Structure of an HIV-2 gp120 in Complex with CD4
Yunji W. Davenport,*Anthony P. West, Jr., Pamela J. Bjorkman
Division of Biology and Biological Engineering, California Institute of Technology, Pasadena, California, USA
HIV-2 is a nonpandemic form of the virus causing AIDS, and the majority of HIV-2-infected patients exhibit long-term nonpro-gression. The HIV-1 and HIV-2 envelope glycoproteins, the sole targets of neutralizing antibodies, share 30 to 40% identity. As a first step in understanding the reduced pathogenicity of HIV-2, we solved a 3.0-Å structure of an HIV-2 gp120 bound to the host receptor CD4, which reveals structural similarity to HIV-1 gp120 despite divergence in amino acid sequence.
H
uman immunodeficiency virus type 1 (HIV-1) remains a threat to global public health. With no vaccine available, it dispropor-tionately causes AIDS in the developing world. While the related virus HIV-2 is capable of generating full-blown AIDS, infection rates are more localized and on the decline (1–4). In contrast to patients in-fected with HIV-1, the majority of HIV-2-inin-fected individuals re-main asymptomatic, with low plasma viral loads and normal CD4⫹T cell counts (5–7). HIV-2 is a robust immunogen in natural infec-tion, and patients readily make high-titer, broadly reactive, and strongly neutralizing antibodies against primary strains, some-thing rarely seen in HIV-1 infections (1–3). In addition, HIV-2 appears slower to evolve the neutralization-resistant variant strains commonly found in HIV-1 infections (3,8). The rate of mother-to-child transmission is also much lower for HIV-2 (9), and differential neutralizing activities found in patients dually in-fected with HIV-1 and HIV-2 suggest the presence of distinct and divergent immune responses against these related viruses (10).The bulk of current investigative efforts are focused on HIV-1 due to its higher virulence and transmissibility. However, the im-mune response in the context of HIV-2 infection may provide a natural model of effective HIV control. While HIV-2 is not an attenuated virus, infected patients seem to naturally exhibit char-acteristics of a desired response to an effective therapy against HIV. Therefore, a deeper understanding of the immune response against HIV-2 could provide insight into how to elicit a similar response against HIV-1. HIV-1 and HIV-2 share 30% overall se-quence identity and 40% amino acid identity in their envelope (Env) glycoprotein gp160 (1,2,11), which is the sole target of neutralizing antibodies. Through structural studies of broadly neutralizing antibodies against HIV-1, these antibodies have been shown to target a variety of epitopes on the gp120 and gp41 sub-units of HIV-1 gp160 (12) but do not generally cross-react be-tween HIV-1 and HIV-2 (2,13). Some HIV-1-induced CD4-in-duced (CD4i) antibodies, which bind to the coreceptor binding site exposed via a CD4-induced conformational change (14), cross-react to neutralize several HIV-2 strains (13), suggesting structural similarity in this portion of HIV-1 and HIV-2 Env. However, some HIV-2 isolates can enter cells in a CD4-indepen-dent manner (15), leading to the hypothesis that HIV-2 may have a more open coreceptor binding domain (3).
Insights gained from the HIV-2 model of elite viral control may inform prophylactic and immunogen design for both forms of HIV. However, to date, no three-dimensional (3D) structures of HIV-2 Env have been reported. To investigate potential explana-tions for its decreased pathogenicity and increased immunogenic-ity compared with HIV-1 Env, we solved a 3.0-Å crystal structure
of an HIV-2 gp120 core from the ST strain (gp120ST) (16) in complex with the first two domains of the host receptor CD4 (sCD4) and compared the structure to HIV-1 and SIV gp120 and CD4/gp120 structures.
A gene corresponding to HIV-2 gp120ST (Los Alamos HIV sequence database [http://www.hiv.lanl.gov]) was modified based on the HIV-1 gp120 core used for crystallographic studies (11,
17–19). Core gp120STcontained deletions of the V1/V2 and V3 variable loops, 15 residues of the C terminus (11), and a mutation (T89I) of a conserved potential N-linked glycosylation site (PNGS) distal from predicted binding interfaces (Fig. 1). Core gp120STand sCD4 were expressed and purified as previously de-scribed (20, 21) and then cocrystallized via sitting drop vapor diffusion in a 1:1 mixture with 0.15 MDL-malic acid 7.0, 0.1 M imidazole (pH 7.0), and 22% polyethylene glycol (PEG) MME 550. Crystals were cryopreserved in Fomblin Y (Sigma), and 3.0-Å diffraction data (as determined by CC1/2[22]) were collected at the SSRL beamline 12-2 and processed using XDS (23) (Table 1). Structure determination was performed by molecular replace-ment in Phaser (24) using structures of HIV-1 gp120CAP210(PDB code3LQA, chain G, 35% identical) and sCD4 D1D2 (2NXY, chain B, 100% identical) as search models. (Molecular replace-ment using the structure of an SIV gp120 [2BF1; 72% identical to gp120ST] as a search model yielded no solutions.) Manual rebuild-ing guided by simulated annealrebuild-ing composite omit maps calcu-lated using CNS (25) or Phenix.autosol (26) to minimize model bias (27) was performed using COOT (28). Iterative rounds of refinement were performed using Phenix.refine (26) and Refmac (29) within the PDB_REDO server (30), including but not limited to rigid body, group B factor, translation, libration, and screw-rotation (TLS), real-space, and Rosetta-guided refinement algo-rithms (31,32) (Table 1).
