Copyright01974 American Society for Microbiology Printed inU.S.A.
Hepatitis B Core Antigen:
Immunology
and
Electron
Microscopy
LEWELLYSF.BARKER, JUNE D.ALMEIDA,JAY H. HOOFNAGLE,ROBERT J. GERETY,DANIEL R. JACKSON, AND PHILIP P. McGRATH
Divisionof Blood andBloodProducts, BureauofBiologics,Food andDrugAdministration, Bethesda,
Maryland,
and the WellcomeResearchLaboratories,Beckenham,
Kent,England
Received forpublication 20August 1974
Two distinct viral antigens are associated with the hepatitis B virus: the
hepatitis B surface antigen (HB8Ag, Australia antigen) and the hepatitis B core
antigen
(HB,Ag). HB.Ag,
purified from the serum of asymptomatic humanHB8Ag carriers, and
HBcAg,
purified from the liver of a chimpanzee acutelyinfected with hepatitis B virus, were examined by serological and immune electron microscopic methods. Antisera raised against HB8Ag reacted with the outer, surfacecomponentof the Daneparticleandwith the 20-nmspherical and tubular particles present in HB8Ag-positive serum, but not with the internal componentof theDaneparticleorwithpurified
HBcAg
particles.Antiseraraisedagainst purified
HB,Ag
particles reacted with the internal component of the Dane particle and withHBcAg,
butnotwiththesurfaceof the Daneparticleorwith the 20-nmspherical and tubular particlesassociatedwith
HB"Ag.
PurifiedHBcAg
particles, 27 nm in diameter, demonstrated distinct subunits. Theinfectious form of hepatitis B virus appears to be represented by the 42-nmDaneparticle composedofa27-nmnucleocapsidcorecomponent
(HBcAg)
surrounded byanantigenically andmorphologically distinct lipoprotein surface
component
(HB,Ag).
Evidence
fromseveral laboratories
hasdem-onstrated that the
hepatitis
B virus(HBV)
possesses at least two separate
and distinct
antigens: the
hepatitis
B surfaceantigen
(HB.Ag,
Australia antigen) and the
hepatitis
B coreantigen
(HB,Ag)
(1, 3, 6,
10).
HB8Ag,
the
lipoprotein
surface component ofHBV,
canbe
readily detected
by
a variety ofimmunological
techniques
inthe
sera ofpatients
acutely
orchronically
infected with this virus
(4, 17).
HB,Ag,
the
nucleocapsid
component ofHBV,
is notfound
free in serumbut
isfound
in the nuclei ofhepatocytes
frompatients with type Bhepatitis
and canalso be extracted
fromHB8Ag-reactive
seracontaining Dane particlesby detergent
treatment (1, 11-13, 16, 18; R. H.Purcell,
J. L.Gerin,
J. D.Almeida,
and P. V.Holland,
Intervirology, in press). Both antigens are associated with distinctvirus-like structures visible on electronmicroscopy (1, 2, 11, 12, 18).HB.Ag
inserum is associatedwith 20-nmspher-ical
particles,
elongated
tubularparticles
(20-nm inwidth and several hundred nanometers inlength),
and with the outer, surface component of thecomplex,
42-nm Dane particles (1, 7).HBcAg
is associated with the inner, 27-nm,electron-dense
core component of the Daneparticle
seen in HBsAg-positive serum and withthe
27-nm nucleocapsid found in the nuclei ofhepatocytes
of patientsinfected with HBV (1-3, 11, 12, 18).This
reportdescribes
the purification andcharacterization
ofnucleocapsid core particles fromthe
liver of an experimentally infectedchimpanzee
(3; J. A.Markenson,
R. J. Gerety,J. H. Hoofnagle, and
L. F. Barker, J. Infect.Dis.,
in press), and the demonstration by im-muneelectron
microscopy that these particles areantigenically
identical to the inner, core component(HB,Ag)
but
antigenically distinct fromthe
outer, surface component(HB8Ag)
of the Daneparticle.MATERIALS AND METHODS
Serological testing. HB8Agwasdetectedby coun-terelectrophoresis (9) and by solid-phase radioim-munoassay (Ausria; Abbott Laboratories, North Chi-cago,Ill.) using theroomtemperature procedure (15). AntibodytoHB8Ag
(anti-HB.)
