JOURNALOF VIROLOGY, Oct. 1974,p.821-833 Copyright0 1974 AmericanSociety for Microbiology
Vol. 14, No. 4 Printed in U.S.A.
Polypeptide
Composition
of
Urea- and
Heat-Resistant
Mutants
of
Poliovirus Types
1
and
2
ROBERT FENNELL' AND BRUCE A. PHILLIPS
Department of Microbiology, University ofPittsburgh, School of Medicine, Pittsburgh, Pennsylvania15261
Received for publication 16May 1974
Five
urea-resistantand
twoheat-resistant
mutants ofpoliovirus types 1 and 2 wereisolated and theirstructural
and
nonstructuralpolypeptides compared
tothose of
theirwild-type, parental
strains in an attempt to correlate mutantphenotypes with
alterations
inspecific
capsid polypeptides.
Four of the sevenmutantswere
found
tocontain
polypeptides
whichdiffered
inmolecularweight
from
their
respectiveparental viruses.
However, resistance of
virionstoheat-orurea-inactivation
could
not beattributed
tochanges
inparticular capsid
polypeptides
because alterations were detected in all but one of thecapsid
components. For two of the urea-resistant mutants and one heat-resistant
mutant, no
differences
werefound
in the molecularweights
of thecapsid
andnoncapsid polypeptides. These results,
and the fact that at least 12 selectivetreatments were
required
to obtain stable mutants, indicate that:(i)
suchphenotypes probably
canbeexpressed
by
mutationsaffecting
one ormoreof thelarger
capsid
polypeptides,
and(ii)
suchphenotypes
reflectmultiple
mutational
steps.Urea-resistant and heat-resistant
mutantsof
poliovirus
can
be
isolated
by
the
repeated
inactivation
(with heat or urea) and
propaga-tion of the surviving virus
population (6,
16).
The resistance to inactivation
is agenotypic
character of virus (2, 3, 6, 10, 12,
17).
Acomparison of the inactivation kinetics of the
RNA
obtained
from
the resistant mutants and
the sensitive
parental viruses showed that the
resistance to inactivation
could
be attributed
to analteration in the
capsid
structureand
not to anincreased
stability
of
the nucleic acid
(3,
6).
This
study
wasundertaken
toinvestigate
(i)
whether the capsid polypeptides from
urea-resistant
and heat-resistant mutants could
be
differentiated from their
parental
viruses
by
polyacrylamide gel electrophoresis;
and
(ii)
whether alterations in
specific
viral
polypep-tide could be
correlated
with resistance
to urea orheat inactivation.
MATERIALS AND METHODS
Cellculture. S-3HeLacellswere usedexclusively.
Suspensioncultures were maintained in Eagle
mini-mal essentialmedium (MEM) made with spinner salts
(Grand Island Biological Co., Grand Island, N.Y.) supplemented with 5% calf serum (Flow Laboratories,
Rockville, Md.) and2.0mMglutamine.AA/50 MEM
isminimalessential medium(Eagle)withspinnersalts
IPresent address: Microbiological Associates, Inc.,
Walk-ersville, Md. 21793.
containing
l/50th
the normal amount of each aminoacid.
Viruses. The wild-type poliovirus strains used in
theseexperimentsweretype 1,LSc;type 1,Mahoney
(Mah);type2,MEF,;and type 2, Sabin(2-Sab).The
type1viruseswerelaboratorystocks. The type 1,LSc
strain and its urea-resistant mutant, LSc-UR (see
below), were obtained from J. Hallum and J. S.
Youngner. The type 2, Sabin vaccine strain was
obtained from LederleLaboratories,PearlRiver,N.Y.
In all cases, uncloned virus stockswere used.
Urea-resistant (UR) and heat-resistant (HR) mutants of
these virus stockswereisolatedasdescribed below.
Propagation ofviruses. HeLa cells in suspension
culture(4x 101 perml)wereinfected with virus with
an input multiplicity of 10 PFU per cell. Thirty
minutes afterinfection, 5% calfserum wasadded.Six
and one-half hours after infection, the cells were
centrifuged and thecell pelletwasresuspendedina
small volume of 0.02 M phosphate buffer, pH 7.0
(PB).Afterrepeated (threetofourtimes)freezingand
thawing to rupture the cells, the cell debris was
removedbycentrifugationand the supernatantfluid
wascollected. The viruswaspelleted(70,000 x gfor 3
h), sonicallytreatedin aRaytheonsonicoscillatorfor
2min,resuspendedinPB, and assayed for infectivity.
Isolation ofurea- and heat-resistant mutants.
Urea-resistant and heat-resistantmutantsof thefour
wild-type (nonresistant) strains of poliovirus were
obtained by the method described by Hallum and
Youngner (6). Thewild-typeviruseswerepropagated
to titers ofapproximately 109PFU per ml. Prior to
urea-inactivation, the virus stockwasdiluted 10-1 in
PBandsonicallytreatedfor60sinaRaytheon sonic
821
on November 10, 2019 by guest
http://jvi.asm.org/
oscillator. The virus then was added to an equal
volume of 6 M urea in PB and the mixture was
incubated for 2 h at 37 C withoccasional mixing. The
virus wasdiluted 10-2in 20ml of MEM and added to
monolayer cultures. The procedure was repeated and
after each treatmentand passage, the virus
popula-tionwastested for urea sensitivity.Approximately 12
passages wererequired to obtain a stable population
of virus which showed no significant drop in titer
whentested for ureasensitivity.
The same experimental design was used for the
isolation ofthe heat-resistant mutants. Virus stocks
were sonically treated, diluted 10- 2 in PB, and
exposed to 50 C for2h. The virus then wasdiluted
10-fold in 20 ml of MEM and passaged in cell
monolayers as above. The isolation of heat-resistant
viruses likewiserequired about 12passages oftreated
viruspopulations.