The HIV-2 core gp120STstructure (numbered according to the SIVmac239 convention [33]) comprises 23-strands (1 to23),
Received19 October 2015Accepted19 November 2015
Accepted manuscript posted online25 November 2015
CitationDavenport YW, West AP, Jr, Bjorkman PJ. 2016. Structure of an HIV-2 gp120 in complex with CD4. J Virol 90:2112–2118.doi:10.1128/JVI.02678-15.
Editor:G. Silvestri
Address correspondence to Pamela J. Bjorkman, [email protected].
*Present address: Yunji W. Davenport, Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, Boston, Massachusetts, USA.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
crossmark
on November 7, 2019 by guest
http://jvi.asm.org/
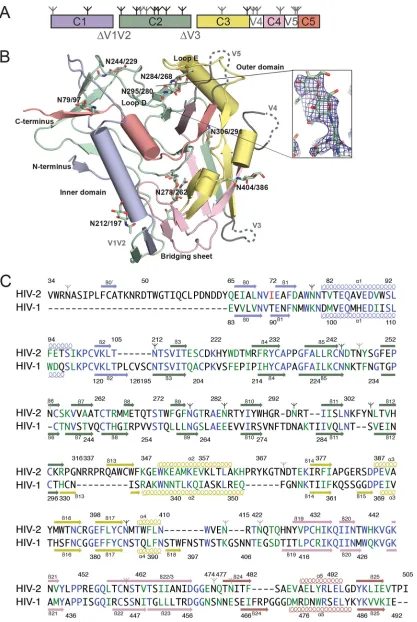
FIG 1Structure and sequence of HIV-2 gp120 core. (A) Diagram of the domain structure of the gp120 core, with locations of deleted loops indicated. Branch symbols mark potential glycosylation sites. (B) Structure of the HIV-2 gp120 core, with polypeptide chains as ribbons, alpha helices as cylinders, and carbohydrates as stick models. Disordered residues are shown as dashed lines. Domains are colored as in panel A. (Inset) Annealed omit electron density map in the vicinity of theN-glycan attached to N295gp120-STcontoured at 1.5. (C) Alignment of the gp120 core sequences of HIV-2 and HIV-1 HxB2. Residues in blue are identical; those in green are
conservative substitutions (scoring⬎0.5 in the Gonnet PAM 250 matrix [49]). The mutation to remove a PNGS in HIV-2 is in red. Colors of secondary structure elements correspond to colors in panels A and B. PNGS are marked as in panel A; sites with ordered sugars are indicated with black branch symbols, while sites with disordered sugars are indicated with gray branch symbols.
on November 7, 2019 by guest
[image:2.585.86.503.30.652.2]five␣-helices (␣1 to␣5), five loop regions (A to E), truncated variable loops V1 to V3, and disordered loops V4 and V5 (Fig. 1). In comparisons with available HIV-1 and SIV gp120 structures, we found that the gp120STstructure was more similar to gp120 structures from HIV-1 (PDB code2NXY[34],4P9H[20],3LQA
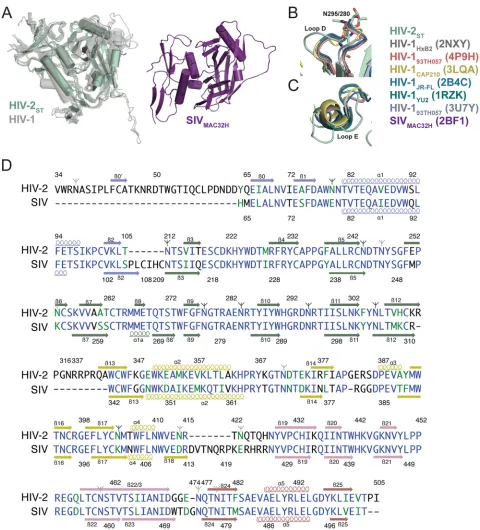
[21],2B4C[35],1RZK[36],3U7Y[37]) than to the structure of SIVmac32H gp120 (2BF1[19]), despite sharing only⬃35% se-quence identity with HIV-1 gp120s compared with 72% identity with SIVmac32H gp120 (11, 19) (Fig. 1Cand2D). Indeed, the overall architecture of HIV-2 gp120STwas nearly identical to both liganded and unliganded (PDB code3TGT [17]) HIV-1 gp120 cores, with secondary structure elements comprising the inner and outer domains adopting common conformations (Fig. 1). When superimposed with HIV-1 gp120 core structures using se-quence-based alignment in PyMOL (38), the core gp120ST struc-ture yielded root mean square deviation (RMSD) values of 1.3 to 1.6 Å (Fig. 2). In contrast, the HIV-2 gp120STstructure aligned poorly with the structure of unliganded SIVmac32H gp120 (19) (RMSD⫽5.6 Å for 230 C␣atoms), and the-strands comprising the bridging sheet were shifted by 13 to 25 Å, the V1/V2 stems were shifted by⬃27 Å, the␣1 helix was shifted by 15 Å, and the three-stranded-sheet at the C terminus was offset by a 90° rotation (Fig. 2A). We therefore used HIV-1 core gp120 structures for comparisons with HIV-2 gp120ST.
