Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Rotavirus RNA Replication Requires a Single-Stranded
3
9
End for Efficient Minus-Strand Synthesis
DAYUE CHENANDJOHN T. PATTON*
Laboratory of Infectious Diseases, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Bethesda, Maryland 20892
Received 19 March 1998/Accepted 22 May 1998
The segmented double-stranded (ds) RNA genome of the rotaviruses is replicated asymmetrically, with viral mRNA serving as the template for the synthesis of minus-strand RNA. Previous studies with cell-free repli-cation systems have shown that the highly conserved termini of rotavirus gene 8 and 9 mRNAs contain cis-acting signals that promote the synthesis of dsRNA. Based on the location of the cis-acting signals and computer modeling of their secondary structure, the ends of the gene 8 or 9 mRNAs are proposed to interact in cis to form a modified panhandle structure that promotes the synthesis of dsRNA. In this structure, the last 11 to 12 nucleotides of the RNA, including the cis-acting signal that is essential for RNA replication, extend as a single-stranded tail from the panhandled region, and the 5*untranslated region folds to form a stem-loop motif. To understand the importance of the predicted secondary structure in minus-strand synthesis, muta-tions were introduced into viral RNAs which affected the 3*tail and the 5*stem-loop. Analysis of the RNAs with a cell-free replication system showed that, in contrast to mutations which altered the structure of the 5*
stem-loop, mutations which caused complete or near-complete complementarity between the 5*end and the 3*
tail significantly inhibited (>10-fold) minus-strand synthesis. Likewise, incubation of wild-type RNAs with oligonucleotides which were complementary to the 3* tail inhibited replication. Despite their replication-defective phenotype, mutant RNAs with complementary 5*and 3*termini were shown to competitively interfere with the replication of wild-type mRNA and to bind the viral RNA polymerase VP1 as efficiently as wild-type RNA. These results indicate that the single-strand nature of the 3* end of rotavirus mRNA is essential for efficient dsRNA synthesis and that the specific binding of the RNA polymerase to the mRNA template is required but not sufficient for the synthesis of minus-strand RNA.
Rotaviruses, members of the family Reoviridae, are the ma-jor cause of acute dehydrating diarrhea in infants and young children (16). The rotavirus genome consists of 11 segments of double-stranded (ds) RNA and is contained in the viral core along with the minor structural proteins, VP1 and VP3 (10). These two proteins function as the RNA-dependent RNA polymerase (7, 41) and guanylyltransferase (31), respectively, of the virus. The shell of the core is made up of 60 dimers of VP2 arranged as a T51 icosahedral lattice (20). VP2 has non-specific RNA-binding activity (18) and directly interacts with dsRNA lining the VP2 shell (33). Cores which are purified from virions and then disrupted by incubation under hypotonic conditions (open cores) have been shown to possess replicase activity which catalyzes the synthesis of dsRNA from rotavirus mRNA (5). Analysis of recombinant proteins in cell-free rep-lication systems has demonstrated that VP1 and VP2, when combined, have replicase activity and are able to catalyze the synthesis of dsRNA (30, 46).
Rotavirus mRNAs are largely monocistronic, possessing 59 cap structures but lacking 39 poly(A) tails (8). Cores sur-rounded by a protein matrix consisting of 260 trimers of VP6 are referred to as double-shelled particles (32) and have an associated transcriptase activity which directs the synthesis of viral mRNA (3, 6). The fact that transcriptase activity is asso-ciated with double-shelled particles, and not with cores, indi-cates that VP6 may play an important role in mRNA synthesis.
Infectious virions (triple-shelled particles) consist of double-shelled particles which are covered by a shell of protein com-posed of 780 molecules of the glycoprotein VP7 and 120 mol-ecules of the spike protein VP4 (38).
Among the 11 segments of dsRNA that make up the viral genome, conserved sequences are present only at the 59and 39 termini. The consensus sequence at the 59terminus is 59-GGC-poly(A/U), and that at the 39terminus is 59-aUgugaCC-3,9with fully conserved residues shown in uppercase letters (8). Se-quence analyses of different strains of rotavirus have shown that the entire 59and 39untranslated regions (UTRs) of ho-mologous genome segments are highly conserved, sometimes more so than the nucleotide sequence of the open reading frame of the segments (8). The functions of the conserved termini of the viral RNAs are not entirely clear, but these regions may contain cis-acting signals which are important for replication, transcription, and packaging. Indeed, recent work with cell-free replication systems has shown that within the conserved sequence at the 39end of viral mRNA is a cis-acting signal which is essential for promoting minus-strand synthesis, i.e., the minimal essential promoter (29, 43, 44). Analysis of mutant viral RNAs in vitro has also revealed that rotavirus mRNAs contain two other signals that, although not essential, enhance minus-strand synthesis (29, 44). One of these en-hancement signals is located immediately upstream of the min-imal essential promoter at the 39end of the mRNA, while the other is located at the 59 end of the mRNA. Because the enhancement signals are located in regions of the viral mRNA that are not highly conserved, it is possible that secondary structures common to all 11 species of viral RNA, as opposed to the primary nucleotide sequence of the regions, are respon-sible for the enhancement of minus-strand synthesis.
* Corresponding author. Mailing address: Laboratory of Infectious Diseases, National Institute of Allergy and Infectious Diseases, Na-tional Institutes of Health, 7 Center Dr., MSC 0720, Room 117, Be-thesda, MD 20892. Phone: (301) 496-3372. Fax: (301) 496-8312. E-mail: [email protected].
7387
on November 9, 2019 by guest
http://jvi.asm.org/
mRNA to function as an efficient template for minus-strand synthesis but that the strandedness of the minimal essential promoter does not affect the ability of the polymerase to rec-ognize the viral RNA.
MATERIALS AND METHODS
Preparation of open cores.Rotavirus SA11-4F virions were propagated in MA104 cells and purified by CsCl centrifugation (29). The method of Chen et al. (5) was used to prepare open cores from purified virus. The concentration of protein in open core preparations was determined with a Bio-Rad protein assay kit.
Removal of genomic dsRNA from open cores.Open cores (100mg) were treated for 60 min at 30°C with 340 U of micrococcal nuclease (Worthington Biochemical Co., Lakewood, N.J.) per ml in a reaction mixture (500ml) con-taining 10 mM Tris-HCl (pH 8.0), 10 mM NaCl, and 1 mM CaCl2. After complete digestion of dsRNA in the open core preparation was confirmed by electrophoresis on a 1% agarose gel, the nuclease was inactivated by the addition of EGTA to 3 mM.
