0022-538X/79/08-0494/12$02.00/0
Further
Characterization
of
the
Replicative Complex
of
Vesicular Stomatitis Virus
CHRISTIAN C. SIMONSEN,* VIRGINIA M. HILL,AND DONALDF. SUMMERS
Departmentof Cellular, Viraland MolecularBiology, Universityof Utah Medical Center, Salt LakeCity,
Utah 84132
Receivedfor publication 23 March 1979
Replicating vesicular stomatitis virus ribonucleoprotein (RNP) complexeswere
isolated in nonequilibrium Renografin density gradients. These nascent RNPs
had the same buoyant densityas virionnucleocapsids in both isopycnic
Renq-grafm and CsClgradients. Both transcribing and replicating RNP complexeswere
showntobestable insucrosegradients,whereasonly replicatingRNPcomplexes
werestable inRenografin gradients.Sizeanalysisof the5-min-pulse-labeledRNA
speciesfrom the replicatingRNPs using methylmercury gels revealed that the
nascentstrandswere primarily less thanfull-lengthmolecules. Longertimesof
radiolabeling demonstrated that the nascent RNA accumulated as 42S RNA,
whichwasprimarily ofthesamesense asthe virion strand when itwas
radiola-beledat5hpostinfection.The percentage of this radiolabeled RNA whichwas
plusstrandedwashigherat 2.5hpostinfection, reflective of the shift inplus-to
minus-strandfull-length 42SRNAsynthesiswhichoccursinthecell. Addition of
cycloheximidetotheinfectedcellsbefore the addition of the radiolabelprevented
the formation of these RNP complexes. Both the changein the percentage of
minus strands found in the RNPcomplexesatthe different timespostinfection
and the sensitivitytocycloheximide indicate that the RNP complexwhichwas
isolatedwasindeed thereplicative complex.
During the infection of HeLa cells by vesicular
stomatitis virus(VSV),both virion minus-strand
andcomplementary plus-strand full-length42S
RNAcanbe isolated from intracellular
ribonu-cleoprotein (RNP) complexes (34, 36). These
RNPs are composed of 42S RNA and three
VSV-specific proteins: the N, or nucleocapsid,
protein; the NSprotein,aphosphorylated
poly-peptide;and the Lprotein(17, 18, 36). The virion
nucleocapsid is able todirect the in vitro
syn-thesis of the five VSV-specific mRNA's which,
likethe in vivoVSV mRNA's,arecappedatthe
5'end, methylated,andpolyadenylated(2, 7, 20,
25-27,32).
Reconstitution experimentshave shown that
both the L and NSproteins are required to form
thevirion-associatedRNA-dependentRNA
po-lymerase (17,18,28). Thetemplate for
transcrip-tion has been demonstratedtobecomposed of
minus-strand 42SRNA and Nprotein (11, 17).
Therequirements forreplication, however, are
notwellknown. Thetemperature-sensitive
mu-tant tsG114, which possesses a thermolabile L
protein (21), has been usedtoprovide indirect
evidence that theLproteinisinvolvedin
repli-cation (30). Several othermutants suggest arole
inreplication forthe N andNSproteins aswell
(15, 16). For thesereasonsit has been assumed
that replication occurs on an RNP template
similartothetranscriptiveRNP.
Fractionation techniques previously used to
isolate VSV RNPs do notseparate replicating
RNPs from transcribing RNPs. Sedimentation
velocity centrifugation analyses have indicated
thatintracellular VSV RNPs sedimentas 140S
molecules, whether or not nascent mRNA is
associated with the RNP complexes (31, 35).
The lack of resolution is furthercomplicated by
the fact thatreplicationaccounts for only 10%
of the total VSV-specific RNA synthesized in
infectedcells (38);thus,sucrosegradients have
notbeenabletoseparatereplicatingRNPsfrom
transcribing RNPs. CsCl gradients have been
used toisolate glutaraldehyde-fixed messenger
RNPs (19) and intracellularnucleocapsids(34);
however, themessengerRNPsare notstablein
CsCl withoutprior fixation withglutaraldehyde.
RNPsisolated with CsClgradientsdo notretain
the L and NS proteins, which are known to
comprisepart of thefunctionalRNPs inthecell
(17, 18).Inaddition,allof the intracellularRNPs
band at the same density in CsCl. For these
reasons, CsCl gradients have not been used to
separate the RNPcomplexes.
lodinateddensitygradientmedia suchas
me-trizamide and Renografm have beenemployed
494
on November 10, 2019 by guest
http://jvi.asm.org/
toobtainedresolution of proteins, nucleic acids, and RNPs in the same gradients (6, 22). These
media arehypertonic and minimize nonspecific
RNA-protein interactions, yet do not dissociate
mostRNPs. Severalinvestigatorshave used
me-trizamnide and Renografin to examine chromatin
(12),influenza RNPs (10), and proteins (8). We
haverecently described the isolation in
Reno-grafingradients of RNP complexes from
VSV-infected HeLa cells (V.M.Hill, C. C.Simonsen,
and D. F. Summers,submitted for publication).
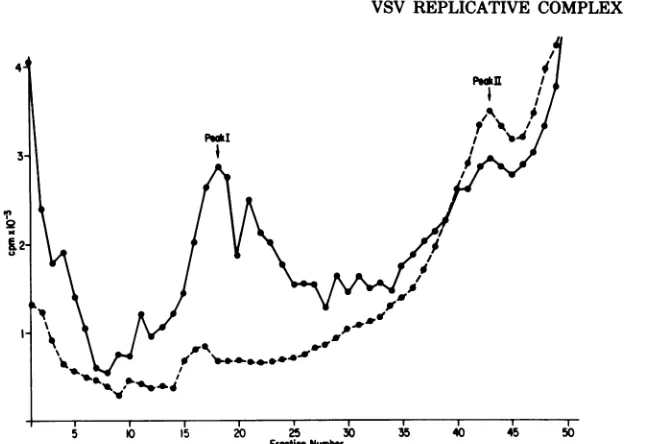
Several peaks of radioactivity were observed
when an infected cell extract radiolabeled from
3 to5hpostinfection (p.i.) wascentrifuged in a
Renografin gradient. One of those peaks
con-tained predominantly minus-strand 42S RNA
and was termed peak I. A similar pattern of
radioactivitywasseenafteraninfectedcell
ex-tract pulse-labeled for 5 min at 5 h p.i. was
analyzed, except that peak I exhibited a
pro-nounced trailing edge. Virion nucleocapsids
formedonlyahomogeneouspeakIwhichlacked
atrailingedge.
