.65.275.2]100
A G G T G A C A T G T T A A T T C
GGCTTTTAAA CGAAGTCTTC AAC ATG GAT GTC CTA TAC TCT TTG TCA AAG ACT CTT AAA GAC GCT AGA GAC AAA ATT GTC GAA GGC ACT TTG TAT TCT MC GTG AGT GAT
M D V L Y S L S K T L K D A R D K I V E G T L Y S N V S D
E
200
T G C G A T G T A T G T C C G T T
CTA ATT CM CAA TTT MT CAA ATG ATA ATT ACT ATG AAT GGA AAT GAA TTT CM ACT GGA GGA ATC GGT AAT TTG CCA ATT AGA MC TGG MT TTT MT TTC GGG
L I Q Q F N Q M I I T M N G N E F Q T G G I G N L P I R N W N F N F G
V D V T D
300
C TA T A C TT G T G AT C T A T A T C A T T T A
TTA CTT GGA ACA ACT TTG CTGMC TTA GAC GCT AAT TAT GTT GAA ACG GCA AGA MT ACA ATT GAT TAT TTC GTG GAT TTT GTA GAC MT GTA TGC ATG GAT GAG
L L G T T L L N L D A N Y V E T A R N T I D Y F V D F V D N V C M D E
N T I E I I
400
CA T A T G A G A T A G T G T A G G GA T G C A A
ATG GTT AGA GM TCA CAA AGGMC GGA ATT GCA CCT CM TCA GAC TCG CTA AGA MG CTG TCA GCC ATT AAA TTC AAA AGA ATA AAT TTT GAT MT TCG TCG GAA
M V R E SQ R N G I A P Q S D S L R K L S A I K F K R I N F D N S S E
A V E A A G
500
T C A C C C A T GT T A A C T T T T
TAC ATA GM MC TGG MT TTG CM MT AGA AGA CAG AGG ACA GGT TTC ACT TTT CAT AAA CCAMC ATT TTT CCT TAT TCA GCA TCA TAT ACA CTA MAT AGA TCA
Y I E N W N L Q N R R Q R T G F T F H K P N I F P Y S A S Y T L N R S
V F
600
A ATG A A C C T T T A G C C A T G C
CM CCC GCT CAT GAT AAT TTG ATG GGC ACA ATG TGG TTAMC GCA GGA TCG GM ATT CM GTC GCT GGA TTT GAC TAC TCA TGT GCT ATT MC GCA CCA GCC AAT
Q P A H D N L M G T M W L N A G S E I Q V A G F D Y S C A I N A P A N
M
700
G A C C AG T A G C C CA C G T T A T T T A G A A T
ATA CAA CM TTT GAG CAT ATT GTG CCA CTC CGA AGA GTG TTA ACT ACA GCT ACG ATA ACT CTT CTA CCA GAC GCG GAA AGG TTT AGT TTT CCA AGA GTG ATC AAT
I Q Q F E H I V P L R R V L T T A T I T L L P D A E R F S F P R V I N
Q A
800
T C G C T T T A A A C T A G A A T G A T T T A r,
TCA GCT GAC GGG GCA ACT ACA TGG TTT TTC AAC CCA GTG ATT CTC AGG CCG AATAAC GTT GAA GTG GAG TTT CTA TTG MT GGA CAG ATA ATA AAC ACT TAT CAA
S A D G A T T W F F N P V I L R P N N V E V E F L L N G Q I I N T Y Q
900
T T A C A G A C T T T T G C T T T T T AAT C G T A
GCA AGA TTT GGA ACT ATC GTA GCT AGAAAT TTT GAT ACT ATT AGA CTA TCA TTC CAG TTA ATG AGA CCA CCA AAC ATG ACA CCA GCA GTA GCA GTA CTA TTC CCG
A R F G T! V A R N F D T I R L S F Q L M R P P N M T P A V A V L F P
A L N A
1000
C A A T T C G C T C T TAC T A G A A G T G C A G A
AAT GCA CAG CCA TTC GAA CAT CAT GCA ACA GTG GGA TTG ACA CTT AGA ATT GAG TCT GCA GTT TGT GAG TCT GTA CTC GCC GAT GCA AGT GAA ACT CTA TTG GCA
N A Q P F E H H A T V G L T L R I E S A V C E S V L A D A S E T L L A
Q Q N
1100
G C GG G C T A T C G A C T A T T C G A A A
AAT GTA ACA TCC GTT AGG CAA GAG TAC GCA ATA CCA GTT GGA CCA GTC TTT CCA CCA GGT ATG AAC TGG ACT GAT TTA ATC ACC AAT TAT TCA CCG TCT AGG GAG
N V T S V R Q E Y A I P V G P V F P P G M N W T D L I T N Y S P S R E
A E
1200