Construction of templates for T7 transcription.PCR was used as follows to introduce mutations in the gene 8 cDNA and to directly link the gene 8 cDNA to a promoter for T7 RNA polymerase. The plasmid SP65g8R, containing a full-length gene 8 cDNA of simian rotavirus SA11, was digested with BamHI and
HindIII to release the gene 8 cDNA insert (35). The insert was gel purified with
Qiaex II reagents (Qiagen, Valencia, Calif.) and used as the template in ampli-fication reaction mixtures containing 25 U of Pfu DNA polymerase (Stratagene, La Jolla, Calif.) per ml under the following conditions: 94°C at 1 min, 48°C at 1 min, and 72°C at 2 min (30 cycles). The amplified DNA produced by Pfu polymerase is blunt ended and does not contain 39-overhang A residues. To generate the G8-59-B1 mutant DNA under the control of a promoter for T7 RNA polymerase, the plus-sense primer T7G8/59-B1 (59-TAATACGACTCA CTATAGGtcacatAAGCGTCTCAGTCGCCGTTTG-39) and the minus-sense primer G8/39-wt (59-GGTCACATAAGCGCTTTCTATTCT-39) were used for PCR. To produce the G8-59-B2 and G8-59-B4 DNAs, the plus-sense primers T7G8/59-B2 (59-TAATACGACTCACTATAGGCTacatAAGCGTCTCAG TCGCCGTTTG-39) and T7G8/59-B4 (59-TAATACGACTCACTATAGAAG CGTCTCAGTCGCCGTTTG-39), respectively, and the minus-sense primer G8/ 39-wt were used instead. The G8-59/39-B1 and G8-39-B1 DNAs were generated by including the plus-sense primers T7G8/59-B1 and T7G8/59-wt (59-TAATACGA CTCACTATAGGCTTTTAAAGCGTCTCAGTCG-39), respectively, in ampli-fication reactions along with the minus-sense primer G8/39-B1 (59-GgctgtgcAA GCGCTTTCTATTCTTGC-39). Sequences within primers that define the pro-moter for T7 RNA polymerase are underlined, and sequences in the primers that represent mutations of the wild-type gene 8 sequence are shown in lowercase letters. The amplified DNAs were purified by phenol-chloroform extraction and then used in T7 transcription reactions to produce gene 8 RNAs. Unless other-wise noted, the 59and 39ends of the transcripts made from the amplified DNAs are identical in sequence to those of authentic (wild-type) gene 8 mRNA.
The T7 transcription vectors SP65g8R, SP65g8Rd45-543, and pT7g6 were constructed as described before (29, 30). Following linearization with SacII and blunt ending with T4 DNA polymerase, SP65g8R, pT7g6, and SP65g8Rd45-543 were used in transcription reactions to produce, respectively, wild-type gene 8 RNA, wild-type gene 6 RNA, and gene 8 RNA which was wild type in sequence except for lacking residues 45 to 543.
To produce the T7 transcription vector, SP72g8R40, the complementary oli-gonucleotides, 59-aaTGATGATGGCTTAGCAAGAATAGAAAGCGCTTAT GTGACCgcggtgca-39(plus-sense) and 59-ccgcGGTCACATAAGCGCTTTCTA TTCTTGCTAAGCCATCATC (minus-sense), were treated with kinase and annealed, forming a short DNA hybrid with EcoRI and PstI cohesive ends and an internal sequence which is identical to that of the 39-terminal 40 nucleotides of the gene 8 mRNA (29). The annealed oligonucleotides were ligated to SP72
m
assays contained 0.1mg each of reporter RNA and competitor RNA. Reaction mixtures were incubated for 2 h at 32°C. Replication products were resolved by electrophoresis on 12.5% polyacrylamide gels containing sodium dodecyl sulfate (19) and detected by autoradiography. Bands of32P-labeled dsRNA were quan-tified with a Molecular Dynamics PhosphorImager (model 445SI).
Oligonucleotide inhibition assay.Three deoxyoligonucleotides were tested for the ability to inhibit the synthesis of dsRNA in replication assays, G8-WT-59
(59-GGCTTTTAAAGCGTCTCA-39), which corresponds in sequence to the first 18 nucleotides of the gene 8 mRNA; G8-B1-59(59-GGtcacatAAGCGTCT CA-39, mutant residues shown in lowercase letters), which corresponds in se-quence to the first 18 nucleotides of the G8-59-B1 RNA and is fully complemen-tary to the 39-terminal 14 nucleotides of the gene 8 mRNA; and G8-507 (59-T CCTTCTGCAGTTATTGTAGTTTC-39), which is complementary in sequence to nucleotides 484 to 507 of the gene 8 mRNA. In inhibition assays, oligonucle-otides were added to standard reaction mixtures lacking open cores. After incu-bation for 1 h at 32°C, open cores were added and the reaction mixtures were incubated an additional 2 h at 32°C. The dsRNA products were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and quantified with a PhosphorImager.
Electrophoretic mobility shift assay.To generate the RNA probe for the mobility shift assays, SP72g8R40 was first cleaved with SacII and blunt ended with T4 DNA polymerase (5). The32P-labeled RNA probe, SP72-v39-40, was then synthesized from the linearized plasmid with the Ambion MEGAshort transcription system according to the protocol of the manufacturer, except that the concentration of UTP was reduced to one-fourth and 50mCi of [a-32P]UTP (800 Ci/mmol; Amersham) was included per 20ml of reaction mixture. The RNA product was 73 nucleotides in length, with the sequence of the last 40 residues identical to that found at the 39terminus of the gene 8 mRNA. The remaining 33 nucleotides of the probe represent SP72-specific sequences. The RNA probe was gel purified as described elsewhere (27).
Mobility shift assays contained 50 mM Tris-HCl (pH 7.2), 5 mM magnesium acetate, 5 mM dithiothreitol, 1.5% polyethylene glycol 8000, 20 U of RNasin, 1.2 to 10 pmol of32P-labeled probe, 0.5mg of open cores, and the indicated amount of competitor RNA in a final volume of 20ml (27). The average size of the competitor yeast RNA used in the shift assays was 1,000 nt. Reaction mixtures were incubated for 60 min at 32°C. Complexes formed between the probe and VP1 were resolved by electrophoresis at 250 V on nondenaturing 8% polyacryl-amide gels containing 50 mM Tris-glycine, pH 8.8 (17). The gels were analyzed by autoradiography, and band intensities were determined by phosphorimaging. Data interpretation.All experiments described in this study were repeated a minimum of three times, and the results obtained from the repetitions were consistent with one another. Unless otherwise noted, the values provided in the figures are the nonaveraged results of a single experiment and are representative of the values obtained for the other repetitions of the same experiment.
Secondary structure of rotavirus RNAs.The secondary structures of rotavirus RNAs were computed with the mfold program, version 2.3, developed by Zuker and Turner (37, 42, 48, 49) and made available on the home page of Michael Zuker of Washington University, St. Louis, Mo. (http://www.ibc.wustl.edu/
;zuker/).
Nucleotide sequence accession numbers.The GenBank accession numbers of the NSP2 genes analyzed with the mfold program are LO4529 (DS1), X57944 (Hu-SG2), X94562 (IS2), LO4530 (NCDV), X06722 (OSU), AB009625 (PO-13), Z21640 (RF), LO4530 (SA11), LO4533 (Ty-1), J02420 (UK), and LO4534 (Wa). The accession number for OSU gene 9 is X04613.
