JOURNALOFVIROLOGY, May 1978,p.522-531 Vol. 26, No. 2 0022-538X/78/0026-0522$02.00/0
Copyright©1978 AmericanSocietyforMicrobiology PrintedinU.S.A.
Interstrain Variation of the
Major Internal
Structural
Component
(p30gag)
of Two
Murine
Oncornaviruses:
Comparative
Immunochemical, Biochemical, and
Biophysical
Analysis
W. NEALBURNETTEt ANDWILLIAM M. MITCHELL*
DepartmentofMedicine, Albert Einstein College ofMedicine, Bronx,New York 10461, andDepartment of
Pathology,
VanderbiltUniversity
Schoolof
Medicine, Nashville, Tennessee 37232*Received forpublication 3 October1977
The
major
internal structural protein
(p30a9') of the Moloney leukemia virus
and theendogenous Y-1 murine oncornavirus was examined for biochemical and
biophysical manifestations of interstrain
antigenic variation. Although the
twoviral
proteins
share murinegroup-specific
antigenic determinants,
the Y-1
virus
p30
appeared
tohaveboth a lowerrelative
number of such determinants and
adecreased
affinity
atthe cross-reactive sites forMoloney
virusp30
monospecific
antibodies.
Further, immunological
analysis
indicated the presence ofunique
antigenic sites on the Moloney virusp30 not shared
by
theanalogous
Y-1 virusmolecule. Thetwo
polypeptides copurified
andhadsimilar
isoelectricpoints (pH
6.2 to
6.3) and sedimentation coefficients
(2.47S).
However, equilibrium
sedimen-tation
yielded
asignificant
mass differencebetween the twoproteins,
28,300
±600and
31,000
± 300daltons for theMoloney
and Y-1 virusmolecules,
respec-tively.
Amino acidanalysis indicated
aconcomitant increase
intotal
residues for
the Y-1 virus
p30, although
anumber ofresiduesappeared
tohavebeen conserved
between the two viral
proteins.
Conformationalstudies
andhydrodynamic
cal-culations demonstrated marked
secondary
andtertiary
structuraldifferences;
with the
Y-1virus
p30
being
anasymmetric
prolate
ellipsoid containing
27to28%
a-helix and
Moloney
virusp30being
somewhatmorespherical
andpossessing
ana-helical
content of 50to 55%.Two-dimensional
mapping
of'2II-labeled
tryptic
peptides
of eachp30
suggested
thatconsiderable
sequenceheterogeneity is
responsible
formany of thebiophysical,
biochemical,
andimmunochemicaldif-ferences in these twoanalogous structural
proteins.
The
provirus
genomeof mammalian
oncor-reactive determinants
inthe
greatest relativenaviruses
code
for anumber
ofstructural
poly-
concentration.
Itdoes, however,
alsopossessde-peptides (2) whose
primary
and
secondary
struc-monstrable
type-specific reactivity,
and
p30
mol-tures
immunologically
define the
interspecies
ecules from various strainsof murine virus
ex-(13, 14,
41),
species-specific (15,
18,28, 30-33,
36,hibit
unique antigenic
determinants (42). Unlike
42),
andtype-specific (17,35,39,42,44)
antigenic
the
hemagglutinating
and
interference
proper-reactivities
of various virus strains
ingeneral,
ties of
gp69/71enV (6,
20), the RNA-dependent
and
analogous proteins
shared amongstrains
in DNApolymerase activity
of the virion reverseparticular.
Themajor internal
structuralprotein
transcriptase
enzyme(3,
46),
and thenucleic
of the murine oncornaviruses is the
approxi-
acid-binding ability
ofp12
(37)
and plO(12;
M.mately
30,00-dalton polypeptide,
p309ag(2,
8, 9,Schulein,
W. N.Burnette,
and J. T.August,
J.18, 26, 30, 36). The p309ag and the
polypeptides
Virol.,
inpress),
thep3O
hasasyet nodefinitive
p155a5, p125a5,
andp109a9
arecleavage products
function,
although
itsinvolvement
in the processof a
65,000-dalton
polyprotein
precursor (1, 38,of
hostrange restriction hasbeen
suggested (19), 39),Pr65SaS,
which is itself theproduct
of theand,
because of itsproclivity
forself-association,
gag(group-specific
antigen)
gene constituent(4,
itmayplay
a role in viralcoreassembly
atthe5) of the
integrated
provirus
genome(45).
The cell membrane(8).
p30
hasbeen termed thegroup-specific antigen
We
havepreviously
described
a number of(15) because it
possessed
murine-specific
cross-biophysical
andbiochemical
properties
ofp30
tPresent address: Fred Hutchinson Cancer Research Cen- from the laboratory strain of Moloney murine
ter,Seattle,WA 98104. leukemia
virus
(8).
In thisreport
we compare522
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 ONCORNAVIRUS p30"a9 INTERSTRAIN VARIATION 523
many of
those
properties
with the p30 of the Y- the slices wereminced;and theproteins wereeluted
1
N-ecotropic endogenous murine
oncornavirus overnight at370C
in a buffer containing 50 mM(9)
toelucidate the
primary and secondary struc- NH4HCO3(pH8.3), 0.05%sodiumdodecyl sulfate,andtural
characteristics
responsible for theantigenic
1 mMphenylmethylsulfonyl fluoride. Proteinrecoveryvariation exhibited
by
thisevolutionarily
con- wasroutinely 80 to 90% by thisprocedure. The eluantsserved
polypeptide.
werecentrifuged briefly
toremovegel
debris,
KCl
wasaddedtothe supernatants to a final concentration of 0.2 M,and, after 15min onice, the precipitates were MATERIALS AND METHODS
collected
by
centrifugation
(7).