Genomelengthand smaller RNAswerefound
in thepeak fractions of thepulse-labeled peak I,
whereas onlythe smaller RNAswere found in
the trailing fractions. This RNA, pulse-labeled
for 5 min at 5 hp.i., was 80%minus stranded,
the same percentage observed with
pulse-la-beled42S RNA obtained from intracellular
nu-cleocapsidsat 5hp.i.,whenreplicationis maxi-mal (34). The nascent minus-strand RNA was
shown tobe identicaltothe 5'end of the VSV
genome RNA. We also observed that thenascent
RNAwasRNase resistant,wassinglestranded,
andwasisolatedat adensity of 1.240
g/cm3
inRenografin. All of this suggested that peak I
contained nascent replicating VSV RNA
asso-ciated withprotein as areplicating RNP
com-plex. Herein we extend these observations and
further characterize the complexes to show by
several different criteria that these RNPs are
indeed VSVreplicativecomplexes.
MATERIALS AND MErHODS
Cells,virusinfection,andradioactivelabeling.
Suspension culturesofHeLa S3 cellsweregrownin
Joklikmodifiedminimal essential medium(Flow
Lab-oratories) supplementedwith 2 mM glutamine plus
5%fetal calfserum(FlowLaboratories) at a concen-tration of 2 x 105 to 8 x 10' cells per ml. Stock
preparationsof VSV(Indiana serotype) were grown in
HeLacells, purified, and assayed asdescribed previ-ously (3, 4). Cells were collected by centrifugation, resuspended in growth medium minus serum at a
concentration of6 x106cellsper ml, and then infected with 10PFUofVSVper cell. At 1hp.i. serum was addedto5% and actinomycin D (a gift fromMerck, Sharp & Dohme) was added to 1 yg/ml. At times
ranging from 2.5 h to 5 h p.i.
['H]uridine
or['H]-Corp., Boston, Mass.) or both were added to a concen-trationof 200,tCi/mlfor the indicated labeling periods, after which crushed, frozen medium was added and
thecellswerepelletedbycentrifugation for 3min at
900xg.Thecell pelletwaslysed by the addition of 1
mlof 1% Nonidet P-40 in NT buffer (0.1 M NaCl, 0.01 MTris,pH7.5)and kept on ice for 10 min. The nuclei were removed by centrifugation as described above andwerethenresuspended in 1 ml of 0.2% deoxycho-late-0.3% Nonidet P-40 in NT buffer and recentri-fuged. Thesupernatants from the 1% Nonidet P-40 treatmentand the deoxycholate-Nonidet P-40 wash werethencombined andlayered onto preformedCsCl
orRenografingradients.
Isolationof RNPcomplexes.RNPs were isolated
by using eitherRenografin,CsCl, or sucrose gradients. Forisolation of the replicativeRNPs,the cytoplasmic extractsfrom above were layered onto preformed
36-ml, 15to50%or20 to60%Renografin (E. R. Squibb & Sons) gradients in NT buffer and centrifuged at 23,000 rpmand4°C in an SW27 rotor for 16 h (for
nonequilibriumgradients) or for 80 h (for equilibrium
gradients). Thegradients were fractionated by pump-ingfrom the bottom, and radioactivity was determined bydirectly assaying50-1lportions in7mlof
scintilla-tion cocktail in aBeckman liquidscintillation spec-trometer.
Preformed 20 to 40% (wt/wt) CsCl (Varlacoid
ChemicalCo.) gradientsin TNEbuffer (0.025 M
Tris-hydrochloride, pH7.5, 0.05 MNaCl, 0.002 M EDTA)
were overlayed with fractions from the Renografin
gradient which had been diluted 1:3 in NT buffer. Thesegradientswerecentrifugedin anSW41 rotor at 33,000 rpmfor16hat4°C.Aftercentrifugation, the
gradientswerefractionated,andacid-precipitable
ra-dioactivitywasdeterminedasdescribedpreviously(3, 4). Thedensitiesof the fractionsweredeterminedby measuring the refractive indexes withaBausch and Lombrefractometer.
Cytoplasmic extractswerealso layeredonto 15 to
30% sucrosegradientsin NTbuffer,whichwere
cen-trifugedinanSW27rotorfor16h at16,000 rpm and
4°Cand fractionatedasdescribedabove.
Phenol extractionof RNA. Anequalvolume of
phenolsaturated with NETS buffer(0.1MNaCl,0.001
MEDTA,0.01 MTris, pH7.5,0.2% sodiumdodecyl
sulfate) wasaddedto the samplesin 3 ml of NETS
buffer andmixed,and thetwo layerswereseparated
by centrifugation. The aqueous layerwas collected,
and thephenol layerwasextractedoncemorewith 3 ml of NETS buffer. The aqueous fractionswere
com-bined,sodiumacetate wasaddedto afinal
concentra-tionof 0.2 M,2volumes of ethanolwereadded,and
theRNAwasprecipitatedat-70°C overnight. RNA
waspelletedbycentrifugationat16,000xgfor 30min
at40C.
Purification ofVSV-specificRNA. Minus-strand 42S RNAwasprepared frompurifiedvirionsby dis-rupting the virus in NETS buffer andcentrifugingthe
samplesin 15to30% sucrose-sodiumdodecylsulfate
gradients inanSW41rotor. The 42SRNApeakwas
ethanolprecipitatedanddissolved in2xAbuffer(0.3
M NaCl,0.02MTris,pH 7.4,0.002MEDTA). VSV
mRNAwaspreparedfrom infectedcellsat4.5hp.i.A
on November 10, 2019 by guest
http://jvi.asm.org/
cytoplasmicextract waspreparedasdescribed above
and was phenol extracted, ethanolprecipitated,and
chromatographed on an oligodeoxythymidylic
acid-cellulose column.
Oligodeoxythymidylic acid-cellulose
chroma-tography ofRNA.Oligodeoxythymidylic
acid-cellu-lose (Collaborative Research, Inc., Waltham, Mass.)
columns werepreparedandelutedby themethod of
Banerjeeand Rhodes(7).Thehigh-saltelution buffer
contained 0.01 M Tris-hydroxychloride, pH 7.4, and 0.5 MNaCl,and thelow-saltbuffer contained 0.01 M
Tris-hydrochloride,pH7.4.RNA wasloadedontothe
columninhigh-salt buffer.Fractionscontaining bound
materialwerepooled andethanolprecipitatedas
de-scribed above.
Hybridization of VSVRNA.Hybridizationswere
performedin sealedcapillarypipettes in0.02-mIl
vol-umesbyusingthe conditionsofKolakofsky (24).To
eachsampleof[3H]RNAisolated fromnucleocapsids
were added increasing amounts of unlabeled VSV
virion 42S RNA or mRNA. The sampleswere then heat denatured by heating to 115°C for 3 min and incubated for 3 h at730C. Eachsamplewastreated with a 30-,ug/ml solution of RNase A (Worthington
Biochemicals Corp.)in2xAbuffer for 30 min at250C
and wasthenassayed foracid-precipitable
radioactiv-ity asdescribed above. The self-annealing reactions
were identical, except that no unlabeled RNA was
added to thesamples.
