Herpesvirus Latent Origin of Replication That Are Essential for the
Efficient Establishment and Maintenance of Intact Plasmids
Prabha Shrestha, Bill Sugden
McArdle Laboratory for Cancer Research, University of Wisconsin—Madison, Madison, Wisconsin, USA
ABSTRACT
The maintenance of latent Kaposi’s sarcoma-associated herpesvirus (KSHV) genomes is mediated in
cis
by their terminal repeats
(TR). A KSHV genome can have 16 to 50 copies of the 801-bp TR, each of which harbors a 71-bp-long minimal replicator element
(MRE). A single MRE can support replication in transient assays, and the presence of as few as two TRs appears to support
estab-lishment of KSHV-derived plasmids. Why then does KSHV have such redundancy and heterogeneity in the number of TRs? By
determining the abilities of KSHV-derived plasmids containing various numbers of the TRs and MREs to be established and
maintained in the long term, we have found that plasmids with fewer than 16 TRs or those with tandem repeats of the MREs are
maintained inefficiently, as shown by both their decreased abilities to support formation of colonies and their instability,
result-ing in frequent rearrangements yieldresult-ing larger plasmids durresult-ing and after establishment. These defects often can be overcome by
adding the Epstein-Barr virus (EBV) partitioning element, FR (i.e., family of repeats), in
cis
to these plasmids. In addition we
have found that the spacing between MREs is important for their functions, too. Thus, two properties of KSHV’s origin of latent
replication essential for the efficient establishment and maintenance of viral plasmids stably are (i) the presence of
approxi-mately 16 copies of the TR, which are needed for efficient partitioning, and (ii) the presence of at least 2 MRE units separated by
801 bp of center-to-center spacing, which are required for efficient synthesis.
IMPORTANCE
KSHV is a human tumor virus that maintains its genome as a plasmid in lymphoid tumor cells. Each plasmid DNA molecule
encodes many origins of synthesis. Here we show that these many origins provide an essential advantage to KSHV, allowing the
DNAs to be maintained without rearrangement. We find also that the correct spacing between KSHV’s origins of DNA synthesis
is required for them to support synthesis efficiently. The identification of these properties illuminates plasmid replication in
mammalian cells and should lead to the development of rational means to inhibit these tumorigenic replicons.
K
aposi’s sarcoma-associated herpesvirus (KSHV) is an
onco-genic human herpesvirus causally associated with the
endo-thelial cell-derived tumor, Kaposi’s sarcoma (KS), and
lym-phoproliferative disorders, including primary effusion lymphoma
(PEL) and multicentric Castleman’s disease (MCD) (
1–4
). KSHV
is present in KS and PELs primarily in the latent phase of its life
cycle as a multicopy plasmid (
2
,
5–7
). KSHV belongs to the
gam-maherpesvirus subfamily and is related to another oncogenic
gammaherpesvirus, Epstein-Barr virus (EBV). Various latently
expressed genes of both EBV and KSHV have been shown to
con-tribute directly to cell survival and proliferation. Forcing the loss
of EBV genomes from EBV-positive Burkitt’s lymphoma,
post-transplant lymphoproliferative disorder (PTLD), and
PEL-de-rived cell lines can induce apoptosis and affect cell growth,
indi-cating that the lymphoma cells depend upon EBV for survival
and/or proliferation (
8–10
). Attempts to isolate PEL cells after
eviction of KSHV have not been successful (
11
), suggesting that
the PELs are dependent on latent KSHV genomes for their survival
and/or proliferation. Hence, given the importance of the
persis-tence of latent KSHV genomes in the associated tumors, it is
de-sirable to understand the factors that allow their maintenance
sta-bly in infected cells.
The maintenance of latent KSHV genomes in proliferating
host cells involves synthesis or replication of the viral DNA during
the S phase of the cell cycle and subsequent partitioning of the
newly synthesized viral DNA into the daughter cells during
mito-sis. Synthesis is mediated in
cis
by its origin of latent replication
located within its terminal repeat (TR) (
12
) and in
trans
by a viral
protein, LANA1 (latency-associated nuclear antigen 1) (
12–16
).
The TR is an 801-bp highly GC-rich unit and harbors a
71-bp-long minimal replicator element (MRE). The MRE, consisting of
two LANA1-binding sites (LBS 1 and 2) and an upstream GC-rich
replication element (RE), is the minimal element that supports
synthesis of KSHV plasmids (
17
). LANA1 mediates licensed
syn-thesis of the viral plasmid by binding to LBS 1 and 2 through its
DNA-binding and dimerization domain located in the C terminus
and recruiting the cellular origin recognition complex (ORC)
(
18–22
). The partitioning of KSHV plasmids is thought to be
me-diated by tethering of viral plasmids to cellular chromosomes by
LANA1. Binding and localization of LANA1 to the cellular
chro-matin occur through protein-protein interactions between
Received14 March 2014Accepted11 May 2014
Published ahead of print14 May 2014
Editor:R. M. Longnecker
Address correspondence to Bill Sugden, [email protected]. Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.00742-14
on November 7, 2019 by guest
http://jvi.asm.org/
harbor the plasmids stably in the long term (
29
,
30
). The process
of establishment is not fully understood, but it appears to depend
on the efficiency with which a plasmid can be synthesized and
partitioned (
31–34
) and may involve epigenetic modifications to
the viral genomes (
30
,
35
).
KSHV genomes as measured in tumor biopsy specimens can
have 16 to 50 tandem repeats of the TR (
36–38
), and the presence
of as few as two copies of the TR appears to support establishment
of KSHV-based plasmids in proliferating cells (
12
). Why then
does KSHV have 16 to 50 copies of the TR? Two findings with EBV
have suggested a plausible answer to this question. In EBV, Rep*
acts as an auxiliary origin of Epstein-Barr nuclear antigen 1
(EBNA1)-dependent DNA synthesis (
39
). A single unit of Rep*
can support DNA synthesis only in the short term, but an octamer
of Rep* can support both the establishment and maintenance of
the EBV DNA stably in the long term (
40
). Based on these findings
with Rep*, we hypothesized that an increasing number of TRs
provides KSHV plasmids a selective advantage by increasing the
efficiency of their establishment. Further, a single MRE unit, in the
absence of rest of the TR, can support the synthesis of the
KSHV-based plasmids in transient assays (
17
). However, based on studies
done with EBV, an origin that can support synthesis in transient
assays may not support establishment and long-term
mainte-nance of the plasmids (
31
). Whether the MRE is also sufficient for
the establishment and long-term maintenance of the plasmid in
addition to synthesis is not known.
To characterize the
cis
-acting elements involved in KSHV’s
establishment and long-term maintenance, we generated
KSHV-based replicons containing different numbers of the TR
or MRE units and assessed their abilities to give rise to
drug-resistant colonies and be maintained in cells stably (i.e.,
with-out yielding rearrangements). We have found that tandem
re-peats of the MRE unit can indeed support establishment of
KSHV-based plasmids. Additionally, we have identified two
properties of the origin of latent replication that are essential
for the efficient establishment and maintenance of the KSHV
plasmids stably. The first is the requirement of approximately
16 copies of the TR, without which the plasmids are established
inefficiently and are unstable, likely reflecting a defect in their
ability to be partitioned. The second is the requirement of at
least 2 units of the MRE separated by an 801-bp
center-to-center spacing between each for optimal synthesis. Our studies
show also that only the spacing between each MRE unit and not
the actual sequence is important for the function of the TR as a
maintenance element.
MRE, respectively, were constructed by multimerizing 2 MREs between XbaI and NheI in plasmid 4033. Fragments containing 2, 8, or 16 copies of the MRE between BglII and NheI sites in plasmids 4033, 4036, and 4037, respectively, were inserted into BglII and XbaI sites in 1782 to construct MRE plasmids. To generate the MRE-spacer plasmid, plasmid 4126, which contains two copies of the 73-bp (region from bp 539 to 611 of the TR) MRE separated by a 729-bp spacer sequence, was constructed as a pIDTSMART-KAN minigene (IDT). The 729 bp of spacer sequence con-sists of one NheI site followed by 723 bp ofEnterobacteriaphageDNA (GenBank accession no.J02459.1[bp 5716 to 6438]) that cannot support replication in mammalian cells (FR--Luc) (39). A fragment containing the MRE-spacer unit from 4126 was inserted between the EcorV and XbaI sites of 1782 to generate the MRE-spacer plasmid.
Plasmids containing FR are based on plasmid 994, which contains EBV OriP in a G418 resistance backbone. The 8-TR/FR, 2-MRE/FR, 8-MRE/FR, 16-MRE/FR, and MRE-spacer/FR plasmids were constructed by replacing the dyad symmetry (DS) between the SpeI and BamHI sites in plasmid 994 with 8 TRs between SpeI and BglII sites from the 8-TR plas-mid, 2 MREs between BglII and NheI sites from plasmid 4033, 8 MREs between BglII and NheI sites from plasmid 4036, 16 MREs between BglII and NheI sites from plasmid 4037, and MRE-spacer between BamHI and XbaI from the MRE-spacer plasmid, respectively. The 2-TR/FR plasmid was constructed on a different vector, pLON-33k (42), which contains EBV OriP and lac operator (lacO) sites in a G418 resistance backbone. DS between SnaBI and SpeI sites in pLON-33k was replaced with 2 TRs be-tween NheI and HpaI from Z6-2TR to generate plasmid 4067. 4067 was digested with XbaI to delete lacO repeats and ligated back to generate the 2-TR/FR plasmid. The 2-TR/FR plasmid is thus larger than the 8-TR/FR plasmid due to a different backbone.