was assayed by coun-terelectrophoresis, passive hemagglutination (19), and solid-phase radioimmunoassay (Ausab; Abbott Laboratories [Reagents kindly provided by Abbott Laboratories.]). Selectedsampleswerealsotested foranti-HB.
by radioimmunoprecipitation (14). HBcAg andantibodytoHB,Ag
(anti-HB,)
weredetectedbya 1552on November 10, 2019 by guest
http://jvi.asm.org/
further processing. Electron microscopyof liver tions revealed numerous 27-nm intranuclear core
particles. Because of this, attempts were made to purify core particles from this liver by differential centrifugation. Liver (80g) wassuspendedin320ml of hypotonic (0.45%) saline and homogenized in a Waring blender. The 20% (wt/vol) homogenate was clarified by centrifugationat2,500 rpm for30min in an International PR-2 centrifuge (1,100 x g). The supernatant fluidwascentrifugedat25,000 rpmfor 2 hina no.30 rotorina Beckman L-2ultracentrifuge (75,000 x g). Theresultingpelletwasresuspendedin 35 ml ofdistilled water, and thehigh-speed centrifu-gationwasrepeated. Thepelletwasthenresuspended
in 35 ml of distilled water and clarified again by centrifugation at 2,500 rpm for30min(1,100 x g).
Theresultingsupernatantwasusedastheantigen
in serological testing for both complement fixation and counterelectrophoresis to detect
anti-HB,.
To furtherpurify thissupernatant, itwaslayeredonto a continuousCsClgradient,density1.2to1.5g/ml.The gradient mixturewascentrifugedat22,000 rpm for16 h in an SW25.2rotor in a Beckman L-2 ultracentri-fuge (75,000 x g). Fractionswerecollecteddropwise from the bottom ofthetube in approximately 1-ml amounts. Thespecific gravity ofthe resultant frac-tionswas determined byusing anAmericanOptical refractometer. Each fractionwas assayedfor HB.Ag byradioimmunoassayandforHBcAg
by complement fixation and examined forvirus-like particles under the electronmicroscope. Fractionscontainingtypical27-nm core particles were dialyzed for 1 to 3 days againstphosphate-buffered saline and usedto immu-nize animals and for immune electron microscopic studies.
HB.Ag
was purified from the plasma ofhumanchronic
HB.Ag
carriersbyaseries ofisopycnic band-ings in CsCl and rate zonal ultracentrifugations in sucrose (5). Samples weredialyzed
againstphos-phate-buffered salinepriorto use.
Immuneelectronmicroscopy.
Specimens
for elec-tron microscopy were prepared by dialyzing either purifiedHB.Ag
orHBcAg
againstphosphate-buffered saline. Specimens were then diluted with an equal quantity of phosphate-buffered saline, and 0.5-ml portions ofeither antigen(HB.Ag
orHB,Ag)
weremixed with 0.02 ml of the antisera to be tested
(anti-HB.,
anti-HB,,
or normal control serum). Forstudies of the Dane particles, purified
HB.Ag
was treated with Tween 80priortomixingwith antisera (1). Antisera were tested under code, identified by species only. The antigen-antibody mixtures wereallowedtoreactovernightat4C and thencentrifuged for1h at18,000 xg.Thesupernatantwas
discarded,
HB,Ag
(anti-HBc, anti-core) were prepared in guineapigs (10; R. J. Gerety, J. H. Hoofnagle, and L. F. Barker, J. Immunol., in press). Hartley guinea pigs weighing 200 to 400 g were inoculated subcutaneously with 0.2 ml of HB8Ag or
HB,Ag
(diluted 1:3 to 1:10in normal saline), emulsified in an equal volume of complete Freund adjuvant, and were boosted with a similar amount of antigen emulsified in incomplete Freund adjuvant at 14 days. The animals were bled by cardiac puncture at 21 days and at weekly intervals thereafter. In addition, antisera to both HB.Agand HBcAgwere prepared in rabbits andrhesusmonkeys using similar immunization schedules. Naturally oc-curring antibodies toHB.Ag
and HBcAg were ob-tained from chimpanzees experimentally infected with HBV and from humans at varying stages of convalescence from type B hepatitis.Anti-HB.