Methylmercury-agarose gel electrophoresis.
Methylmercury-agarose gel electrophoresis was
per-formed byamodificationof the method ofBaileyand
Davidson (5). Seakemagarose powder was obtained from Marine-Colloids, Inc., and methylmercury (II)
hydroxidewasobtainedas a 1 Msolution from Alfa
Products, Danvers, Mass. Because of the toxic and
volatile nature of the mercury compound, operations
wereperformedunder ahood. Horizontal slab gels (19
by13.3by0.4cm; 100-mlbedvolume)were1%agarose inborate buffer (0.05 M boric acid,0.005 M sodium
borate, 0.01 M sodium sulfate, 0.001 M trisodium
EDTA, pH 8.0) and contained 0.005 Mmethylmercury
hydroxide.Sampleswereappliedin 40,ulof 1:1
sam-ple-samplebuffer(boratebuffercontaining50%
glyc-eroland 0.1%bromophenol blue),withmethylmercury
hydroxide addedto a concentration of 0.005 M just
beforeloading onto the gel. Thegel reservoirs con-tained boratebuffer,and this buffer was recirculated
atapproximately80ml/h throughoutthe
electropho-resisperiod.Gels were run at 75 to 100 V (50 mA) for
6h(dyefrontmigrated 14cm).
Fluorographyofmethylmercurygels. Gels
con-taiing [3H]RNAwerefixedfor 10 min in 10% acetic
acid containing 0.01 M cysteine. They were
dehy-drated in 100% methanolfor two successive 1-h pe-riods.Afterthey were dried to paper thinness under a vacuum, the gels were soaked in a 10%(wt/vol)
solu-tion of 2,5-diphenyloxazole (New England Nuclear
Corp.).inmethanolfor3h.Gelswere soaked inwater
for10mintoprecipitatethe2,5-diphenyloxazole,
blot-ted dry, and mounted on a, piece of 3 MM paper (Whatman, Ltd., England).Afterbeingcovered with SaranWrap, gelswereexposed to Kodak SB-5 X-ray filmat-70°C anddevelopedafterappropriateperiods of time.
J. VIROL.
RESULTS
Kinetics of
labeling
ofpeak
I. We wishedtodetermine whether 42S RNA would
accumu-late in the Renografin-isolated peak I if the
length of the pulse-labeling period was
in-creased. We also wished to learn whether the
material in thetrailing fractions ofpeakI would
also accumulate duringalongerpulse-labeling
period. VSV-infected HeLa cells were
radiola-beled for 5, 10, 15, and 30 min at 4.5 h p.i.
Cytoplasmic extracts wereprepared from each
sample and centrifuged in identical 15 to 50%
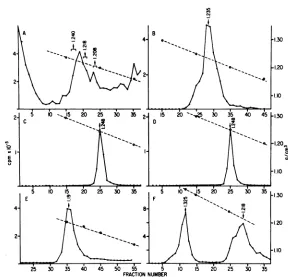
Renografin gradients. The results (Fig. 1)
dem-onstratethat thetrailing fractions of the 5-min
sample (fractions26 to31) were not as apparent
in thel0-minsample and wereevenless
notice-able in the 15-and30-min samples. All samples
werecontinuously labeled; therefore, the
mate-rial in the trailing fractionswas still presentin
the longer-labeled fractions, but comprised a
minorproportionof the totalradiolabeled
ma-terial duetotheincrease in the amount of
radi-olabel inthe peakfractions (fractions 26 to 31,
allsamples).
We nextpooledthe fractions of thereplicative
complexfromFig.1.Thetrailing fractions of the
5-min-pulse-labeled sample were included
be-causetheradiolabeled RNAcontainedinthese
fractionsrepresentedanappreciable amountof
thetotal radiolabeledRNA.Only the peak
frac-tions from the remaining samples were pooled
becausethematerial inthetrailing fractions did
notrepresentasignificant proportionof the total
radiolabeled RNA. The RNA was extracted
from these pools and analyzed in
methylmer-cury-agarose gels.Thedensitometertracingsof
theautoradiographsfrom these gels are shown
inFig.2.The RNAfromthe5-min-pulse-labeled
sampleswasheterogeneous insize,rangingfrom
less than18Sto42S,althoughthe average size
of thissamplewasapproximately26S. The size
distribution ofthe 10-minsample was also
het-erogeneous; however, the average size of the
labeled RNA increased to a value of
approxi-mately 35S,andtherewas anobvious42S band.
Both the15-and30-minsamples contained
pri-marily42SRNA, althoughthereweresignificant
amountsofheterogeneous material less than 42S
RNA in size. This suggests that the radiolabel
accumulatedasgenomelengthRNAasthepulse
timewasincreased.Figure2alsoimplies that it
tooklonger than5min for 42SRNA tobegin to
accumulate.It is notclearwhythis was thecase,
sinceonewouldexpect thata5-minpulse-label
woulduniformly label all nascent RNA. Perhaps
thesynthesis of VSV 42SRNA does not occur
at a uniform rate throughoutthe template; as
wehave notedpreviously, thereisanapparent
on November 10, 2019 by guest
http://jvi.asm.org/
T 0
FRACTION NUMBER
FIG. 1. Renografin gradient centrifugationof cytoplasmic extractsfrompulse-labeled VSV-infected cells.
Infectedcells at 4.5 hp.i.were labeled with 100,uCi each of[3H]adenosineand[3H]uridineperml.Samples
wereremovedat5, 10, 15, and 30 minaftertheadditionofthe labels. Cytoplasmicextracts wereprepared,
layeredonto 15 to50%Renografin gradients, andcentrifuged for17h at 23,000 rpmand 4°C in an SW27
rotor.Fractions (0.5ml)werecollected by pumping out from the bottom, and the radioactivity in each fraction
wasdeterminedbycounting50-,iuportions in a toluene-based scintillation fluid.
pause site near the juncture of the L- and
G-proteincistrons (Hilletal.,submitted for publi-cation).
Hybridizationof pulse-labeledRNAfrom RenografinRNPs. We have recently reported
that the ratio ofplus-tominus-strand 42S RNA
synthesisin infected cellsisgreaterearly in the
infection (2.5 h p.i.) than when replication is
maxinal at 5 h p.i., at which time 80% ofthe
newly synthesized 42S molecules are minus
stranded (34). Wehave also shown that
pulse-labelednascentRNAspecieswhich areisolated
from putative replication complexesat 5 hp.i.
are approximately80% minus stranded (Hillet
al., submitted forpublication).