Theprecipitates
werewashed once with ice-cold acetone containing 0.1 N
Celis and viruses. Moloney leukemia virus was HCI, washed again with acetone alone, and dried.
propagatedinhigh-passage-levelSwissmouseembryo They were suspended in20
Al
of 0.5 M sodiumphos-cells maintained in Eagle minimal essential medium phate (pH 7.5)containing 0.1% sodium dodecyl sulfate, with 10% fetal calfserum.Murine adrenal carcinoma heated for2 min at 100°C, and allowed to cool. The cells(clone Y-1, ATCCno.CCL-79) producing the N- proteinpreparationswerethen radioiodinated by the tropic Y-1 virus were grown in Ham F-10 medium chloramine T method (21), and free radioiodine was containing15% fetal calfserumand 2.5% horse serum. removed from the quenched reaction mixtures by pas-Virus was harvested and purified as previously de- sageovercolumns(0.8by14.5cm) ofSephadex G-25
scribed (9). superfine equilibratedinabuffer of 1.0M
Tris-hydro-Antisera. Heterospecific goat anti-Moloney leu- chloride (pH 8.6) with 0.1% sodium dodecyl sulfate. kemia virus (1S-166) and goat anti-AKR virus (2S- Reduction and S-aminoethylation were performed by 296) sera were obtained through the courtesy of R. modificationsofpreviously published procedures (10, Wilsnack, Huntingdon Laboratories, Md. Monospe- 11).Briefly, theradioiodinated proteins from the G-cific antiseraagainst p30wereobtainedby hyperim- 25-excluded volumes (1 ml each) were reduced by munization of rabbits with the purified protein (see addingdithiothreitolto afinal concentration of 0.1 M,
below) incompleteFreundadjuvant followedbytwo flushing the reaction vessels with N2, capping them,
fortnightly boosters in incomplete Freund adjuvant. and incubating them for3 h at 370C. The reduced
Goat anti-rabbit immunoglobulin G was purchased proteinpreparationswerethenS-aminoethylated by fromCalbiochem(LaJolla, Calif.). thesequential addition ofone10-,lI portion and two 5-p30 Purification. 5-p30was purifiedunder nonde- ,ul portions ofethyleneimine (ICN Pharmaceuticals, naturing conditions fromcorepreparations of virus by Inc.)at10-min intervals underaN2 atmosphere. A
50-phosphocellulose ion exchange andSephadex G-100 jig quantity of carrier bovine immunoglobulin was
(Pharmacia) gel filtration chromatography as de- addedtoeachpreparation (16),and the mixtures were
scribedbyStrand andAugust (43).Column effluents precipitatedat4°Cwith 10%trichloroaceticacid.The
were monitored for relative fluorescence at 287 precipitates were collected by centrifugation and
nm/348nm. washed twicewithice-coldacetone.Thedried
precip-Gelelectrophoresis.Sodiumdodecylsulfate-poly- itateswere
resuspended
in50,lt
of50mM NH4HCO3acrylamide gelelectrophoresiswasperformed in cylin- (pH 8.3), 1 ,ug of tolylsulfonyl phenylalanyl
chloro-drical 10%polyacrylamide gelsaspreviouslydescribed methyl ketone-treated trypsin (Worthington
Bio-(8). chemicals) was added to each suspension, and the
Radioiodination.Purifiedp30wasradioiodinated preparationswereincubated for3to4hat370C.An
forradioimmunoassaybythechloramine T method of additional 1,ug oftrypsinwasaddedtoeach
prepara-Hunter(21)andchromatographedonSephadexG-25 tion,and incubationwascontinuedovernight.
Diges-superfinetoremoveunbound radioiodine. tionwasroutinelyfoundtobemorethan 90%complete
Immunodifusion
andradioimmunoassay. Im- as measuredby
inclusion onSephadex
G-25. Aftermunodiffusion wasperformed in 1% Difco agarcon- briefcentrifugationtoremoveinsolublematerial,the
taning100mMNaCl,1mM
EDTA,
and10mMTris-samples
werespotted
atonecornerofPolygram
Celhydrochloride(pH 7.2) for24 to 72hatroomtemper- 300plastic-backedcellulose sheets(Brinkman
Instru-atureand thenfor24hat4°C. Thegelswerewashed ments). The sheetswerewetted withapH3.5buffer
extensivelyinphosphate-buffered saline and stained ofpyridine-aceticacid-water(1:10:89)and
electropho-withCoomassie brilliant blue R-250. Competition ra- resed under Varsol
(Savant
Instruments, Inc.)forap-dioimmunoassay was performed as described by proximately30 min at1,000Vwith acid fuchsinas a
Strand and August (42); unlabeled p30 competed migration marker. The sheetswere dried and chro-againstradioiodinated p30for antibodies inamono- matographedatascendingright anglestothe direction
specificantiserum dilution sufficienttobindapproxi- ofelectrophoresis inanatmosphere-equilibrated tank
mately50% of thetrichloracetic acid-precipitablein-
containing
apH
5.3 buffer of n-butanol-acetic putradioiodinatedprotein.Precipitationwaseffected acid-water-pyridine (15:3:12:12). The sheets were bytheaddition of goat anti-rabbitimmunoglobulin G, again dried,and autoradiography wasperformed byand theprecipitatewaswashedextensivelytoremove exposuretoKodakno-screenmedicalX-rayfilm.
nonspecific radioactivity. Radioactivityin theprecip- Other biochemical and biophysical analyses.
itate was determinedby gamma counting. Extinctioncoefficient determinations, liquid column
Peptideanalysis.Y-1andMoloney leukemiavirus andpolyacrylamide gelisoelectricfocusing,
sedimen-p30 bands, each containing approximately 5 ug of tation equilibrium andvelocity analysis, amino acid
protein,were cutfrom driedCoomassiebrilliant blue- analysis, and far-UV circular dichroism were per-stained sodium dodecyl sulfate-polyacrylamide gels; formedaspreviouslydescribed(8).
on November 10, 2019 by guest
http://jvi.asm.org/
524 BURNETTE AND MITCHELL J. VIROL.
RESULTS
5
Purity of
the protein
preparations.
Frac-0
xtionation
of the
solubilized
coreproteins
from
the
Moloney and Y-1 viruses
onphosphocellu-
z
40lose
with
acontinuous
gradient
of
0 to 1.0M
+KCI revealed
amajor
peak for both viruses
atabout 0.2 M
(Fig.
1).
When thesepeaks
were 3subsequently
rechromatographed
by gel
filtra- ! 30tion,
asingle
homogeneous
peak
wasobserved
wfor each
preparation
inthe
30,000-molecular-
zweight region of the column effluent
(Fig. 2).