was obtained from achimpanzee (chimp 959) who devel-oped this antibody 5 weeks after exposure to infec-tious serum and inwhomHB.Ag
was neverdetected (Markenson et al., J. Infect. Dis., in press).Chimpan-zee
anti-HB,
was obtained from three animals(chimps 23, 920, and 921) all in the immediate convalescent period of typical, experimentally in-duced type B hepatitis (after the disappearance of
HB.Ag
and prior to the appearance ofanti-HB.).
Human
anti-HB.
was obtained from a hemophiliac patient, and human anti-HBc wasobtained from a patient recently convalescent from type B hepatitis (1). Control sera consisted of prebleeds from uninocu-lated guinea pigs, rabbits, rhesus monkeys, and one chimpanzee (chimp 23). Human control serum con-sisted of plasma negative forHB.Ag,anti-HB.,
and anti-HBc from a person with no known history or exposure to type Bhepatitis.RESULTS
Electron
microscopic
examination of intact liver sections from thechimpanzee
revealed many intranuclear core particles. Both the crude homogenate and pellet fromthe
secondhigh-speed
ultracentrifugation
revealed 27-nmHBcAg
particles and typicalHB.Ag
particles(20-nm spheres and
tubules)
(Fig. 1).Examina-tion of fractions from CsCl gradients revealed typical 27-nm
nucleocapsid
core particles in fractions at adensity
from1.30 to1.33g/ml
and the typicalHB.Ag
particles in fractions at adensity
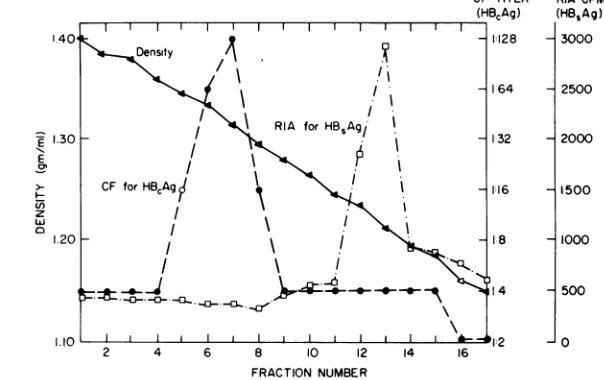
from 1.20 to 1.23 g/ml.Figure
2 illus-trates the patterns ofreactivity
ofHB.Ag
(byradioimmunoassay)
andHB,Ag
(by comple-ment fixation) found in each fraction from aon November 10, 2019 by guest
http://jvi.asm.org/
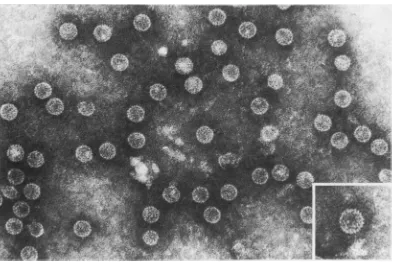
--FIG. 1. Pellet of second high-speed ultracentrifugation ofthe
chimpanzee
liverhomogenate.
BothHB,Ag
particles (20-nm spheres andtubules) andHBAgparticles (27-nm
cores)
are seen. x140,000.2 4 6 8 10 12 14 16
FRACTION NUMBER
FIG. 2. Patternof serological reactivity inCsCIgradientfractions ofthe materialshown inFig.1.HB,Ag,as
detectedbyradioimmunoassay (0),was seenatdensities(A) of1.20 to1.23g/ml,whereasHBeAg,asdetected
by complement fixation (0), was seenat densitiesof1.30to1.33g/ml.ElectronmicroscopyrevealedHB8Ag particles infractions12and 13andHBAg particlesinfractions 6 and 7ofthisgradient.
CsCl gradient.