Plasmids used to characterize the Gardella gel contain either OriP or FR from EBV in backbones of various sizes. These plasmids include p4151 (6.7 kb; the MRE-spacer/FR plasmid described above), p994 (7.5 kb), p4147 (20.7 kb), and p4066 (40 kb).
Cell lines.The cell lines used for the replication assays include two PEL-derived cell lines, BCBL-1 and JSC-1; one epithelial cell line, SLK (43); and one human embryonic kidney-derived cell line, 293. Cell lines were cultured in either Dulbecco’s modified Eagle’s medium (SLK and 293) or RPMI 1640 medium (PELs) supplemented with 10% fetal bovine serum, 200 U/ml of penicillin, and 200g/ml of streptomycin sulfate at 37°C in a humidified 5% CO2atmosphere. SLK/LANA cells that stably express LANA1 were generated by transducing SLK cells with a retroviral vector encoding LANA1 driven by a cytomegalovirus (CMV) promoter and hygromycin B phosphotransferase. Selection with puromycin was performed at final concentrations of 1g/ml (SLK and BCBL-1) or 0.3 g/ml (JSC-1); selection with G418 was performed at final concentrations of 800 to 1,250g/ml (SLK), 600g/ml (JSC-1), or 400g/ml (BCBL-1); selection with hygromycin was performed at a final concentration of 400 g/ml (SLK).
Short-term replication assay.SLK/LANA cells were cotransfected with 20g of sample plasmids (2-, 8-, and 16-TR or -MRE plasmids and
on November 7, 2019 by guest
plasmid 1782) and 3g of p2134 (enhanced green fluorescent protein [eGFP] expression vector) using Lipofectamine 2000 (Life Technologies) as per the supplier’s recommended protocol. At 4 days posttransfection, cells were harvested, and the percentage of GFP-positive cells was deter-mined as a measure of the transfection efficiency. Low-molecular-weight DNA was isolated by the method of Hirt (44) with a few modifications and digested with 80 U of HindIII (plasmids with TRs) or XbaI (1782 and plasmids with MREs) overnight to linearize the plasmids. After the diges-tion was complete, 70% of the sample was digested with 120 U of DpnI overnight to digest bacterially methylated DNAs. Plasmids that have un-dergone at least one round of replication in the cells are resistant to diges-tion by DpnI. Complete digesdiges-tions with both enzymes were ensured by parallel digestions of 1.5g test plasmid in 10-l aliquots of each of the sample digests. Concentrations of 2⫻106to 2.5⫻106GFP-positive cell equivalents for positive (⫹) and negative (⫺) DpnI fractions of each sam-ple were used for Southern blotting.
Colony formation assay for measurement of establishment effi-ciency.Adherent cells (SLK) in 60-mm-diameter dishes were cotrans-fected using Lipofectamine with equimolar (approximately 3-g) amounts of sample plasmids and 1g of p2134 (eGFP expression vector). A total of 107PEL cells (JSC-1 and BCBL-1) were cotransfected with equimolar (approximately 10-g) amounts of sample plasmids and 2g of p2134 by electroporation. One or 2 days posttransfection, the percent-age of GFP-positive cells was determined as a measure of transfection efficiency, and adherent cells were plated at 104, 103, or 100 GFP-positive cells per 15-cm-diameter dish in triplicate for each dilution. PELs were plated at 103, 100, 10, or 1 GFP-positive cell per well of 96-well plates (48 wells for each dilution). Selection was applied for 3 to 4 weeks using an appropriate antibiotic (puromycin for the TR and pPUR plasmids and G418 for the MRE, TR/FR, MRE/FR, MRE-spacer, MRE-spacer/FR, 1782, and 994 plasmids), and the number of drug-resistant colonies (adherent cells) or the number of wells with drug-resistant cells (PELs) was counted to determine the CFU for each plasmid. For adherent cells, the CFU was measured as CFU⫽(no. of drug-resistant colonies/no. of GFP-positive cells plated)⫻100%.
For PELs, the CFU was calculated using the Poisson distribution as CFU⫽{[⫺ln(no. of negative wells/total no. of wells)]/no. of cells per well}⫻100%.
Statistical analysis.Any relationship between the number of TRs or MREs and the CFU was tested using the Jonckheere-Terpstra statistic for trend, a nonparametric test for ordered differences between groups. A two-sidedPvalue ofⱕ0.05 was assigned as statistically significant. Statis-tical analyses were performed using Mstat software, version 5.5 (N. Drink-water, McArdle Laboratory for Cancer Research, School of Medicine and Public Health, University of Wisconsin) and is available for downloading athttp://www.mcardle.wisc.edu/mstat.
Gardella gel.Gardella gels were performed as described previously (45) with modifications (46). A horizontal 20- by 28-cm (width by length) gel was prepared consisting of (i) resolution gel made of 0.75% agarose in 1⫻Tris-borate-EDTA (pH 8.2) (TBE) buffer and (ii) lysis gel made of 0.8% agarose, 2% SDS, and 1 mg/ml proteinase K in 1⫻TBE buffer. The lysis gel occupied the top 5 cm of the horizontal gel and included 10- by 1-by 6-mm (width 1-by depth 1-by height) wells for loading samples. Cells resuspended in 50l resuspension buffer (7% Ficoll, 100g/ml RNase A, and 0.01% bromophenol blue in phosphate-buffered saline) were loaded into the wells and electrophoresed initially at 0.8 V/cm for 3 to 4 h and then at 4.5 V/cm for approximately 16 h at 4°C. Upon completion, the gel was stained with 0.5g/ml ethidium bromide (EtBr) in 1⫻TBE for 20 min, visualized under UV, and then analyzed by Southern blotting or with a second gel run at 90° to the first gel as described below. A total of 2⫻106 to 3⫻106cells were used for Gardella gels unless otherwise noted.
2D gel.The first dimension of the two-dimensional (2D) gel was a Gardella gel, performed as described above. Following the Gardella gel, the lane containing the desired sample was excised with minimal spacing around the lane. The fragment was then embedded into a 1⫻
Tris-acetate-EDTA (TAE) gel with 0.6% agarose and 0.5g/ml EtBr on a 20- by 28-cm (width by length) gel tray such that the length of the fragment lies across the top of the gel, perpendicular to the direction of electrophoresis in the Gardella gel. This second gel was electrophoresed at approximately 1.5 V/cm for 20 to 24 h at room temperature. The gel was then analyzed by Southern blotting.
Isolation and digestion of total genomic DNA for Southern blotting. For Southern blotting of plasmids in cells after long-term replication as-says, total genomic DNA was isolated from approximately 3⫻107cells. Pellets were resuspended in 5 ml of 0.15 M sodium acetate (pH 7.5 to 8.0) and lysed with an equal volume of lysis buffer (50 mM EDTA, 0.4 M sodium acetate, 2% SDS, and 1 mg/ml proteinase K) with minimal vor-texing to prevent shearing of high-molecular-weight DNA. The samples were incubated at 45°C overnight and extracted sequentially with phenol and chloroform by rocking the samples for 2 to 4 h on a rotator, centri-fuging them at 4,000 rpm for 30 min, and transferring them to new tubes using wide-bore pipette tips. The samples were then ethanol precipitated and resuspended in 2 ml Tris-EDTA (TE) at 4°C overnight. They were incubated with 0.1% SDS and 50g RNase A at 37°C for 2 h and then with 100g proteinase K at 45°C for 2 h. The samples were extracted with phenol and chloroform as described above, precipitated with ethanol, and resuspended in TE. Twenty-five to thirty percent of the isolated DNA from cells was digested with 120 U of HindIII (TR and pPUR plasmids) or XbaI (MRE plasmids and p1782) to linearize the plasmids, and the com-pletion of digestion was ensured using a test plasmid in parallel digests as described above. Approximately 20g of digested DNA was loaded on a 0.8% agarose gel in 1⫻TAE buffer containing 0.5g/ml ethidium bro-mide and electrophoresed at 1.4 V/cm for 16 to 20 h for analysis by South-ern blotting as described below.
Southern blotting.Southern blotting was performed as described pre-viously (31). DNA was denatured and transferred to Gene Screen Plus hybridization membrane (NEN Life Sciences) using 10⫻SSC (1.5 M NaCl, 150 mM sodium citrate [pH 7]). A32P-radiolabeled probe was prepared using the Rediprime II random prime labeling system (GE Healthcare) and hybridized to the membrane in ULTRAhyb hybridiza-tion buffer (Ambion). The hybridized membrane was exposed to a storage phosphor screen, and the signals were captured using Storm 860 Phos-phorImager (Molecular Dynamics). TR and pPUR plasmids were de-tected with pPUR DNA; the MRE, TR/FR, MRE/FR, spacer, MRE-spacer/FR, and 1782 plasmids were detected with 1782 (pcDNA3.1) DNA, unless otherwise noted. Quantification of signals in Southern blots was done using ImageJ software (NIH). The final intensity for any particular signal was obtained by subtracting the neighboring background intensity within the same lane.