Test ofurea sensitivity. Virus in PB(37 C) was
added to anequal volume of6MureainPB, pH 7.0,
at 37C. At designated times, 0.1-ml samples were
diluted 10-2 in cold MEM andassayedforinfectivity.
The results of virus inactivations were expressed as
the
log,0
of the surviving infectivity, PIPo, where Pequals the titer of survivorsattime, t, andPo is the
titer of virusat zero time. Theinfectivity was
mea-sured by the agar cell-suspension technique using
HeLacells (1).
Testofheatsensitivity. Virus,whichwas
preincu-bated at22C,wasdiluted 10-2 in 9.9 ml of PB at 50 C
(± 0.25C). At the designatedtimes, 0.1-mlsamples
were diluted 10-2 in cold MEM and assayed for
infectivity. The results of heat-inactivation
experi-ments wereexpressedalsoasthe
log,0
ofthesurviving infectivity.Preparation ofcytoplasmic extracts containing radioactivelylabeledvirus-specificproteins. Cells
insuspensionculture (4 x 106/ml)containing 4
jg
ofactinomycin D per ml and 3 mM guanidine were
infected withaninputmultiplicityof50PFU per cell.
Two hours after infection thecellswerewashed free of
guanidine (reversal) andresuspendedinamino
acid-deficient medium (AA/50 MEM). Radioactive amino
acids were added 90 to 120 min after guanidine
reversal. Seven hours after infection the cells were
centrifuged into a pellet (800 x g for 10 min), resuspended in reticulocyte standard buffer (RSB;
0.01MNaCl,0.0015MMgCl2,0.01MTris, pH 7.4)to
aconcentrationof 107cells perml,anddisruptedina
glass Dounce homogenizer. Nuclei and intact cells
were removedby centrifugation (1,000 x g for4min).
Thesupernatant fraction wasusedas acytoplasmic
extract.
Preparation ofpurifiedviruslabeled with
radio-active amino acids. Suspension cultures of cells
(4 x 107perml)were infectedat amultiplicityof50
PFU per cell. Calf serum was added 45 min after
infection. Four hours after infection, the cells were
centrifuged andresuspendedto8 x 106cellsperml in
AA/50 MEM containing 4 mM glutamine, 5% calf
serum, and labeled amino acidmixture (10uCi/ml).
Six andone-half hours after infection thecellswere
centrifuged into apelletand resuspendedin PBto a
concentration of4 x 107 cellsperml. Thecellswere
disrupted byfreeze-thawing. Cellular debris was
re-moved by centrifugation and the supernatant fluid
was layered on an 11 ml, 10 to 30% sucrosegradient
containing PB and 10mM EDTA. Thegradientwas
centrifuged in an SW-41 rotor, 52,000 x g for 18 h.
After centrifugation the gradient was continuously
scanned at 260nmby pumping thegradient througha
continuous flowcellin aGilford recording
spectropho-tometer.Usually, about 20 fractions werecollected. A
portion of each fractionwasassayed for radioactivity.
The virus and empty capsid bands were located as
peaks of radioactivity, optical density, and, in the
case ofvirions, infectivity.
The virus or empty capsid fractions were pooled
anddialyzed overnight against 1,000 volumes of PB.
Thedialyzed material was brought up to 4.0 ml with
PB. Two grams ofCsClwas added and the mixture
was centrifuged to equilibrium in a SW 39 rotor
(37,000 RPM for 18-24 hours at 5 C). Five drop
fractions were collected from thebottom of the tube.
Aportion of each fraction wasassayed for
radioactiv-ity. The virus or noninfectious empty capsids were located as peaks of infectivity and radiactivity, re-spectively.
Polyacrylamide gel electrophoresis of viral polypeptides. Purified virus, empty capsids, or cyto-plasmic extracts were dissociated by the addition of
one-tenth volumes of glacial acetic acid, 5 M urea,
and 10% sodiumdodecyl sulfate (SDS). After 30 min
of incubation at 37C, the dissociated polypeptides
weredialyzed overnight against 6,000 volumes of 0.01
Mphosphatebuffer, pH 7.0, containing 0.1% SDS, 0.5
M urea, and 0.01 M2-mercaptoethanol (14).
Immedi-ately prior to electrophoresing, the samples were placed in boiling water for approximately 5 min.
Samples containing the dissociated radioactive
viralpolypeptides in 5% sucrose were layered on 10%
acrylamide gels (20 cm by 5 nm) containing 0.5 M
urea, 0.1%SDS, and 0.1 M phosphate buffer, pH 7.0.
After electrophoresis, the gels were fractionated and
collected automatically into scintillation vials by a
SavantAutogeldivider and fraction collector.
Approx-imately 100 fractions were collected for each gel.
Thegelfractions weredried and 0.3 ml of hydrogen
peroxide (30%) was added to each vial. All vials were
incubated at 37 C overnight. Threedropsof
Soluene-100(PackardInstrument Co, Downers Grove, Ill.) or
NCS reagent (Amersham-Searle, Arlington Heights,
Ill.) wereadded and the sealed vials incubated
over-nightat 37C.Onedropof saturatedascorbicacid and
12.5 ml ofAquasol scintillation fluid (New England
Nuclear Corp., Boston, Mass.) were added and the
fractionswerecounted in a Packard model3375
scin-tillation spectrometer. Discrimination settings were
adjusted to give 10% spillover of
["4C]CPM
intothetritium channel and0.1%spilloverof [3H]CPM into
the
"4C-channel.
With these discrimination settingsthe counting efficiency was approximatey 45 and
65% for 3Hand
'4C,
respectively, whencompared totheradioactivity determinedwhenthe channelswere
set for optimal, nondiscriminating counting. The
re-covery of radioactivity from the fractionated gels
varied between70and 95%.