HB,Ag
reactivity was seen in fractionswithdensities of from1.30to1.33g/ml andHB8Ag reactivity at densities of from 1.20 to 1.23g/ml. Thispattern wasfound repeatedly onsubsequent isopycnic bandings in CsCl.Because of the high degree of purity ofthe
HBcAg
particles, itwaspossibletoobtain bettermorphological characterization than has been
possible with previous serum-derived
prepara-tions. The great majority of particles were 27 nm in diameter. From the viewpoint of
sub-structure, the core particles displayed distinct
subunits (Fig. 3). Although subunits could be resolved, it was not possible to determine the exactnumber ofcapsomeresforming the capsid.
Insomeareas,particles showed
breakdown,
and1554
E 1.30
E
z
0
1.20.
RIACPM
(HBSAg)
3000
2500
2000
1500
1000
500
0
O
on November 10, 2019 by guest
http://jvi.asm.org/
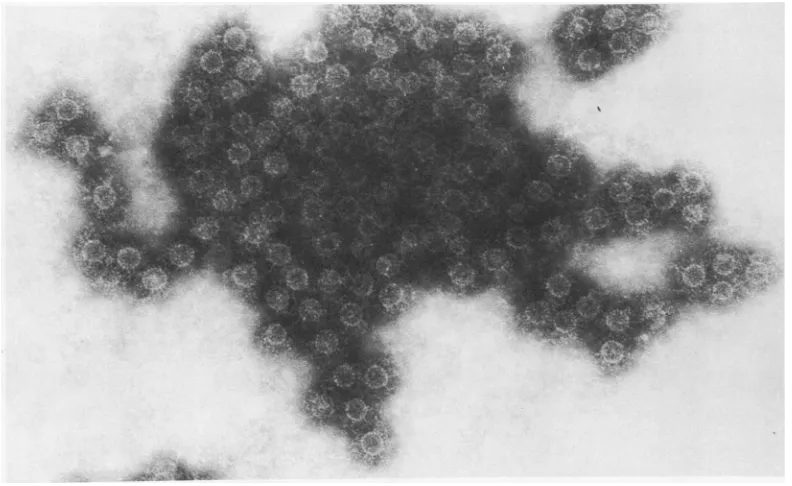
[image:3.499.117.397.69.287.2] [image:3.499.96.398.329.519.2]FIG. 3. PurifiedHBAg particlesseen at adensityof1.32 inaCsCIgradientshown inFig.2. Thepurity ofthe preparation and morphologyofthecoreparticles is shown. The average diameter
of
theparticles
was27nm, withoccasionalparticles only20 nmin diameter. x230,000. The insert showsasinglecoreparticleat ahigher
magnification (x350,000). This demonstrates the distinctive characteristicoftheHB¢Ag particle, the subunits whichcomprisetheparticle.Itwas notpossibleto establish theexactnumber
of
subunitspresent.in
these
itappeared
thatpartially disrupted
core
particles coalesced
toformhighly aberrant
forms
(Fig.
4).Results
ofserological testing
andimmune
electron microscopy on antisera from
guinea
pigs,
rabbits, rhesus monkeys, chimpanzees,
and humans
aresummarized
inTable
1.Immu-nization of guinea pigs with
HB,Ag
yielded
antisera
that contained
anti-HB,
by
comple-ment fixation
and
counterelectrophoresis
inthe
absence
ofdetectable
anti-HB8
by passive
he-magglutination, radioimmunoassay, and
radi-oimmunoprecipitation. These antisera failed to
agglutinate
HB.Ag
particles
or attach to the surface component ofthe
Dane particle(HB.Ag),
but distinct
antibody attachment wasseen to
the
internal core component ofthe
Daneparticle previously released by
Tween 80 treat-ment ofthe
serum (1). Antisera raised in guineapigs against purified
HB8Ag
gave the opposite pattern ofreactivities. Although
a high titer ofanti-HB8
was found in these antisera, noanti-HB,
wasdetectable
by complement
fixation orcounterelectrophoresis. Furthermore, when
guinea pig
anti-HB8
wasreacted with purified,
disrupted
Daneparticles,
alarge
amount ofantibody
was seenattached
tothe outersurface component of the Daneparticle
aswellas tothe
20-nm
spherical
andtubular
particles,
butnone was seenattached
tothe
inner, core component ofthe Daneparticle (Fig.