' 20-Electrophoresis
ofthese
peak
fractions
in the lpresence
of sodium
dodecyl
sulfate
demon-
°strateda
single
bandfor eachp30 preparation
at J labout
30,000
daltons
(Fig. 3),
although
it
is
ap- w 10parent
that the
Moloney
virus
p30
migrates
Pslightly
faster, and thus
has alower
molecular
-Jmass,
than the
corresponding
Y-1 virus
protein.
Isoelectric
focusing
in
asucrose-ampholyte
col-
20 40 6080
umn(Fig. 4)
yielded
anisoelectric
point
of
ap-FRACTION
NUMBERproximately pH
6.3for
both viral
proteins.
The
FIG 2.Purification of
themajor
structuralprotein
minor peak
seen as ashoulder
atabout
pH
7of
ofteMoloney
theI
2. P andanYmno
Y-1
murine
eoncrnaviruses
oncornaviruses
by may beanalogous
to theheterogeneously
molecular sievechromatography.
The peak fractionscharged species of p30 observed
by
Oroszlanet elutingfromphosphocellulose
atapproximately
0.2al.
(33)
incomplement
fixationassaysof isoelec-
M KCI were concentrated by precipitation withtric-focusing
column effluents.
High-resolution
(NHJ)2S04
at 75%saturation andchromatographed
inparallel columnsof Sephadex
G-1X00
inabuffer of1.0M
NaCl,
1 mM EDTA, and 10mMN,N-bis(2-If,
I I | I |.hydroxyethyl)-2-aminoethanesulfonic
acid (pH6.5).
O Moloney leukemia virusprotein (A);Y-1virusprotein
x
().
Z40_
30-
_I.O
0~~~~~~
S
_
_-
C0
BOVINE
ALBUMIN-o-
J
z
(67,000)
w
20C
2 6 1 0w 0
U.
10
CYToCHRioMEC--
ma sU
a>
~~~~~~~~~(12,4
00)
-aJ
20
406080 100
1 2
34 56
FRACTION NUM BER FIG. 3. Sodiumdodecyl
sulfate-polyacrylamide
gel
FIG. 1. Ion-exchange chromatographic fractiona- electrophoresis ofthemajorstructuralproteinofthe tion oftheproteinsof Moloneyand Y-1 murineon- Moloney and Y-1 murine oncornaviruses. Electro-cornaviruses. Moloneyleukemiavirusand Y-1 virus phoresis was performed in 10%polyacrylamidegels weredisruptedin Triton X-100, the soluble proteins in thepresenceof 0.1% sodium dodecyl sulfate and 3
were appliedtoparallelcolumnsofWhatman P-11 Murea asdescribedin thetext.
(1)
and(6), molecularphosphocellulose, andp30waselutedwithalinear weightmarkers; (2) wholedisrupted Moloney
leuke-gradient of0 to 1.0 MKCI in 10 mM N,N-bis(2- miavirus; (3) purified Moloney virusp30;(4)purified
hydroxyethyl)-2-aminoethanesulfonic acid (pH 6.5), Y-1virusp30;(5) wholedisruptedY-1virus.Protein
and1 mMEDTA.Moloneyleukemia virusproteins bandswerestained withCoomassiebrilliantblue
R-(A); Y-1 virusproteins(0). 250.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.509.274.466.66.275.2] [image:3.509.85.249.346.561.2]VOL. 26, 1978 ONCORNAVIRUS p30gag INTERSTRAIN VARIATION 525
<,
zT115.0
~ ~ ~ ~ ~ 80z 6p
LL0
10 20 30 40 50
FRACTION NU MBER
FIG. 4. Columnisoelectric focusing of the purified
major structural protein of the Moloney and
Y-l1
,/\ 26murine oncornaviruses. The viral proteins were fo- \
cused in a 1.2%XampholytepH gradient of 3.5 to 10at/\
4°C. The pHgradient was stabilized with a 0 to40%o4
sucrose gradient to prevent convection, and focusing n3)
was at 500 Vfor the time indicated. Moloney leukemia
pH(A.viu poein 268
hC0;Ylvrsprti,9
;
(g B P39Wfocusing in a
polyacrylamide-ampholyte
matrix
_________________
(not
shown) demonstrated a difference of aboutFG.5Im
ndifsoaalisfthmune
0.1 pH unit between the isoelectric pointsofthe
spcFIc.
crs-eatvtImndfsonalsOf
Mooethemn
-iusie
two e . w p30protens.
Immunodiffusion
wasperformed asde-being the slightly more basic component. No scribed in the text with the purified viralcomponents.
evidence of minor speciesin these gelswas ob- JS-166,goat anti-Moloney virus serum; 2S-296, goat
served.
Thus, the minor chargeheterogeneity
anti-AKR virus serum. Gels were stained withCoo-obtained with column isoelectric focusing may massie brilliant blue R-250. (A) Immunodiffusion
secondary to self-association (8) at the higher plate; (B) illustration ofimmunodiffusion plate to
concentrations required
for analysis. highlight spur.Immunological
analysis.
Inimmunodiffu-sion (Fig. 5), the
Yim andite Moloney
virus p30polypeptides
extensively cross-reacted withboth_______________
anti-Moloney
virusand
anti-AKRvirushetero- _specific
sera. Thepresence
of a small spurin the
Z aprecipitin
bandpointing
toward the Y-1 p30 wellm 80with the anti-Moloney
virusserum
and the ap-
FIG. 5 Iparent
absence
ofspurs in the
cross-reaction 60 _ a YI ruswith the
antiserum
directed against AKR virus,5
p \ xa
virus verysimilar
to
Y-1 (9), indicated the j; X 40 - w \presence
ofdetemninants
on theMoloney
virus s 2 goap30 not shared by
the
Y-1 virusprotein.