RESULTS
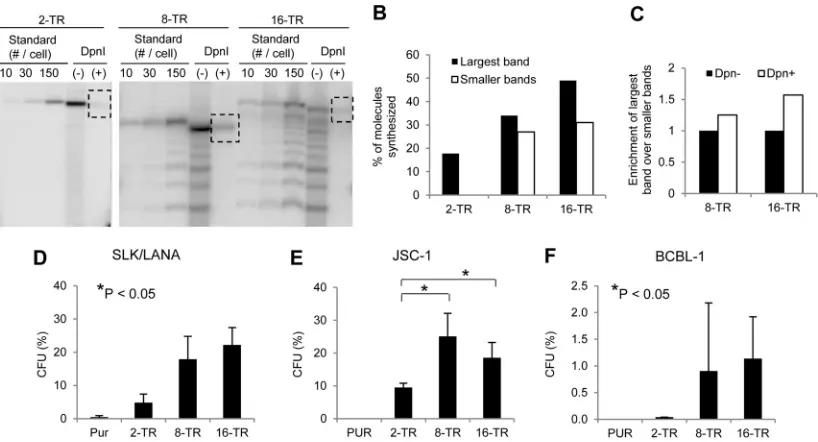
Increasing the numbers of TRs increases the efficiencies of
syn-thesis and establishment supported by KSHV plasmids.
We
constructed KSHV-based plasmids containing 2, 8, or 16 copies of
the TR and measured their abilities to support synthesis by
per-forming short-term replication assays in SLK cells stably
express-ing LANA 1 (SLK/LANA). We found a positive correlation
be-tween the number of TRs and the efficiency of synthesis (
Fig. 1A
and
B
). Additionally, we also observed that plasmids with higher
numbers of TRs are preferentially synthesized when a heterogeneous
mixture of plasmids containing different numbers of TRs is
intro-duced into cells. Plasmids with multiple copies of TRs from KSHV are
unstable in
Escherichia coli
, likely due to their high GC content and
repetitiveness. We have propagated plasmids containing TRs in
eight strains of
E. coli
variously deficient in recombination.
Plas-mids harvested from all strains displayed heterogeneity in the
number of TRs they contained. A single clonal
E. coli
population
obtained after transformation with homogeneous plasmid DNA,
containing 16 TRs, for example, harbored a mixture of plasmids
on November 7, 2019 by guest
http://jvi.asm.org/
with 16 or fewer TRs (
Fig. 1A
) (data not shown). When this
het-erogeneous population of plasmids was introduced into SLK/
LANA cells, the plasmids with higher numbers of TRs were
pref-erentially synthesized, as shown by an enrichment of the larger
plasmids over smaller plasmids in the DpnI (
⫹
) lane (
Fig. 1A
and
C
). Given that the synthesis of KSHV genomes is licensed (
22
),
these observations likely indicate that a higher number of TRs
provides KSHV plasmids with an increased probability that they
will undergo synthesis.
We also measured the abilities of these plasmids to support
establishment using colony formation assays. In addition to SLK/
LANA cells, we performed assays in JSC-1 and BCBL-1 cells,
pri-mary effusion lymphoma (PEL)-derived cell lines harboring
en-dogenous KSHV genomes, to simulate the environment in which
KSHV plasmids are maintained
in vivo
. We cotransfected cells
with a GFP expression plasmid and equal amounts of 2-, 8-, or
16-TR plasmids. Two days posttransfection, the cells were plated
at various dilutions of the GFP-positive cells and cultured for
ap-proximately 3 weeks in puromycin-containing media. The CFU
(percentage of successfully transfected cells that can give rise to
drug-resistant colonies) was used as a measure of efficiency of
establishment for each of the plasmids. In SLK/LANA and BCBL-1
cells, we observed a positive correlation (
P
⬍
0.05) by Jonckheere
Terpstra test between the number of TRs per plasmid and the
efficiency of establishment they support (
Fig. 1D
and
F
). In JSC-1
cells, plasmids with 8 and 16 copies of the TR were established with
significantly higher efficiencies (
P
⬍
0.05) than those with 2 copies
of the TR (
Fig. 1E
). It is important to note that the CFU for 2- and
8-TR plasmids observed here likely deviate from their actual
es-tablishment efficiencies due to recombination events of unknown
frequencies (shown below). Thus, these observed efficiencies for
2- and 8-TR plasmids should be viewed as overestimates of the
establishment efficiencies of the unrecombined, parental
plas-mids. The average efficiencies of establishment of 16-TR plasmids,
for which the observed CFU more accurately reflect the actual
establishment efficiency, varied between the recipient cell lines
with SLK and JSC-1 supporting about 20-fold-higher efficiencies
than BCBL-1 cells.
Plasmids with 16 copies of the TR are stable, while those with
2 and 8 copies of the TR are often maintained only as
recombi-nants.
We performed Gardella gels to determine the
extrachro-mosomal status and structure of the TR plasmids in cells after
establishment. Gardella gels involve the lysis of live cells in the
wells of the gel and allow separation of extrachromosomal and
chromosomal DNAs upon electrophoresis (
45
). We characterized
this technique to inform our experiments by analyzing the
migra-tion patterns of various forms of plasmids of different sizes from
different numbers of cells (
Fig. 2
). We found that the covalently
closed circular (CCC) form of plasmids derived from
in situ
lysis
of cells can migrate as two distinct bands in a Gardella gel (
Fig. 2A
FIG 1Efficiencies of short-term replication and establishment of KSHV-based plasmids with various numbers of TRs. (A) Southern blot analysis of a short-term replication assay of 2-, 8-, and 16-TR plasmids in SLK/LANA cells performed 4 days posttransfection. DpnI (⫺), DNA from⬃2⫻106GFP-positive cell equivalents digested with HindIII but not with DpnI; DpnI (⫹), HindIII- and DpnI-digested DNA from⬃2⫻106GFP-positive cell equivalents. Dotted boxes indicate bands obtained for DpnI-resistant replicated DNA. The standards were free parental plasmids digested with HindIII and loaded at 10, 30, or 150 plasmids per cell calculated for⬃2⫻106cell equivalents. Multiple bands in the lanes for the 8- and 16-TR DNAs were derived by recombination of the parental plasmids inE. coli. The largest bands in these lanes have 8 or 16 TRs, while the smaller bands have fewer than 8 or 16 TRs, respectively. The probe was pPUR (backbone of TR plasmids). (B) Quantification of signals in panel A. The percentage of synthesized DNA for each TR plasmid was calculated by measuring the intensities of the bands corresponding to total and replicated DNA in the (⫺) and (⫹) DpnI lanes, respectively, using the ImageJ program (NIH). White bars represent the synthesis of plasmids with fewer than 8 or 16 TRs in the lanes with 8- and 16-TR plasmids, respectively. (C) Enrichment of the largest plasmids over smaller plasmids in the lanes for 8- and 16-TR plasmids in panel A. Ratios of signal intensities of the largest band to the smaller bands were calculated in the (⫹) and (⫺) DpnI lanes, and the ratio in the (⫺) DpnI lane was set to 1. (D, E, and F) Establishment efficiencies of 2-TR, 8-TR, and 16-TR plasmids expressed as CFU in SLK/LANA (D), JSC-1 (E), and BCBL-1 (F) cells. CFU was determined as the percentage of transfected cells resulting in puromycin-resistant colonies⬃3 weeks posttransfection. Cells transfected with pPUR (Clontech) were used as the negative control. The error bars represent the standard deviations obtained from three independent experiments. Correlation analysis was performed using the Jonckheere-Terpstra test for a trend for increasing CFU, and statistically significant (Pⱕ 0.05) trends (D and F) and differences (E) are indicated and marked with asterisks.
on November 7, 2019 by guest
[image:4.585.93.502.66.287.2]and
B
). A faster-migrating band (band a) and a slower-migrating
band (band b) observed for a plasmid 4151 from JSC-1 cells (
Fig.
2A
), were both confirmed to be the CCC form of the plasmid by a
2D analysis following the Gardella gel (
Fig. 2B
). Both the CCC and
nicked forms of plasmids derived from cells migrate more slowly
than those of free plasmids, and this difference becomes greater as
more cells are used in Gardella gels (
Fig. 2C
and
D
). The slower
migration is likely a result of delayed exit of the plasmids from cells
due to their gradual
in situ
lysis and increased viscosity from the
increased number of cells. The migration of plasmids became
ab-errant at more than 3
⫻
10
6cells; thus, we have used up to 3
⫻
10
6cells in all of our Gardella gel analyses. We extended this study to
293 cells newly transfected with plasmids of various sizes and
found a similar trend (
Fig. 2E
) (data not shown). Under the
con-ditions of the Gardella gels we used in this study, plasmids of all
sizes derived from up to 3
⫻
10
6cells migrate between 77 and
100% as far as the free parental plasmids of the same size. Plasmids
that are retarded by more than 23% relative to their free parental
plasmids under these conditions therefore represent distinct,
larger species.