Reagents and isotopes. Radioactive amino acid
on November 10, 2019 by guest
http://jvi.asm.org/
MUTANTS OF POLIOVIRUSTYPES 1AND 2
tmixtures
(14C and 3H) were obtained from New England Nuclear Corp., Boston, Mass. ActinomycinD wasobtained from Merck Sharp & Dohme, West
Point, Pa. Guanidine-hydrochloride was purchased
from Eastman Organic Chemicals, Rochester, N.Y.
The urea used for the inactivation kinetics was an
ultra-pure grade purchased from Schwartz-Mann,
Orangeburg, N.Y.
RESULTS
Inactivation kinetics. The
exposureof
wild-typestrains of
poliovirus
type 1 to3 M
ureaat37
C
resulted
inamulti-hit
type of inactivation
kinetics
(Fig.
1).
The maximum
drop
in
titer wasobtained by
15 minof
inactivation,
afterwhich time the
surviving
fraction of virus
re-mained
unchanged.
Type
2strains
weresignifi-cantly
moreresistant
tourea-inactivation
(Fig.
1),
aninteresting
finding
because
mosttype
2strains
appear tobe
moreheat resistant
than
type 1strains
(16).
Their inactivation
kineticsmore
closely
resembled
single-hit
kinetics.
De-+1.0
0
-.1.0
t
-2.0 +
0
0
9-0
0
-3.0
-4.0 t
-5.0 +
spite the
greaterinitial resistance of the
type 2strains,
ascompared
to type 1viruses,
the
samenumber
of urea exposuresand
passages wasrequired before
astable
population
ofre-sistant
mutants wasobtained.
Itwasfound that
if a viruspopulation
exhibited,
for
example,
a0.5
log10
drop
in titer in 15min, such
a viruspopulation,
uponfurther
passage inthe
ab-senceof the
selection
teatment,would
revertrapidly
to aless
resistant state.In
contrast,all of the urea-resistant
mutantsexhibited
nosignificant inactivation
after 15 min in 3M
urea at 37C.
Theinactivation
kinetics of the
mutants weredetermined
afteratleast
twopassages of
the
viruswithout
ureatreatment
and reflect the kinetics of the
popula-tions
used
insubsequent
experiments.
Two strains of viruses
wereused
toobtain
heat-resistant
mutants.Heat inactivation of
these viruses
appeared
tofollow
single-hit
ki-netics
and
again the
wild-type
2strain
was moreresistant
than the
wild-type
1strain
(Fig. 2).
As0 5 10 15 20 25 30
MINUTES IN 3M UREA
FIG. 1. Urea-inactivationkineticsofthe urea-resistantmutantsand theirwild-typeviruses. Virus inPB,at
37C,wasaddedtoanequalvolumeof6 MureainPB and incubatedat37C.Sampleswerediluted inMEMfor
aplaque assay ofthesurviving infectivity. P = PFUIml at the sample time; P0 = PFU/ml at zero time.
Symbols:LSc, 0;LSc-UR,0;MEF1, x;
MEF1-UL,
®;Brun,0;Brun-UR, U;Mah, A;Mah-UR, A; 2-Sab,O;2-Sab-UR, O.
2-BAB
\>
\s
"
s
~~~~-L
-"B
X% _ _ O
s
~~~BRUN
823
VOL.14,1974
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.502.107.399.324.617.2]AND PHILLIPS
MINUTES AT 500C
1 2 3
+1.0 0 -1.0 04 -2.0 0 -3.0 1 -4.0 -Le0 4
FIG. 2. Heat-inactivation kinetics of the
heat-resistantmutantsand theirwild-type viruses.A 1-ml
amountof virus (at22C)wasadded to9.0 mlofPB
preincubatedat50C. At various times, sampleswere
diluted in MEM (at 22C) and assayed for the
survivingvirus.
noted above, about 12 passages ofvirus, each following heatexposure, werenecessaryinorder togetstable mutants.
Polyacrylamide gel electrophoresis of
vi-rion
polypeptides. Representative electro-pherograms are shown in Fig. 3 and 4. Highly purified, radioactivelylabeled virions werepre-pared for electrophoresis as described. The polypeptides from the mutant viruses were co-electrophoresed with those of theirrespective parental strains. Controls consisted of co-elec-trophoresing 3H- and "4C-labeled preparations of the same virus, as exemplified in Fig. 3A.
Differences in the polypeptide patterns were detected in fourof thesevenpairsofviruses. A
split in the VP-2a polypeptide of the Brun-UR virus was consistently found (Fig. 3D). The VP-1 polypeptide of the LSc_UR virus was approximately 5,000 daltons smaller than the
VP-1 of the parental LSc virus; and the VP-3
polypeptide of the LSc_UR virus was approxi-mately 4,000 daltons larger thanthat ofthe LSc
virus (Fig. 3E). The 2-Sab_UR virus contained
both a VP-1 and a VP-2 component smaller
than theirrespectivecomponentspresentinthe
parental virus, 2-Sab (Fig. 3F). The
heat-resist-ant mutheat-resist-ant of the LSc strain, LSc-H ,
con-tained a BP-3component of greater molecular
weight (migrated slightly slower)thanthe VP-3
polypeptide of the parental,
LSc virus (Fig.
4A).No differences
weredetected
inthe polypeptide
compositions of the urea-resistant
mutant of type 1, Mahoney(Mah-UR),
oreither of
the
mutantsderived
from type 2,MEF, (MEF,-HR
and
MEF1_UR).
No
differences
were everde-tected in the VP-4
polypeptides when the
elec-trophoresis
wascarried
outfor
shorter times so asto retainthis
component onthe gel
(Fig. 3A).Polyacrylamide
gel electrophoresis of
emptycapsid
polypeptides.