Analysis of structural and sequence conservation between gp120STcore and the seven most similar HIV-1 gp120 core struc-tures in the PDB (4DVR,3TGR,1G9N,3U7Y,4JM2,2NY0, and
1GC1) using ENDScript (39) showed high degrees of structural conservation in most ordered regions. Notably, the coreceptor binding footprint, which comprises the bridging sheet and sur-rounding residues, showed no major differences between gp120ST
and HIV-1 gp120s. In the HIV-2 gp120STstructure, six conserved disulfide bonds superimposed with their counterparts in HIV-1 gp120 core structures. Eight PNGSs showed ordered sugar resi-dues (Fig. 2). Only two (N262HXB2and N386HXB2) are also PNGSs in HIV-1 gp120s.
It has been postulated that HIV-2 has a more open V3 loop conformation than HIV-1 due to the HIV-2 V3 loop having fewer PNGSs (40). Core gp120SThas a truncated V3 stem with the same number of residues as the truncated stem in HIV-1 gp12093TH057 (3U7Y) and 10 more residues than the V3 stem in HIV-1 gp120HXB2 (2NXY). The ST and HIV-1 93TH057 V3 stems of equal length were superimposed with an RMSD of 0.46 Å. Thus, characterizing differences in this region between HIV-1 and HIV-2 would require future comparison of structures with intact and ordered V3 loops. The V4 and V5 loops, which are shorter and less glycosylated in HIV-2 than in HIV-1, were completely disor-dered in the gp120ST core structure. One of the most distinct structural differences between HIV-2 and HIV-1 gp120 is the ex-tended loop E of HIV-2 gp120, which emanates from the junction between the␣2 helix and14 strand in the gp120STcore structure. In alignments with loop E from HIV-1 gp120 (Fig. 2C), ST loop E was 3 to 5 residues longer and featured a turn not seen in HIV-1 gp120 structures.
Superposition of the gp120ST/sCD4 structure on three HIV-1 gp120/sCD4 structures revealed no notable global differences in the orientation of sCD4 binding to gp120ST(RMSD values of 2 to 2.1 Å after aligning 350 to 380 C␣atoms from gp120 and CD4) (Fig. 3). HIV-2 and HIV-1 gp120s share the same core interactions with CD4, represented on CD4 by residues 40 to 60, including the C⬙strand and residues F43CD4and R59CD4, and on gp120 by the CD4 binding loop, the F43CD4-binding pocket, and the D386gp120-ST/368gp120-HXB2(Fig. 4). However, gp120 loop D, which makes close contacts with sCD4, differed greatly between HIV-1 and HIV-2 (Los Alamos HIV sequence database [http://www.hiv.lanl.gov]). Differences included overall sequence, conformation, and glycosylation pattern. HIV-1 and HIV-2 gp120 residue 280gp120-HXB2/295gp120-STfeatures a highly conserved asparagine, which is the N-terminal residue of a PNGS in nearly all HIV-2 strains but never in HIV-1 strains (Los Alamos HIV sequence database [http://www.hiv.lanl .gov]). Instead, HIV-1 strains usually have a PNGS at position 276gp120-HXB2. The different loop D sequences are reflected in several differences in the gp120-sCD4 interaction. HIV-1 loop D and the pre-V523 strand interact with CD4 residues 29 to 35, but these interactions are absent in HIV-2. Conversely, HIV-2 gp120 has CD4 interactions that do not occur in HIV-1: loop D residue R293gp120-STinteracts with E87CD4and D88CD4. Also, the BC loop of CD4 (residues 19 to 25), which in other CD4 structures (11,20,
36) (including structures of CD4 alone [41–43]) is packed against the CD4 DE loop, is displaced⬃6 Å toward HIV-2 gp120, inter-acting with E84gp120-STand N79gp120-STnear the beginning of the ␣1 helix.