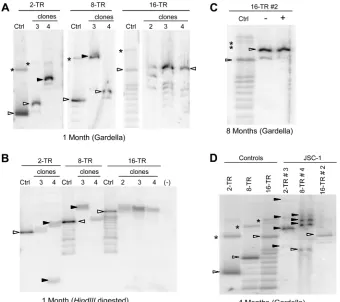
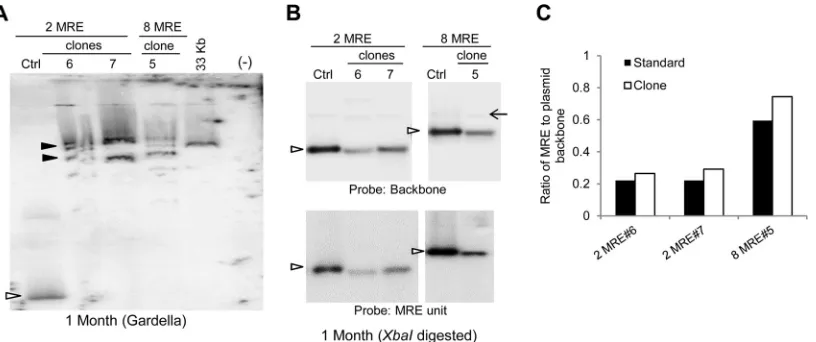
Gardella gels of some selected clones of JSC-1 cells harboring
TR plasmids showed that these plasmids were present as
extrach-romosomal DNAs (
Fig. 3A
). Clones of JSC-1 cells transfected with
2- and 8-TR plasmids often harbored plasmid DNAs that were
larger than the size of the plasmids that were initially introduced
into them (
Fig. 3A
, clones 2-TR 4 and 8-TR 3). These larger species
are not aggregates of smaller plasmids produced as artifacts of the
Gardella gel technique, as confirmed by probing for
mitochon-drial DNA in the same cells, which migrated as DNAs of
approx-imately 16 kbp (data not shown). This observation indicates that
these larger, recombined species have a selective advantage in the
recipient cells. In contrast to 2- and 8-TR plasmids, 16-TR
plas-mids were stable in that they were the same size as the parental
DNA in all clones tested and lacked detectable rearranged
prod-ucts (
Fig. 3A
and
Table 1
). Similar results were obtained when we
examined bulk populations of SLK/LANA and BCBL-1 cells
har-boring the TR plasmids (
Table 1
) (data not shown). To gain
in-sights into the structures of the recombined plasmids, we isolated
total genomic DNA from the cells 1 month posttransfection and
performed Southern blot analyses after digesting them with an
enzyme (HindIII) having a unique site in the plasmids (
Fig. 3B
).
Some of the recombined plasmids had gained an additional
Hin-dIII site (
Fig. 3B
, clone 2-TR 4), while some did not (
Fig. 3B
, clone
8-TR 3). In both of these cases, digestion with HindIII did not
FIG 2Characterization of Gardella gels. (A) Gardella gel showing migration of plasmid DNA derived fromin situlysis of JSC-1 cells. Ctrl (control), 0.3 ng of free parental plasmid p4151; JSC-1⫹4151,⬃3⫻106JSC-1 cells harboring established p4151 (6.7-kb) plasmid; O, position of wells; *, nicked circle form of the plasmid; L, linear DNA; arrowhead, covalently closed circular (CCC) form of the plasmid. CCC DNA from cells migrates as 2 bands: a faster-migrating band (a) and a slower-migrating band (b). E, CCC DNA of⬃170-kbp endogenous EBV plasmids. The probe was an⬃1.7-kb OriP fragment. (B) 2D gel of JSC-1 cells harboring p4151 showing that bands a and b in panel A are both derived from the CCC form of the plasmid and are of the same size. In the first dimension (1st), p4151 was run on a Gardella gel as for panel A. In the second dimension (2nd), the corresponding lane from the Gardella gel was embedded in a 1⫻TAE gel and electrophoresed perpendicular to the first dimension, as shown. Ctrl, mixture of plasmid and linearized forms of p4151 loaded at the beginning of the 2nd gel. (C) Gardella gel of various numbers of JSC-1 cells harboring p4151 showing that plasmids from cells have reduced mobility likely due to viscosity of the cells. Ctrl, 0.3 ng of free parental plasmid p4151. (D) Migration of plasmids in Gardella gels as a function of cell number, measured from panel C. Distances from the wells migrated by different DNAs, including the CCC and nicked forms of p4151 and the CCC form of EBV were measured. The percentage of migration indicates the distance moved by cell-derived plasmids expressed as a percentage of the distance moved by the same DNA/form in the Ctrl lane. (E) The difference in the migrations of cell-derived plasmids and their corresponding free parental plasmids is greater for larger plasmids. Plasmids of various sizes from newly transfected 293 cells were run on Gardella gels, and their migration from⬃3⫻106cells compared to that for free parental plasmids was measured. Gel 1 was electrophoresed for approximately 2 h longer than gel 2.on November 7, 2019 by guest
http://jvi.asm.org/
yield unit-length DNAs of the expected sizes. In contrast, plasmids
from cells transfected with 16-TR plasmids gave the expected
unit-length DNAs (
Fig. 3B
, 16-TR clones).
[image:6.585.123.463.66.368.2]In order to test further the finding that 16-TR plasmids are
structurally stable, we subjected clone 16-TR 2 of JSC-1 cells to
further selective pressure by removing antibiotic selection from
the culture media for 2 months and then subcloning the cells in
the presence of selection. If this clone contained minor,
recom-bined species below the level of detection at earlier time points, the
absence and then subsequent reintroduction of selection could
provide such minor species an opportunity to expand in the cell
population above the detection limit. A Gardella gel performed on
the subcloned cells a month after reintroduction of selection (8
months posttransfection) showed no evidence of recombination
or change in the composition of plasmids (
Fig. 3C
). Consistent
with our observation from short-term replication assays, we
ob-served a preferential establishment of plasmids with higher
num-bers of the TRs when a heterogeneous population of plasmids
containing various numbers of the TR was introduced into cells.
FIG 3Larger plasmids produced by recombination of 2- and 8-TR plasmids are selected during establishment in JSC-1 cells. (A) Gardella gels of representative clones of puromycin-resistant JSC-1 cells 1 month after transfection with 2-, 8-, or 16-TR plasmids. (Control and sample lanes for the 16-TR plasmid are from the same gel.) Ctrl, 0.3 to 1.5 ng of free parental plasmids corresponding to each TR plasmid; open arrowheads, bands obtained from CCC DNA of the free parental plasmid or from plasmids from cells that are the same size as the parental plasmid; closed arrowheads, bands obtained for CCC DNA of demonstrably larger, recombined plasmids from cells; *, nicked form of plasmids in control and sample lanes when distinctly detected. Multiple bands in the control lanes for 8- and 16-TR DNAs below the open arrowheads are derived by recombination of the parental plasmids inE. coli. (B) Southern blot analysis of the total genomic DNA from cells in panel A isolated 1 month posttransfection and digested with a single cutter enzyme (HindIII) of the plasmids. Control (Ctrl) lanes, 0.3 to 1.5 ng of free parental plasmids digested with HindIII; open arrowheads, bands in the ctrl lane that represent the expected migration of HindIII-digested parental TR plasmids; closed arrowheads, fragments of recombined plasmids from cells generated by HindIII digestion; (⫺), nontransfected parental cells. (C) Gardella gel of JSC-1 cells, clone 16-TR 2, 8 months posttransfection grown with or without G418. In the – lane, cells were grown in the absence of G418 for 2 months and then grown in limited dilution for a month after reintroduction of G418. In the⫹lane, cells were grown continuously in the presence of G418. (D) Gardella gel of cells in panel A 4 months posttransfection. Clones 2-TR 3 and 8-TR 4 have recombined, producing larger species at this time. The probe was pPUR (backbone of TR plasmids).TABLE 1Replicons with MREs and 2 or 8 copies of the TRs are often rearranged and/or integrated
Type of plasmid
No. of clones:
Testeda
With rearranged plasmidsb
With integrantsc
2-TR 8 5 3
8-TR 10 8 2
16-TR 8 0 0
2-MRE 5 3 2
8-MRE 4 3 1
16-MRE 6 3 3
a
Clones from SLK, JSC-1, and BCBL-1 cells.
bResults from Gardella gels and/or Southern blot analyses of DNAs isolated from cells
at 1 to 8 months posttransfection.
cClones that continue growing in the presence of antibiotic selection provided by the
plasmids, yet have no detectable presence of extrachromosomal DNAs by Gardella gel analyses when probed for the plasmid backbone of each plasmid.
on November 7, 2019 by guest
[image:6.585.39.287.568.669.2]This effect was most apparent for samples containing 16-TR
plas-mids (
Fig. 3A
,
B
, and
C
).
2- and 8-TR plasmids continue to recombine and yield larger
molecules after establishment.
It has been shown that
introduc-tion of DNA into mammalian cells through transfecintroduc-tion promotes
recombination (
47
,
48
). We therefore asked if the recombined,
larger plasmids derived from those initially having 2 or 8 copies of
TRs formed during transfection or continued to be formed and
selected over time. One month after transfection is sufficient for
the detectable presence of recombined plasmids with selective
ad-vantages; therefore, any species that is detectable only at time
points beyond 1 month should represent recombinants that were
formed subsequent to transfection. Clones of cells with apparently
homogeneous 2-TR and 8-TR plasmids at 1 month
posttransfec-tion yielded cells with larger, recombined plasmids at 4 months
posttransfection, while those with 16-TR plasmids carried
simi-larly didn’t harbor any recombined species (
Fig. 3D
). The clones
with recombined plasmids contained multiple species, the
major-ity of which were larger than the parental plasmids. Clone 8-TR 4
also contained nonrecombined 8-TR plasmid, albeit as a minor
species. Similar changes were observed in SLK/LANA and BCBL-1
cell populations harboring 2- and 8-TR plasmids (data not
shown). Some of the cells with 2- or 8-TR plasmids evolved to
harbor these plasmids as integrants at a later time point (
Table 1
),
as evidenced by their continued growth in the presence of
antibi-otic but lack of detectable presence of extrachromosomal DNAs
by Gardella gel analyses (data not shown). This observation
sug-gests that integration occurs and maintains these KSHV plasmids
more efficiently than extrachromosomal replication even after
es-tablishment. These findings indicate not only that the
recom-bined, larger plasmids continue to be formed over time but also
that the cell populations harboring them evolve in a dynamic
pro-cess favoring recombined species that are selected for.