The73S
emptycapsids of the
mutantsand their respective
parental viruses
werealso examined
for differ-ences inpolypeptide composition. The
electro-pherograms showed the
samealterations of the
polypeptides
asseenin
the virion
polypeptides
(Fig. 5). The
LSc-UR
emptycapsid
polypep-tides,
VP-1 and
VP-3,
were, respectively,smaller and
larger than the corresponding
poly-peptides
of
the
parental LSc strain (Fig. 5C).
There
was somedifficulty in the identification
of certain
polypeptides (see
arrowFig.
5C).
When
such aberrant
polypeptides
arose, it wasdifficult
toadhere
tothe standard definition
ofpoliovirus polypeptides described by Maizel
etal.
(8).
So the question remained
as towhether
agiven
polypeptide could be defined
as one of theVP-2
orVP-3
polypeptides.
The
VP-1 and VP-2 polypeptides
ofthe
2-Sab-UR
emptycapsids
weresmaller than those
ofthe
parental
2-Sab
virus(Fig.
5E). The
2-Sab-UR
emptycapsids, unlike the
virions,contained several
componentsmigrating the
region of its
VP-1;
one ofthese polypeptides
migrated coincidently with the VP-1
of itsparental
type(see bracketer region, Fig. 5E).
The
type 1,Brunhilde
emptycapsids contained
verylittle VP-2
component;therefore,
anypos-sible differences
inthe
VP-2polypeptides
could
notbe detected
(Fig. 5D). The
emptycapsids of
all the strains
examined,
except type 1,Brun-hilde,
contained
amajor VP-2 component(s).
The
electrophoresis
of
the
polypeptides
from
the
type2,
MEF1
strain
repeatedly showed
asingle
major VP-2 polypeptide (Fig. 5B). None of the emptycapsids
from the resistant mutantsshowed
anydifference
from theirrespective
parental
types inthe
mobility
oftheir
NCVP-6
polypeptide.
It
proved difficult
toobtain
partially purified
emptycapsids
fromthe
heat-resistant mutants.No
significant differences between
parental and
mutant virusescouldbe detected; however,
the amount ofradioactivity
in the emptycapsid
polypeptides
wasvery small(Fig. 6).
Polyacrylamide
gel
electrophoresis of
cyto-plasmic
polypeptides. Radioactively labeled,
virus-specific
polypeptides
frominfected cells
Ift O~0MIEF1
HR_
K
LSO-HR\
No
"I- N NN %MP
N\
.I ^° _ X E
F-e~~~~~loW,MnP
\~~~~1..----N.. LS
824
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.502.59.249.75.309.2]800
6001
10 20 30 40 50 60 70 80 FRACTION
90
O 10 20 30 40 50 60 70 80 90 FRACTION
E
OLSC NL
LSc-UR ,. > >
_
LS
1400-200 1000
IL8oo
600-
400-200 100
0.
UL
a
IL
u
0 10 20 30 40 50 60 70 80 90 1O0 0 10 20 30 40 50 60 70 80 90 100
[image:5.502.60.447.43.572.2]FRACTION FRACTION
FIG. 3. Polyacrylamide gel electrophoresis ofthe dissociated virionpolypeptides from the urea-resistant
mutantsand theirwild-type viruses. Theradioactively labeledpolypeptides fromdissociated virionswere
lay-eredon 10%oacrylamide-SDS gels (10cm by6mm), electrophoresed, fractionatedintoscintillation vials,and
counted for radioactivity. Panel A. Co-electrophoresis of two preparations of type 1. Mahoney virion
polypeptides. Electrophoresis was for 15 hat 80 V, 8 mA. Symbols: [3H]Mah, 0; [14C]Mah, 0. Panel B.
Co-electrophoresis oftype1,Mahoney,andMah-U' virionpolypeptides. Electrophoresiswasfor22 h at 80V,8
mA.Symbols:
["4C]Mah,
0; [SH]Mah-UR,0.Panel C.Co-electrophoresis oftype2,MEF1 and MEF1-LR virionpolypeptides.Electrophoresiswasfor22hat80V,8mA.Symbols: ['4C]MEF,, 0; [sH]MEF,-UR,0.PanelD.
Co-electrophoresis oftype1,Brunhildeand Brun-U' virionpolypeptides. Electrophoresiswasfor16 h at 80V,7
mA. Symbols: [14C]Brun,0; [SH]Brun-UR, 0.PanelE. Co-electrophoresis oftype 1,LSc and LSc-UR virion
polypeptides. Electrophoresis wasfor 21 h at80 V,8 mA. Symbols: [14C]LSc, 0; [3H]LSc-UR, 0.Panel F.
Co-electrophoresis oftype2, Sabin and2-Sab-UI virionpolypeptides.Electrophoresis wasfor15 h at 80V, 7
mA. Symbols: ["4C]2-Sab, 0; [3H]2-Sab-UR, 0.
825 a.
A
oMoh *Mah
0.
I
It
B
o Moh *Moh-UR
0.
