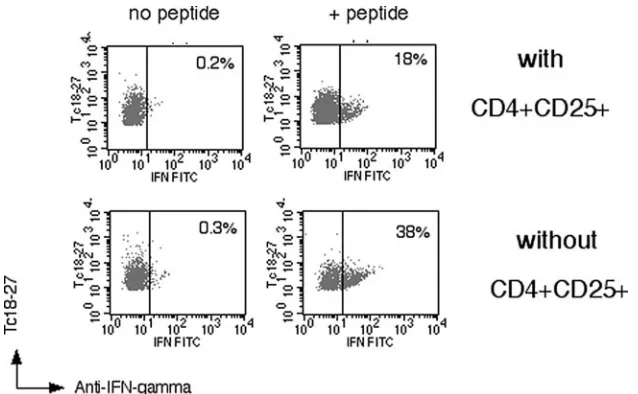
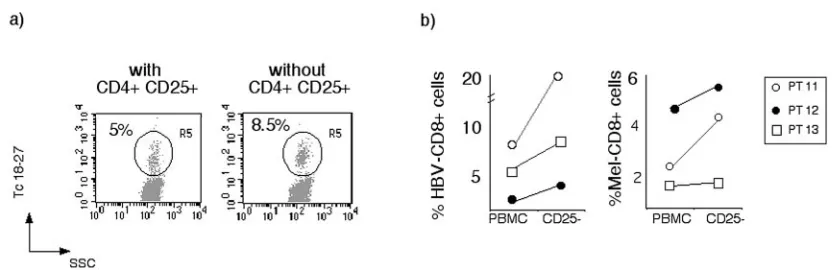
Modulation of the CD8+-T-Cell Response by CD4+ CD25+ Regulatory T Cells in Patients with Hepatitis B Virus Infection
7
0
0
Full text
Figure


Related documents