Todetermine whether pulse-labelednascent
RNA extractedfrom thereplicationcomplexes
wouldreflect the in vivofindings,infected HeLa
cellswerelabeled for5minwith[3H]uridineat
2.5 or 5 h p.i. Cytoplasmic extracts were
pre-pared from these cellsand were centrifugedin
15 to.50% Renografin gradients. The nascent
RNA contained in the RNP complexes (at a
peakdensityof1.240g/cm3)washybridizedwith
unlabeled VSV 42SRNAandVSVmRNA.The
RNPslabeledat2.5hp.i.containedsignificantly
more plus-strand RNA than the RNPslabeled
at5hp.i. (Table1). The changeinthe ratio of
plus- tominus-strand RNA contained in these
RNPcomplexesfromapproximately35:65at2.5
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.104.403.65.443.2]498 SIMONSEN, HILL, AND SUMMERS
origin 42S 28S
I I I
185
E
0
to
cli
.o
cn
2 4 6 E
DISTANCE MIGRATED(cm)
FIG. 2. Densitometer tracings of fluo methylmercury-agarose gels. The RNA co
thepulse-labeled RNPswasextractedfrom tionspooledas indicated in Fig. 1. Appi 50,000cpmofpurified RNA from eachst
electrophoresed inamethylmercury-agaro
fluorographedasdescribed in thetext. T
diograph was analyzed with an Ortec n
densitometer. Thescale isidenticalfor a,
hp.i.to 15:85at5hp.i.is similarto ti
in the ratio ofplus-to minus-strand
synthesiswe have previously observe
indicatingthat theplus-strandRNAsl
lated inpeakIwereproductsofreplic
Effectofcycloheximideonthere
RNA. A further means of differenti
replicative processfrom thetranscrip
ess is by using the protein synthesis
cycloheximide. The synthesis of
mRNA's is notaffected appreciably t
dition ofcycloheximide, whereasrepl
42S RNA israpidlyinhibited (30, 38)
shows that the synthesis of the pea
complex from Renografinwasinhibit
additionofcycloheximide, whereas t:
tion of the slower-sedimenting peak
containsexclusivelyplus-strandseque
etal.,submitted forpublication),wasi
by the addition of the drug. This result also
demonstrates that the cycloheximide-sensitive
5min RNPcomplex isareplicative complex.
Isopycnic gradient centrifugation of the
replicative complexes.Wenextexamined the
RNP complexes in isopycnic Renografin
gra-_ _ dientstostudythebasisforthe isolation of the
heterogeneouspeak I.A5-min-pulse-labeled
cy-10min toplasmicextractwaspreparedfromcellsat5h
p.i. asdescribedaboveandwascentrifugedina
15to50%Renografm gradient for17hat23,000
rpm in an SW27 rotor. Fractions from three
portions of the RNPpeakwerepooledas
indi-cated(Fig. 4A)andcombined, and sampleswere
recentrifuged on identical 20 to 60% gradients
15min for 17, 40, and 80 h (Fig. 4B, C, and D). The
replicative RNP peak, which was very
hetero-geneousafter beingrecentrifuged for 17h(Fig.
4B), formedahomogeneous peakatadensityof
1.248g/cm3 in the isopycnic gradients (Fig. 4C
andD). Under thesameconditions of
centrifu-30min gation, VSV42SRNAwasfoundat 1.19g/cm3
(Fig. 4E). Poliovirustop component
(nucleocap-sidwhich lacksRNA) andpoliovirus infectious
particleswerefoundintheequilibrium gradients
at 1.325and 1.218 g/cm3,respectively (Fig. 4F).
Theresult indicated that theratio ofproteinto
RNA in the nascent RNPs was the same
throughout the peak and trailing fractions of
peakI,sincethesmallerRNPscontained in the
,rographed
trailing
edge
ofpeak
I had the sameisopycnic
intained in densityinRenografinastheRNPs in thepeak
mthefrac- fractions.
roximately
ample was TABLE 1. Hybridization analysis of the RNA
ge
getoand
isolatedfrom replicative
RNPsfrom cytoplasmic
'he autora- extractspulse-labeledat2.5and5hp.i.a
Moael 4MlU
11samples. he change 42S RNA
d invivo,
pecies
iso-ation. bplicative
iating the
itive
proc-inhibitor
the VSV
bythe
ad-lication of
I.Figure3
tk I RNA
;edbythe
he
forma-II, which
-nces(Hill
anaffected
Self-annealed Unlabeled RNA added Tixme Total material
p.i. mRNA 42S RNA
(h) cpm %
cpm % cpm %
2.5 776 298 38 545 70 266 34
2.5 707 277 39 488 69 247 35
5 1,247 407 33 1,175 94 225 18
5 572 192 33 544 95 60 10
a Infected cells werepulse-labeled at either 2.5 or 5 h p.i. as described in thelegendtoFig.1.The replicative RNP complex
wasisolated fromnonequilibriumRenografingradients, and the RNA contained in the RNP was isolated byphenol ex-traction. Thepulse-labeled RNA was mixed with increasing
amounts of unlabeled VSV mRNA or 42S RNA in 20-Il reactionvolumesby using the conditions ofKolakofaky(24). Thesampleswereheatedto115°Cfor1min, annealedfor3
h at730C,and then treated with30ygof RNaseAperml in
2xAbuffer for 30 min at 220C. Plateau levels of RNase resistanceareexpressed after being corrected forthe RNase-resistantmaterial whichwas present when thesamplewas
heatdenatured butnotannealed. Thisfigure varied from2 to
4%oftheinput radioactivity.Self-annealingof thematerial
wasdeterminedasdescribedabove, exceptthatnounlabeled RNAwasaddedtothesamples.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.274.463.456.536.2]Pe E
[image:6.504.102.425.56.278.2]n Number
FIG. 3. Effect of cycloheximideontheproductionofreplicative RNP complex. A culture of infected cells
wassplitinhalfat 4.5hp.i.,and toone-half ofthe culture cycloheximide was added to a final concentration
of100
tLg/ml.
After 15min in thepresence ofthe drug, [3H]uridine wasadded to each culture to afinal concentration of 100 ,aCi/ml. After a 5-min incubation, the cells were removed by centrifugation andcytoplasmicextractswerepreparedasdescribed in the text.The extracts were layered onto separate identical
15 to50%Renografin gradientsand werecentrifuged for16 hat23,00( rpm and 4°C in anSW27 rotor. The
radioactivity from the fractionated gradients was analyzed as described previously. Symbols: -4,
untreatedcontrol;0---S cycloheximide-treated sample.
ThenascentRNPswerefurther analyzed by
centrifugingfractions from thepeakandtrailing
fractions of peak I in CsCl gradients (Fig. 5).
The nascent RNPs had identical equilibrium
densities inCsCl. This result alsosuggeststhat
the nascent RNPs have a protein-RNA ratio
sunilartothat of virionnucleocapsids, since all
of the radioactivity was found at a density of
1.31
g/cm3
(19). The nascent RNA was thenremoved from the RNPs and analyzed in
su-crose-sodium dodecyl sulfate gradients, which
showed that the RNA contained in the
CsCl-banded RNPswasshorter than fulllength and
notjust42S RNA (Fig. 6). The demonstration
thatnascentRNPsarestable inCsCl is further
evidence suggesting that these nascent RNPs
arederived fromareplicativeintermediate since
transcriptive productsdonotband inCsCl (19).