10 20 30 40 50 60 70 80 90 1I FRACTION
C
0 MEF,
* MEF,-UR ra.
b-I
g_\\_~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
sw
400
2001
400
3001
a2
u200 100 F
300
200 a1
1001
FRACTION
7
CL
q
on November 10, 2019 by guest
http://jvi.asm.org/
600
400
20
200
F RACTJON
U
100 F R ACTION
FIG. 4. Polyacrylamide gel electrophoresis of the dissociated virion polypeptides from theheat-resistant
mutantsand theirwild-type virions. Preparation andelectrophoresis of virionpolypeptideswasthesame asin
Fig.6.PanelA.Co-electrophoresis of type 1, LSc and LSc-HR virionpolypeptides.Electrophoresiswasfor22h
at 80 V, 8 mA. Symbols: [3H]LSc, 0;
[14C]LSc-HR,
*. Panel B. Co-electrophoresis of type 2, MEF1 andMEF1-HR virion polypeptides. Electrophoresis was for 15 h at 80 V, 7 mA. Symbols:
[14C]MEF,,
0;[3H]MEF,-HR,
*.826 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.502.56.450.61.614.2]VOL. 14, 1974 MUTANTSOFPOLIOVIRUSTYPES 1AND2
JV\J T
A
If&
O MOh Q
~CL
>L
* Mah-U"
o9 200
CL
0 10 20 30 40 50 60 70 80 94
FRACTION
0 10 20 30 40 50
FRACTION
60 70 80 90
0~~~ ~~
-~
~~~0
0 10 20 30 40 50 60 70 80 90
[image:7.502.53.448.63.579.2]FRACTION
FIG. 5. Polyacrylamide gel electrophoresis ofthe dissociatedemptycapsidpolypeptides fromthe urea-resist-ant mutants and their wild-type viruses. The purified empty capsids were dissociated and prepared for
electrophoresison10%,oacrylamide-SDS gelsasdescribedinMaterials andMethods.Panel A.
Co-electrophore-sis oftype 1,Mahoney and Mah-U" empty capsidpolypeptides. Electrophoresis wasfor20 hat80 V, 7 mA.
Symbols: [3H]Mah, 0; [14C]Mah-UR, 0. PanelB. Co-electrophoresis oftype2,MEF1 andMEF1-URempty
capsidpolypeptides. Electrophoresiswasfor15.5 hat80V,8 mA.Symbols: [3H]MEF-UIR, 0; [14C]MEF,,0.
PanelC.Co-electrophoresis oftype1,LSc and LSc-URemptycapsid polypeptides. Electrophoresiswasfor20 h at80 V,8 mA.Symbols: [3H]LSc, 0; ['4C]LSc-UL, 0. PanelD. Co-electrophoresis oftype1,Brunhilde and
Brun-UR empty capsid polypeptides. Electrophoresis was for20 h at80 V, 8 mA. Symbols: [14C]Brun, 0;
[3H]Brun-UL, 0. Panel E. Co-electrophoresis of type 2, Sabin and 2-Sab-UR empty capsid polypeptides.
Electrophoresis wasfor20hat80V, 8 mA.Symbols: ['4C]2-Sab, 0; [3H]2-Sab-UR, 0.
827
I I
C
O LSC
*LSC- IR
A~~~~~~~~~~~~~~~~~~~1
100
a.
C-) 0.C)
FRACTION
FRACTION
.9
a c
0
50
on November 10, 2019 by guest
http://jvi.asm.org/
80
60
A.
UL
40
20
0
100
80
60
A.
VL
40
20
0
0 10 20 30 40 50 60 70 80
F RACTION
0 10 20 30 40 50 60 70 80
[image:8.502.62.465.57.568.2]F
RACTION
FIG. 6. Polyacrylamide gelelectrophoresis ofthe dissociatedemptycapsidpolypeptides fromthe
heat-resist-ant mutheat-resist-ants and their wild-type viruses. The purified empty capsids were dissociated and prepared for electrophoresison10%acrylamide-SDSgels asdescribed in
Materials
andMethods. PanelA.Co-electrophore-sisoftype2,
MEF,
and MEF-HR emptycapsid polypeptides. Electrophoresiswasfor22hat80V,8 mA.Sym-bols:
[3HIMEF1,
Q;
[14C]MEF1HR, *. Panel B. Co-electrophoresis oftype 1,LSc andLSc-ULP empty capsidpolypeptides. Electrophoresis wasfor22h at80 V,8 mA.
Symbols: [14C]LSc,
0;[3H]LSc-HR,
*.were
prepared
andelectrophoresed
asdescribed
Mah-UR-infected cells repeatedly migratedabove. Unlike
the VP-3polypeptide
seeninthe slightly faster than the VP-3polypeptide
oftheelectropherograms
of the virion andempty cap- type 1,Mahoney-infected
cells(Fig. 7A).
Asid
polypeptides,
the VP-3 component from thesimilar
disparity
wasdetected
in theon November 10, 2019 by guest
http://jvi.asm.org/
MUTANTS OF POLIOVIRUS TYPES 1AND2
1750
1000
0
500
°IL
a
7
6
a.
2
40 50 60 70 80 90 100
FRACTION
0 10 20 30 40 50 60 70 80 90
FRACTION
600
400
IL
200
1200
1000
a Soo
600I
400 200
0 10 20 30 40 50 60 70 80 90 100
FRACTION
FIG. 7. Polyacrylamide gel electrophoresis of radioactively labeled, viral-specific, cytoplasmic polypeptides
from cells infected with the urea-resistantmutantsand theirwild-type viruses. Cytoplasmicextractscontaining
radioactively labeled, viral-specific polypeptideswereprepared forelectrophoresison10%acrylamide-SDS gels
as described in Materials and Methods. Panel A. Co-electrophoresis of type 1, Mahoney and Mah-UR
cytoplasmic polypeptides. Electrophoresiswasfor15hat80V,8mA.Symbols: [l4C]Mah-UR, Q;
[3H]Mah,
*.Panel B. Co-electrophoresis oftype1,Brunhilde and Brun-U" cytoplasmic polypeptides. Electrophoresiswas
for20 hat80V, 8mA.Symbols: [14C]Brun-U5, 0;
[3H]Brun,
*. Panel C. Co-electrophoresis oftype2,MEF,and
MEF,-UR
cytoplasmic polypeptides. Electrophoresiswasfor16hat80V,8mA.Symbols: [14CJMEF1,0;['H]MEF,-U", 0. Panel D. Co-electrophoresis of type 1, LSc and LSc-U'1 cytoplasmic polypeptides.