5).Immune
electron
microscopy using
purified
core
particles
andpurified HBSAg
particles
confirmed these
findings.
Guinea
pig,
rabbit,
chimpanzee, and human
serapositive
foranti-HB,
by serological
testsdemonstrated both
agglutination
andantibody
attachment topuri-fied core
particles (Fig. 6). However, when these
anti-HB,
reactiveserawerereacted
withHB.Ag
purified
from serum, noagglutination
oranti-body attachment
was seen.Conversely,
guinea
pig,
chimpanzee, and human
anti-HB.
did
not reactwith thepurified
HB,Ag
preparations,
but
demonstrated
agglutination
and
antibody
at-tachment
toHBSAg
particles (Fig. 5). Control
sera
(prebleeds and human
serumnegative
forantibodies
toHBVantigens) showed
no reactiv-ity with eitherHB8Ag
orHBcAg
particles.
DISCUSSION
The
immune electron
microscopic and
sero-logical data presented
here reinforcesthe
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.499.48.443.72.334.2]FIG. 4. Malformed HB^Agparticles. Occasionallygroupsof malformedandseemingly coalesced
particles
were seen. Theseparticles mayhave been altered anddisruptedbythepurification procedure. Alternatively, such particlesmay represent incompletely formedvirus. x238,000.
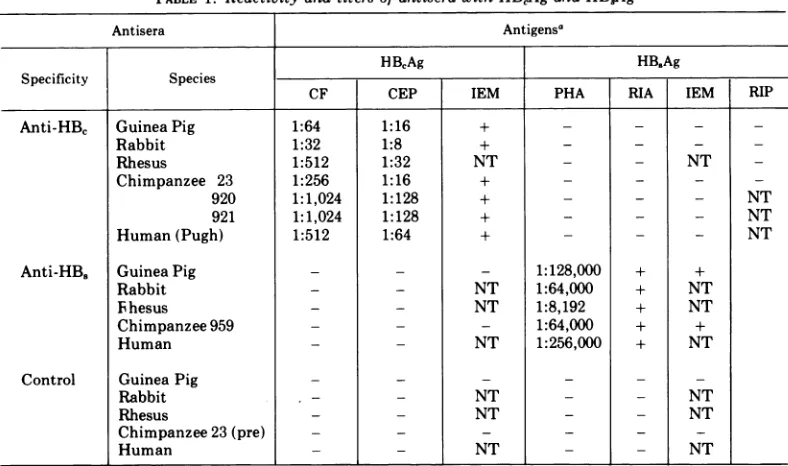
TABLE 1. Reactivityand titersofantiserawithHBAgand
HB8Ag
Antisera Antigensa
HBCAg HB,Ag
Specificity Species
CF CEP IEM PHA RIA IEM RIP
Anti-HBc GuineaPig 1:64 1:16 + - - _
Rabbit 1:32 1:8 + - - _
Rhesus 1:512 1:32 NT - - NT
Chimpanzee 23 1:256 1:16 + - -
-920 1:1,024 1:128 + - - - NT
921 1:1,024 1:128 + - - - NT
Human(Pugh) 1:512 1:64 + - - - NT
Anti-HB.
GuineaPig - - - 1:128,000 + +Rabbit - - NT 1:64,000 + NT
Ihesus - - NT 1:8,192 + NT
Chimpanzee959 - - _ 1:64,000 + +
Human - - NT 1:256,000 + NT
Control GuineaPig _- -
-Rabbit - - NT _ - NT
Rhesus - - NT _ _ NT
Chimpanzee23(pre) - - _ _ _
Human - - NT NT
aAbbreviations:CF,complementfixation;CEP,counterelectrophoresis; IEM,immuneelectronmicroscopy; PHA, passivehemagglutination; RIA,radioimmunoassay; RIP, radioimmunoprecipitation; NT,nottested.
mulating
evidenceregarding
the existence oftwo
independent
antigen-antibody
systemsas-sociated
with type Bhepatitis
(1, 3, 6, 10,
12).