Itthusappearslikelythat theheterogeneity of
the 5-min-pulse-labeled RNP complex which
hasbeen isolated is not due to adifference in
densitybetween the RNPs in the peakand the
trailingfractions.
Examination of the trailing fractions of
peak I. The results shown in Fig. 1
demon-strated that the pulse-labeled material in the
trailingfractions didnotaccumulateduring long
pulse-labeling periods. Furthermore,it hasbeen
shown thatnascent RNA is found in both the
peak and the trailing fractions of peak I, but
detectable newly replicated 42S molecules are
found in the peak fractions only (Hill et al.,
submitted for publication). These results
per-haps suggest that the material in the trailing
fractions isnascentRNA which has dissociated
from thetemplateRNA. We have observed that
thenonequilibrium Renografin gradientsresolve
RNA molecules on the basis ofsize; thus, it is
possible that the trailing fractions contain
smaller RNPsseparatedonthe basis of size.
At the present time there are two possible
waysthat thisproblemcanbestudieddirectly.
Electronmicroscopic examinationof the
mate-rial contained in thepeak andtrailingfractions
wasperformed; however, duetothelarge
num-ber of nonradiolabeledstructurespresentin the
same area aspeakI,wecouldnotprovethat the
trailing edge representednascentstrands
disso-ciated fromtheir templates (C. Naeve,
unpub-lisheddata). We didobservepossiblereplication
formsinthepeak fractions (Naeve, manuscript
inpreparation),but duetothehigh background
of nonreplicating RNPs, it was impossible to
deal conclusively with the problem of nascent
strand attachment to template RNP. Another
wayofdirectly showingthat thepeakfractions
on November 10, 2019 by guest
http://jvi.asm.org/
500 SIMONSEN, HILL,
7d - E
5 3 5 20 25 30 35 5 1015 20 25 30 35
E FF C
4-
~~~~~~~~~~~~8-- -1.20
2- ..
4--1.10
[image:7.504.117.405.66.343.2]25 30 35 40 45 50 55 5 10 '15 20 25 30 '35 FRACTION NUMBER
FIG. 4. EquilibriumRenografindensitygradientcentrifugation ofthe 5-min-labeled RNP complex. Infected cells at 5 hp.i. were labeled bythe addition of100,LCi each of[3H]uridine and [3H]adenosineper ml.
Crushed,frozenmedium was added to thesampleafter5min in thepresenceofthelabels,and the cells were
removed bycentrifugation. A cytoplasmic extract wasprepared andlayered onto a 15 to50%Renografin gradientwhich wascentrifuged for16h at 23,000 rpm and4°C in an SW27 rotor. The radioactivity was
quantitated asdescribed in thetext.Thefractionsindicatedfiomthepeakandtrailingfractionsin(A) were
combined and diluted. Samples were layered onto identical 20 to 60% Renografin gradients and were
centrifugedat23,000 rpm and4°CinanSW27rotorfor17h(B)40h(C),or80h(D).3H-labeled42S RNA (E)
andpoliovirusnucleocapsidand top component(F) werecentrifugedin identicalRenografin gradientsat
23,000 rpm and4°C for80h.
8-A PeokI PeakH2-B C D
g |~12"23':~ 5 1"
6- *140
E
.4 , , , . , 2 . , 5 130
2- .120
5 10 152025 30 35 5 10 15 5 8 15 5 10 15
FRACTION NUMBER
FIG. 5. CsClgradient centrifugation of peak Ifrom a Renografin gradient. Acytoplasmic extractfrom
infected cells pulse-labeled for 5 min at 5hp.i. was layered onto a 15 to50% Renografin gradientand
centrifugedasfor Fig.1. The RNPpeakwassplit into three poolsasindicated in(A).Thepoolswerethen
layered onto 20 to40%(wt/wt)CsClgradients in TNEbufferandcentrifuged for16hat33,000 rpm and4°C
in an SW41 rotor. Fractions (0.5 ml) were collected from the bottom, trichloroacetic acid-precipitable
radioactivity was determinedfor250itlofeachfraction, and thedensitywasrecorded bymeasuring the
refractiveindexesof thefractionswith a Bausch and Lombrefractometer. (B)CsClgradientofpool1from
(A).(C) Pool 2. (D) Pool 3.
on November 10, 2019 by guest
http://jvi.asm.org/
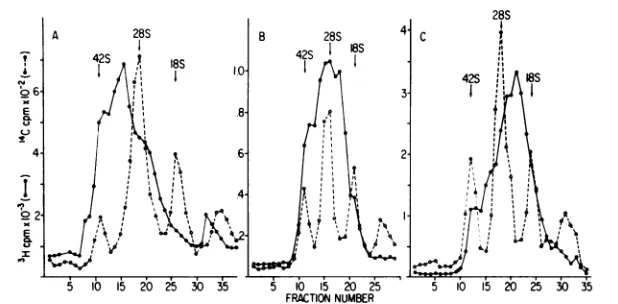
[image:7.504.87.428.448.582.2]FRATIONNUMBER
FIG. 6. Size analysis of the pulse-labeled RNA contained in CsCl-bandedreplicative RNPs. The pulse-labeled RNPs which were banded inCsClinFig.5B, C,and D werephenolextracted,and the RNAcontained inthesecomplexeswascentrifugedin 15 to30%sucrose-sodiumdodecylsulfategradientsfor16 hat20,000
rpm in an SW41 rotor. Trichloroacetic acid-precipitable radioactivity was determined for each of the
fractions,which were collectedby drippingfromthe bottomofthe tube.Symbols:0,['4C]uridine-labeled42S,
28S, 18S, and 4S markers; *, [3H]uridine-labeled RNA from Fig. 5B, C, and D (panels A, B, and C,
respectively).
contain nascent strands attached to their
tem-plate is tocompletethe nascent strands invitro;
however, at the present time this has not been
possible.
It wastherefore necessary to study this
prob-lemindirectly.Asizeanalysisof the RNPs
con-tained in the peakandtrailing fractions of the
Renografin gradient was attempted by using
sucrosegradients,but due to aggregation of the
RNPs when removed from Renografin, the
re-sultswereinconclusive.Thesizingof the RNPs
was nextattempted by comparing the
sedimen-tation characteristics of RNPs from the peak
andtrailing fractions ofpeakI tothose of RNPs
derived from a strain of defective-interfering
(DI)particles containingagenomewhich isonly
15%of thewild-typegenome. We reasoned that
the DI RNPs , which also band inCsClat 1.31
g/cm3 (M. Leppert, personal communication),
should be found in the sameregion of a
none-quilibrium Renografin gradientasnascentRDYPs
similar in sizetothe DI RNPs iftheRenografin
gradients could separate RNPs on the basis of
size. Figure 7 shows that the nucleocapsids
de-rived fromthe DIparticles are found in exactly
the sameregionofanonequilibriumRenografin
gradientas nascentRNPs, which are about 15%
replicated (Fig.7A and B; density, 1.18g/cm3).