Electrophoresiswasfor24hat80V,8 mA.Symbols:
[3H]LSc,
0; [14C]LSc-UR, 0.Panel E.Co-electrophoresisoftype2, Sabin and 2-Sab-U' cytoplasmic polypeptides. Electrophoresiswasfor15hat80V,8mA.Symbols:
[3H]2-Sab,
0; [l4C]2-Sab-UR, Q.VOL. 14, 1974
IL
829
-B | St_N - o EBrun
4 >- 0 Brun-U 2>>>
0
FRACTION
4000
3000
II
C oMEF,
0* MEF-UR
Ii
1-F
Ne>
>_~~~~~~~o
0 10 20 30 40 50 60 70 80 90 IC FRACTION
00
300
200 '
tL
100
8
6
*0
4 2
3
12
10o
I r, T
D A. oLSC
4C *LSC-UV
0.0.>7 r
>.l (.0. a.
u>z
j
z >>
'I
II~~~~~~~~~~~~~~~~~~~~~~~~~~, III
II -I
I
Iu
E ~~& o 2-Sob
I I IL
>
>^>f
(L lL022-Sob-UnI0 >
I
r
I
11
O 10 20 30
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.502.50.451.57.557.2]0 10 20 30 0 5, 60 70 So 90
FRACTION
* 1i 26 36 40 s5 To
t6
90FRACrON
83() 1400
1200
100W
IL u 600
400
200
0
12
1400
12W
a.
4W0
200
S
on November 10, 2019 by guest
http://jvi.asm.org/
MUTANTS OFPOLIOVIRUS TYPES 1AND 2
phoresis
ofthe LSc-UH
cytoplasmicpolypep-tides. The
electrophoresis of the LSc-UR virionand
emptycapsids polypeptides
had shownthatthe
VP-3polypeptide(s)
waslarger
than the VP-3component
oftheLSc virus. No differencebetween
thecytoplasmic
VP-3components wasdetected (Fig. 7D). Three
of the resistant vi-ruses had shownalterations
inthe
mobility
ofthe
virion VP-1polypeptide
(Brun-U',
LSc_UR
,2-Sab-UHl). Examination
of thecytoplasmic
extractsrevealed that cells infected
withthese
virusescontained multiple
components whichmigrated
in theVP-1 regions
(Fig.
7B,D, and
E). In addition, the NCVP-lb polypeptide
fromthe LSc_UR-infected cells migrated slightly
faster than
theNCVP-lb polypeptide
of theparental LSc
virus(Fig. 7D).
Nodifference wasdetected
inthe
NCVP-la
components andagain
nodifference
wasdetected
in theNCVP-6
polypeptides.
A
similar
analysis
ofthe
cytoplasmic
poly-peptides synthesized
inMEF1-HR- andLSc-HR-infected cells showed that there
were nodetect-able differences between the
mutants andtheir
parental
types(Fig. 8).
DISCUSSION
Five urea-resistant
and twoheat-resistant
mutants wereisolated
from five strains ofpolio-virus
types 1and
2.The
polyacrylamide
gel
electrophoresis
of thepolypeptides
ofthese
virusesindicated
that four ofthe
seven mutantscontained
apolypeptide(s) which differed
inmolecular weight
fromthe
corresponding
[image:11.502.257.452.91.246.2]poly-peptide
oftheir
respective
parental
virions. A summary ofthese
findings
isshown
inTable
1.The urea-resistant
orheat-resistant
phenotype
of
the
virioncould
notbe attributed
to analteration
in oneparticular polypeptide
be-cause,depending
onthe
virusstrain, differences
were seen inall but
oneofthe
capsid
polypep-tides
(namely,
VP-4).
Nodifferences
werede-tected in
anyof the
noncapsid viral
polypep-tides with the
exception
ofthe
NCVP-lb
compo-nent ofthe
LSc-UR
virus(Fig. 7D). These
findings
are notsurprising
in that there is nocovariation with
respect tothe
urea-and
heat-resistant markers (6). It should be emphasized thatonly
differences in the molecular weights of the viralpolypeptides
weredetectable
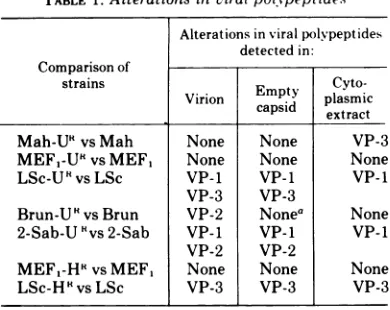
in. these experiments. Amino acid substitutionsTABLE 1.Alterations inviralpolypeptides
Alterationsinviralpolvpeptides detected in: Comparisonof
strains
Empty
Cyto-Virion
capsid
plasmic casd extractMah-U5
vs Mah None None VP-3MEF,-U5
vsMEF, None None NoneLSc-U5
vsLSc VP-1 VP-1 VP-1VP-3 VP-3
Brun-UHvsBrun VP-2 Nonea None
2-Sab-U 'vs 2-Sab VP-1 VP-I VP-1
VP-2 VP-2
MEF,-H5
vsMEF, None None NoneLSc-H'vsLSc VP-3 VP-3 VP-3
aThere was insufficient amount of VP-2 in extracts
to reliably detect differences between wild-type and
mutantviruses.
in the
polypeptides
thatdid not affect cleavage reactionswould
notbe detected.