In XiAR20 _ \competition
radioimmunoassay
withmonospe-lateXto
cific
anti-Moloney
virus p30 serum, thediffer-ences
between thetwo
proteins became more 'l-5
loo
o-obvious
(Fig. 6). Although
Y-1 p30 couldCOMnyiuNGPROTEINmg)
com-pete
with
Moloney p30, indicative ofthe
group-specific
and type-specific cross-reactivity ob- FIG. 6. Analysis of the antigenicdeterminants
ofserved inimmunodiffusion, the
displacement
of the p30 of Moloney and Y-1 murine oncornaviruses.ed
cpeition
c thedslo
Competition radioimmunoassay was performed witht sa final dilution of monospecific anti-Moloney virus
relative
to the homologousMoloneyvirus
pro-p30serum
50%4
sufficient to precipitate approximately tein competition, and the failure of theY-1
Of theradioiodinatedMoloney
p30 intheassay. Thepolypeptide
to compete fully for all the antibody amount ofcompetingproteinis asindicated.Moloneyspecies
in theanti-Moloney
p30
serumpopula-
virus
p30(-);
Y-1 virus p30().
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.509.55.250.51.221.2] [image:4.509.261.451.73.330.2] [image:4.509.265.454.448.582.2]526
BURNETTE AND MITCHELL J. VIROL.tion
clearly
demonstrated unique
determinants formed in the analytical ultracentrifuge, withpresent on
the
Moloney virus p30 that
werenot
initial loading concentrations and ionic strength
exposed
onthe
analogous
Y-1protein.
favoring the monomer. Under these
conditions
Amino acid
composition.
The amino acid thes'20w
for both the Y-1 andMoloney virus
p30
compositions of the Moloney virus and the Y-1
proteins
wasfound
tobe
2.46 to2.47S
(data
notvirus
p30 polypeptides
arepresented
inTable 1.shown). However,
high-speed,
meniscus-deple-The best integral fit of the recovery data was
tion
equilibrium sedimentation revealed
asig-obtained with three residues of histidine
for the nificantdifference
in the massof
the
twopro-Y-1
and four residues for the Moloney
virus p30. teins(Fig. 7),
confirming the
initial observation
Because of its
greater
apparent mass, the Y-1 in sodiumdodecyl
sulfate-polyacrylamide
gels.virus
polypeptide
was found tocontain
more Using partial specific volumes of 0.719 ml/g forresidues than
the Moloney protein. However, Moloney virus p30 and 0.722 ml/g for Y-1p30
the
total number
oflysine and alanine residues (determined from amino acid analysis) for theappeared
tohavebeenconserved, and histidine calculation of weight average molecularweights
and
methionine were actually in excess in Mo- and extrapolating to zero concentration, weob-loney
virusp30.
Two to three half-cystine resi-tained
molecular weights of28,300
+ 600 forthedues
wereassigned
tobothpolypeptides
since in Moloney virus protein and 31,000 ± 300 fortheeach
casethe calculated value wasfound to be Y-1p30.
nonintegral.
On thebasis of the mass andvelocity
param-Sedimentation
analysis. Although p30
ex- eters, africtional ratio relative to a perfectly
hibits
amarked
proclivity
for
concentration-de-
spherical ellipsoid could
becalculated.
Aratio
ofpendent self-association (8), sedimentation
ve- 1.38 for Moloneyvirus p30 and 1.45 for Y-1
p30
locity and molecular weight analysis
canbe per-
corresponded to axial ratios of about 5 and
6,TABLE1.Amino acidcompositionof the major internal structural protein of the Y-1 andMoloney oncornavirus
Apparentresidues per mol ofprotein'
Aminoacid Y-1 virusprotein Moloneyvirus
protein(
Aminoacid_
~~~~~~~~~~~~~~~~~Observed
(Y-1)Avg or extrapo- Assumed compo- Avgorextrapo- Assumed compo- minusMoloney lated value sition lated value sition
Lysine... 16.7 17 17.3 17 0
Histidine.3.0 3 4.0 4 -1
Arginine.31.2 31 28.6 29 +2
Tryptophanc.3.8 4 3.0 3 +1
AsparticAcid.29.1 29 26.9 27 +2
Threonine .18.0 18 14.5 15 +3
Serine .17.0 17 15.8 16 +1
Glutamic Acid.51.2 51 44.5 45 +6
Proline.20.1 20 16.3 16 +4
Glycine.18.3 18 16.0 16 +2
Alanine .14.4 14 13.6 14 0
Half-cystined.2.6 3(2) 2.3 2(3) (0-1)
Valine .9.2 9 7.5 8 +1
Methionined.2.0 2 3.2 3 -1
Isoleucine.6.6 7 4.8 5 +2
Leucine.35.0 35 27.8 28 +7
Tyrosine... 6.3 6 5.3 5 +1
Phenylalanine 5.6 6 5.9 6 0
Calculatedmolwt 33,400 30,000
"Calculated with histidineastheintegral residue. Analyseswereperformedin
triplicate
onsamples dialyzed extensivelyagainstdistilledwater,lyophylized,andhydrolyzedat110°Cfor20,40, and 72 h, respectively, in 6 NHCIcontaining0.01%(vol/vol) phenoltopreservetyrosineresidues. Therecovery values ofthreonineand serinewereobtainedbyextrapolationof the recoveries from the varioushydrolysistimestozerotime.Valine, leucine, and isoleucine valuesweretakenastheirmaximum recoveriesfrom the timed HCIhydrolyses. Theremaining residuerecoveryvalueswereobtained fromanaverage of the timedHCIhydrolyses.
bDataforMoloneyvirus
p30
taken fromBurnette etal.(8).cTryptophananalyzed after hydrolysisat115°Cfor 22 hin4Nmethanesulfonic acid.
dHalf-cystine determinedascysteic acid and methionineasthesulfone byperformic acidoxidation
followed
by20hof HCIhydrolysis.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.509.73.458.332.569.2]VOL. 26, 1978 ONCORNAVIRUS p305&S INTERSTRAIN VARIATION 527
respectively,
assuming
ahydration value of
0.3ordered
secondary
structure asreported
previ-g
of
water per gof protein. These ratios indicate
ously
for
the
Moloney
virusp30
alone (8),
that,
although both
areapparently asymmetric
namely,
aresolution of the
patterns into
amin-ellipsoids, the Y-1 p30 is
a moreelongated glob-
imum number of
gaussian
curves toachieve
aular molecule.