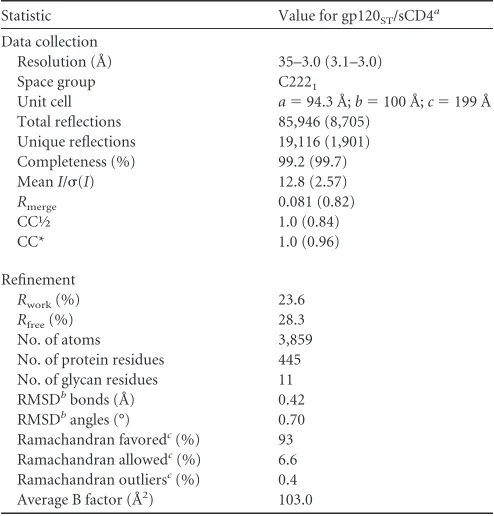
[image:3.585.40.287.77.335.2]Given the difference in pathogenicity between HIV-1 and HIV-2 (3) and the highly distinct unliganded SIV gp120 struc-ture (19), we expected larger differences between the gp120 structures of the two human viruses. Because the unexpected similarities between HIV-1 and HIV-2 gp120s do not explain the difference in virulence between HIV-1 and HIV-2 (1–4), the decreased virulence of HIV-2 may relate to structural dif-ference outside the gp120 core or to other difdif-ferences between TABLE 1Data collection and refinement statistics for gp120ST/sCD4
Statistic Value for gp120ST/sCD4a
Data collection
Resolution (Å) 35–3.0 (3.1–3.0) Space group C2221
Unit cell a⫽94.3 Å;b⫽100 Å;c⫽199 Å Total reflections 85,946 (8,705)
Unique reflections 19,116 (1,901) Completeness (%) 99.2 (99.7) MeanI/(I) 12.8 (2.57)
Rmerge 0.081 (0.82)
CC½ 1.0 (0.84)
CC* 1.0 (0.96)
Refinement
Rwork(%) 23.6
Rfree(%) 28.3
No. of atoms 3,859 No. of protein residues 445 No. of glycan residues 11 RMSDbbonds (Å) 0.42
RMSDbangles (°) 0.70
Ramachandran favoredc(%) 93
Ramachandran allowedc(%) 6.6
Ramachandran outliersc(%) 0.4
Average B factor (Å2) 103.0
a
Statistics for the highest-resolution shell are shown in parentheses. bRMSD, root mean square deviation.
c
Defined by MolProbity. Davenport et al.
on November 7, 2019 by guest
http://jvi.asm.org/
the two classes of virus. If decreased virulence is related at least in part to Env structure, differential immune activation could result from differences in the variable loops, which are trun-cated or disordered in gp120 core structures, and/or fewerN -linked glycans resulting in decreased glycan shielding of the
HIV-2 V3 and V4 loops (40). As demonstrated in X-ray and EM structures of trimeric HIV-1 Env (44–48), quaternary interac-tions outside the context of monomeric gp120 likely also play key roles in the behavior of HIV-2.
Due to the robust neutralizing antibody response to HIV-2 FIG 2Superposition of HIV-2, HIV-1, and SIV gp120 structures. (A) (Left) Sequence-based alignment of HIV-2STgp120 core (green) with liganded and
unliganded HIV-1 gp120 cores (gray) shows structural similarity. (Right) The SIVmac32H gp120 core from same viewpoint shows several secondary structural elements with different conformations. (B and C) Alignment of loops D and E from HIV-1 (various colors) and HIV-2 (mint green). (D) Alignment of the gp120 core sequences of HIV-2 and SIV. Residues in blue are identical; those in green are conservative substitutions (scoring⬎0.5 in the Gonnet PAM 250 matrix [49]). Colors of secondary structure elements correspond to colors inFig. 1A. PNGSs are marked as inFig. 1C; sites with ordered sugars are indicated with black branch symbols, while sites with disordered sugars are indicated with gray branch symbols.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.52.532.62.592.2](1–3), it is likely that differences in pathogenicity are due, at least in part, to the role of the Env glycoprotein. This is encouraging for the future growth of HIV-2 knowledge, as the HIV field has long been focused and is well equipped for conducting comprehensive
structural studies of the interactions between Env and its binding partners. Further X-ray crystallographic or EM studies involving neutralizing antibodies against HIV-2 Env should provide greater insight into this intriguing model of viral control.