The
electropherograms of the viral
polypep-tides
from emptycapsids and cytoplasmic
ex-tractsgenerally supported the similarities
ordifferences found
by examination of
virionpoly-peptides. One should
notethat misleading
re-sults
might be obtained
ifonly the
cytoplasmic
extractsfrominfected cells
wereexamined. For
example, the electrophoresis
of thecytoplasmic
extractof
Mah-UR-infected cells often revealed
aslight alteration
inthe
VP-3polypeptide;
purified
virionsshowed
nosuch difference.
Similarly, the electrophoresis
ofthe
cytoplas-mic
extractsof
LSc-UR-infected cells did
notshow
the altered VP-3
polypeptide repeatedly
detected
inthe
purified
virions.Therefore, the
examination ofthe
structural polypeptides
ap-pearsbest
accomplished by using
purified
vi-rions
and,
inaddition, other virus-related
parti-cles. The
use ofcrude
cytoplasmic
extractsalone
maygive incomplete
ormisleading
re-sults.
We
detected
inthe
emptycapsids
of mostviral
strains a significant amount of the VP-2polypeptide(s). The
amount ofthis
componentvaried
frombeing
amajorcomponent in type 1,LSc and
type 2,MEF1
emptycapsids
(Fig.5B
and C) to a very minor component in type 1,Brunhilde
empty capsids (Fig. 5D). Early re-ports on themorphogenesis
of type 1, Mahoneypoliovirus
claimed that the emptycapsids
didFIG. 8. Polyacrylamidegelelectrophoresis of radioactively labeled, viral-specificcytoplasmicpolypeptides
from cells infected withtheheat-resistant mutantsand theirwild-type viruses. Procedurewasthesame asin
Fig. 7.PanelA. Co-electrophoresis of type 2, MEF, andMEF,-HR cytoplasmic polypeptides.Electrophoresis
wasfor16hat80 V,8mA.Symbols: [14C]MEF,,0;
[3H]MEF,-HR,
*. Panel B.Co-electrophoresis of type 1,LSc and LSc-HR cytoplasmic polypeptides.Electrophoresiswasfor16hat80V,8 mA.Symbols: [14C]LSc,0;
[sHILSc-HR,*.
8.31
VOL.14,1974
on November 10, 2019 by guest
http://jvi.asm.org/
notcontain the VP-2
polypeptide
(7). The VP-2and
VP-4polypeptides
werepresumably
formed fromthe
cleavage
ofthe
NCVP-6 polypeptide
during the final association of the
emptycapsid
withthe viral RNA
toform virions. Aninterme-diate
structure,the
provirion, was recentlycharacterized by Fernandez-Tomas and
Bal-timore
(5).
Evidence presented here indicates
that
the
emptycapsids
of mostpoliovirus
strainsdo
contain aVP-2
polypeptide
(also see ref.15).
It maybe
thatthe
VP-2polypeptides
found
inthe
emptycapsids
are notthe
same asthose found in the virions after encapsidation
of theviral RNA.
Of
particular interest was the VP-2polypeptide found
in the empty capsidsand virions of
the
2-Sab-UR
virus (Figs. 3F and5E). This
VP-2polypeptide
was amajor compo-nentof
the
emptycapsid particle and had the
samemolecular weight
astheVP-2
polypeptide
found in its parental virus. Assuming that
emptycapsids
areprecursors tothe virion,
these
data
suggestthat the
emptycapsids
possess aVP-2 component which
maybecome
part ofthevirion
capsid. Vanden Berghe and Boeye
(15)reported
thatpurified poliovirions
containedthree VP-2 polypeptides.
These investigatorsalso
reported the isolation
of twotypes of emptycapsids containing different
amounts of theVP-2
polypeptide. Our experiments
alsoindi-cated
multiple VP-2 components
in certainstrains of
poliovirus.
Atleast
one ofthe VP-2 components maybe derived from the
emptycapsid.
It
isclear that the cleavage of the NCVP-6
polypeptide
(41,000
daltons)
cannotproduce
twodistinct VP-2
polypeptides,
eachwith
amolecular weight
of 28,000 to26,000 and a VP-4polypeptide with
amolecular weight
of5,500.One
possible explanation
isthat the
twoVP-2
polypeptides
areformed
by
the
ambiguous
cleavage of individual NCVP-6 molecules.
Therefore, the VP-2 polypeptides detected
inthe virions
represent acomposite of
amixture of
particles;
somevirions contain
the smaller VP-2
polypeptide
and others contain thelarger
one.Cooper
etal.
(4)
firstsuggested
thatambiguous
cleavage
of precursorpolypeptides
wasrespon-sible
forthe
formation ofalteredcopies
of thecapsid
polypeptides.
Rekosh
(13) determined
that thegene order ofthe
capsid
polypeptides
wasVP-4-VP-2-VP-3-VP-1. If
these capsid
polypeptides require
the entirelength
of theNCVP-la
precursor, thenthe
cleavage
ofNCVP-la occurs at twopoints:
between NCVP-6
andVP-3,
and between VP-3and
VP-1. The VP-4 and VP-2polypeptides
subsequently
form from thecleavage
ofNCVP-6. It
wasexpected,
therefore, that
a decrease ofseveral thousand daltons in
themolecular
weight of the VP-1
polypeptide (as in
the LSc-URand 2-Sab_UR
virions) would
pro-duce
acorresponding increase in the size of the
VP-3polypeptide.
Likewise,
adecrease in the
sizeof the VP-3
polypeptide (as in the LSc-HR
virus) should be
paralleled by
an increase inthe
size
of the VP-1
and/or NCVP-6 polypeptides.
The
LSc-U R virus showed both
adecrease in the
size of its
VP-1
polypeptide
and
anincrease in
the size of its VP-3
polypeptide (Fig.
3E). The
2-Sab-UR virus showed
noincrease in the size of
itsVP-3 component
despite the decreased size
of itsVP-1
component(Fig. 3F).