T C G T C G A T G G GA G C TC TC A T C C
GAC AAT TTG CAA CGC ATA TTT ACA GTG GCT TCC ATT AGA AGC ATG CTC ATT AM TGAGGA CCAAGCTAAC AACTTGGTAT CCAACTTTGG TGAGTATGTA GCTATATCAA
D N L Q R I F T V A S I R S M L I K
V
1300
TCA CAG TC CAT A T T ATG GT CTGTCT GA A G
GCTGTTTGM CTCTGTMGT AAGGATGCGT ATACGCATTC GCTACACAGA GTAATCACTC AGATGGTATA GTGAGAGGAT GTGACC
FIG. 2. Nucleotide sequence and inferred amino acid sequences ofSAl and Wa rotavirus genomic segments6. Thecomplete nucleotide
and amino acid sequences of the coding strand of theclonedSAl segment arepresented togetherwithnucleotide andamino acid changes
found in the corresponding Wa sequence.
high concentrations of calcium chloride (1), accessibility to Only 34 amino acid changes are found when the inferred iodination (P. Gunn and A. R. Bellamy, unpublished data), amino acid sequences of VP6 proteins of SAl and Wa and marked immunogenicity (10, 12) suggest that this protein rotaviruses are compared. Major amino acid sequence con-probably constitutes the peripheral spikes of the icosahedral servation between VP6 proteins of rotaviruses of the two
rotavirus core. Although immunogenic, VP6 must be almost different subgroups is consistent with the known ability of completely masked in the virion because little or no VP6- antibody primed by one rotavirus to detect most other specific antibodies were formed when rabbits were chal-
rotaviruses
(12), with the exception of the newly defined lenged with intact human, simian, or bovine rotaviruses(5a).
pararotaviruses (2), which are distinct (22). Thegroup-The minimal changes in protein sequence found here for specific determinants that enable rotavirus VP6 proteins to the two VP6 proteins implies that variation in this major cross-react probably are located on the regions of the structural protein may be limited by the constraints imposed molecule in which the amino acid sequence is extensively
by the conformational requirements of the rotavirus core. conserved. Incontrast, thoseepitopes that enablesubgroups
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.59.556.73.558.2]._
2
X:
.2
Z220 240 20O 3U30 34396J 3O0
Amino
Acid Number
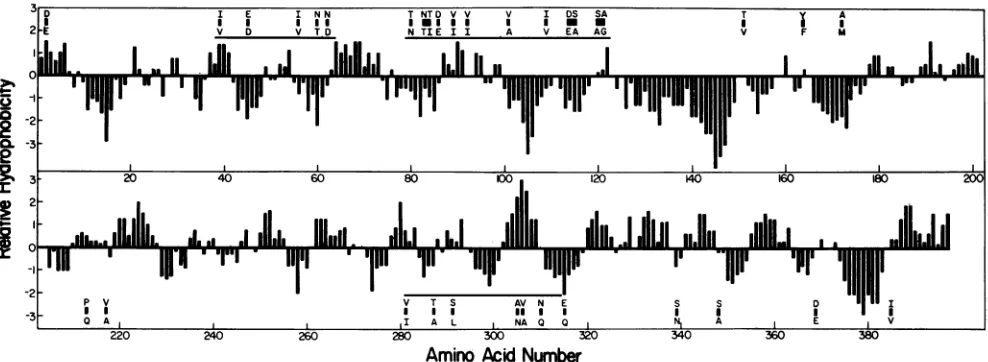
FIG. 3. Amino acid substitutions found between subgroup 1 (SAil) and subgroup 2 (Wa) rotaviruses. The amino acid changes are
indicatedby solidsquaresandarelocatedon ahydrophobicity plot of the SAil polypeptide. The SAil amino acidpresentateach position is shownabove thesquareand theamino acidpresentintheWasequenceis shown belowthesquare.Therelativehydrophobicity indexwas
generated by computer, using the method of KyteandDoolittle(18). Analysis of the Wasequence by the samemethodyieldedasimilar
hydrophobicity profile. Horizontal bars indicate the three putative antigenic regions which wouldencompass morethan 70% of theamino acid changes found between the twopolypeptides.