FIG 3Conservation of the CD4 binding site. (A) Superposition of HIV-2STgp120 core in complex with sCD4 (green) with analogous HIV-1 cocrystal structures
(gray). (B and C) Polar contacts within the gp120ST/CD4 (left) and gp120HIV-1/CD4 (right) binding interface. (D) Superposition of CD4-binding loops from
HIV-2 and HIV-1 gp120 core/CD4 complex structures (colors are as inFig. 2B).
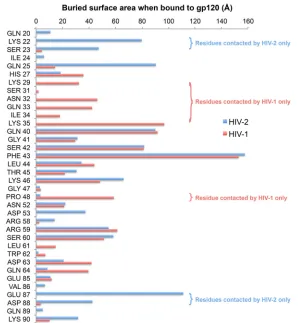
FIG 4Buried per-residue accessible surface area of CD4 upon binding to HIV-2 (blue) or HIV-1 (pink) gp120. Although much of the buried interface is shared, there are residues that are contacted by only HIV-1 or HIV-2 gp120. The CCP4 program AREAIMOL (mode DIFFMODE COMPARE) was used to calculate the per-residue difference in accessible surface area (1.4-Å probe radius) for CD4 upon complex formation with Env. HIV-1 coordinates were from PDB entry1RZJ. Davenport et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.152.439.65.250.2] [image:5.585.151.450.369.692.2]Protein structure accession number.Coordinates and X-ray crystallographic data for ST gp120/sCD4 have been deposited in the Protein Data Bank with accession number5CAY.
ACKNOWLEDGMENTS
We thank Jost Vielmetter and the Caltech Protein Expression Center for assistance with protein expression, Jens Kaiser, Pavle Nikolovski, the Caltech Molecular Observatory, and the staff at Beamline 12-2 (SSRL) for assistance with crystallographic data collection, the developers of PHENIX, CCP4, and XDS software, and Robbie Joosten of PDB_REDO for assistance and advice concerning X-ray crystallogra-phy.
The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
FUNDING INFORMATION
This work was supported by Collaboration for AIDS Vaccine Discovery grants from the Bill and Melinda Gates Foundation (grants 1040753 and 1124068 to P.J.B.) and by the National Institute of Allergy and Infectious Diseases, NIH, grant HIVRAD P01 AI100148 (P.J.B.). The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
REFERENCES
1.Kong R, Li H, Georgiev I, Changela A, Bibollet-Ruche F, Decker JM, Rowland-Jones SL, Jaye A, Guan Y, Lewis GK, Langedijk JP, Hahn BH, Kwong PD, Robinson JE, Shaw GM.2012. Epitope mapping of broadly neutralizing HIV-2 human monoclonal antibodies. J Virol86:12115– 12128.http://dx.doi.org/10.1128/JVI.01632-12.
2.Kong R, Li H, Bibollet-Ruche F, Decker JM, Zheng NN, Gottlieb GS, Kiviat NB, Sow PS, Georgiev I, Hahn BH, Kwong PD, Robinson JE, Shaw GM.2012. Broad and potent neutralizing antibody responses elic-ited in natural HIV-2 infection. J Virol86:947–960.http://dx.doi.org/10 .1128/JVI.06155-11.
3.de Silva TI, Cotten M, Rowland-Jones SL.2008. HIV-2: the forgotten AIDS virus. Trends Microbiol 16:588 –595.http://dx.doi.org/10.1016/j .tim.2008.09.003.
4.Hahn BH, Kong LI, Lee SW, Kumar P, Taylor ME, Arya SK, Shaw GM.1987. Relation of HTLV-4 to simian and human immunodefi-ciency-associated viruses. Nature 330:184 –186.http://dx.doi.org/10 .1038/330184a0.
5.van der Loeff MF, Larke N, Kaye S, Berry N, Ariyoshi K, Alabi A, van Tienen C, Leligdowicz A, Sarge-Njie R, da Silva Z, Jaye A, Ricard D, Vincent T, Jones SR, Aaby P, Jaffar S, Whittle H.2010. Undetectable plasma viral load predicts normal survival in HIV-2-infected people in a West African village. Retrovirology7:46.http://dx.doi.org/10.1186/1742 -4690-7-46.
6.Berry N, Ariyoshi K, Jaffar S, Sabally S, Corrah T, Tedder R, Whittle H. 1998. Low peripheral blood viral HIV-2 RNA in individuals with high CD4 percentage differentiates HIV-2 from HIV-1 infection. J Hum Virol 1:457– 468.