RESULTS
Predicted secondary structure of the gene 8 mRNA.Previous studies with a number of viruses, including picornaviruses (24), tobamoviruses (12, 21), human immunodeficiency virus type 1
on November 9, 2019 by guest
FIG. 1. The optimal secondary structures of wild-type and mutant SA11 gene 8 RNAs and wild-type OSU gene 9 RNA were predicted based on free-energy minimization with the mfold program. Shown are the portions of the secondary structures illustrating the computed interaction between the 59and 39ends of the RNA. Mutations in the sequences are presented in red, lowercase letters. The conserved residues that are part of the 39essential cis-acting replication signal are shown in blue, and the location of deletions is shown in green. kc/m, kcal/mol.
on November 9, 2019 by guest
http://jvi.asm.org/
(13, 23), and Qb(2), have shown that secondary structures in viral RNAs can play important roles in genome replication, assembly, and translation. As an initial step toward identifying secondary structures in rotavirus plus-strand RNAs that might be involved in the synthesis of minus-strand RNAs, the RNA encoding NSP2 of simian (SA11, gene 8), avian (PO-13 and Ty-1), bovine (NCDV, RF, and UK), human (DS1, Hu-SG2, IS2, and Wa), and porcine (OSU) strains of rotavirus was analyzed with the mfold program (version 2.3) made available by M. Zucker. The mfold program predicts the secondary structures of RNAs based on their minimal computed free energies according to the rules established by Turner and oth-ers (37, 42). All of the 20 most-stable secondary structures calculated for the SA11 gene 8 mRNA predicted that the 59 and 39 ends of the RNA would interact in the manner illus-trated in Fig. 1A. The major features of the structure produced by the interaction of the ends of the RNA are that (i) the 39-terminal 12 residues are non-base paired, including the min-imal essential promoter; (ii) the 59 UTR (residues 1 to 45) forms a stem-loop motif; (iii) the initiation codon (residues 46 to 48) is not base paired and is located immediately down-stream of the stem-loop motif of the 59UTR; and (iv) residues downstream of the initiation codon (residues 49 to 80) and upstream of the non-base-paired 39-terminal stretch (residues 1015 to 1045) interact to form a panhandle that juxtaposes the 59and 39ends of the RNA.
Given that the sequences of the first 75 and last 29 bases of the NSP2 RNA are nearly identical for all group A rotaviruses (28), the NSP2 RNAs of virus strains other than SA11 were also expected to fold such that their 59and 39termini would interact. Indeed, of the 10 most-stable secondary structures computed for each DS1 (10 of 10), Hu-SG2 (10 of 10), NCDV (6 of 10), OSU (10 of 10), RF (7 of 10), UK (5 of 10), and WA (10 of 10), at least one-half showed that the termini of the NSP2 RNAs would interact like that of the SA11 gene 8 RNA (Fig. 1A). Likewise, 9 of the 10 most-stable secondary struc-tures computed for avian virus Ty1 indicated that the termini of its NSP2 RNA would interact in a manner similar, although not identical, to that of the SA11 gene 8 RNA. Unlike the other 9 strains analyzed, only a minority of the most-stable secondary structures predicted for the NSP2 RNAs of IS-2 (1 of 10) and PO-13 (1 of 10) showed that the 59and 39ends of the RNA would interact. However, in terms of computed free energy, only a small difference (,4 kcal/mol) was found be-tween the most-stable structure computed for the NSP2 RNAs of IS-2 and PO-13 and the most-stable structure computed, predicting an interaction between the 59 and 39 termini like that found for SA11 gene 8 RNA.
Previous studies showed that gene 8 RNAs in which the last 12 bases were deleted no longer served as efficient templates for minus-strand synthesis in vitro (29, 44). The mfold program predicts that except for the loss of the 39-terminal single-stranded stretch, deletion of the 12 nucleotides does not alter the secondary structure of the gene 8 RNA (Table 1 and Fig. 1C). This indicates that neither the putative stem-loop struc-ture of the 59UTR nor the panhandled region of the gene 8 RNA alone is sufficient to promote the synthesis of minus-strand RNA. Earlier work also showed that deletion of the 59 UTR did not prevent the gene 8 RNA from serving as an efficient template for minus-strand synthesis in noncompetitive replicase assays (29). However, in competitive replicase assays, where wild-type gene 8 RNA was also present, gene 8 RNAs containing 59deletions were replicated less efficiently, indicat-ing that optimal minus-strand synthesis requires the 59 UTR (29). Thus the putative stem-loop structure formed by the 59 UTR may play an important role in promoting RNA replica-tion.
Structural similarity of SA11 gene 8 and OSU gene 9 RNAs.
Analysis of mutated SA11 gene 8 RNAs in a cell-free replica-tion system has shown that both the 59and 39ends of the RNA contain cis-acting signals that are important for minus-strand synthesis (29). Folding of the gene 8 RNA in a way that allows interaction of the 59and 39termini would provide a mechanism by which the cis-acting signal at the 59 end could be brought close to those at the 39end to form a structure that serves as the promoter for minus-strand synthesis. Besides the gene 8 RNA of SA11, cell-free replication assays have shown that the 59and 39end of the VP7 RNA (gene 9) of OSU also contain
cis-acting signals that enhance minus-strand synthesis (43, 44).
To address the possibility that the OSU gene 9 RNA, like the SA11 gene 8 RNA, might fold such that its 59- and 39-terminal replication signals were close to each other, the mfold program was used to predict the folded structure of the gene 9 RNA. Of the 22 most-stable structures predicted, all showed that the 59 and 39 ends of the OSU gene 9 RNA would interact in a manner nearly identical to that exhibited by the SA11 gene 8 RNA (Fig. 1B). These results support the hypothesis that in-teractions between the termini of these RNAs could lead to the formation of a structure that promotes minus-strand synthesis.
Base pairing of the 3*terminus of the RNA template inhib-its minus-strand synthesis. A key feature of the computed secondary structure for the wild-type gene 8 RNA is that the last 12 bases of the RNA are non-base paired (Fig. 1A). To determine whether synthesis of minus-strand RNA on the gene 8 template RNA requires that this region be single-stranded, the 59-terminal sequence was mutated such that the 59and 39
on November 9, 2019 by guest
[image:4.612.50.550.81.199.2]ends of the gene 8 mRNA were either fully (G8-59-B1) or partially (G8-59-B2) complementary (Table 1). The proposed secondary structure of the mutant RNA, G8-59-B1, predicts that the first and last 14 bases of the RNA will base pair and that the stem-loop of the 59UTR of the wild-type RNA will be lost, with a new stem-loop generated beginning with residue 15 (Fig. 1D). The structure of G8-59-B2 is predicted to be like that of G8-59-B1, except that the third residues from the 59and 39 ends of the RNA cannot base pair (Fig. 1E). The ability of G8-59-B1 and G8-59-B2 to function as templates for dsRNA synthesis was evaluated in a cell-free replication system con-taining SA11 open cores. The results showed that compared with assays containing wild-type gene 8 RNA, assays contain-ing G8-59-B1 and G8-59-B2 synthesized approximately 100-and 10-fold-less dsRNA, respectively (Fig. 2 100-and Table 2).