1 and 2 rotaviruses to be distinguished probably reside in threeregions (Fig. 3,horizontal bars) which contain mostof the amino acid changes.
In theabsenceofanyinformationonthetertiarystructure ofVP6, itobviously isnotpossibletoidentifyprecisely the regions of the molecule that constitute the subgroup
epi-topes: widely separated regions of the molecule could be brought into close proximity by folding of thepolypeptides,
as has been shown elegantly for the influenzavirus
hemag-glutinin (31). However, by analogy with the amino acid changes found to occur among H3 influenza viruses (8, 29, 31), the best studied system involving variation in a viral
antigen, the amino acid differencesatresidues 62(Asn-Asp), 213 (Pro-Gln), and 315 (Glu-Gln) could contribute to the antigenic differences between the rotavirus subgroups. Oth-er changes, for example, those located at residues 84 and 172, which arealso likelytohave surface locations in VP6 (Fig. 3) could also contribute totheantigenic determinants. Since most of the amino acid changes found between subgroup 1 and 2 proteins are clustered in a fewrelatively restrictedregions of the molecule, peptide synthesis should provideafeasiblemethod by which thenatureof the epitopic determinantscould be furtherinvestigated. The synthesis of peptides corresponding to putative epitopic regions should enable thegeneration of site-specific antibodiesthatcouldbe
tested for subgroup specificity. Alternatively, blocking
ex-periments could be carried out with selected synthetic peptides and the subgroup-specific monoclonal reagents
already developed (11). Either of these two experimental approaches should in the future enable the major subgroup-specific epitopes to be unambiguously located on the VP6
molecule.
ACKNOWLEDGMENTS
Wethank Jeanette StreetandMark Lin forassistanceinscreening theSA-11androtavirus(Wa)cDNA libraries forgenomic6 clones.
Thisworkwassupportedinpartbygrantsfrom the NewZealand MedicalResearchCouncil and theWorld HealthOrganization.
LITERATURE CITED
1. Bican, P., J. Cohen, A. Charpilienne, and R. Scherrer. 1982. Purification and characterization ofbovine rotaviruscores. J.
Virol.43:1113-1117.
2. Bohl, E. H., L. J. Saif, K. W. Theil, A. G. Agnes, and R. F.
Cross. 1982. Porcine pararotavirus: detection, differentiation from rotavirus, and pathogenesis in gnotobiotic pigs. J. Clin. Microbiol. 15:312-319.
3. Both, G. W., A. R. Bellamy, J. E.Street, and L. J. Siegman. 1982. A general strategy for cloning double-stranded RNA: nucleotide sequence of the simian-11 rotavirusgene8. Nucleic Acids Res. 10:7075-7088.