best fit of the data and aleast-squares
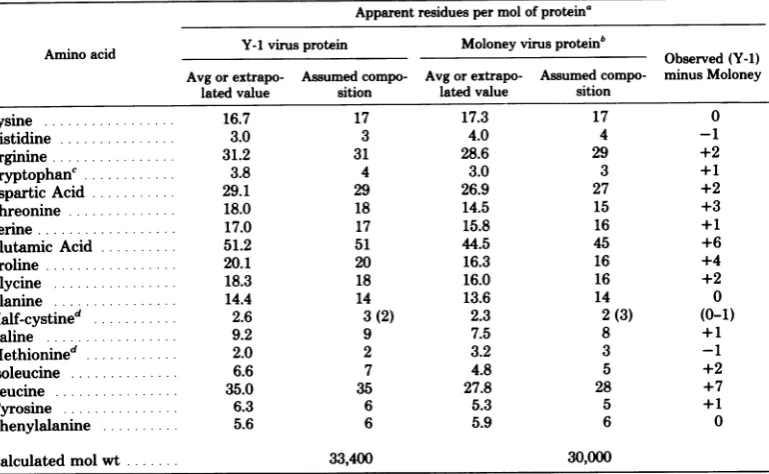
fit ofConformational
analysis.
Figure
8illus-
ellipticity
to alinear combination
ofthe
possible
tratesthe
major conformational differences be-
structural contributions.
Atabulation
ofthe
ro-tween
the
analogous
Moloney and Y-1 virus p30
tational
strengths
of the resolved
gaussian bands
proteins
asdetermined
by far-UV circular di-
by the former method is presented in Table
2,chroism. We have utilized
twoindependent
although both
procedures
gaveclosely
compa-methods for
estimating the
typeand
amountof
rable results. While both
polypeptides have
1
I
I
I
In6 - 600 ll l
I
0~~~~~~~
0.05 0.10 0.15 0.20 0.25 /
Ci(OPTICAL
DENSITY AT 280nm) E0
E E 10
B.
~~~~~~~~~~~~~~B
300 EU
0~~~~~~~~~~~~2
0.05 0.10
Ci
(OPTICAL DENSITYAT280nm) 200 210 220 230 240 250FIG. 7. Plotsof l/molecular weight versus initial WAVELENGTH (nm) loading concentration in analytical ultracentrifuge
cellfor the major internal structuralproteinof the FIG. 8. Far-UV circular dichroic spectra of the
Moloneyand Y-1 murine oncornaviruses.High-speed majorinternal structuralprotein oftheMoloneyand
sedimentation equilibrium wasperformed asprevi- Y-1 murine oncornaviruses. Circular dichroismwas
ously described (8), and the molecular weights at performed in 0.5-mmpath length fused-silica cells various initialprotein concentrations wereextrapo- from 250nm to200 nmin0.2Mborate(pH8.0) and latedto zero concentration by the method ofleast 0.15Mpotassiumfluoride. (A)Moloneyvirusprotein,
squares. (A) Moloney leukemia virusprotein; data 360 pg/ml. (B) Y-1 virusprotein, 350 ug/ml. [e]',
takenfromBurnetteetal. (8). (B) Y-1 virus protein. Reducedmeanresidueellipticity.
TABLE 2. Meanresidue rotational strengths of the far- UV circular dichroic bands of the major internal structuralprotein of Moloney leukemia virus and Y-1 virus
Virusprotein and
XOa
[OImaxb
Rc Transition Conformation %Conformation'Y-1
223 nm -10,500 -5.81 n-X* a-helix 28
215nm -3,500 -0.83 n-S7*
fl-structure
11Moloneyleukemia
224 nm -17,000 -10.88 n-7T* a-helix 52
216rum -3,000 -0.80 n-f* fl-structure 10
192nm +41,000 +17.25 IT-IT* a-helix 50
A., Wavelengthofcircular dichroictransition.
b
[6]max,
Maximummeanresidueellipticity.
R, Rotational strength in cgs units, x
10-,
of each resolved gaussian band determined as previouslydescribed(8).
dBasedon acomparison of the resolved gaussianband rotationalstrengths calculatedaspreviously described (8).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.509.65.236.188.391.2] [image:6.509.270.440.195.407.2] [image:6.509.57.451.514.604.2]528 BURNETTE AND MITCHELL J. VIROL.
about 10 to 14%
fl-structure,
Moloney
p30
ex-hindered,
andproline-adjacent arginyl
andlysyl
hibits
anexceptional
50 to 55%a-helicity
(8), bonds thatare nothydrolyzed by trypsin.
whereas the Y-1
p30 is predicted
to possess asignificantly
lower 27 to28% a-helix. The
re- DISCUSSIONmainder
ofthe
structure ispresumably
ape- Theprincipal physical
andchemical
charac-riodic.
teristics
ofMoloney leukemia
virusp30
and theTryptic
peptide
analysis.
Two-dimensional
endogenous
Y-1virus
p30
aresummarized
inpeptide
mapping
(Fig.
9) demonstrates that the
Table 3. Foranalogous proteins
from twostrains
Y-1and
Moloney
virus
p30 proteins share
two of murineoncornavirus possessing
group-spe-major
radioiodinated
tryptic
peptides.
However,
cificcross-reactivity, they display
asurprising
the
Y-1virus
p30
appearstopossessfive
unique
dissimilarity.
Although
they sharecommon
iso-major iodinated
peptides, and the
Moloney
leu-
electric points
andsedimentation
coefficients,
kemia virus
p30,
three.
Inview of
thisobserved
distinct
differences exist
inmolecular
weight,
sequence
heterogeneity,
it is notsurprising
that amino acidcomposition, tryptic peptides,
andthe two
proteins exhibit
significant type-specific
conformation.
Theconformational disparity
ob-antigenic variation
andsecondary
structural
dis- served bycircular dichroic analysis
is, incon-parity.
junction
with thepeptide maps,
the mostsignif-Assuming that
eachradioiodinated
tyrosine
icantinterstrain variation detected
as anexpla-residue is
located
on a separatepeptide,
the
nation of theobserved antigenic
heterogeneity.
number of labeled
major
peptides corresponds
Thehelical
differences
are most likely due torather well
tothe amino acid
analyses.