HBcAg
particles
have beenpurified
from the liver of achimpanzee
that diedduring
acute viralhepatitis,
type B. Immune electronmi-croscopy using this purified
HBcAg
and deter-gent-treated, Dane-richHB.Ag-positive
prepa-rationshave
revealed the antigenic identity of the 27-nm intranuclear core particle with the internal component of the Dane particle. Immu-nization of a variety of laboratory animals with 1556on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.499.61.455.337.570.2]4sikS: * : S ; Aj;w;
FIG. 5. Reactionseenwhenguinea pig antiserum against HBAg
(anti-HB,)
was mixed with a preparation of Dane particles disrupted by detergent treatment. The antibody agglutinated and attached to the surface component of the Dane particle and to the 20-nm spherical and tubular particles. Antibody was not seen attachedtothe intemal component of the disrupted Dane particles, and when this antiserum was mixed with purified HBAg no agglutination was seen, but the particles appeared dispersed and uncoated as shown in Fig. 3. x230,000.FIG. 6. Reaction seen when chimpanzee antiserum against HBAg
(anti-HBc)
was mixed withpurified HB,Agparticles. Thepreparation shown inFig. 3wasreacted withchimpanzeeanti-HB8naturallyacquired
fromanHBVinfection. The particlesareagglutinated andareobscuredbyahaloofattachedantibody.Similar resultswereobtainedusing antiserum from animals immunized withHBAg. Thesameantisera hadnoeffect
on
HB,Ag particles.
x158,000.
1557
[image:6.499.47.443.378.621.2]BARKERET AL.
this
purified
HB,Ag
preparation
yielded
anti-sera
which
hadhigh
titers ofanti-HBc,
butwhich
were negative foranti-HB.
bythe
mostsensitive
techniques (14). Also
demonstrated
wastheability
ofpurified
HB.Ag
toinduce
high titersof
anti-HB.
in the absence ofanti-HB,
detectable by complement
fixation,coun-terelectrophoresis,
or immuneelectron
micros-copy.Finally,
theantigenic duality shown
byserological techniques
wasconfirmed visually
by immune electron microscopy using
both
purified
HBcAg
andpurified, disrupted
serum-derived
Daneparticles.
The
purity
ofthe
corepreparation
described
here has
allowed
for a bettermorphological
description
of the coreparticle.
Distinctsub-units were
visualized, although
theexactnum-ber of capsomeres
forming
thenucleocapsid
couldnot
be
determined.
The presenceofsub-units
differentiates
this agent fromen-teroviruses and
rhinoviruses
which alsofall
in the 27-nmrangeindiameter.
Indeed,
the struc-ture, size, andcharacteristics
ofHBV
appeartoplace it in aviruscategoryof itsown.Failure of anti-HB8 and
anti-HBc
to react with there-centlydiscovered
hepatitis
Aantigen instools of patients acutely ill with viralhepatitis,
typeA, makes it unlikely that the hepatitis A and B viruses areimmunologically
related (8). The recentdemonstration
of a specific DNA-dependent, DNA polymeraseintheheavycorefractions from
detergent-disrupted
Dane parti-cles isstrongevidencethat HBV isaDNA virus(13).
Immunological
and electron microscope datanow allows the description ofthe
physical
andantigenic madeupofthehepatitisBvirus. The 42-nm Dane
particle
is most likely the intact, infectious form of the virus andismadeupofa27-nm
nucleocapsid
core(HBcAg)
surroundedby a
lipoprotein
surface component(HB.Ag).
The morenumerous20-nm
spherical
and tubu-lar structures seen inHB8Ag-positive
serumprobably
representthelipoprotein
surfaceanti-gen
(HB.Ag)
produced
by infected hepatocytesfar inexcessof what isnecessarytoencapsulate the number ofnucleocapsid forms. The exact role of
HBcAg
andanti-HBcinthecourseoftype Bhepatitis
isnotknown.Hopefully,
thediscov-eryand
characterization
ofthissecondantigen-antibodysystemin typeBhepatitis will
permit
aclearerunderstanding of this still uncontrolledandsometimes fatal viral disease.