4. Both, G. W., J. S. Mattick, and A. R. Bellamy. 1983. The serotype-specific glycoprotein of simian-11 rotavirus: coding assignmentandgene sequence. Proc. Natl. Acad. Sci. U.S.A. 80:3091-3095.
5. Both, G. W., J. S. Mattick, L. J. Siegman, P. H. Atkinson, S. Weiss, A. R. Bellamy, J. E. Street, and P. Metcalf. 1983. Cloning ofSAil rotavirusgenes:genestructureandpolypeptide assign-mentfor the type-specific glycoprotein, p. 73-82. In D. H. L. Bishop and R. W. Compans (ed.), Thedouble-stranded RNA Viruses.Elsevier-Biomedical, New York.
Sa.Both, G. W., L. J. Siegman,P. H.Atkinson,M. S.Poruchynsky, A.Kabcenell, J. E. Street,P.R.Gunn,F.Sato,K. F.H.Powell, and A. R. Bellamy. 1984. Analysis ofcloned genes encoding rotavirustype-specific neutralising antigens, p.53-58. InR. A.
Lernerand R. M. Chanock(ed.), Modern approaches to vac-cines: molecular and chemical basis of virus virulence and immunogenicity. Cold SpringHarborLaboratory, ColdSpring Harbor,N.Y.
6. Both,G.W.,L.J. Siegman,A.R.Bellamy,and P. H. Atkinson.
1983. Coding assignment and nucleotide sequence of simian rotavirus SAil genesegment10: location ofglycosylationsites suggeststhat thesignalpeptideisnotcleaved. J. Virol. 48:335-339.
7. Both, G. W., and M. J. Sleigh. 1980. Complete nucleotide
sequenceof the haemagglutinin gene froma human influenza virus of the Hong Kong subtype. Nucleic Acids Res. 8:2561-2575.
8. Both, G.W.,M.J. Sleigh,N.J. Cox,and A.P. Kendal. 1983. Antigenicdriftin influenza virus H3hemagglutininfrom1968to
1980: multiple evolutionary pathwaysandsequentialamino acid changesatkeyantigenic sites.J. Virol. 48:52-60.
D I E I NN TNTD V V v DS SA T
A-3 I2040 60 80 0 120 140 60 180 200
-I
*2
3-2 _1 ,0 1_
4
060l
T0
1'''
120il'
1§I
140
l
16
180lil
-3L 7A A° T AVN E s s D Vl
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.67.561.75.256.2]9. Cohen, J. 1977.Ribonucleic acidpolymeraseactivityassociated with purified calf rotavirus. J. Gen. Virol. 36:395-402. 10. Estes, M. K.,E. L.Palmer, andJ. F.Obijeski. 1983.
Rotavir-uses: areview. Curr. Top.Microbiol. Immunol. 105:123-184. 11. Greenberg, H. B., V. McAuliffe, J. Valdesuso, R. Wyatt, J.
Flores, A.Kalica, Y. Hoshino,and N. Singh. 1983. Serological
analysis of the subgroupprotein ofrotavirus, using monoclonal antibodies.Infect. Immun. 39:91-99.
12. Holmes, I. H. 1983. Rotaviruses, p. 359-423. In W. K. Joklik (ed.), TheReoviridae. PlenumPublishingCorp.,NewYork.
13. Imai, M., M. A. Richardson, N. Ikegami, A. J. Shatkin, and Y.
Furuichi. 1983. Molecular cloning of double-stranded RNA virus genomes. Proc. Natl. Acad. Sci. U.S.A. 80:373-377.
14. Joklik,W. K.1983. The membersof thefamily Reoviridae, p. 1-78. In W. K. Joklik (ed.), The Reoviridae. Plenum Publishing Corp., NewYork.
15. Kalica,A.R.,H. B.Greenberg,R.G. Wyatt, J. Flores,M. M.
Sereno, A. Z. Kapikian, and R. M. Chanock. 1981. Genes of human(strain Wa) and bovine (strain UK) rotaviruses that code forneutralization and subgroup antigens.Virology 112:385-390.