The
presence
of minor
peptides that
amount toless
than
10%
ofthe total
radioactivity
may be the
TABLE
3.
Summary of the
physical
and chemical
result of a number of
secondary
reactive proc- properties of the major internal structural protein ofesses,
such
asdiiodination of
tyrosine
and for-
Moloney leukemia virus and
Y-1
virus
mation
ofmonoiodohistidine
(22),
incomplete
S-
Majorstructuralproteinaminoethylation
of
cysteinyl
residues
(23, 24)
Property Moloneyleuke-and other chemical modifications
(27),
arginyl
miaS
virusav
and lysyl residues that are
conformationally
Extinction coefficient(E1cm,28)
13.7 9.5CAbsorption maximum
(nm) 280-281 279
Isoelectricpointd 6.2 6.3
Molecular
weighte
28,300
±60031,000
±300Partial
specific
volumeo G
~~~~~~~~~~~(ml/g)f
0.1...0.71 0.722,_. ,.m Sedimentation
coeffi-s.
_952
,,--'cient (so,w)
Z..;
2.47 2.47Diffusion coefficient
(D2w)
(cm2/s)....
7.33xi07 6.69x 10-v a-,
,-~~,-:_
- -~Frictional
ratio(f/fW)
1.38 1.45o
Amino acidresiduesh
259 290I'--',_a-Helicity (%).50 27
/3-Structure (%). 10-13 11-14
Major radioiodinated tryptic peptides
(shared/unique) 2/3 2/5
aDataonMoloneyvirus
p30
taken fromBurnette<|
~~~~~~~~~~~~~~~et
al.(8).ELECTROPHORESIS bDeterminedboth bydryweightmeasurementand
[image:7.509.268.458.317.545.2]synthetic boundary formation.
FIG. 9. Two-dimensional trypticpeptideanalysis cDeterminedbydryweightmeasurementonly.
ofthe Y- 1andMoloneyleukemiavirusp30proteins. d Difference estimatedon thebasis offocusing in Thepurifiedproteins wereiodinated,
S-aminoethyl-
polyacrylamidegels.ated,digestedwithtrypsin,andmappedonthin-layer eDeterminedbyhigh-speedsedimentation
equilib-sheetsasdescribed in the text.
Electrophoresis
was rium.in the horizontal plane and
chromatography
was fCalculatedfrom amino acidcomposition.
vertical (ascending). Symbols:
0,
sharedmajor
pep- g Calculated from sedimentationequilibrium
and tides;,, shared minorpeptides;0,
uniquemajor
velocity data.Moloney viruspeptides;
*0,
unique major Y-1 virus h Basedontimedhydrolysis
and aminoacidrecov-p30 peptides. ery data.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.509.67.255.366.560.2]VOL. 26, 1978 ONCORNAVIRUS p309ag INTERSTRAIN VARIATION 529
the
presenceof amino acid sequences and par-
kemia virus(W.
N.Burnette,
S.Duttagupta, and
ticular residues (e.g., proline) that
act ashelix-
R. Soeiro, manuscript
inpreparation).
Suchin-breakers
atcritical
regions
inthe
Y-1 virus p30.tragenic
exclusion could account for theapprox-These sequentially different
and/oradditional
imately
9% difference in molecularweight
seenresidues in the Y-1
polypeptide
henceinterfere between theN-tropic Y-1 virus p30 and thep30
with the formation of
a-helix
asobserved
inthe from theNB-tropic Moloney
virus used in ourMoloney
virusmolecule.
How theconforma-
studies. Intramolecular recombination
has beentional differences affect the
tertiarystructure ofpreviously
described inoncornaviruses
by Wyke
these
proteins is
notknown, although the hydro-
etal. (47)
fortransformation-dependent,
tem-dynamic studies indicate
someminorratiodif-
perature-sensitive
mutantsof the Rous sarcomaferences.
Steric hindrance
or otherfactors duevirus
genome.to
regional conformational
differences about the The sequenceheterogeneity
observed in theseimmunologically
cross-reactive sites
may con-investigations is also compatible with
pointmu-tribute
tothe
observed decrease
inantibody
tations.
Such
point mutations would give rise
toaffinity for the
Y-1virus
p30 molecule.
helix-forming amino acid residues and
sequencesOn
abiological level,
the
Moloney leukemia
and thus
tothe
increased
helicity of the Moloney
virus
and the
Y-1endogenous
virusalso
differ virus p30, since the 9% massexclusion
probablygreatly. The Moloney virus is highly oncogenic
cannotfully
accountfor the
22 to28%
difference
in vivo
and
displays high
titerinfectivity
within
helicity between the proteins from the
endog-NB-tropic host
range invitro
(C. Riggin,
W. N. enousand
exogenousviruses. Further, point
mu-Burnette, and W.
Mitchell,
unpublished
data). tationscould also be the
sourceof the
massSince the
laboratory strain
ofMoloney leukemia
difference
by leading
toaltered
processing and
virus arose
from
anendogenous murine
viruscleavage
of the
p30 from the
gag genepolypro-(24, 25) as a
result
ofthe
application
ofevolu-tein precursor. If altered processing
were thetionary
pressuresdesigned
toselect for enhanced
case, onemight
expect aconcomitant
massinfectivity
andoncogenicity,
it isreasonable
tochange
inthose
polypeptides (e.g., pl10a9) that
expect
these
pressures tobe
reflected
inboth
arecontiguous with the p30agin the polyprotein.
the
structureand
functional
natureof
its
gen-Alternate cleavage of the
precursorfor Rauscher
omic
products. The
natureof the variations that
leukemia virus
envelope glycoproteins is thought
give rise
tothe
immunochemical differences, for
tobe the
sourceof
the
twocomponentsof
theexample,
canbe ascribed
tothe chemical and
gp69/71efv complex (W. N. Burnette and J.
T.physical disparities noted in this study. Con-
August,Fed.
Proc.35:1736).
versely, the conservation
of the intrinsic charge Afunctional
approach
tothe effect of
evolu-and
general
shape between the p30 molecules
tionary
pressure onthe
major internal structural
argues
for
animportant functional
role, such
asprotein
componentof mammalian
oncornavi-molecular association
(8) and subviral
coreself-
ruseshas
notyetbeen
possible,
since
the
func-assembly.
tions of this
polypeptide
have
yet tobe
estab-Our data
suggestconsiderable
sequence het-lished.