7.Jaffar S, Wilkins A, Ngom PT, Sabally S, Corrah T, Bangali JE, Rolfe M, Whittle HC.1997. Rate of decline of percentage CD4⫹cells is faster in HIV-1 than in HIV-2 infection. J Acquir Immune Defic Syndr Hum Retro-virol16:327–332.http://dx.doi.org/10.1097/00042560-199712150-00003. 8.MacNeil A, Sankale JL, Meloni ST, Sarr AD, Mboup S, Kanki P.2007.
Long-term intrapatient viral evolution during HIV-2 infection. J Infect Dis195:726 –733.http://dx.doi.org/10.1086/511308.
9.O’Donovan D, Ariyoshi K, Milligan P, Ota M, Yamuah L, Sarge-Njie R, Whittle H.2000. Maternal plasma viral RNA levels determine marked differences in mother-to-child transmission rates of HIV-1 and HIV-2 in The Gambia. MRC/Gambia Government/University College London Medical School working group on mother-child transmission of HIV. AIDS14:441– 448.
10. Schim van der Loeff MF, Jaffar S, Aveika AA, Sabally S, Corrah T, Harding E, Alabi A, Bayang A, Ariyoshi K, Whittle HC.2002. Mortality of HIV-1, HIV-2 and HIV-1/HIV-2 dually infected patients in a clinic-based cohort in The Gambia. AIDS16:1775–1783.http://dx.doi.org/10 .1097/00002030-200209060-00010.
11. Kwong PD, Wyatt R, Robinson J, Sweet RW, Sodroski J, Hendrickson WA.1998. Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature393: 648 – 659.http://dx.doi.org/10.1038/31405.
12. Kwong PD, Mascola JR.2012. Human antibodies that neutralize HIV-1: identification, structures, and B cell ontogenies. Immunity37:412– 425.
http://dx.doi.org/10.1016/j.immuni.2012.08.012.
13. Decker JM, Bibollet-Ruche F, Wei X, Wang S, Levy DN, Wang W, Delaporte E, Peeters M, Derdeyn CA, Allen S, Hunter E, Saag MS, Hoxie JA, Hahn BH, Kwong PD, Robinson JE, Shaw GM.2005. Anti-genic conservation and immunoAnti-genicity of the HIV coreceptor binding site. J Exp Med201:1407–1419.http://dx.doi.org/10.1084/jem.20042510. 14. DeVico AL.2007. CD4-induced epitopes in the HIV envelope glyco-protein, gp120. Curr HIV Res5:561–571. http://dx.doi.org/10.2174 /157016207782418560.
15. Thomas ER, Shotton C, Weiss RA, Clapham PR, McKnight A. 2003. CD4-dependent and CD4-independent HIV-2: consequences for neutraliza-tion. AIDS17:291–300.http://dx.doi.org/10.1097/00002030-200302140 -00002.
16. Kong LI, Lee SW, Kappes JC, Parkin JS, Decker D, Hoxie JA, Hahn BH, Shaw GM.1988. West African HIV-2-related human retrovirus with at-tenuated cytopathicity. Science240:1525–1529.http://dx.doi.org/10.1126 /science.3375832.
17. Kwon YD, Finzi A, Wu X, Dogo-Isonagie C, Lee LK, Moore LR, Schmidt SD, Stuckey J, Yang Y, Zhou T, Zhu J, Vicic DA, Debnath AK, Shapiro L, Bewley CA, Mascola JR, Sodroski JG, Kwong PD. 2012. Unliganded HIV-1 gp120 core structures assume the CD4-bound confor-mation with regulation by quaternary interactions and variable loops. Proc Natl Acad Sci U S A109:5663–5668.http://dx.doi.org/10.1073/pnas .1112391109.
18. Kong L, Wilson IA, Kwong PD.2015. Crystal structure of a fully glyco-sylated HIV-1 gp120 core reveals a stabilizing role for the glycan at Asn262. Proteins83:590 –596.http://dx.doi.org/10.1002/prot.24747. 19. Chen B, Vogan EM, Gong H, Skehel JJ, Wiley DC, Harrison SC.2005.
Structure of an unliganded simian immunodeficiency virus gp120 core. Nature433:834 – 841.http://dx.doi.org/10.1038/nature03327.
20. Scharf L, Scheid JF, Lee JH, West AP, Jr, Chen C, Gao H, Gnanapra-gasam PN, Mares R, Seaman MS, Ward AB, Nussenzweig MC, Bjork-man PJ.2014. Antibody 8ANC195 reveals a site of broad vulnerability on the HIV-1 envelope spike. Cell Rep7:785–795.http://dx.doi.org/10.1016 /j.celrep.2014.04.001.
21. Diskin R, Marcovecchio PM, Bjorkman PJ.2010. Structure of a clade C HIV-1 gp120 bound to CD4 and CD4-induced antibody reveals anti-CD4 polyreactivity. Nat Struct Mol Biol17:608 – 613.http://dx.doi.org/10.1038 /nsmb.1796.