ACKNOWLEDGMENTS
Wethank Audrey J. Shawver and DavidE. Gilbertfor technical assistance and Robert Purcellfortestingselected
samples byradioimmunoprecipitation. LITERATURE CITED
1. Almeida, J. D.,D.Rubenstein,and E. J. Stott. 1971. New antigen-antibodysysteminAustralia-antigen-positive hepatitis. Lancet 2:1225-1227.
2. Almeida, J. D.,A. P. Waterson, J.M.Trowell, and G. Neale. 1970. The findingofvirus-likeparticlesintwo
Australia-antigen positive human livers. Microbios 2:145-153.
3. Barker, L. F., F. V. Chisari, P. P. McGrath, D. W. Dalgard, R. L. Kirschstein, J. D. Almeida, T. S. Edgington, D. G. Sharp, and M. R. Peterson. 1973. Transmission ofviral hepatitis, typeB, to
chimpan-zees.J.Infect. Dis.127:648-662.
4. Blumberg,B.S., A.I.Sutnick, and W. T. London. 1968.
Hepatitis and leukemia. Their relation to Australia antigen. Bull.N.Y.Acad.Med.44:1566-1586. 5. Bond, H. E., and W. T. Hall. 1972. Separation and
purification ofHepatitis-associated antigen into
mor-phologic typesby zonalultracentrifugation.J. Infect. Dis. 125:263-268.
6. Brzosko,W.J.,K. Madalinski,K.Krawczynski, and A. Nowoslawski. 1973. Dualityofhepatitis B antigen and itsantibody.I.Immunofluorescence studies.J. Infect. Dis.127:424-428.
7. Dane, D. S., C. H. Cameron, and M. Briggs. 1970. Virus-likeparticlesinserumofpatients with Australia-antigenassociated hepatitis. Lancet 1:695-698. 8. Feinstone, S. M., A. Z. Kapikian, and R. H. Purcell.
1973. Hepatitis A: detection byimmuneelectron
mi-croscopyofavirus-like antigen associated withacute
illness.Science 182:1026-1028.
9. Gocke, D. J., and C. Howe. 1970. Rapid detection of Australia antigen bycounterimmunoelectrophoresis.J. Immunol. 104:1031-1034.
10. Hoofnagle, J. H., R. J. Gerety, and L. F. Barker. 1973. Antibody to hepatitis-B-virus core in man. Lancet 2:869-873.
11. Huang,S.N. 1972.Hepatitis-associatedantigen hepati-tis. An electronmicroscopic study of virus-like particles inlivercells. Amer. J. Pathol. 64:453-470.
12. Huang, S.N., andV.Groh.1973. Astudyonantibodies
produced with liver tissue containing Australiaantigen andvirus-like particles. Lab.Invest.29:743-750. 13. Kaplan, P. M., R. L. Greenman, J. L. Gerin, R. H.
Purcell, andW.S. Robinson. 1973. DNA polymerase associatedwith human hepatitis B antigen. J. Virol. 12:995-1005.
14. Lander, J. J., H. J. Alter, and R. H. Purcell. 1971. Frequency of antibodytohepatitis-associatedantigen
asmeasured bya newradioimmunoassay technique. J. Immunol. 106:1166-1171.
15. Ling, C. M., and L. R. Overby. 1972. Prevalence of hepatitisBvirusantigenasrevealed by direct radioim-mune assay with 1125 antibody. J. Immunol.
109:834-841.
16. Lipman,M.B., W. J.Hierholzer,Jr.,and A. Schlueder-berg. 1973. Isolation ofcoresfrom hepatitis B Dane particles.J. Infect.Dis. 128:664-667.
17. Prince, A. M. 1968. An antigen detected in the blood duringtheincubationperiodofserumhepatitis.Proc. Nat.Acad.Sci.U.S.A. 60:814-821.
18. Stannard, L. M.,A.Polson,A.Kipps, andJ.R.Parker. 1973. Twoantigen-antibodysystemsinserum
hepati-tis:preparationofimmunoglobulins andtheir reaction with Australia antigen and particles in liver cell homogenates.Microbios 8:87-100.
19. Vyas,G.N.,and N. R.Shulman.1970.Hemagglutination
assay for antigen and antibody associatedwith viral hepatitis.Science170:332-333.