However, investigations
toelucidate the
erogeneity
between the
major
structural
pro-functions of viral
p30,
particularly
inrelation
toteins of these
twoviruses, although
Oroszlan
etits
possible
role
invirion
self-assembly
and host
al.
(29) have described
only
asingle
amino
acid
rangerestriction,
arecurrently
in
progress.difference in the first
24N-terminal residues of
the
p30
proteins
from six different
murine on- ACKNOWLEDGMENTScomaviruses. Itispossiblethat theN-terminus Thisstudy was supported by PublicHealthServiceGrant
isthemosthighlyconservedportionofthemol- CA 14792 from the National Cancer Institute and bythe
ecule and
maybe the
portion responsible
for the
American Cancer Society.
species-specific
specirthees-specific immuhanologic
immunological cross-reactivity.
whicrss- heati
Theinvaluable assistance of LeslieHolladay
intheanalysisof thecircular dichroic datais
gratefully
acknowledged. The Nevertheless, the mechanism by which the het- iodinated peptideanalysis procedure was developed jointlybyerogeneityarises is notcurrently understood.On W.N.B. and MarkKrantz in theDepartment ofMolecular
astructurallevel, unequal crossingoverwithin Biology, Albert Einstein College ofMedicine, Bronx, New
the p30 gene during recombinational events York.Quantitative optical density scans of thetrypticpeptide
couldhaveledto the exclusionof genetic infor- mapswereprovided byW.Lutin,VanderbiltUniversity.
mation from the
endogenous virus
that is notLITERATURE
CITED
essential,
andperhaps inimical,
to exogenous 1. Arcement,L.
J., W.L.
Karshin, R. B. Naso, G.viral infectivity
andoncogenicity
functions.
This Jamjoon, and R. B.Arlinghaus.
1976.Biosynthesisisparticularly intriguing with respect to recent ofRauscher leukemia viralproteins:presence ofp30
observationsof massvariationinp30molecules and envelopep15 sequences in precursorpolypeptides.
inhostangerevrtants o Moloneyleukemia Virology69:763-774.
in host range revertants of Moloney leukemia 2. August, J. T.,D. P. Bolognesi, E. Fleissner, R.V.
virus
(19)
and hostrange clones of Friend leu-Gilden, and
R. C.Nowinski. 1974.
A proposedon November 10, 2019 by guest
http://jvi.asm.org/
530 BURNETTE AND MITCHELL J. VIROL
menclature for the virion proteins of oncogenic RNA Weir(ed.), Handbook of experimental immunology. F. viruses.Virology 60:595-601. A.DavisCo., Philadelphia.
3. Baltimore, D. 1970. RNA-dependent DNA polymerase 22. Lamoureux,G., P. R. Carnegie, and T. A. McPherson. in virionsof RNA tumour viruses. Nature (London) 1967. Experimental allergic encephalomyelitis-226:1209-1211. propertiesofaniodinatedencephalitogenicpolypeptide. 4. Baltimore, D. 1974. Tumor viruses. Cold Spring Harbor Immunochemistry4:273-281.
Symp.Quant.Biol. 39:1187-1200. 23. Lindley,H. 1956. Anewsyntheticsubstrate fortrypsin 5.Barbacid, M., J. R. Stephenson, and S. A. Aaronson. and itsapplicationto thedetermination of the amino-1976. gag Geneofmammalian type-C RNA tumour acid sequence of proteins. Nature (London) viruses. Nature(London)262:554-559. 178:647-648.
6. Bilello, J. A., M. Strand, and J. T. August. 1977. 24. Moloney, J. B. 1960. Biological studies on a lymphoid Expression of viralenvelope glycoproteinand transfor- leukemia virus extracted from sarcomaS37.I.Origin mationgenes in cells transformedby a defective kirsten and introductory investigations. J. Natl. Cancer Inst. murine sarcoma virus. Virology77:233-244. 24:933-951.
7. Bray,D., and S. M. Brownlee. 1973.Peptidemapping 25. Moloney, J. B. 1966. A virus-inducedrhabdomyosarcoma of proteins from acrylamide gels. Anal. Biochem. ofmice.Natl.Cancer Int. Monogr.22:139-142. 55:213-221. 26. Nowinski, R. C., E.Fleissner,N. H.Sarkar, and T. 8. Burnette, W.N.,L.A.Holladay,andW. M. Mitchell. Aoki. 1972. Chromatographic separationandantigenic 1976.Physicaland chemicalpropertiesofMoloneymu- analysis of proteins of theoncornaviruses. II. Mamma-rineleukemia virusp30protein:amajorcorestructural lianleukemia-sarcomaviruses. J.Virol.9:359-366. componentexhibitinghighhelicity and self-association. 27. Offord, R. E.1966.Electrophoreticmobilitiesofpeptides J.Mol. Biol. 107:131-143. onpaper and their use in thedetermination of amide 9. Burnette,W. N., C. H.Riggin, and W. M. Mitchell. groups. Nature (London)211:591-593.
1974.Physicalandchemicalpropertiesofanoncorna- 28. Oroszlan, S., D. Bova, R. J. Huebner, and R. V. virusassociatedwithamurine adrenal carcinoma cell Gilden. 1972. Major group-specific protein of rat type line.J. Virol. 14:110-115. Cviruses. J.Virol. 10:746-750.
10. Chen, K. C. S., T. J. Kindt, and R. M. Krause. 1975. 29. Oroszlan, S., T. Copeland, M. R. Summers, G. Smy-Primary structure of the L chain fromarabbit homo- thers, and R. V. Gilden. 1975.Amino acid sequence geneousantibodytostreptococcal carbohydrate. I. Pu- homology of mammalian type C RNA virus major in-rification ofantibodyandsequence determination of temalproteins. J. Biol. Chem.250:6232-6239. peptidesfroma-cnymotrypticandthermolyticdigests. 30. Oroszlan,S., C. L. Fisher, T. B. Stanley, and R. V. J.Biol. Chem. 250:3280-3288. Gilden. 1970. Proteins of the murine C-type RNA 11. Cole, R. D. 1967.S-aminoethylation.MethodsEnzymol. tumorviruses: isolation of a group-specific antigen by
11:315-317. isoelectric focusing. J. Gen. Virol. 8:1-10.