22. Karplus PA, Diederichs K.2012. Linking crystallographic model and data quality. Science336:1030 –1033.http://dx.doi.org/10.1126/science .1218231.
23. Kabsch W.2010. XDS. Acta Crystallogr D Biol Crystallogr66:125–132.
http://dx.doi.org/10.1107/S0907444909047337.
24. McCoy AJ, Grosse-Kunstleve RW, Adams PD, Winn MD, Storoni LC, Read RJ.2007. Phaser crystallographic software. J Appl Crystallogr40: 658 – 674.http://dx.doi.org/10.1107/S0021889807021206.
25. Brunger AT.2007. Version 1.2 of the Crystallography and NMR system. Nat Protoc2:2728 –2733.http://dx.doi.org/10.1038/nprot.2007.406. 26. Adams PD, Afonine PV, Bunkoczi G, Chen VB, Davis IW, Echols N,
Headd JJ, Hung LW, Kapral GJ, Grosse-Kunstleve RW, McCoy AJ, Mo-riarty NW, Oeffner R, Read RJ, Richardson DC, Richardson JS, Terwilliger TC, Zwart PH.2010. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr D Biol Crystallogr66: 213–221.http://dx.doi.org/10.1107/S0907444909052925.
27. Brunger AT, Rice LM.1997. Crystallographic refinement by simulated annealing: methods and applications. Methods Enzymol277:243–269.
http://dx.doi.org/10.1016/S0076-6879(97)77015-0.
28. Emsley P, Lohkamp B, Scott WG, Cowtan K.2010. Features and devel-opment of Coot. Acta Crystallogr D Biol Crystallogr66:486 –501.http: //dx.doi.org/10.1107/S0907444910007493.
29. Murshudov GN, Vagin AA, Dodson EJ.1997. Refinement of macromolec-ular structures by the maximum-likelihood method. Acta Crystallogr D Biol Crystallogr53:240 –255.http://dx.doi.org/10.1107/S0907444996012255. 30. Joosten RP, Long F, Murshudov GN, Perrakis A.2014. The PDB_REDO
server for macromolecular structure model optimization. IUCrJ1:213– 220.http://dx.doi.org/10.1107/S2052252514009324.
on November 7, 2019 by guest
http://jvi.asm.org/
31. Adams PD, Baker D, Brunger AT, Das R, DiMaio F, Read RJ, Rich-ardson DC, RichRich-ardson JS, Terwilliger TC.2013. Advances, interactions, and future developments in the CNS, Phenix, and Rosetta structural biol-ogy software systems. Annu Rev Biophys42:265–287.http://dx.doi.org/10 .1146/annurev-biophys-083012-130253.
32. Afonine PV, Grosse-Kunstleve RW, Echols N, Headd JJ, Moriarty NW, Mustyakimov M, Terwilliger TC, Urzhumtsev A, Zwart PH, Adams PD. 2012. Towards automated crystallographic structure refinement with phe-nix.refine. Acta Crystallogr D Biol Crystallogr68:352–367.http://dx.doi .org/10.1107/S0907444912001308.
33. Calef C, Mokili J, O’Connor DH, Watkins DI, Korber B.2005. Num-bering Positions in SIV Relative to SIVMM239. Los Alamos National Lab-oratory, Los Alamos, NM, USA.
34. Zhou T, Xu L, Dey B, Hessell AJ, Van Ryk D, Xiang SH, Yang X, Zhang MY, Zwick MB, Arthos J, Burton DR, Dimitrov DS, Sodroski J, Wyatt R, Nabel GJ, Kwong PD. 2007. Structural definition of a conserved neutralization epitope on HIV-1 gp120. Nature445:732–737.http://dx .doi.org/10.1038/nature05580.
35. Huang CC, Tang M, Zhang MY, Majeed S, Montabana E, Stanfield RL, Dimitrov DS, Korber B, Sodroski J, Wilson IA, Wyatt R, Kwong PD. 2005. Structure of a V3-containing HIV-1 gp120 core. Science310:1025– 1028.http://dx.doi.org/10.1126/science.1118398.
36. Huang CC, Venturi M, Majeed S, Moore MJ, Phogat S, Zhang MY, Dimitrov DS, Hendrickson WA, Robinson J, Sodroski J, Wyatt R, Choe H, Farzan M, Kwong PD.2004. Structural basis of tyrosine sulfation and VH-gene usage in antibodies that recognize the HIV type 1 coreceptor-binding site on gp120. Proc Natl Acad Sci U S A101:2706 –2711.http://dx .doi.org/10.1073/pnas.0308527100.