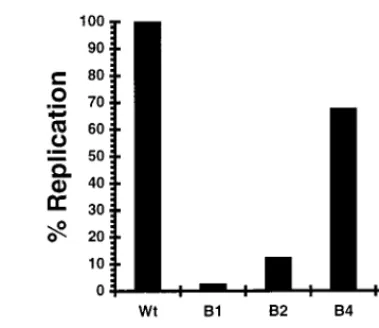
To examine the possibility that the 59B1 and 59B2 RNAs replicated poorly because they lacked the 59stem-loop of the wild-type RNA (and not because of base pairing between the 59and 39termini), two mutant RNAs (G8-59-B4 and G8-59-B6) were prepared which also lacked the wild-type 59 stem-loop but, unlike the B1 and B2 RNAs, did possess single-stranded 39 tails. In comparison to the wild-type RNA, the G8-59-B4 RNA
contains a deletion of residues 2 to 8, and the G8-59-B6 RNA contains the sequence G15AGUCG instead of C15UCAGU
(Table 1). The G8-59-B4 RNA is predicted to fold such that it has the same 59stem-loop structure as the 59-B1 and G8-59-B2 RNAs, but because of the deletion lacks the residues which can base pair with the 39-terminal eight nucleotides of the RNA (Fig. 1F). Except for lacking the wild-type 59 stem-loop, in all other respects, the predicted secondary structure of G8-59-B6 is like that of the wild-type RNA (Fig. 1G). By assay in the cell-free replication system, both G8-59-B4 and G8-59-B6 were shown to replicate at levels 60 to 70% those of wild-type RNA, and thus the synthesis of dsRNA was only slightly de-creased (,twofold) by the absence of the wild-type 59 stem-loop (Fig. 3). Considered together, these data indicate that the 100-fold and 10-fold decreases seen in replication with G8-59-B1 and G8-59-B2, respectively, stemmed not from the lack of the wild-type 59 stem-loop but, rather, from base pairing involving the 39-terminal 12 nucleotides of these RNAs. The fact that G8-59-B2 replicated significantly better than G8-59-B1 suggests that the presence of even a single non-base-paired nucleotide at the 39 terminus increases the replication effi-ciency of the RNA template.
[image:5.612.331.521.68.232.2]The mutant G8-59/39-B1 was constructed to determine if mutations could be introduced into the 39 terminus which could compensate for the inhibitory effects of mutations at the 59 end of G8-59-B1 on replication. The sequence of the 59 terminus of G8-59/39-B1 is the same as that of G8-59-B1, but residues 3 to 8 from the 39 end of G8-59/39-B1 are such that they are not complementary to residues 3 to 8 from the 59end of the RNA (Table 1). As a result of this sequence difference, the proposed secondary structure for G8-59/39-B1 indicates that the 59terminus of the RNA is not base paired to the 39tail and that 5 of the last 12 nucleotides of the RNA are not base paired (Fig. 1H). Since previous work has shown that muta-tions of the 39-terminal seven nucleotides of the gene 8 mRNA can significantly reduce its ability to function as a template (29), we also constructed the mutant G8-39-B1 as a control for the analysis of G8-59/39-B1 (Table 1 and Fig. 1I). The results showed that replication assays containing G8-59/39-B1 pro-duced approximately eight times as much dsRNA product as assays containing G8-59-B1 (Fig. 4). Hence, the introduction of FIG. 2. Synthesis of dsRNAs from wild-type and mutant template RNAs by
[image:5.612.97.247.74.306.2]the cell-free replication system. Reaction mixtures contained micrococcal nucle-ase-treated open cores, [32P]UTP, and either 0.1mg of the indicated template RNA or no RNA (Without). The32P-labeled dsRNA products were resolved by electrophoresis on a 12.5% polyacrylamide gel and detected by autoradiography.
FIG. 3. Effect of mutagenesis at the 59end of the gene 8 mRNA on synthesis of dsRNA in vitro. Wild-type RNA (Wt) and the mutant gene 8 mRNAs G8-59-B1, -B2, -B4, and -B6 were used as templates in the cell-free replication system. The dsRNA products of the reaction mixtures were resolved by gel electrophoresis and quantified with a PhosphorImager. The relative values of dsRNA are shown with the value for the wild type normalized to 100%.
TABLE 2. Effect of 59mutation on the level of dsRNA synthesis
Template RNA
% dsRNA synthesizeda
Expt 1 Expt 2 Expt 3
G8-wt 100 100 100
G8-59-B1 0.9 0.3 0.5
G8-59-B2 9.1 4.6 7.8
aPercentage of dsRNA product5(integrated intensity of the band of dsRNA product/integrated intensity of the dsRNA product for G8-wt)3100. Results are given for three independent experiments.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.49.290.638.702.2]mutations into the 39 terminus of G8-59-B1, which decreased the extent of base pairing between the 59and 39ends of the RNA, increased its efficiency as a template for replication. Assays containing the control RNA G8-39-B1 produced two to three times more dsRNA than assays containing G8-59/39-B1 but approximately 10-fold less product than assays containing wild-type gene 8 mRNA (Fig. 4). The inefficiency of replication of G8-39-B1 in comparison to wild-type gene 8 mRNA is consis-tent with earlier findings showing that the 39-terminal eight nucleotides form the essential cis-acting replication signal of the RNA and that mutations introduced in this region can significantly inhibit replication of the RNA (29, 44).
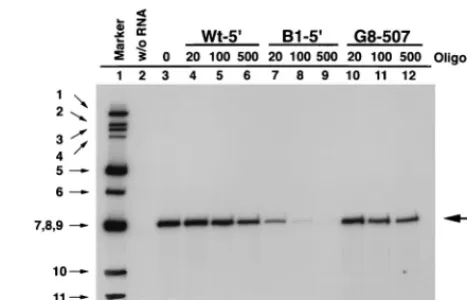
Inhibition of RNA replication by complementary oligonucleo-tides.The change of six nucleotides at the 59end of wild-type gene 8 RNA hypothetically allows the ends of G8-59-B1 to fully anneal and thus form a “closed” panhandle structure (Fig. 1D). The fact that G8-59-B1 replicates poorly suggests that the pan-handle structure of the wild-type RNA must include a single-stranded 39terminus in order for the viral RNA polymerase to efficiently replicate the RNA. If this is the case, annealing of a complementary oligodeoxynucleotide to the 39terminus of the RNA would be expected to inhibit replication. To test this pos-sibility, the Wt-59, B1-59, and G8-507 oligonucleotides were syn-thesized. The sequence of the Wt-59oligonucleotide is identi-cal to the first 18 residues of the wild-type gene 8 RNA, while that of the B1-59oligonucleotide is identical to the first 18 res-idues of the mutant G8-59-B1 RNA and therefore has the potential to form a complete duplex with the 39terminus of the wild-type RNA. The control oligonucleotide G8-507 is comple-mentary to residues 484 to 507 of the gene 8 mRNA. The oligo-nucleotides were added in varying concentrations to replica-tion assays containing wild-type gene 8 RNA, and the dsRNA products were detected by sodium dodecyl sulfate-polyacryl-amide gel electrophoresis and quantified with a PhosphorImag-er (Fig. 5). At concentrations of 20 nM, the B1-59 oligonucle-otide inhibited replication by more than fivefold, whereas the Wt-59and G8-507 oligonucleotides inhibited replication only slightly (,twofold). At 500 nM, the presence of the B1-59 oli-gonucleotide reduced replication by more than 100-fold, while the level of inhibition caused by the Wt-59and G8-507 oligo-nucleotides was less than threefold (Fig. 5). These results sup-port the conclusion that base pairing of the predicted 39 single-stranded terminus of gene 8 wild-type mRNA interferes with its ability to function as a template for replication.