Based on our observations that (i) the recombined species selected
over time are larger than the input parental plasmids, (ii) the plasmids
with higher numbers of TRs are preferentially synthesized and
estab-lished when a mixture of plasmids with various numbers of the TR is
introduced into cells, and (iii) plasmids with 16 copies of the TR are
structurally stable, we favor the notion that the recombined species
derived from parental plasmids with 2 or 8 TRs have an increased
number of TRs, potentially approaching 16 per molecule.
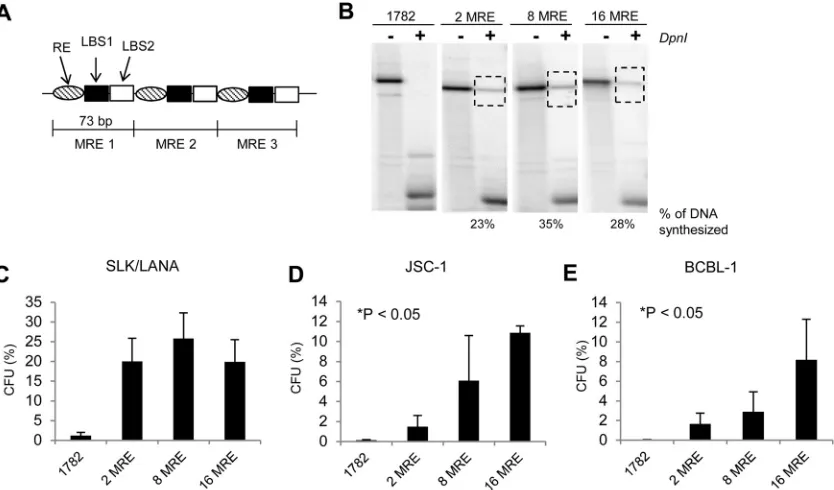
Efficiency of transient replication of KSHV genomes is
inde-pendent of the number of MRE units.
A single unit of the 71-bp
MRE within the TR can support synthesis of KSHV plasmids in
transient assays (
17
). We therefore asked whether the ability of
MREs to support synthesis also increases with the number of
MREs per replicon. We constructed plasmids containing 2, 8, or
16 tandem repeats of the MREs (
Fig. 4A
) and measured their
abilities to support synthesis by performing short-term
replica-tion assays in SLK/LANA cells (
Fig. 4B
). Unlike the plasmids with
intact TRs, the efficiency of synthesis of MRE-containing plasmids
did not correlate with the number of MREs per plasmid. This
FIG 4Efficiencies of short-term replication and establishment of KSHV-based plasmids with various numbers of MREs. (A) Schematic representation of the plasmids with MREs. The 73-bp units of the TR (bp 539 to 611) containing MRE are repeated in a head-to-tail fashion without any spacing between each unit. (B) Southern blot analysis of a short-term replication assay of 2-, 8-, and 16-MRE plasmids in SLK/LANA cells performed 4 days posttransfection. –DpnI, DNA from⬃2⫻106GFP-positive cell equivalents digested with XbaI but not with DpnI;⫹DpnI, XbaI- and DpnI-digested DNA from⬃2⫻106GFP-positive cell equivalents. The dotted boxes indicate bands obtained for DpnI-resistant replicated DNA. The numbers below the Southern blot represent the percentage of synthesized DNA for each MRE plasmid, calculated by measuring the intensities of the bands corresponding to total and replicated DNA in the⫺and⫹DpnI lanes, respectively, using the ImageJ program (NIH). The probe was p1782 (backbone of MRE plasmids). (C, D, and E) Establishment efficiencies of 2-MRE, 8-MRE, and 16-MRE plasmids expressed as CFU in SLK/LANA (C), JSC-1 (D), and BCBL-1 (E) cells. CFU was determined as a percentage of transfected cells resulting in G418-resistant colonies 1 month posttransfection. Cells transfected with p1782 (pcDNA3.1 from Invitrogen) were used as a negative control. The error bars represent standard deviations obtained from three independent experiments. Correlation analysis was performed using the Jonckheere-Terpstra test for a trend for increasing CFU, and statistically significant (Pⱕ0.05) trends are indicated and marked with asterisks.on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.85.502.67.312.2]observation suggests that even though the MRE is the minimal
element that can support synthesis, the additive effect of
addi-tional MREs on synthesis can only be obtained when the MREs are
present in the context of intact TRs.
Increasing numbers of MREs support increasing efficiencies
of establishment.
Studies from EBV show that an origin that can
support synthesis in the short term may not support
establish-ment. Raji ori, a second viral origin of replication of EBV that is
used predominantly for its maintenance in Raji cells, is unable to
support the establishment of EBV genomes even though it can
support maintenance of EBV genomes once established (
31
). We
therefore determined whether the MRE of KSHV, in addition to
supporting synthesis in the short term, also supports the
establish-ment and long-term maintenance of KSHV genomes. We
per-formed colony formation assays of plasmids containing 2, 8, and
16 tandem repeats of MREs in JSC-1, BCBL-1, and SLK/LANA
cells and found that all three MRE-plasmids can be established in
all three cell lines (
Fig. 4C
,
D
, and
E
). The efficiency of
establish-ment was positively correlated with the number of MREs in the
two PEL cell lines (
P
⬍
0.05) but not in SLK/LANA cells. It is
important to note, however, that all three MRE plasmids
under-went recombination of unknown frequencies (shown below);
hence the CFU observed here are likely overestimates of the
effi-ciencies of establishment of the unrecombined parental plasmids.
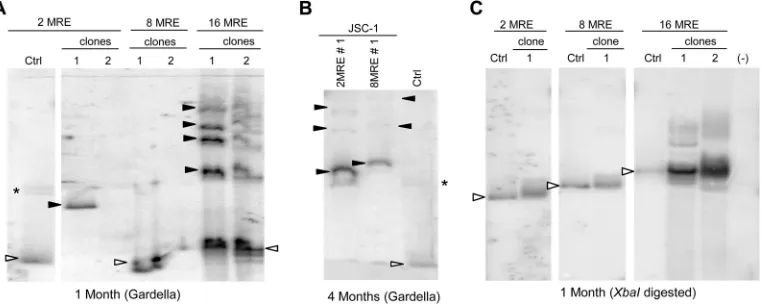
Tandem repeats of 16 MREs are not sufficient for the stable
maintenance of KSHV-derived plasmids.
Representative clones
of cells obtained 1 month after transfection with MRE plasmids
were analyzed by Gardella gels, which showed not only that the
MRE plasmids were maintained extrachromosomally but also
that they frequently recombined, producing larger species that
became established (
Fig. 5A
and
6A
). Similar to plasmids with 2
and 8 copies of the TR, plasmids with MREs also continued to
recombine postestablishment, producing larger and
heteroge-neous species (
Fig. 5B
). In some instances, cells harboring MRE
plasmids evolved to maintain them as integrants (
Fig. 5A
, clone 2
for the 2- and 8-MRE lanes;
Table 1
).
We analyzed the structures of the larger plasmids at 1 month
posttransfection by Southern blotting of the total genomic DNA
after linearization of the plasmid DNAs with XbaI (
Fig. 5C
and
6B
). A majority of recombined plasmids when digested with XbaI
yielded unit-length DNAs of the expected size, similar to the
pa-rental plasmids, indicating that the recombined plasmids were
head-to-tail concatemers of the input MRE plasmids. We
mea-sured the ratio of MRE units to the plasmid backbone in
recom-bined plasmids from SLK/LANA cells by quantifying the signals
obtained by probing for either the MRE unit or the plasmid
back-bone (
Fig. 6B
and
C
). We found that the ratio of MRE to backbone
DNA in recombined plasmids is similar to that in unit-length
parental plasmids (
Fig. 6C
), thus confirming that the recombined
plasmids are concatemers of the input MRE plasmids and have
more MREs per plasmid molecule relative to the input plasmids.
Hence, as is the case with 2- and 8-TR plasmids, the establishment
efficiencies obtained for the MRE plasmids are overestimates of
their actual efficiencies as a result of concatemerization and other
recombination events of unknown efficiencies.
Interestingly, unlike plasmids with 16 TRs, which were
main-tained stably (
Fig. 3
), plasmids with 16 MREs were unstable and
recombined to yield larger plasmids, often heterogeneous in size.
This finding indicates that tandem repeats of 16 MREs cannot
support maintenance stably, suggesting that they are deficient in
some aspect of maintenance compared to the TRs.
Addition of FR limits the instability of plasmids containing 2
and 8 copies of TRs but not of those containing MREs.