16. Kapikian, A.Z., W. L. Cline, H. B. Greenberg, R. G. Wyatt,
A. R.Kalica, C.E.Banks,H. D.James, Jr., J.Flores,and R. M. Chanock. 1981.Antigenic characterization of human and animal rotaviruses by immune adherence hemagglutination assay (IAHA): evidence for distinctness of IAHA and neutralization antigens. Infect. Immun. 33:415-425.
17. Kozak,M.1981.Possible role offlanking nucleotides in recogni-tion of the AUG initiator codon by eukaryotic ribosomes. Nucleic AcidsRes. 9:5233-5252.
18. Kyte, J.,and R. F.Doolittle.1982.Asingle method for display-ing the hydropathic character of a protein. J. Mol. Biol. 157:105-132.
19. Mason, B.B., D. Y.Graham, andM. K. Estes. 1980. Invitro
transcription and translation of simian rotavirus SA1l gene products. J.Virol. 33:1111-1121.
20. Maxam, A. M., and W. Gilbert. 1977. A new method for
sequencingDNA. Proc.Natl.Acad. Sci. U.S.A. 74:560-564. 21. Messing, J., R. Crea, and P. H. Seeburg. 1981. A system for
shotgunDNAsequencing. Nucleic AcidsRes. 9:309-321. 22. Pedley,S., J.C.Bridger, J.F.Brown,and M. A.McCrae. 1983.
Molecular characterization of rotaviruses with distinct group antigens. J. Gen. Virol. 64:2093-2101.
23. Proudfoot,N. 1982. The end of themessage. Nature (London)
298:516-517.
24. Sanger, F., S. Nicklen, and A. R.Coulson. 1977. DNA sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74:5463-5467.
25. Sleigh,M.J., G. W. Both, P. A.Underwood, andV.J. Bender. 1971. Antigenic drift in the hemagglutinin of the Hong Kong influenza subtype: correlation of amino acid changes with alterations in viral antigenicity. J. Virol. 37:845-853.
26. Street, J. E., M. C. Croxson, W. F. Chadderton, and A. R.
Bellamy. 1982. Sequence diversity of human rotavirus strains investigated by Northern blot hybridization analysis. J. Virol
43:369-378.
27. Thouless, M. E., G. M. Beards, and T. H. Flewett. 1982.
Serotying and subgrouping of rotavirus strains by the ELISA test. Arch. Virol. 73:219-230.
28. Urasawa, S., T. Urasawa, and K.Taniguchi. 1982. Three human rotavirus serotypes demonstrated by plaque neutralization of isolated strains. Infect. Immun. 38:781-784.
29. Webster, R. W., G. Laver, G. M. Air,and G. C. Schild. 1982. Molecular mechanisms of variation ininfluenza viruses. Nature (London)296:115-121.
30. White, C. K., andH.J. Zweerink. 1976.Studiesonthestructure
of reovirus cores: selective removal ofpolypeptide A2. Virology 70:171-180.
31. Wiley, D. C.,I. A. Wilson, and J. J. Skehel. 1981. Structural identification oftheantibody-binding sites of Hong Kong
influ-enzahemagglutinin and their involvement inantigenic variation.
Nature(London) 289:373-378.
32. Wyatt, R. G., H. B. Greenberg, W. D.James, A. L. Pittman,
A. R. Kalica, J. Flores, R. M. Chanock, and A. Z. Kapikian. 1982. Definition of human rotavirus serotypes by plaque reduc-tion assay.Infect. Immun. 37:110-115.
33. Wyatt,R.G.,H. D.James, Jr.,A. L.Pittman, Y. Hoshino, H. B. Greenberg, A. R. Kalica, J. Flores, and A. Z. Kapikian. 1983. Direct isolation in cell culture ofhuman rotaviruses and their characterization into four serotypes.J.Clin.Microbiol.
18:310-317.