37. Diskin R, Scheid JF, Marcovecchio PM, West AP, Jr, Klein F, Gao H, Gnanapragasam PN, Abadir A, Seaman MS, Nussenzweig MC, Bjork-man PJ.2011. Increasing the potency and breadth of an HIV antibody by using structure-based rational design. Science334:1289 –1293.http://dx .doi.org/10.1126/science.1213782.
38. Anonymous.The PyMOL molecular graphics system, version 1.7.4. Schrodinger, LLC, New York, NY.
39. Robert X, Gouet P.2014. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res42:W320 –W324.http: //dx.doi.org/10.1093/nar/gku316.
40. Shi Y, Brandin E, Vincic E, Jansson M, Blaxhult A, Gyllensten K, Moberg L, Brostrom C, Fenyo EM, Albert J.2005. Evolution of human immunodeficiency virus type 2 coreceptor usage, autologous neutraliza-tion, envelope sequence and glycosylation. J Gen Virol86:3385–3396.
http://dx.doi.org/10.1099/vir.0.81259-0.
41. Garrett TP, Wang J, Yan Y, Liu J, Harrison SC.1993. Refinement and analysis of the structure of the first two domains of human CD4. J Mol Biol 234:763–778.http://dx.doi.org/10.1006/jmbi.1993.1625.
42. Wang JH, Yan YW, Garrett TP, Liu JH, Rodgers DW, Garlick RL, Tarr GE, Husain Y, Reinherz EL, Harrison SC.1990. Atomic structure of a fragment of human CD4 containing two immunoglobulin-like domains. Nature348:411– 418.http://dx.doi.org/10.1038/348411a0.
43. Wu H, Kwong PD, Hendrickson WA.1997. Dimeric association and segmental variability in the structure of human CD4. Nature387:527–530.
http://dx.doi.org/10.1038/387527a0.
44. Do Kwon Y, Pancera M, Acharya P, Georgiev IS, Crooks ET, Gorman J, Joyce MG, Guttman M, Ma X, Narpala S, Soto C, Terry DS, Yang Y, Zhou T, Ahlsen G, Bailer RT, Chambers M, Chuang GY, Doria-Rose NA, Druz A, Hallen MA, Harned A, Kirys T, Louder MK, O’Dell S, Ofek G, Osawa K, Prabhakaran M, Sastry M, Stewart-Jones GB, Stuckey J, Thomas PV, Tittley T, Williams C, Zhang B, Zhao H, Zhou Z, Donald BR, Lee LK, Zolla-Pazner S, Baxa U, Schon A, Freire E, Shapiro L, Lee KK, Arthos J, Munro JB, Blanchard SC, Mothes W, Binley JM, et al. 2015. Crystal structure, conformational fixation and entry-related inter-actions of mature ligand-free HIV-1 Env. Nat Struct Mol Biol22:522–531.
http://dx.doi.org/10.1038/nsmb.3051.
45. Julien JP, Cupo A, Sok D, Stanfield RL, Lyumkis D, Deller MC, Klasse PJ, Burton DR, Sanders RW, Moore JP, Ward AB, Wilson IA.2013. Crystal structure of a soluble cleaved HIV-1 envelope trimer. Science342: 1477–1483.http://dx.doi.org/10.1126/science.1245625.
46. Lyumkis D, Julien JP, de Val N, Cupo A, Potter CS, Klasse PJ, Burton DR, Sanders RW, Moore JP, Carragher B, Wilson IA, Ward AB.2013. Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer. Sci-ence342:1484 –1490.http://dx.doi.org/10.1126/science.1245627.
47. Pancera M, Zhou T, Druz A, Georgiev IS, Soto C, Gorman J, Huang J, Acharya P, Chuang GY, Ofek G, Stewart-Jones GB, Stuckey J, Bailer RT, Joyce MG, Louder MK, Tumba N, Yang Y, Zhang B, Cohen MS, Haynes BF, Mascola JR, Morris L, Munro JB, Blanchard SC, Mothes W, Connors M, Kwong PD.2014. Structure and immune recognition of trimeric pre-fusion HIV-1 Env. Nature514:455– 461.http://dx.doi.org/10 .1038/nature13808.
48. Scharf L, Wang H, Gao H, Chen S, McDowall AW, Bjorkman PJ.2015. Broadly neutralizing antibody 8ANC195 recognizes closed and open states of HIV-1 Env. Cell162:1379 –1390.http://dx.doi.org/10.1016/j.cell.2015 .08.035.
49. Thompson JD, Gibson TJ, Higgins DG.2002. Multiple sequence align-ment using ClustalW and ClustalX. Curr Protoc Bioinformatics Chapter 2:Unit 2 3.
Davenport et al.