Plasmids
with MREs and lower numbers of TRs support synthesis less
effi-ciently than those with 16 TRs (
Fig. 1
and
4
). Hence, the inability
of plasmids with MREs and lower numbers of TRs to be
main-tained stably could be due to defects in their synthesis.
Addition-ally, these plasmids might also have defects in their partitioning
abilities. Maintenance of latent EBV genomes is mediated by two
distinct elements in its OriP: the dyad symmetry (DS) and the
family of repeats (FR). Four specifically positioned EBNA1-binding
sites in the DS mediate synthesis of the EBV genomes, while 20
FIG 5MRE plasmids are maintained in the long-term extrachromosomally, but larger recombinants are often selected. (A) Gardella gel of representative clones of G418-resistant JSC-1 cells 1 month posttransfection with 2-, 8-, or 16-MRE plasmids. All lanes are from the same gel. Ctrl (control), 0.3 to 0.5 ng of free parental plasmids. The sizes of the 2-, 8-, and 16-MRE plasmids vary by only⬃500 bp; hence, only one of these plasmids is used as the parental plasmid in the control lane. Open arrowheads, bands obtained from CCC DNA of the free parental plasmid or from plasmids from cells that are the same size as the parental plasmid; closed arrowheads, bands obtained for CCC DNA of larger, recombined plasmids from cells; *, nicked form of the control plasmids. No signals can be detected in the 2- and 8-MRE clones 2 even after multiple Gardella gel analyses. (B) Gardella gel of cells in panel A 4 months posttransfection. Clone 2MRE1 has become heterogeneous, and clone 8MRE1 has recombined, producing larger species at this time. (C) Southern blot analyses of the total genomic DNA from cells in panel A isolated 1 month posttransfection and digested with a single cutter enzyme (XbaI) of the plasmids. All lanes are from the same gel. Ctrl, 0.3 to 0.5 ng of parental plasmids digested with XbaI; open arrowheads, bands in the Ctrl lane that represent the expected migration of the XbaI-digested parental MRE DNA; (⫺), nontransfected parental cells. The probe was p1782 (backbone of MRE plasmids).on November 7, 2019 by guest
[image:8.585.104.485.68.220.2]EBNA1-binding sites in the FR mediate tethering to chromosomal
DNAs, which is thought to be required for partitioning of EBV
ge-nomes (
32
,
33
,
49–52
). It is plausible that similar to 20
EBNA1-bind-ing sites in the FR of EBV, approximately 16 pairs of LANA-1 bindEBNA1-bind-ing
sites in the TRs of KSHV are required for efficient partitioning of
KSHV genomes. We thus asked whether the instability of plasmids
with MREs and 2 or 8 TRs resulted from defects in their partitioning
abilities.
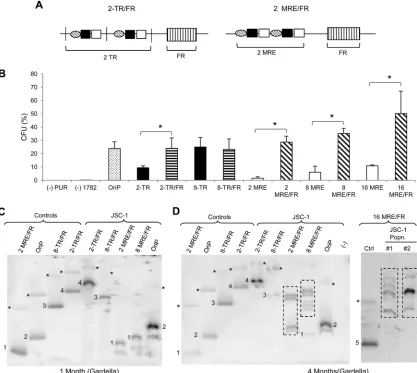
Elements supporting synthesis and partitioning of KSHV are
not distinct as those in EBV. To uncouple these processes in
KSHV-derived plasmids, we generated hybrid plasmids by
intro-ducing FR from EBV into plasmids with 2 or 8 copies of TRs or
MREs (
Fig. 7A
). FR on binding EBNA1 mediates the partitioning
of plasmids but not their synthesis, potentially complementing
any partitioning defects that plasmids with low numbers of TRs or
MREs might have. Colony formation assays performed in JSC-1
cells (dually positive for KSHV and EBV) revealed that the
pres-ence of FR in
cis
increased the efficiency with which 2 TRs
sup-ported establishment (
P
⬍
0.05) (
Fig. 7B
). In addition, the
pres-ence of FR also rendered plasmids with both 2 and 8 TRs stable:
that is, they did not yield larger, recombined plasmids over time
(
Fig. 7C
and
D
). This finding indicates that plasmids with few
copies of TRs are established with low efficiencies and are
struc-turally unstable because they have defects in partitioning. Hybrid
plasmids with 2, 8, or 16 copies of the MRE, on the other hand,
were not completely rescued by the presence of FR. The MRE
plasmids were established approximately 5-fold more efficiently
in the presence of FR than in its absence (
Fig. 7B
) but recombined
to yield larger species over time (
Fig. 7C
and
D
). This finding
indicates that although MREs have defects in partitioning similar
to plasmids with few copies of TRs, they also have some defect in
replication not shared with the TRs.
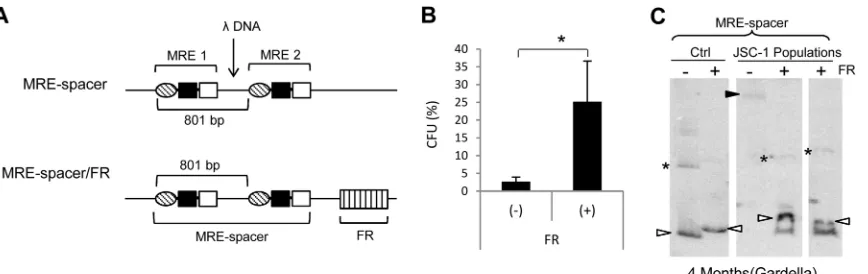
The 801-bp spacing between each MRE unit is essential for
the stable maintenance of KSHV plasmids.
MRE plasmids differ
from TR plasmids in that they lack the DNA sequence between
each unit of the MRE. The defect in replication exhibited by the
MRE plasmids even when complemented with FR could be due to
the absence of an unidentified element in the region of the TR
outside the MRE. Alternatively, the defect could be due to the
absence of an 801-bp center-to-center spacing between each MRE
unit. We explored the latter possibility by generating the
MRE-spacer plasmid containing a MRE-spacer sequence derived from DNA of
phage
between 2 MRE units such that the center-to-to center
spacing between each MRE unit becomes 801 bp (
Fig. 8A
). We
performed colony formation assays with the MRE-spacer plasmid
in the presence or absence of FR in
cis
. While the MRE-spacer
plasmid supported a low level of establishment and exhibited
in-stability similar to those of the 2-TR and 2-MRE plasmids, the
addition of FR increased its establishment efficiency and also
ren-dered the plasmid stable (
Fig. 8B
and
C
). This observation not
only indicates that the 801-bp spacing between each MRE unit is
essential for the stable maintenance of KSHV plasmids but also
suggests that the actual sequence between the MRE units does not
play an active role in the maintenance of KSHV genomes.
DISCUSSION
We have generated plasmid derivatives of KSHV varying in both
the number and the structure of its latent origin of replication to
define elements required for the efficient establishment and stable
long-term maintenance of KSHV genomes. We measured each
plasmid’s ability to support establishment and maintenance
effi-ciently and stably as (i) its ability to give rise to drug-resistant
colonies in colony-forming assays and (ii) its ability to be
main-FIG 6MRE plasmids are present as concatemers in cells. (A) Gardella gel of representative clones of G418-resistant SLK/LANA cells 1 month posttrans-fection with 2- and 8-MRE plasmids. Ctrl, 0.3 to 0.5 ng of free parental plasmids; open arrowhead, bands obtained from CCC DNA of the free parental plasmid; closed arrowheads, bands obtained for CCC DNA of demonstrably larger recombined plasmids from cells; (⫺), nontransfected parental cells; 33 Kb, 33-kb plasmid (pLON-33K, see Materials and Methods) used as a size marker. The probe was p1782. (B) Southern blot analyses of the total genomic DNA from cells in panel A isolated 1 month posttransfection and digested with a single cutter enzyme (XbaI) of the plasmids. Ctrl, 0.3 to 0.5 ng of parental plasmids digested with XbaI; open arrowheads, bands in the Ctrl lane that represent the expected migration of the XbaI-digested parental MRE DNA. The upper panel was probed with p1782 (backbone of MRE plasmids), and the lower panel was probed with approximately 150 bp of 2 MRE units obtained by NheI/BglII digestion of the 2-MRE plasmid. The same amounts of DNA were loaded onto both panels. Arrow, unspecific band. (C) Ratio of MRE to backbone sequence in MRE plasmids measured from panel B. Band intensities of MRE units (lower panel) and backbone DNA (upper panel) in panel B were measured using the ImageJ program (NIH), and the ratios of MRE to backbone DNA were obtained for control DNA (Standard) and DNAs from cells (Clone). The ratios of MRE to backbone sequence for all plasmids from cells are similar to those for control plasmids, showing that the larger plasmids are recombined head-to-tail concatemers.on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.92.500.66.237.2]tained in cells without yielding rearrangements displaying
selec-tive advantages.