G8-5*-B1 and G8-5*-B2 RNAs interfere with replication of wild-type gene 8 mRNA. The G8d45-543 RNA retains the complete sequence of the 59and 39UTRs of the wild-type gene
8 mRNA but lacks residues 45 to 543. The mfold program predicts that the G8d45-543 RNA contains the 59stem-loop of the wild-type RNA but that this structure is not positioned opposite the 39 UTR of the RNA and therefore that G8d45-543 does not contain a 59-to-39panhandle (Fig. 6A). Like the G8-59-B1 and G8-59-B2 RNAs, we observed that the G8d45-543 RNA replicates efficiently in replication assays when the mutant RNA is the only template that is present in the reaction mixture (Fig. 2). However, in assays containing equal amounts of G8d45-543 RNA and wild-type gene 8 mRNA, replication of the mutant template was reduced by approximately 10-fold and therefore is inefficient in comparison to replication of the wild-type template (Fig. 6B). This same phenotype has been described for other gene 8 RNAs that contain deletions at or near their 59ends (29, 44). The three most-likely possibilities to explain the inhibitory effect that wild-type RNA has on the replication of the G8d45-543 RNA are (i) the viral RNA poly-merase, i.e., VP1, binds the wild-type RNA more efficiently than G8d45-543 RNA; (ii) the viral RNA polymerase initiates minus-strand synthesis more efficiently on the wild-type RNA than the G8d45-543 RNA; or (iii) a combination of (i) and (ii). To determine whether the G8-59-B1 and G8-59-B2 RNAs were like wild-type RNA and were able to competitively interfere with the replication of the G8d45-543 RNA, replication assays were performed with equal amounts of G8d45-543 RNA and either G8-59-B1 or G8-59-B2 RNA. The analysis showed that despite their replication-defective phenotypes, the G8-59-B1 and G8-59-B2 RNAs inhibited the replication of G8d45-543 to at least the same extent (.10-fold) as wild-type gene 8 RNA (Fig. 6B). Thus, the mechanism by which wild-type RNA and the mutant RNAs, G8-59-B1 and G8-59-B2, interfere with rep-lication of G8d45-543 is probably the same and does not cor-relate with the level of replication of the competitor RNA in the assay. The inhibitory effects are rotavirus specific given that the replication of G8d45-543 was not affected by the addition of 10-fold excess of yeast RNA in reaction mixtures (Fig. 6B). The results described above were consistent with the idea that even though they were replication defective, the G8-59-B1 and G8-59-B2 RNAs bound the viral RNA polymerase with the same efficiency and affinity as the wild-type gene 8 RNA. If so, then these mutant RNAs should also competitively interfere termined by phosphorimaging. gene 8 RNA in vitro. Wild-type gene 8 template RNA (0.1mg) was replicated either alone (lane 3) or in the presence of the indicated concentrations of the oligonucleotides Wt-59, B1-59, or G8-507 (lanes 4 to 12). The32P-labeled dsRNA products were resolved on a 12.5% polyacrylamide gel and quantified by phos-phorimaging. The amount of dsRNA synthesized in the absence of oligonucle-otide (lane 3) was considered to be 100%. Lane 1,32P-labeled genomic dsRNA of SA11-4F; lane 2, no template RNA was added to the reaction mixture.
on November 9, 2019 by guest
[image:6.612.311.545.71.221.2]with the replication of wild-type gene 8 RNA. To test this possibility, replication assays were carried out with32P-labeled
wild-type gene 8 mRNA but no [32P]UTP, and as competitor,
unlabeled wild-type gene 8 RNA, G8-59-B1 RNA, or yeast RNA. The results showed that all concentrations of G8-59-B1 RNA significantly interfered with the replication of the 32
P-labeled gene 8 wild-type RNA, with the higher concentrations of the mutant RNA (0.5 to 2.0mg) reducing the synthesis of wild-type dsRNA 5- to 10-fold (Fig. 7). Interestingly, G8-59-B1 was approximately twice as effective in inhibiting replication of the labeled wild-type RNA as was the unlabeled wild-type RNA. The competitive interference of the unlabeled wild-type and mutant RNA was rotavirus specific, as yeast RNA had little or no effect on the replication of the32P-labeled wild-type
RNA (Fig. 7).
G8-5*-B1 efficiently binds RNA polymerase.We have shown that the G8-59-B1 mutant, although poorly replicated, is able to inhibit the replication of both wild-type and G8d45-543 RNAs. The simplest explanation for this observation is that the mutations in G8-59-B1 RNA do not affect the ability of the RNA to bind the polymerase, and therefore the RNA is able to specifically interfere with the replication of other template RNAs. In a previous study, Patton (27) used short gene 8-spe-cific RNA probes in electrophoretic mobility shift assays to show that the viral RNA polymerase, VP1, specifically recog-nizes and binds to the 39end of rotavirus mRNA. Because of the length of the RNA, it was not possible to use G8-59-B1 as the probe in electrophoretic mobility shift assays and thereby directly explore the question of whether the mutant RNA efficiently binds VP1. To overcome this limitation, competition assays were performed instead to evaluate the ability of the G8-59-B1 RNA, gene 8 wild-type RNA, and yeast RNA to competitively interfere with the binding of VP1 to the 32
P-labeled probe, SP72-v39-40. This probe is 73 nucleotides in length, and the sequence of its last 40 nucleotides is identical to
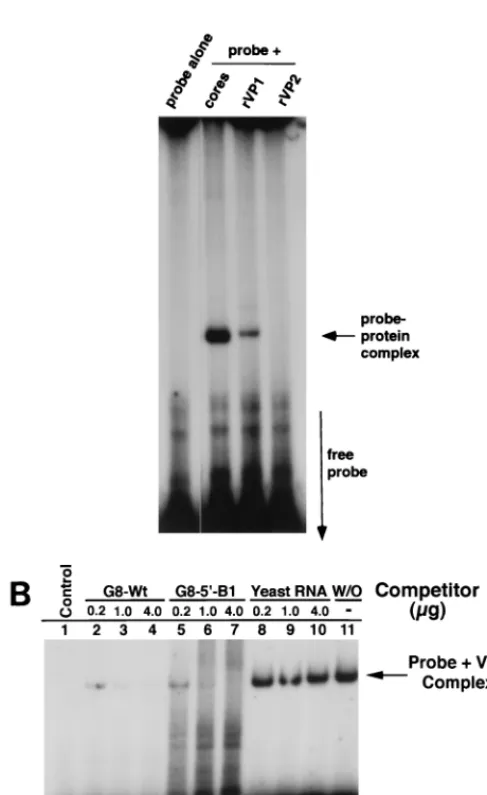
that present at the 39 end of the gene 8 RNA. As shown by assay with baculovirus-expressed recombinant VP1, 32
[image:7.612.85.512.71.285.2]P-la-beled SP72-v39-40 is able to interact with VP1 to form a com-plex that can be detected in the electrophoretic mobility shift assay (Fig. 8A). The results of the competition assays showed that for all concentrations of competitor RNA examined (0.2, 1.0, and 4.0mg), wild-type RNA and G8-59-B1 interfered with the formation of the VP1-SP72-v39-40 complex to a similar extent (Fig. 8B). This interference was specific given that yeast RNA had no effect on the interaction of VP1 and the probe. FIG. 6. Inhibition of the replication of the G8d45-543 RNA by wild-type and mutant RNAs. (A) The optimal secondary structure for the 59and 39ends of the G8d45-543 RNA is shown. Note that the 59end of the RNA forms the same stem-loop structure as predicted for SA11 wild-type gene 8 mRNA but that the 59and 39ends of the RNA do not interact due to the deletion of residues 45 to 543 (Fig. 1A). (B) Replicase assays contained 0.1mg of the internal deletion mutant RNA, G8d45-543 and no competitor RNA (Without), 0.1mg of competitor RNA (G8-Wt, G8-59-B1, G8-59-B2, or Yeast 1X), or 1.0mg of competitor RNA (Yeast 10X). The 32P-labeled dsRNA products were resolved on a 12.5% polyacrylamide gel and quantified with a PhosphorImager. The amount of G8d45-543 dsRNA synthesized in the absence of competitor RNA was considered to be 100%, and the relative amount of G8d45-543 dsRNA synthesized in the presence of the competitor RNAs is given (percentage of replication of reporter RNA).