Virion nucleocapsids cosedimented with the
peak fractions ofpeak I at a density of 1.22 g/
cm3 (data not shown). TheDIRNPspresumably
have the same structure as the nascentRNPs,
sinceDI RNPsandnascent RNPs have identical
densitiesin both isopycnic Renografin (Fig. 7C)
andCsCl gradients. This suggests, but does not
prove, that the RNPs contained in the trailing
fractions are dissociated from their templates
andarebeingseparatedfrom thepeakfractions
onthe basis of size.However,we cannotexclude
thepossibility that conformational factors cause
someof the nascent RNPs to move more slowly
through the Renografingradient, thus forming thetrailingedge.
Effect of Renografin on transcribing
complexes. The total intracellular viral RNPs
consist oftranscribingcomplexes,nonenveloped
virionRNPs to be assembled into virus particles,
andreplicatingcomplexes. Previous studies have shown that intracellular RNPs cosediment with
virion RNPs insucrosegradientsas120S to 140S
molecules. NascentmRNAcanbe detected
as-sociated withsomeof these intracellular RNP
complexes; thus,itis
likely
that theassociationof nascent product strands to template RNA
doesnotgreatlyaffect the sedimentation
coeffi-cient of the transcribing complex (31, 35). We
wanted to determine whetherreplicative
com-plexeswerefound in the 140Speak,aswellas to
determinethe effect ofRenografinon
transcrib-ingcomplexes.
A 5-min-pulse-labeled cytoplasmic extract waspreparedfrom VSV-infected cellsat5hp.i.
The extractwas divided into twosamples; one
was centrifugedin a 15 to 47%
Renografin
gra-dient (Fig. 8A), and theother samplewas
cen-trifuged in a 15 to 30% sucrose gradient (Fig.
8B). The peak fractions from the Renografm
gradientin Fig. 8A, whichcontained the
repli-cativecomplex,werecombinedasindicatedand
recentrifugedin a 20 to47%Renografin gradient
on November 10, 2019 by guest
http://jvi.asm.org/
502 SIMONSEN, HILL, AND SUMMERS
3
2-aE~~~~~~~~~~~~~~~~~~~~~~~a
C1.246 -1.30
6-61.20
4
[image:9.504.58.246.52.384.2]FRACTIONNO
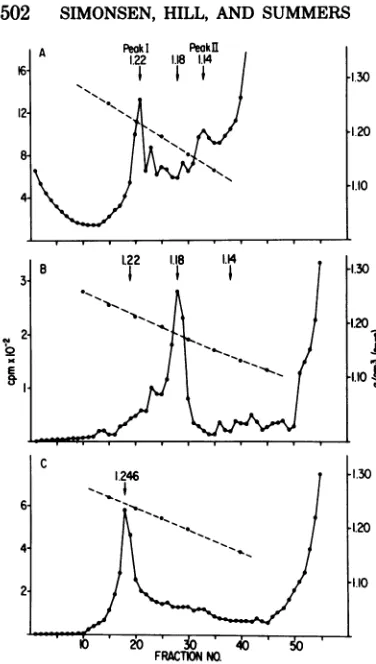
FIG. 7. Comparison of RNPsderivedfrom
DIpar-ticdeswith thereplicativeRNPcomplexes.3H-labeled
DIparticles ofthe MS-T strain werepurified byrate
zonal sedimentation insucrosegradients. The
puri-fied preparationwasadjustedto aconcentration of
0.2% deoxycholate-0.3% Nonidet P-40 in NTbuffer
anddivided into twosamples. Onesamplewas
lay-ered ontoa 15to50%Renografin gradient,andthe other was layeredonto a20to60%gradient. Both
werecentrifugedat23,000 rpm and4°Cin anSW27.1
rotor.FractionswereanalyzedasdescribedforFig.
1. (A) Cytoplasmicextractfrom 5-min-pulse-labeled
cellscentrifugedina 15 to50%Renografin gradient for16h at23,000 rpm in theSW27.1 rotor. (B) DI
RNPcentrifugedfor16h in the15to50%Renografin
gradient.(C)DIRNPcentrifuged for80h in the 20 to
60%Renografin gradient. Virionnucleocapsidswere
foundtocosediment withpeak I(fractions 19 to 23)
atadensity of 1.22g/cm3(datanot shown).
(Fig. 8C). All of the radioactivity wasfound in
theareaofpeak I, andno material was found in
the region of the gradient where the nascent
mRNA species have been isolated (peak II).
Fractionsfrom differentregions ofthe 140S peak
in thesucrosegradient (Fig.8B)were also pooled
asindicated andrecentrifugedonidentical20 to
47% Renografin gradients (Fig. 8D, E, and F).
The results of these three gradients were
vir-tually identical; in addition to the material at
peak I, a peak at the position of peak II was
observed. Although transcription accounts for
nearly 90% of the total RNAsynthesis in
VSV-infected cells (38), peak II does not represent
90%of theinputradioactivity because the fully
synthesizedmessages arefoundatthetopof the
nonequilibriumgradients asfree RNA and not
inpeak II (datanotshown). These results
dem-onstratethatthe 140S RNPs isolated insucrose
gradients contain bothtranscribingand
replicat-ing RNP complexes in the same proportions
throughout the 140S peak and that replicating
products can be separated from transcribing
products in Renografin gradients due to some
undetermined compositional differences or
structuraldifferencesorboth.
DISCUSSION
The ability of the VSV RNP core isolated
from virions to transcribe in vivo and in vitro
has beenwell documented (1, 2,7,20, 25-27,32).
Thetranscriptaseenzymehasbeen showntobe
composed of the L and NS proteins, and the
template has been demonstrated to consist of
minus-strand 42S RNA and N protein (17, 18,
28). Theenzyme isinactive with 42Sdevoid of
proteinorin thepresenceofantibody directed
againstthe Nprotein-RNAcomplex (11, 17). It
thus seems well established that the template
fortranscription isanRNPcomplex.Ithasbeen
assumed that thetemplateforreplicationisalso
anRNPcomplexonthe basis ofgeneticstudies with temperature-sensitive mutants. Several
mutantsingroupI(Lprotein)andgroupsIIand
IV (presumablyNand NS proteins) have
tem-perature-sensitive replicative abilities (15, 16,
30). Direct evidence for the role of theseproteins
inreplicationislackingduetothe absence ofan
invitroreplicationsystem. The datapresented
aboveand in anotherreport(Hilletal.,
submit-ted forpublication)supportthehypothesisthat
VSVreplicationoccurs on anRNPtemplate.