12. Davis,J.,M.Scherer, W. P.Tsai,and C.Long. 1976. 31. Oroszlan,S., C. Foreman, G. Kelloff, and R. V. Gil-Low-molecular-weightRauscher leukemia virusprotein den. 1971. Thegroup-specific antigen and other struc-withpreferential bindingforsingle-strandedRNA and tural proteins of hamster and mouse C-type viruses. DNA.J. Virol. 18:709-718. Virology 43:665-674.
13. Geering, G.,T.Aoki,andL. J. Old.1970.Shared viral 32. Oroszlan,S., R. J. Huebner, and R. V. Gilden. 1971. antigen ofmammalian leukemia viruses. Nature(Lon- Species-specific and interspecific antigenic determi-don)22:265-266. nants associated with thestructural protein of feline C-14. Geering,G.,W. D.Hardy, Jr., L.J.Old,E. deHarven, type virus. Proc.Natl. Acad. Sci. U.S.A. 68:901-904.
andR. S.Brodey.1968.Sharedgroup-specificantigen 33. Oroszlan, S., M. R. Summers, C.Foreman, and R. V. of murine and feline leukemia viruses. Virology Gilden. 1974. Murine type-C virus group-specific anti-36:678-707. gens: interstrain, immunochemical, biophysical, and 15.Geering,G.,L. J.Old,and E. A.Boyse.1966.Antigens amino acidsequence differences. J. Virol.14:1559-1574. of leukemiasinduced by naturally occurringmurine 34. Raftery, M. A., and R. D. Cole. 1963. Tryptic cleavage leukemia viruses: their relationtotheantigens of Gross at cysteinyl peptide bonds. Biochem. Biophys. Res. virus and other murine viruses. J. Exp. Med. Commun. 10:467472.
124:753-772. 35. Sarma, P.S., and T. Log. 1971. Viral interference in 16. Gibson,W. 1974.Polyomavirusproteins:adescription felineleukemia-sarcoma complex. Virology 44:352-358. ofthestructuralproteinsof the virion basedonpoly- 36. Schafer,W., F. A. Anderer, R.Bauer, andL.Pister. acrylamide gel electrophoresis and peptide analysis. 1969.Studies on mouse leukemia viruses. I.Isolation Virology 62:319-336. and characterization of agroup-specific antigen. Virol-17.Gomard,E., J. C.Leclerc,and J. P.Levy.1972.Murine ogy 38:387-394.
leukemia andsarcomaviruses: furtherstudiesonthe 37. Sen, A., C. J. Sherr, and G. J. Todero. 1976. Specific antigens of the viral envelope. J. Natl. Cancer Inst. bindingof thetype C viral coreproteinp12 with purified
50:955-961. viral RNA.Cell7:21-32.
18. Gregoriades, A.,and L. J. Old.1969.Isolationandsome 38. Shapiro,S.Z.,M.Strand, and J. T.August.1976. High characteristicsof agroup-specific antigenofthe murine molecularweightprecursorpolypeptidestostructural leukemia viruses.Virology37:189-202. proteinstoRauscher murineleukemia virus.J. Mol 19.Hopkins, N., J. Schindler, and R. Hynes. 1977.Six Biol.107:459-477.
NB-tropic murine leukemiavirusesderivedfrom a B- 39. Stephenson,J.R., S. R.Tronick, and S.A.Aaronson. tropic virus ofBALB/c have altered p30. J. Virol. 1974.Analysisoftypespecific antigenicdeterminants 21:309-318. of twostructuralpolypeptidesof mouseC-typeviruses. 20. Hunsmann,G.,V.Moennig,L.Pister,E.Seifert,and Virology58:1-8.
W. Schafer. 1974. Properties ofmouseleukemia vi- 40. Stephenson,J.R.,S.R.Tronick,and S. A.Aaronson. ruses. VIII. ThemajorglycoproteinofFriend leukemia 1975.Murineleukemia virusmutantswith temperature-virus.Seroimmunological, interferingandhemagglutin- sensitive defects inprecursorpolypeptide cleavage. Cell natingcapacities.Virology62:307-318. 6:543-548.
21. Hunter,W. M. 1967. Thepreparationof radioiodinated 41.Strand, M.,and J.T.August.1973.Structuralproteins
proteinsofhighactivity,theirreactionwithantibody ofoncogenicribonucleic acid viruses.InterspeII,anew
in vitro:theradioimmunoassay, p.608-642.In D.M. interspeciesantigen.J. Biol.Chem.248:5627-5633.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26,1978 ONCORNAVIRUS p309a9 INTERSTRAIN VARIATION 531
42. Strand, M., and J. T. August. 1974.Structuralproteins 14:1575-1583.
ofmammalian oncogenic RNA viruses: multiple anti- 45. Temin, H. M. 1971.Theprotovirus hypothesis: specula-genic determinants of the major internal protein and tions onthe significance ofRNA-directedDNA synthe-envelopeglycoprotein.J.Virol. 13:171-180. sis fornormaldevelopment and forcarcinogenesis. J. 43.Strand, M., and J. T.August. 1976. Structural proteins Natl. Cancer Inst. 46:iii-vii.
ofribonucleicacid tumor viruses.Purification of enve- 46. Temin, H., and S. Mizutani. 1970. RNA-dependent lope, core, and internal components. J. Biol. Chem. DNA polymerase in virions of Rous sarcoma virus.
251:559-564. Nature(London) 226:1211-1213.
44.Strand, M., R. Wilsnack, and J. T. August. 1974. 47. Wyke, J.A.,J. G. Bell,and J. A. Beamand. 1976. Structural proteins of mammalian oncogenic RNA vi- Genetic recombination among temperature-sensitive ruses:immunological characterization of the p15 poly- mutants of Roussarcoma virus. Cold Spring Harbor peptide of Rauscher murine virus. J. Virol. Symp. Quant. Biol. 39:897-905.