We have found that increasing the number of TRs per replicon
provides a selective advantage to the replicon by increasing its
abilities to support synthesis, establishment, and structurally
sta-ble maintenance over the long term. This conclusion is supported
by four lines of evidence. First, increasing numbers of TRs support
increasing efficiencies of synthesis (
Fig. 1A
to
C
). Second, the
number of drug-resistant colonies that grew out in colony
forma-tion assays generally increased with an increasing number of TRs
(
Fig. 1D
to
F
). Third, plasmids with higher numbers of TRs were
both selectively synthesized and established when a heterogeneous
mix of plasmids with various numbers of TRs was introduced into
human cells (
Fig. 1
and
3
). Fourth, plasmids with initially 16
cop-ies of the TR were stable and maintained their size during and after
establishment, while those with initially 2 or 8 copies of the TR
were unstable (i.e., they often recombined, producing larger
mol-ecules that were selected for over time) (
Fig. 3
). We think it likely
that the larger molecules derived from those initially having 2 or 8
copies of the TR have more copies of the TR per molecule than
their parents, providing them with a replicative advantage. We
could test this notion with plasmids having MREs. Larger,
recom-bined species of plasmids containing MREs, which were selected
for over time, proved to be head-to-tail concatemers (
Fig. 6
) and
thus conclusively did have increased numbers of MREs per
mol-ecule compared to their parents.
Our findings and interpretations are consistent with a
previ-ously reported finding that plasmids containing 3 and 8 copies of
FIG 7Addition of FR incisto plasmids with TR or MRE. (A) Schematic representation of 2-TR and 2-MRE plasmids with FR. (B) Establishment efficiencies of TR and MRE plasmids in the presence or absence of FR in JSC-1 cells expressed as CFU. CFU was determined as a percentage of transfected cells resulting in drug-resistant colonies. OriP, plasmid containing EBV’s origin of latent replication, used as a positive control. pPUR (empty vector expressing puromycin resistance from Clontech) and 1782 (pcDNA3.1 from Invitrogen expressing G418 resistance) DNAs were used as negative controls. The error bars represent standard deviations obtained from three independent experiments. Statistically significant (Pⱕ0.05) differences as determined by the Jonckheere-Terpstra test are indicated and marked with asterisks. (C and D) Gardella gels from populations of drug-resistant JSC-1 cells 1 month (C) and 4 months (D) after transfection with TR/FR or MRE/FR plasmids. We have numbered signals from CCC DNA corresponding to the same size in control and sample lanes for clarity. (⫺), nontransfected parental cells; Ctrl (control), 0.3 to 0.5 ng of free parental plasmids. The 2- and 8-MRE/FR plasmids vary by⬃500 bp; hence, only one is used as the input parental plasmid in the control lane. *, nicked forms of the plasmids in the sample and control lanes when detected. Dotted boxes indicate bands obtained for recombined plasmids from cells. The parental 2-TR/FR plasmid is bigger than the 8-TR/FR plasmid due to the different sizes of the backbones of these plasmids (see Materials and Methods). The probe was p1782 (backbone of the plasmids).on November 7, 2019 by guest
[image:10.585.84.501.68.441.2]the TR recombine to yield larger plasmids, which are selected in an
EBV-negative Burkitt’s lymphoma cell line, BJAB (
12
), suggesting
that this phenomenon cannot be attributed to the particular cell
lines or the plasmid backbones used but rather to the low number
of TRs present in these plasmids. A similar phenomenon was also
observed for EBV in some cell lines. EBV OriP deletion mutants
containing 6 or few EBNA1-binding sites in the FR region support
establishment with low efficiencies, and the established plasmids
are maintained as rearranged plasmids containing head-to-tail
multimers of the initial plasmids (
32
). The presence of at least 7
copies of EBNA1-binding sites in the FR, however, completely
rescues the plasmids from their inability to be established by
sup-porting efficient partitioning (
32–34
).
Our observations with TR/FR hybrid plasmids indicate that the
instability and defect in establishment shown by plasmids with
low numbers of TRs reflect their inability to support efficient
par-titioning. The addition of FR in
cis
, along with EBNA1 in
trans
,
increased the ability of 2 TRs to support establishment and also
limited the instability of plasmids containing both 2 and 8 TRs,
rescuing them from their defective phenotype (
Fig. 7
). This
find-ing suggests that the level of synthesis with two copies of the TR is
sufficient for structurally stable maintenance despite more than 2
TRs supporting an increased level of synthesis. These observations
also provide key evidence in support of the model in which a
sufficiently long multimer of LBS 1/2 in the TRs functions as a
cis
-partitioning element of KSHV, analogous to a multimer of
EBNA1-binding sites in the FR of EBV. A recent study has shown
that the efficient synthesis of EBV also requires tethering of EBV
genomes to chromosomal DNAs through FR (
53
). Hence, it is
possible that the addition of FR to 2- and 8-TR plasmids indirectly
increases their efficiencies of synthesis by tethering these plasmids
more efficiently to chromosomal DNAs. This possibility warrants
further testing.
Approximately 16 copies of the TR seems to be sufficient for
KSHV-based plasmids to be established and maintained
effi-ciently and stably. However, that KSHV genomes can have up to
50 copies of the TRs (
36–38
) indicates that the number of TRs per
KSHV genome is modulated by factors in addition to
establish-ment and maintenance. One such factor could be headful
packag-ing of the viral DNA durpackag-ing lytic replication. EBV packages its
DNA as a function of the length of DNA that allows efficient
pack-aging. The length of DNA that is generated by the cleavage of
lytically replicated concatemeric DNA at EBV’s TR is optimized
for its packaging (
54–56
). Lytic replication of KSHV is in many
ways similar to that of EBV. It is possible that the wide range of
TRs per KSHV genome results from a similar need to package an
optimal length of DNA. This explanation is strengthened by
find-ings that the clonality of KSHV genomes with respect to the
num-ber of TRs varies with the level of lytic reactivation supported by
the host cells. For instance, in cells from MCD that exhibit a high
level of lytic reactivation, KSHV genomes are polyclonal with
re-spect to the number of TRs, while in cells from KS or PEL that
exhibit a lower level of lytic reactivation, KSHV genomes are
monoclonal or oligoclonal with respect to the number of TRs (
57
,
58
). We favor a model in which a dual selection imposed by stable
maintenance and headful packaging is responsible for the range in
the number of TRs per KSHV genome.
Interestingly, our findings indicate that the ability of
KSHV-derived replicons to support maintenance also depends on the cell
type. Of the three cell lines we have used, BCBL-1 cells support
establishment of introduced 16-TR plasmids with approximately
5% the efficiency in SLK and JSC-1 cells (
Fig. 1
). The level of
LANA1 is not likely to be the reason for these differences as we
found no significant differences in its level in the three cell lines
(data not shown). The levels of chromatin binding proteins
MeCP2 and DEK have been shown to affect the localization of
LANA1 to cellular chromosomes, thereby affecting the
mainte-nance function of LANA1 (
28
). Genomes of herpesvirus saimiri
(HVS), a close relative of KSHV, are established inefficiently in
low-MeCP2-expressing NIH 3T3 cells, but this inefficiency is
res-cued upon ectopic expression of MeCP2 (
59
). Based on these
re-ports, it is plausible that the differential abilities of cell types to
FIG 8MRE plasmids with a spacer sequence between 2 MRE units are stable in the presence of FR. (A) Schematic representation of MRE-spacer and MRE-spacer/FR plasmids. DNA fromEnterobacteriaphagewas inserted between 2 MREs such that the center-to-center spacing between them becomes 801 bp. (B) Establishment efficiencies of the MRE-spacer plasmid in the presence or absence of FR in JSC-1 cells expressed as CFU. CFU was determined as a percentage of transfected cells resulting in drug-resistant colonies. The error bars represent standard deviations obtained from three independent experiments. A statistically significant (Pⱕ0.05) difference as determined by the Jonckheere-Terpstra test is indicated and marked with an asterisk. (C) Gardella gel from populations of G418-resistant JSC-1 cells 4 months after transfection with MRE-spacer plasmid with or without FR. All lanes are from the same gel. The two sample lanes for the⫹FR group represent separate populations of JSC-1 cells from two independent transfections. Ctrl (control), 0.3 to 0.5 ng of free parental plasmids; open arrowheads, bands obtained from CCC DNA of the free parental plasmid or plasmids from cells that are the same size as the parental plasmid; closed arrowhead, band obtained for CCC DNA of larger, recombined plasmids from cells; *, nicked forms of the plasmids in control and sample lanes when detected. The probe was p1782 (backbone of the plasmids).on November 7, 2019 by guest
http://jvi.asm.org/
[image:11.585.80.514.65.202.2]recombined and selected—is not limited to those occasions in
which LANA1 is expressed in
trans
.
Although the MRE was sufficient for establishment, plasmids
containing tandem repeats of the MRE behaved differently than
those containing repeats of the TR. First, the efficiency of synthesis
of MRE plasmids did not depend on the number of MREs per
plasmid (
Fig. 4
). Second, plasmids with 16 copies of the MRE,
unlike those with 16 copies of the TR, were also unstable, with
recombination producing larger species that were selected for
over time (
Fig. 5A
). Third, the MRE plasmids weren’t completely
rescued by the addition of FR in
cis
(
Fig. 7
). MRE/FR hybrid
plas-mids were established more efficiently than the MRE plasplas-mids,
but unlike TR/FR hybrid plasmids, they did recombine to yield
larger species that were selected after establishment. These
obser-vations indicate that in addition to being defective for
partition-ing, plasmids containing tandem repeats of MREs are also
defec-tive in synthesis.
We found that the defect in synthesis shown by the MRE
plas-mids can be attributed to the absence of 801-bp center-to-center
spacing between each unit of the MRE. Plasmids with a
non-KSHV spacer sequence between 2 units of MRE behaved similarly
to plasmids with 2 copies of the TR in that their inefficient
estab-lishment and instability were both rescued by the addition of FR in
cis
(
Fig. 8
). Thus, the actual sequence in the TR outside the MRE
region does not play a detectable role in the synthesis of KSHV
genomes, while an 801-bp center-to-center spacing between each
MRE does.