The nuclease resistance and density of the
nascentRNPcomplexesareindications that the
nascent RNA species found in the RNPs are
quickly associated with protein. Preliminary
re-sults from this laboratory have indicated that
oneof theproteins whichrapidly associates with
thecomplex is indeed the VSV Nprotein. The
association of thisprotein with thenascentRNA
isintriguing, since thismay betheprocess with
which cycloheximide interferes, and thus
in-hibits, replication. The strong and stable
asso-ciation ofN protein with 42S RNA (9) is not
seenwith theVSVmRNAspecies,eventhough
the nucleotide sequences ofthe mRNA are a
subset of the plus-strand 42S RNA sequence.
The "leader"RNA,which isidenticaltothefirst
I-I-B
J. VIROL.
,<2q 1,0 L
on November 10, 2019 by guest
http://jvi.asm.org/
FIG. 8. Renografingradientcentrifugation of RNPs obtained from Renografin and sucrose gradients. A
5-min-pulse-labeled cytoplasmicextractwasprepared as described for Fig.1.Thesample was divided; one-half
waslayeredonto a 15 to50%Renografin gradient, and theother halfofthesample was layered onto a 15 to
30% sucrosegradient. TheRenografingradient wascentrifugedfor 16 h at 23,000 rpm and 4°C in an SW27 rotor, and the sucrosegradient was centrifuged for 16 h at 16,000rpm in thesame rotor. Fractions were removedfromboth gradients and recentrifuged in 20 to47%Renografin gradientsfor 16 h at 23,000 rpm in an SW41 rotor. (A) Cyplasmicextractcentrifugedin a 15 to50%Renografin gradient. (B) Cytoplasmicextract
centrifugedin a 15 to30% sucrosegradient. (C)Pooledfractions from Renografingradient in (A) rerun in a
20 to47%Renografin gradient. (D)Fraction18fromthe sucrosegradientin(B)recentrifugedin a 20to47%
Renografin gradient.(E) Fraction 21from (B) recentrifugedin a 20 to47%Renografingradient.(F)Fraction
24from (B) recentrifugedin a 20 to 47%Renografin gradient.The sedimentationpositionsofthe60S and 40S
ribosomalsubunits and the virion RNP are indicated in(B) bythearrows.The materialin(C), fractions5 to
7, which wasfound at agreaterdensitythanpeak I,represents aconformer ofpeakI. The natureofthis materialispresently beingexamined.
48 nucleotides atthe 5' end of the plus-strand
42SRNA (13, 14) isnotfound in themessages
and thus couldcontainasequencewhichacts as
abinding site for the N protein. However, the
leader isnotfound in the infected cellas ashort
RNP, which would be expected if the leader
alone were the signal for the association of N
protein with the nascentRNA (Simonsen,
un-published data). The addition of N protein to
the nascentRNA maywell bean integralpart
ofreplicationwhich is associated with either the
replicase enzyme or the template. This would
accountfor thenuclease resistance anddensity
inCsCl ofthe nascentstrands. Wearepresently
examining
the mechanism by whichcyclohexi-mide inhibitsreplication,aswellasthekinetics
of protein association with the nascent
repli-cated RNA in orderto clarifythe role that N
proteinplaysinreplication.
These datamay alsosuggest that most of the
nascent RNPs remain associated with their
RNPtemplates.We have observed that thevery
homogeneous Renografin peaks from samples pulse-labeledfor15and30min(Fig. 1CandD)
and the peak fraction ofa5-min-pulse-labeled
sample (Fig. 6A) contain RNA ranging in size
from 42S downtoless than18S,but thetrailing
fractions ofpeak I contain primarily less than
full-lengthRNA(Fig.60).Theobservation that
RNPs derived from DIparticles which have a
genomeapproximately 15% of thewild-type
ge-nomecosediment with nascentRNPsfrom the
trailing edge, which are also 15% completed,
leads us to believe that perhaps the trailing
fractions do containnascentRNPs whichare no
longer attachedtotheirtemplates.
Our resultsareconsistentinestablishingthat
nascent replicating RNA is found as part ofa
RNP. Wehavenotfoundsignificantamountsof
double-stranded RNA in thesecomplexes (Hill
et al., submitted for publication). We suspect
that double-stranded RNA molecules which
on November 10, 2019 by guest
http://jvi.asm.org/
504 SIMONSEN, HILL, AND
have been examined (33, 37) as possible
repli-cativeintermediatesareperhaps artifacts of
iso-lation. Since the structures we have isolated
appear tocontaintemplateandnascentstrands,
presumablylinked by thereplicative enzyme or
a small region of base pairs, the addition of
phenolto removeprotein wouldlikelyenhance
thehybridization of thesecomplementaryRNA
molecules which are alreadyincloseproximity
(23, 29). Itthusseemsreasonable toacceptthe
phenol-extracted double-stranded RNA
mole-cules as "collapsed" replicative complexes,
which probably approximate certain aspectsof
replication.
Theisolation ofareplicative complex, coupled
with the recent demonstration of full-length
VSV RNA synthesis in vitro (S.
Batt-Hum-phries,C. C. Simonsen, and E. Ehrenfeld,
Virol-ogy, in press), may now make it feasible to
develop anin vitro replication systemby using
the complexes isolated from Renografin. The
reconstitution ofareplication systemisclearly
required to fully understand the regulation of
replication and the role of host factors inVSV
replication, andwe arecurrentlyattemptingto
studythissystemingreaterdetail.
ACKNOWLEDGMENTS
Wethank Steven Humphries for the gift of labeled polio-viruspreparations, and we are grateful for the kind gift of VSVDIpreparations from Sue Moyer. The helpful discussions of D.Kolakofsky,0. Richards, S. Casjens, C. Georgopoulos, D.Carroll, and A. Larsen are also greatly appreciated.
C.S.is arecipientof aUniversityofUtah Graduate Re-searchFellowship.This work was supported by Public Health Service grant AI-12316-04 from the National Institutes of Health and by National Science Foundation grant PMC77-17867A01.
LITERATURE CITED
1. Abraham, G., andA. K.Banerjee. 1976. Sequential transcriptionof the genes of vesicular stomatitis virus. Proc.Natl. Acad.Sci.U.S.A. 73:1504-1508.
2. Abraham, G.,D. P.Rhodes,and A. K.Banerjee.1975. The 5'terminal structure of themethylated mRNA synthesizedin vitrobyvesicular stomatitis virus.Cell 5:51-58.
3. Atkinson,P.H.1975.SynthesisandassemblyofHeLa cellplasm membrane glycoproteins and proteins. J. Biol. Chem.250:2123-2134.
4. Atkinson, P. H., S. A. Moyer, and D. F. Summers. 1976. Assembly of vesicular stomatitis virus glycopro-tein and matrixprotein into HeLa cell plasma mem-branes. J. Mol. Biol. 102:613-631.