Likewise, there
was noincrease
inthe size
ofthe VP-1
polypep-tide of the LSc-H R
virus inspite of the
de-creased size of its VP-3
component(Fig. 4A).
Since
nodifference
was everdetected in the size
of the NCVP-la
precursormolecules, it
seemsthat there
mustbe
portions of the NCVP-la
precursormolecule that
do
notbecome the
capsid
polypeptides
after
cleavage.
It is
possi-ble,
therefore,
that there
are morecleavage
sites
than
orginally hypothesized.
Alternatively,
there
may exist amechanism for
trimming the
initial
cleavage products
tothe
propersize.
Some
ofthe
polypeptides
ofthe
mutantsmayexhibit
analtered
sensitivity
tothe
trimming
mechanism,
possibly
as aconsequencof
confor-mational
changes due
to anamino acid
substi-tution,
thereby producing
polypeptides
ofal-tered size.
ACKNOWLEDGMENTS
We gratefully acknowledge the technical assistance of Marie Urban and Navin Patel.
ThisresearchwassupportedbyaPublic HealthResearch grant Al 08368 from the National Institute ofAllergyand Infectious Diseases.
LITERATURECITED
1. Cooper, P. D. 1961.An improved agar cell-suspension plaque assay for poliovirus; some factors affecting efficiencyofplating. Virology15:153-157.
2. Cooper, P.D. 1962. Studiesonthe structure and func-tions of the poliovirion: effect of concentrated urea solutions.Virology 16:485-495.
3. Cooper,P.D. 1963. Effectofconcentratedureasolution onpoliovirusstrainsadaptedtodifferentgrowth tem-peratures.Virology21:322-331.
4. Cooper, P.D.,D.F.Summers, and J. V. Maizel. 1970. Evidence forambiguityintheposttranslational cleav-age ofpoliovirus proteins.Virology41:408-418. 5. Fernandez-Tomas, C.B.,andD. Baltimore.1973.
Mor-phogenesis ofpoliovirus. II. Demonstration ofanew intermediate, the provirion. J. Virol. 12:1122-1130. 6. Hallum, J. V., and J. S. Youngner. 1966. Urea and
thermal mutants of poliovirus. J. Bacteriol. 91:2305-2308.
7. Jacobson, M., and D. Baltimore. 1968. Polypeptide cleavages in the formation of poliovirus. Proc. Nat. Acad.Sci. U.S.A. 61:77-84.
on November 10, 2019 by guest
http://jvi.asm.org/
MUTANTS OF POLIOVIRUS TYPES 1AND 2
8.Maizel, J. V., and D. F. Summers. 1968. Evidence for differences in sizeand composition of the poliovirus-specific polypeptides in infected HeLa cells. Virology 36:48-54.
9. Maizel, J. V.,B. A. Phillips, andD. F. Summers. 1967. Composition of artificially produced and naturally occurringemptycapsids of poliovirus Type1.Virology 32:692-699.
10. Papaevangelou,G. J., and J. S. Youngner. 1961. Thermal stability of ribonucleic acid from poliovirusmutants.
Virology 15:509-510.
11. Phillips, B. A., and R. H. Fennell. 1973. Polypeptide composition ofpoliovirions, naturally occurringempty capsids, and 14S precursor particles. J. Virol.
12:291-299.
12. Pohjanpelto, P. 1961. Two different variants ofpoliovirus. Virology 15:231-236.
13. Rekosh, D. 1972. Gene order of the poliovirus capsid proteins. J. Virol. 9:479-487.
14. Summers, D. F., J. V.Maizel, and J. E. Darnell. 1965. Evidence for virus-specific noncapsid proteins in poliovirus infected HeLa cells. Proc. Nat. Acad. Sci. U.S.A. 54:505-513.
15. Vanden Berghe, D., and A. Boeye. 1972. New polypep-tides ofpoliovirus. Virology 48:604-606.
16. Youngner, J.S. 1957. Thermal inactivation studies with differentstrainsofpoliovirus. J. Immunol. 78:282-290. 17. Youngner, J. S., and J. V. Hallum. 1964.
Cystine-independent thermoresistant mutants of poliovirus. J. Bacteriol. 88:265-266.
VOL. 14, 1974



![FIG. 4.Fig.MEF1-HRatmutants[3H]MEF,-HR, 80 Polyacrylamide gel electrophoresis of the dissociated virion polypeptides from the heat-resistant and their wild-type virions](https://thumb-us.123doks.com/thumbv2/123dok_us/1579783.110597/6.502.56.450.61.614/hratmutants-polyacrylamide-electrophoresis-dissociated-virion-polypeptides-resistant-virions.webp)
![FIG. 5.antsisPanelElectrophoresiselectrophoresisSymbols:Brun-URatcapsid[3H]Brun-UL, 80 of Polyacrylamide gel electrophoresis of the dissociated empty capsid polypeptides from the urea-resist- mutants and their wild-type viruses](https://thumb-us.123doks.com/thumbv2/123dok_us/1579783.110597/7.502.53.448.63.579/antsispanelelectrophoresiselectrophoresissymbols-uratcapsid-polyacrylamide-electrophoresis-dissociated-polypeptides-mutants-viruses.webp)

![FIG. 7.forElectrophoresisandPanelfromradioactivelyasofcytoplasmic['H]MEF,-U",[3H]2-Sab, type described Polyacrylamide gel electrophoresis of radioactively labeled, viral-specific, cytoplasmic polypeptides cells infected with the urea-resistant mutants and](https://thumb-us.123doks.com/thumbv2/123dok_us/1579783.110597/9.502.50.451.57.557/forelectrophoresisandpanelfromradioactivelyasofcytoplasmic-described-polyacrylamide-electrophoresis-radioactively-cytoplasmic-polypeptides-resistant.webp)