The spacing between each MRE could be important for the
preservation of the chromatin architecture of the TR.
EBNA1-and LANA1-dependent replication of EBV- EBNA1-and KSHV-derived
plasmids, respectively, is affected by nucleosome positioning and
remodeling at the vicinity of their replication origins. For yeast
autonomous replicating sequences (ARS), nucleosomes are
ex-cluded from sites in the ARS bound by its replicators (
60
). Forcing
the assembly of ARS into a nucleosome by deleting sequences
flanking the ARS reduces its ability to initiate replication (
60
,
61
),
highlighting the importance of both the native structure and the
chromatin organization of the origin. The TR of KSHV is also
organized into nucleosomes that are excluded from the LBS1/2
region in the context of an intact TR (
20
). Hence, it is plausible
that the 801-bp center-to-center spacing between 2 units of MREs
is required for proper chromatin structure and nucleosome
posi-tioning in the TR, which in turn might be required for efficient
synthesis through the TRs. Additionally, the process of
establish-ment is thought to include, as yet unknown, epigenetic changes in
cis
to the introduced plasmids (
16
,
35
). It is possible that the
plas-63
). Based on our findings, the persistence of these recombined
endogenous KSHV genomes indicates that they likely have a
se-lective advantage in terms of their genome maintenance in the
host cells harboring them.
ACKNOWLEDGMENTS
We thank Kenneth M. Kaye for kindly providing plasmids Z6-2TR and Z6-BE and Norman Drinkwater and members of the Sugden laboratory for helpful comments and critical readings of the manuscript.
This work was supported by grants from the National Cancer Insti-tute, National Institutes of Health (grants P01 CA022443, R01 CA133027, and R01 CA070723). Bill Sugden is an American Cancer Society Research Professor.
REFERENCES
1.Cesarman E, Moore PS, Rao PH, Inghirami G, Knowles DM, Chang Y.
1995. In vitro establishment and characterization of two acquired immu-nodeficiency syndrome-related lymphoma cell lines (BC-1 and BC-2) containing Kaposi’s sarcoma-associated herpesvirus-like (KSHV) DNA sequences. Blood86:2708 –2714.
2.Cesarman E, Chang Y, Moore PS, Said JW, Knowles DM.1995. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in AIDS-related body-cavity-based lymphomas. N. Engl. J. Med.332:1186 –1191.http://dx .doi.org/10.1056/NEJM199505043321802.
3.Soulier J, Grollet L, Oksenhendler E, Cacoub P, Cazals-Hatem D, Babinet P, d’Agay MF, Clauvel JP, Raphael M, Degos L.1995. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman’s disease. Blood86:1276 –1280.
4.Chang Y, Cesarman E, Pessin MS, Lee F, Culpepper J, Knowles DM, Moore PS. 1994. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science266:1865–1869.http://dx.doi .org/10.1126/science.7997879.
5.Zhong W, Wang H, Herndier B, Ganem D.1996. Restricted expression of Kaposi sarcoma-associated herpesvirus (human herpesvirus 8) genes in Kaposi sarcoma. Proc. Natl. Acad. Sci. U. S. A.93:6641– 6646.http://dx .doi.org/10.1073/pnas.93.13.6641.
6.Renne R, Lagunoff M, Zhong W, Ganem D.1996. The size and confor-mation of Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8) DNA in infected cells and virions. J. Virol.70:8151– 8154.
7.Dictor M, Rambech E, Way D, Witte M, Bendsöe N.1996. Human herpesvirus 8 (Kaposi’s sarcoma-associated herpesvirus) DNA in Kaposi’s sarcoma lesions, AIDS Kaposi’s sarcoma cell lines, endothelial Kaposi’s sarcoma simulators, and the skin of immunosuppressed patients. Am. J. Pathol.148:2009 –2016.
8.Kennedy G, Komano J, Sugden B.2003. Epstein-Barr virus provides a survival factor to Burkitt’s lymphomas. Proc. Natl. Acad. Sci. U. S. A.
100:14269 –14274.http://dx.doi.org/10.1073/pnas.2336099100. 9.Mack AA, Sugden B.2008. EBV is necessary for proliferation of dually
infected primary effusion lymphoma cells. Cancer Res.68:6963– 6968. http://dx.doi.org/10.1158/0008-5472.CAN-08-0627.
10. Vereide D, Sugden B.2009. Proof for EBV’s sustaining role in Burkitt’s lymphomas. Semin. Cancer Biol.19:389 –393.http://dx.doi.org/10.1016/j .semcancer.2009.07.006.
on November 7, 2019 by guest
11. Nishimura K, Ueda K, Sakakibara S, Ishikawa K, Chen J, Okuno T, Yamanishi K.2001. Functional analysis of Kaposi’s sarcoma-associated herpesvirus RTA in an RTA-depressed cell line. J. Hum. Virol.4:296 –305. 12. Ballestas ME, Kaye KM.2001. Kaposi’s sarcoma-associated herpesvirus latency-associated nuclear antigen 1 mediates episome persistence throughcis-acting terminal repeat (TR) sequence and specifically binds TR DNA. J. Virol.75:3250 –3258.http://dx.doi.org/10.1128/JVI.75.7.3250 -3258.2001.
13. Lim C, Seo T, Jung J, Choe J.2004. Identification of a virustrans-acting regulatory element on the latent DNA replication of Kaposi’s sarcoma-associated herpesvirus. J. Gen. Virol.85:843– 855.http://dx.doi.org/10 .1099/vir.0.19510-0.
14. Ye FC, Zhou FC, Yoo SM, Xie JP, Browning PJ, Gao SJ.2004. Disrup-tion of Kaposi’s sarcoma-associated herpesvirus latent nuclear antigen leads to abortive episome persistence. J. Virol.78:11121–11129.http://dx .doi.org/10.1128/JVI.78.20.11121-11129.2004.
15. Ballestas ME, Chatis PA, Kaye KM.1999. Efficient persistence of extra-chromosomal KSHV DNA mediated by latency-associated nuclear anti-gen. Science 284:641– 644. http://dx.doi.org/10.1126/science.284.5414 .641.
16. Grundhoff A, Ganem D.2003. The latency-associated nuclear antigen of Kaposi’s sarcoma-associated herpesvirus permits replication of terminal repeat-containing plasmids. J. Virol.77:2779 –2783.http://dx.doi.org/10 .1128/JVI.77.4.2779-2783.2003.
17. Hu J, Renne R.2005. Characterization of the minimal replicator of Ka-posi’s sarcoma-associated herpesvirus latent origin. J. Virol.79:2637– 2642.http://dx.doi.org/10.1128/JVI.79.4.2637-2642.2005.
18. Hu J, Garber AC, Renne R.2002. The latency-associated nuclear antigen of Kaposi’s sarcoma-associated herpesvirus supports latent DNA replica-tion in dividing cells. J. Virol.76:11677–11687.http://dx.doi.org/10.1128 /JVI.76.22.11677-11687.2002.
19. Lim C, Sohn H, Lee D, Gwack Y, Choe J.2002. Functional dissection of latency-associated nuclear antigen 1 of Kaposi’s sarcoma-associated her-pesvirus involved in latent DNA replication and transcription of terminal repeats of the viral genome. J. Virol.76:10320 –10331.http://dx.doi.org/10 .1128/JVI.76.20.10320-10331.2002.
20. Stedman W, Deng Z, Lu F, Lieberman PM.2004. ORC, MCM, and histone hyperacetylation at the Kaposi’s sarcoma-associated herpesvirus latent replication origin. J. Virol.78:12566 –12575.http://dx.doi.org/10 .1128/JVI.78.22.12566-12575.2004.
21. Verma SC, Choudhuri T, Kaul R, Robertson ES. 2006. Latency-associated nuclear antigen (LANA) of Kaposi’s sarcoma-Latency-associated her-pesvirus interacts with origin recognition complexes at the LANA binding sequence within the terminal repeats. J. Virol.80:2243–2256.http://dx.doi .org/10.1128/JVI.80.5.2243-2256.2006.
22. Verma SC, Choudhuri T, Robertson ES.2007. The minimal replicator element of the Kaposi’s sarcoma-associated herpesvirus terminal repeat supports replication in a semiconservative and cell-cycle-dependent man-ner. J. Virol.81:3402–3413.http://dx.doi.org/10.1128/JVI.01607-06. 23. Piolot T, Tramier M, Coppey M, Nicolas JC, Marechal V.2001. Close
but distinct regions of human herpesvirus 8 latency-associated nuclear antigen 1 are responsible for nuclear targeting and binding to human mitotic chromosomes. J. Virol.75:3948 –3959.http://dx.doi.org/10.1128 /JVI.75.8.3948-3959.2001.
24. Shinohara H, Fukushi M, Higuchi M, Oie M, Hoshi O, Ushiki T, Hayashi J, Fujii M.2002. Chromosome binding site of latency-associated nuclear antigen of Kaposi’s sarcoma-associated herpesvirus is essential for persistent episome maintenance and is functionally replaced by histone H1. J. Virol.76:12917–12924.http://dx.doi.org/10.1128/JVI.76.24.12917 -12924.2002