FIG. 7. Inhibition of gene 8 RNA replication by competitor RNAs.32 P-labeled wild-type gene 8 RNA (0.1mg) was replicated alone (lane 2) or in the presence of the indicated amount (in micrograms) of cold competitor RNA. The 32P-labeled dsRNA products were resolved by gel electrophoresis and detected by autoradiography. The intensities of the32P-labeled bands were determined with a PhosphorImager. The amount of gene 8 dsRNA made in the absence of competitor RNA was considered to be 100%. Lane 1,32P-labeled genomic dsRNA of SA11-4F.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.316.513.506.655.2]These data indicate that although G8-59-B1 is replicated 100-fold less efficiently than wild-type gene 8 RNA, it is able to bind VP1 as efficiently as the wild-type RNA. Hence, events other than merely the binding of the RNA polymerase to the RNA template are important for efficient synthesis of minus-strand RNA.
others have proposed that rotavirus mRNAs fold in a manner such that interaction of the 59and 39termini is allowed, there-by producing a panhandle structure that serves as the promoter for minus-strand synthesis (15, 26, 28, 29). The structure of the rotavirus mRNA is suggested therefore to be similar to the panhandles formed by the RNAs of influenza virus, which have been shown to promote RNA synthesis (11, 14, 39). Besides rotavirus mRNAs, analysis of the secondary structure of other
Reoviridae mRNAs, including those of wound tumor virus (1),
reovirus (4), and rice dwarf virus (40), has indicated that the mRNAs of these viruses may also fold to produce panhandle-shaped molecules.
In this study, we have used current RNA-folding algorithms to predict the secondary structures of all 11 rotavirus NSP2 genes available in GenBank. The computed secondary struc-ture of the SA11 NSP2 mRNA predicts that the last 12 nucle-otides of the RNA are single-stranded and that the 59 UTR folds to form a stem-loop structure. The 59stem-loop and the 39single-stranded tail are proposed to be held adjacent to each other by base pairing between the region of the RNA imme-diately downstream of the 59 UTR and upstream from the 39 single-stranded tail. The mfold program predicted that the most-stable secondary structures for the NSP2 mRNA of six of the other eight mammalian strains of rotavirus (DS-1, Hu-SG2, NCDV, OSU, RF, and Wa) formed a panhandle structure like that of SA11 NSP2 mRNA. The two exceptions were for the UK and IS2 strains of virus. In the case of UK, while the most-stable structures did not show the SA11-type panhandle structure, the two next-most-stable structures did, and these two differed only minimally in free energy (,2 kcal/mol) from the most-stable structure. For the IS2 strain, no dominant structure emerged for the NSP2 mRNA. Instead, of the 10 sta-ble structures computed for the mRNA, four distinct classes of structures were predicted, with one class like that of the SA11 panhandle. Analysis of the predicted secondary structures of other rotavirus genes (e.g., OSU VP7 mRNA, porcine CN86, and CC86 gene 11 mRNAs) suggests that ends of other mRNAs may interact to form a panhandle and a 39single-stranded tail. Of particular note, analysis of the reovirus T1L S2 gene by the
mfold program indicated that the ends of the S2 mRNA
inter-act to form a 59and 39panhandle and a 39single-stranded tail that are remarkably similar to those predicted for the SA11 NSP2 and the OSU VP7 mRNAs.
[image:8.612.51.295.137.535.2]The results of experiments (present and previous) have dem-onstrated that deletion or mutation of residues making up the 59 UTR reduces the efficiency at which the rotavirus NSP2 mRNA replicates, particularly in competition assays that also contain wild-type RNA (Fig. 3) (29, 44). This suggests that the 59UTR and its secondary structure play a key but an as-yet-undefined role in minus-strand synthesis. However, studies on FIG. 8. The replication-defective RNA, G8-59-B1, competitively interferes
with the formation of VP1-probe complexes. (A) The sequence of the 73-nucleotide SP72-v39-40 probe is given, and the portion of the probe that corre-sponds to the last 40 residues of the gene 8 RNA is underlined.32P-labeled SP72-v39-40 probe (10 pmol) and 2.5mg of rabbit liver tRNA were incubated alone, with open cores or with purified recombinant VP1 (60 ng) or VP2 (400 ng) (30). (B)32P-labeled SP72-v39-40 probe (27 ng; 1.2 pmol) was incubated alone (lane 1) or with open cores and no competitor RNA (lane 11), or with 0.2mg (0.6 pmol), 1.0mg (2.8 pmol), or 4.0mg (11.4 pmol) of G8-wt, G8-59-B1 RNAs, or yeast RNA. The molar ratios of competitor RNA to probe in assays containing 0.2, 1.0, and 4.0mg of G8-wt and G8-59-B1 RNA were 0.5, 2.3, and 9.5, respec-tively. VP1-probe complexes were detected in reaction mixtures by electrophore-sis on nondenaturing 8% polyacrylamide gels and by autoradiography.
on November 9, 2019 by guest
the internal deletion mutant G8d45-543 indicate that the mere presence of the 59 UTR and its secondary structure is not sufficient to allow the mRNA to undergo maximum levels of replication, even when the 39 half of the mRNA is present. Instead, analysis of G8d45-543 indicates that along with se-quences in the 59 and 39 UTRs, an internal sequence of the RNA is required for maximal RNA replication, perhaps be-cause it promotes folding of the RNA so that the 59stem-loop and 39 tail are correctly juxtaposed. From these results, it is possible to infer that two types of cis-acting replications exist within the viral template mRNA, those that contain recogni-tion signals that stimulate the binding of viral proteins neces-sary for RNA synthesis (e.g., VP1) and those that are necesneces-sary for the folding of the RNA so that the promoter for minus-strand synthesis is created.