5. Bailey,J.J., and N. Davidson. 1976. Methylmercury as
areversibledenaturing agent for agarose gel electro-phoresis. Anal. Biochem. 70:75-85.
6. Baltimore, D., and A. S. Huang. 1970. Interaction of HeLacellproteins with RNA. J. Mol. Biol. 47:263-273.
7.Banerjee, A. K., and D. P. Rhodes. 1973. In vitro synthesisofRNA that containspolyadenylate by virion-associated RNApolymerase of vesicular stomatitis vi-rus.Proc. Natl. Acad. Sci. U.S.A.70:3566-3570.
8. Birnie,G.D.,D.Brickwood,and A. Hell.1973.
Buoy-antdensities andhydration of nucleic acids, proteins
J. VIROL.
andnucleoprotein complexes in metrizamide. Biochim. Biophys. Acta 331:263-294.
9. Bishop, D. H. L., and P. Roy. 1972. Dissociation of vesicular stomatitis virus and relation of the virion proteins to the viral transcriptase. J. Virol. 10:234-243. 10. Calaguiri, L. A., and H. Gerstein. 1978. Subclasses of ribonucleo-proteins in influenza virus-infected cells. Vi-rology 90:119-132.
11. Carroll, A. R., and R. R. Wagner. 1978. Inhibition of transcription by immunoglobulin directed against the ribonucleoproteinofhomotypic and heterotypic vesic-ular stomatitis viruses. J. Virol. 25:675-684.
12. Chan, R. T. L., and I. E. Scheffler. 1974. Isopycnic centrifugation of chromatin in Renografin solutions. J. Cell Biol. 61:780-788.
13. Colonno, R. J., and A. K. Banerjee. 1976. A unique RNAspecies involved in initiation of vesicular stoma-titis virustranscription in vitro. Cell 8:197-204. 14. Colonno, R.J., and A. K. Banerjee. 1978. Complete
nucleotide sequence of the leaderRNA synthesized in vitroby vesicular stomatitis virus.Cell15:93-101. 15. Combard,A., C. Martinet, C. Printz Ane, A.
Fried-man,and P. Printz. 1974.Transcriptionand replica-tion ofvesicular stomatitis virus: effects of temperature-sensitive mutations incomplementation group IV. J. Virol. 13:922-930.
16.Combard, A., C. Printz Ane,C. Martinet, and P. Printz.1977.Temperature-sensitivedefect of vesicular stomatitis virus incomplementation group II. J. Virol. 21:913-923.
17.Emerson,S.U., and R. R. Wagner. 1972. Dissociation and reconstitutionof the transcriptase and template activities of vesicular stomatitis virus B and T virions. J. Virol. 10:297-309.
18. Emerson,S. U.,and Y.-H. Yu. 1975. Both NS and L proteins arerequired for in vitro RNA synthesis by vesicularstomatitis virus. J. Virol. 15:1348-1356. 19.Grubman,M.J., andD. A.Shafritz.1977.Identification
and characterization of messenger ribonucleoprotein complexes from vesicular stomatitis virus-infected HeLa cells.Virology81:1-16.
20. Huang,A.S.,D.Baltimore, andM.Stampfer. 1970. Ribonucleic acidsynthesisofvesicular stomatitis virus. III. Multiple complementary messenger RNA mole-cules.Virology42:946-957.
21. Hunt,D.M.,S. U.Emerson,and R. R.Wagner.1976.
RNA temperature-sensitive mutantsofvesicular sto-matitis virus:L-proteinthermosensitivityaccountsfor transcriptaserestriction of group I mutants.J. Virol. 18:596-603.
22. Hutterman,A.,and G.Wendlberger-Schieweg.1976.
Studiesonmetrizamide-proteininteractions. Biochim. Biophys.Acta453:176-184.
23. Kohne,D.E.,S. A.Levison, and M. J.Byers. 1977. Room temperaturemethodforincreasingthe rate of DNA reassociationbymanythousand-fold: thephenol emulsion reassociation technique. Biochemistry 16:
5329-5341.
24. Kolakofsky, D. 1976.Isolation and characterization of Sendai virus DI-RNAs.Cell8:547-555.
25. Moyer,S.,and A. K.Banerjee.1975.MessengerRNA species synthesized in vitro bythe virion associated RNApolymeraseof vesicular stomatitis virus.Cell4: 37-43.
26. Mudd, J. A., and D. F. Summers. 1970. Polysomal ribonucleic acid of vesicular stomatitis virus-infected HeLacells.Virology 42:958-968.
27. Murphy, M.F.,and R. A.Lazzarini.1974.Synthesisof
viralmRNA and polyadenylate by an RNP complex from extracts of vesicular stomatitis virusinfectedcells. Cell3:77-84.
28. Naito,S.,and A. Ishihama.1976.Function andstructure ofRNApolymerasefrom vesicular stomatitisvirus. J.
on November 10, 2019 by guest
http://jvi.asm.org/
29. Oberg, B.,and L. Philipson.1971.Replicativestructures
ofpoliovirusRNA in vivo. J. Mol. Biol. 58:725-737. 30. Perlman,S. M., and A. S.Huang.1973.RNAsynthesis
ofvesicular stomatitis virus. V. Interactions between
transcription and replication. J. Virol. 12:1395-1400. 31. Petric, M., and L. Prevec. 1970. Vesicular stomatitis
virus-a new interfering particle, intracellular struc-tures,andvirus-specificRNA.Virology41:615-630.
32. Rhodes, D. P., S. A. Moyer, and A. K.Banerjee.1974.
Invitrosynthesis ofmethylatedmessengerRNAby the virion-associated RNApolymerase of vesicular
stoma-titisvirus.Cell 3:327-333.
33. Schincariol,A.L.,and A. F.Howatson. 1972. Repli-cation of vesicular stomatitisvirus.II.Separationand characterization ofvirus-specific RNAspecies.Virology 49:766-783.
34. Simonsen, C.C.,S.Batt-Humphries,and D. F.
Sum-rus-infected cells: in vivo regulation of replication. J. Virol. 31:124-133.
35. Soria,M.,S.P.Little, and A. S. Huang. 1974. Charac-terization of vesicular stomatitis virusnucleocapsids. I. Complementary 40S RNA molecules in nucleocapsids. Virology 61:270-280.
36. Wagner, R. R. 1975. Reproduction of rhabdoviruses, p. 1-93.In H. Fraenkel-Conrat and R. R. Wagner (ed.), Comprehensive virology, vol. 4, Plenum Publishing Corp., New York.
37. Wertz,G. W. 1978. Isolation of possible replicative inter-mediate structures from vesicular stomatitis virus-in-fected cells.Virology 85:271-285.
38. Wertz,G. W., and M. Levine. 1973. RNA synthesis by vesicular stomatitis virus and a small plaque mutant: effects ofcycloheximide. J. Virol. 12:253-264.
on November 10, 2019 by guest
http://jvi.asm.org/