The last 12 nucleotides of the rotavirus gene 8 mRNA con-tain a cis-acting signal that is essential for minus-strand syn-thesis and is predicted to project as a single-stranded tail from the panhandle region of the RNA. In this study, we used two approaches to show that the predicted single-stranded nature of the 39tail is important to the function of the essential rep-lication signal. Firstly, the introduction of mutations into the 59 end of the gene 8 mRNA that caused complete (G8-59-B1) or near-complete (G8-59-B2) complementarity between the 59end and the 39tail were found to significantly inhibit minus-strand synthesis (.10-fold). Furthermore, compensatory mutations that were introduced into the 39end of the replication-defec-tive G8-59-B1 and G8-59-B2 RNAs that decreased the extent of complementarity between the 59end and 39tail were found to stimulate minus-strand synthesis. The fact that deletion of res-idues 2 to 8 of the gene 8 mRNA (G8-59-B4) produced a template which could be replicated efficiently and, except for the presence of a 39single-stranded tail, was structurally iden-tical to the replication-defective G8-59-B1 RNA provides com-pelling evidence that the 39end of the mRNA must be single-stranded for efficient minus-strand synthesis. Secondly, the presence of an oligonucleotide (B1-59) in the cell-free replica-tion that was fully complementary to the 39tail of the gene 8 mRNA inhibited the synthesis of minus-strand RNA by more than 100-fold (500 nM). In contrast, the same concentration of an oligonucleotide that was not complementary to the 39 tail and that corresponded in sequence to the first 18 nucleotides of the gene 8 mRNA had only a minimal effect on minus-strand synthesis (,threefold). Taken together, these data in-dicate that the 39 end of the template mRNA must be single stranded for the 39cis-acting replication signal to function and
for dsRNA synthesis to occur. The fact that the 39cis-acting
replication signal may not function unless it is single stranded may explain why the viral replicase in open-core preparations is unable to use the endogenous dsRNA genome as a template for the synthesis of minus-strand RNA.
Although replication defective, the mutant G8-59-B1 RNA was shown to be able to competitively interfere with the rep-lication of other rotavirus mRNAs in vitro and to do so as efficiently as wild-type mRNA. Hence, the G8-59-B1 RNA is phenotypically wild type with respect to its ability to recruit at least one of the trans-acting protein factors in the cell-free system that is required for minus-strand synthesis. A compet-itive electrophoretic mobility shift assay was used to show that the G8-59-B1 RNA binds the RNA polymerase VP1 as effi-ciently as wild-type RNA. While the recognition signal for VP1 in the mRNA has not been precisely mapped, these data indi-cate that whether the 39essential replication signal is single or double stranded has no effect on the ability of the polymerase to bind to the RNA. Furthermore, the fact that gene 8 mutant RNAs containing deletions of residues 1 to 10 (g8Rd1 to -10
[29]), 4 to 50 (g8Rd4 to -50 [29]), and 45 to 453 (G8Rd45 to -543) retain the ability to replicate to a level within threefold of the wild-type mRNA suggests that the 59portion of the mRNA does not contain the recognition signal for the RNA polymer-ase. Instead, as was indicated in previous studies with electro-phoretic mobility shift assays (27), the polymerase recognition signal probably resides in the 39-terminal region of the mRNA. The finding that the G8-59-B1 RNA can efficiently bind VP1 but is replication defective supports the conclusion that the association of the polymerase with the template RNA is not the sole factor which determines whether an RNA undergoes replication. Indeed, it is known that VP2 is also essential for RNA replication (22, 46) and that, when combined, VP1 and VP2 possess replicase activity that stimulates the synthesis of dsRNA from wild-type mRNA in vitro (30). VP2 is the most abundant protein in the open-core replication system and has been shown to have RNA-binding activity that is nonspecific and that recognizes both single-stranded RNA and dsRNA (18). As a result, both VP1 and VP2 can be expected to asso-ciate with the mutant template G8-59-B1 in the cell-free rep-lication system but this interaction does not lead to efficient levels of dsRNA synthesis. Thus, the replication-defective phe-notype of G8-59-B1 is probably related not to a defect in the ability to recruit the minimum two proteins required for RNA replication but, instead, to a defect in the ability of these pro-teins to initiate minus-strand synthesis on the template. More precisely, the data provide evidence that minus-strand initia-tion, but not VP1 and VP2 binding, requires that the 39
cis-acting replication exist in a single-stranded form, i.e., an open form.
In summary, the rotavirus gene 8 mRNA is predicted to fold to form a panhandle with a single-stranded 39tail. Data were obtained with a cell-free replication system showing that the 39 tail must exist in a single-stranded form if the mRNA is to function as an efficient template for minus-strand synthesis. While RNA-folding programs do not take into account the effect that proteins may have on the secondary structure of the RNA, the rotavirus core proteins VP1, VP2, and VP3 may serve to promote the formation of the panhandle. In particular, since it is thought that the polymerase VP1 binds the 39end of mRNA (27, 31) and the capping enzyme VP3 binds the 59end of mRNA (31), and both these proteins bind to adjacent sites at the NH2end of VP2 (47), the VP1-VP2-VP3 complex may
catalyze the formation of the panhandle by bringing the 59and 39ends of the RNA close to each other. Given that the 39end of the template RNA for replication must be single stranded for minus-strand synthesis to occur, it remains unclear how the RNA polymerase is able to initiate mRNA synthesis on the double-stranded genome segments. Perhaps a mechanism ex-ists by which the 59 end of the dsRNA segments is partially melted, thus providing a single-stranded cis-acting signal at the 39end of the minus strands that allows initiation of plus-strand synthesis.
ACKNOWLEDGMENTS
We acknowledge the excellent technical assistance of Melinda Jones with this project. We also thank Kim Green, Albert Kapikian, and Robert Chanock for critical reviews of the manuscript.
REFERENCES
1. Anzola, J. V., Z. Xu, T. Asamizu, and D. L. Nuss. 1987. Segment-specific inverted repeats found adjacent to conserved terminal sequences in wound tumor virus genome and defective interfering RNAs. Proc. Natl. Acad. Sci. USA 84:8301–8305.
2. Beekwilder, M. J., R. Nieuwenhuizen, and J. van Duin. 1995. Secondary structure model for the last two domains of single-stranded RNA phage Q beta. J. Mol. Biol. 247:903.
on November 9, 2019 by guest
http://jvi.asm.org/
Fields, D. Knipe, P. M. Howley, et al. (ed.), Fundamental virology, 3rd ed. Lippincott-Raven Publishers, Philadelphia, Pa.
11. Fodor, E., D. C. Pritlove, and G. G. Brownlove. 1994. The influenza virus panhandle is involved in the initiation of transcription. J. Virol. 68:4092– 4096.
12. Gallie, D. R., J. N. Feder, R. T. Schmke, and V. Walbot. 1991. Functional analysis of the tobacco mosaic virus tRNA-like structure in cytoplasmic gene regulation. Nucleic Acids Res. 19:5031–5036.
13. Harrison, G. P., and A. M. Lever. 1992. The human immunodeficiency virus type 1 packaging signal and major splice donor region have a conserved stable secondary structure. J. Virol. 66:4144–4153.
14. Hsu, M.-T., J. D. Parvin, S. Gupta, M. Krystal, and P. Palese. 1987. Genomic RNAs of influenza viruses are held in circular conformation in virions and in infected cells by a terminal panhandle. Proc. Natl. Acad. Sci. USA 84:8140– 8144.
15. Imai, M., M. A. Richardson, N. Ikegami, A. J. Shatkin, and Y. Furuichi. 1983. Molecular cloning of double-stranded RNA viruses genomes. Proc. Natl. Acad. Sci. USA 80:373–377.
16. Kapikian, A. Z. 1996. Overview of viral gastroenteritis. Arch. Virol. 12:7–19. 17. Kattoura, M., L. L. Clapp, and J. T. Patton. 1992. The rotavirus non-structural protein, NS35, is a non-specific RNA-binding protein. Virology 191:698–708.
18. Labbe, M., P. Baudoux, A. Charpilienne, D. Poncet, and J. Cohen. 1994. Identification of the nucleic acid binding domain of the rotavirus VP2 pro-tein. J. Gen. Virol. 75:3423–3430.
19. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680–685.
20. Lawton, J. A., C. Q.-Y. Zeng, S. K. Mukherjee, J. Cohen, M. K. Estes, and B. V. V. Prasad.1997. Three-dimensional structural analysis of recombinant rotavirus-like particles with intact and amino-terminal-deleted VP2: impli-cations for the architecture of the VP2 capsid layer. J. Virol. 71:7353–7360. 21. Leathers, V., R. Tanguay, M. Kobayashi, and D. R. Gallie. 1993. A phylo-genetically conserved sequence within viral 39 untranslated RNA pseudoknots regulates translation. Mol. Cell. Biol. 13:5331–5347. 22. Mansell, E. A., and J. T. Patton. 1990. Rotavirus RNA replication: VP2, but
not VP6, is necessary for viral replicase activity. J. Virol. 64:4988–4996. 23. McBride, M. S., and A. T. Panganiban. 1996. The human immunodeficiency
virus type 1 encapsidation site is a multipartite RNA element composed of functional hairpin structures. J. Virol. 70:2963–2973.
24. Mirmomeni, M. H., P. J. Hughes, and G. Stanway. 1997. An RNA tertiary structure in the 39-translated region of enteroviruses is necessary for efficient replication. J. Virol. 71:2363–2370.
25. Nibert, M. L., L. A. Schiff, and B. N. Fields. 1996. Reoviruses and their replication, p. 1555–1596. In B. N. Fields, D. Knipe, P. M. Howley, et al. (ed.), Fundamental virology, 3rd ed. Lippincott-Raven Publishers, Philadel-phia, Pa.
26. Okada, Y., M. A. Richardson, N. Ikegami, A. Nomoto, and Y. Furuichi. 1984. Nucleotide sequence of the human rotavirus genome segment 10, an RNA encoding a glycosylated virus protein. J. Virol. 51:856–859.
27. Patton, J. T. 1996. Rotavirus VP1 alone specifically binds to the 39end of
D. Knipe, P. M. Howley, et al. (ed.), Fundamental virology, 3rd ed. Lippin-cott-Raven Publishers, Philadelphia, Pa.
35. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1992. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
36. Sanger, F., S. Nicklen, and A. R. Coulsen. 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463–5467. 37. Serra, M. J., D. H. Turner, and S. M. Freier. 1995. Predicting
thermody-namic properties of RNA. Methods Enzymol. 259:243–261.
38. Shaw, A. L., R. Rothnagel, D. Chen, R. F. Ramig, W. Chiu, and B. V. V. Prasad.1993. Three dimensional visualization of the rotavirus hemaggluti-nin structure. Cell 74:693–701.
39. Stoeckle, M. Y., M. W. Shaw, and P. W. Choppin. 1987. Segment-specific and common nucleotide sequences in the noncoding regions of influenza B virus genome RNAs. Proc. Natl. Acad. Sci. USA 84:2703–2707.
40. Uyeda, I., H. Kudo, T. Takahashi, T. Sano, K. Ohshima, T. Matsumura, and E. Shikata.Nucleotide sequence of the rice dwarf virus genome segment 9. J. Gen. Virol. 70:1297–1300.
41. Valenzuela, S., J. Pizarro, A. M. Sandino, M. Vasquez, J. Fernandez, O. Hernandez, J. Patton, and E. Spencer.1991. Photoaffinity labeling of rota-virus VP1 with 8-azido-ATP: identification of the viral RNA polymerase. J. Virol. 65:3964–3967.
42. Walter, A. E., D. H. Turner, J. Kim, M. H. Lyttle, P. Muller, D. H. Mathews, and M. Zuker.1994. Coaxial stacking of helixes enhances binding of oligo-ribonucleotides and improves predictions of RNA folding. Proc. Natl. Acad. Sci. USA 91:9218–9222.
43. Wentz, M. J., C. Q.-Y. Zeng, J. T. Patton, M. K. Estes, and R. F. Ramig. 1996. Identification of the minimal replicase, and the minimal promoter of (2 )-strand synthesis, functional in rotavirus RNA replication in vitro. Arch. Virol. 12:59–67.
44. Wentz, M. J., J. T. Patton, and R. F. Ramig. 1996. The 39-terminal consensus sequence of rotavirus mRNA is the minimal promoter of negative-strand RNA synthesis. J. Virol. 70:7833–7841.
45. Xu, Z., J. V. Anzola, C. M. Nalin, and D. L. Nuss. 1989. The 39-terminal sequence of a wound tumor virus transcript can influence conformational and functional properties associated with the 59-terminus. Virology 170:511– 522.
46. Zeng, C. Q.-Y., M. J. Wentz, M. K. Estes, and R. F. Ramig. 1996. Charac-terization and replicase activity of double-layered and single-layered rotavi-rus-like particles expressed from baculovirus recombinants. J. Virol. 70: 2736–2742.
47. Zeng, C. Q.-Y., M. K. Estes, A. Charpilienne, and J. Cohen. 1998. The N terminus of rotavirus VP2 is necessary for encapsidation of VP1 and VP3. J. Virol. 72:201–208.
48. Zuker, M. 1989. The use of dynamic programming algorithms in RNA secondary structure prediction, p. 159–184. In M. S. Waterman (ed.), Math-ematical methods for DNA sequences. CRC Press, Inc., Boca Raton, Fla. 49. Zuker, M. 1994. Prediction of RNA secondary structure by energy
minimi-zation, p. 267–294. In A. M. Griffin and H. G. Griffin (ed.), Computer analysis of sequence data, part II, vol. 25. CRC Press, Inc., Totowa, N.J.