Comparison
of
the Structure
and Polypeptide
Composition
of
Three
Double-Stranded
Ribonucleic
Acid-Containing
Viruses
(Diplornaviruses):
Cytoplasmic
Polyhedrosis
Virus,
Wound
Tumor
Virus,
and
Reovirus
L. J. LEWANDOWSKI AND B. L. TRAYNOR
DepartmentofMedicalMicrobiology,Stanford UniversitySchooloJMedicinie,Staniford, Californzia 94305anid Departmentof MolecutlarBiology anid Virus Laboratory, UniiversityofCalifornia, Berkeley, California94720
Received for publication 17July1972
Iodinationofreovirus, cytoplasmic polyhedrosisvirus (CPV),and wound tumor
virus(WTV),and theirrespectivesubviralforms,followedby analysisof thelabeled
polypeptides by usingpolyacrylamide gel electrophoresis, has beenused tocompare
theprotein contentsof these three diplornaviruses. Thisapproach, whencombined
with electron microscopy and buoyant density determinations, appears capable of
localizing individual polypeptides in some ofthe viral and subviral forms. CPV
(p = 1.435 g/cm3) seems to resemble reovirus cores (p = 1.440 g/cm3) in both
ultrastructureand polypeptide composition. CPV iscomposedof five polypeptides
with molecular weights of about 151,000, 142,000, 130,000, 67,000, and 33,000.
The polyhedral matrix, which in nature encapsulates the virions, is, in turn,
com-posed mainly oftwo polypeptide species with molecular weights ofabout 30,000
and 20,000, and several minor proteins. The proteins of WTV consist mainly of
four species of polypeptide with molecular weights of about 156,000, 122,000,
63,000, and44,000, and several minor components. These molecular weight
deter-minationsareconsistentwith thehypothesis that,ashasbeensuggested for reovirus,
the viral proteins ofCPV and WTV seemto becoded for by monocistronic mes senger RNA molecules transcribed from distinct segments ofthe double-stranded
RNAviral genomes.
In previous communications it was observed thatthestructureofthenucleic acid of the insect pathogen,
cytoplasmic
polyhedrosis
virus (CPV),was similar to that ofthe mammalian reovirus,
inthatbothgenomeswerecomposed of10distinct
double-stranded ribonucleic acid (dsRNA)
seg-ments present in equimolar amounts within the respective virions (15, 20). Furthermore, these
viruses, as well as a third virus-containing
seg-mented dsRNA (12), the plant oncogenic virus,
namely, wound tumor virus (WTV), contain
enzymatic activitiesin their superstructures
capa-ble of transcribing their respective dsRNA
genomes into discrete corresponding
single-strandedRNAmessengermolecules (3, 4, 13, 22).
The
common
presence ofthese two properties, asegmenteddsRNA genomeandavirus-associated transcriptase, suggests that, despite their rather
diverse host systems, these dsRNA viruses (the
nomenclaturediplornaviruseshas
been
suggestedby Verwoerd [31]) share the samebasic mode of
replication.
In the present study we have continued the
structural comparison of reovirus, CPV, and
WTV with special emphasis on the number and
location of specific
polypeptides.
The combinedtechniques of electron microscopy and the in
vitro labeling ofproteins with 1251 (19) have been
used for this
purpose.
lodination
studies withinfluenza virus have recently been used to suggest
1053
on November 10, 2019 by guest
http://jvi.asm.org/
LEWANDOWSKI AND TRAYNOR
that the rate of such in vitro labeling of viral
polypeptides can be related to the position ofthe
individual polypeptide on the surface of the
virus(26).
MATERIALS AND METHODS
Chemicals. Sodium 125I (250 mCi/ml) in 0.1 M NaOH was obtained from New England Nuclear Corp., Boston, Mass. Chloramine-T and sodium-m-bisulfite were obtained from Eastman Kodak Co., Rochester, N.Y. Sodium dodecyl sulfate (SDS) was from Sigma Chemical Co., St. Louis, Mo. a-Chymo-trypsin was obtained from Worthington Biochemical Corp., Freehold, N.J. Electrophoresis grade acryl-amide, methylene bisacrylamide, ammonium persul-fate, and N,N,N',N'-tetramethylethylene diamine
wereobtained from BioRadLaboratories, Richmond,
Calif. Allotherchemicals werereagentgrade. Preparation ofviruses.ReovirustypeIIIwasgrown
in Lcells and purified by the method of Watanabe
et al. (33). Reovirus cores were prepared following
theprocedureof Smithetal. (25). CPV was grown in Bombyx mori,and bothpolyhedralbodies and virions
were purified as described in an earlier report (13).
WTV-infected tumor root tissue from sweet clover
(Melilatus alba) and virus-infected plant cuttings for vegetativepropagation werekindly providedbyL. M.
Black. Virus was extracted and purified as described by Kimura (3) with threeexceptions. Exposure of the tumor tissue to carbon tetrachloride waseliminated,
virus was concentrated by sedimentationonto a65%
sucrose-D20 cushionin placeof pelleting, and virus
waspurified through twocycles on 30 to60%j>o linear
sucrosegradients.
Equilibrium density centrifugation was performed
using preformed gradients of CsCl instandard buffer
and ranging in densities from 1.453 to 1.25 g/cm3.
Sedimentation was at 36,000 rev/min in an SW50.1 rotorfor 9 hrat4 C.Fractionswerecollected and
re-fractive indicesweredeterminedon aZeiss refractrom-eter. Buoyant density values were calculated accord-ing to theformula, pCsCl = 10.860nd25C - 13.4974
(32).
Electron microscopy. Samplesfornegative staining
weredialyzed against 0.01 Mammonium acetate, pH
7.0, mixed with an equal volume of 2%/o
phospho-tungstic acid (PTA) (pH 7.0), and sprayed with a
nebulizerontoaplatinum,7-hole Siemensgridwhich
hadacarbon-coveredcollodionfilm. Forshadowing,
the samples were sprayed on collodium-covered,
200-meshcoppergrids, air dried, and uranium shad-owed. Forthin-sectioning, samples werefixed in 2%7 glutaraldehyde, postfixed in osmium, embedded in epoxy, and sectioned with the LKB ultratome. Sec-tionswere poststained, first with 20% uranyl acetate inmethanol for20min, followedbylead citrate for5
min, and examined inaSiemens Elmiskop Ielectron microscope.
Dissociation of virus. Samples were dissociated eitherpriorto,orafter,iodination.Intheformercase,
CPV and WTV samples in buffer were dissociated with
17%
SDS at 100 C for 2 min, whereas reoviruswas dissociated with
I1,%c
SDSat 100 Cfor 2min, orwith 0.2% SDS and 6 M urea at 45 C for 15min. For dissociation of iodinated CPV and WTV, samples were exposed to 1 % SDS, 6 M urea, 0.03 M dithio-threitol (DTT), 0.1 M tris(hydroxymethyl)amino-methane (Tris), pH 7.5, at 100 C for 2min.lodinated reovirus was dissociated in 0.6% SDS, 6 M
urea,
0.007 M DTT, and 0.007 M phosphate buffer, pH 7.0, at 37 C for 1 hr. Unlabeled CPV used for molecular weight determinations in Fig. 8 was disrupted in 1% SDS, 0.014 M DTT, and 0.006 M phosphate buffer, by heating at 45 Cfor 20min.
In vitro iodination. The tyrosine residues of the samples under study were iodinated by using chlor-amine-T as the oxidizing agent. The reaction mixture consistedof 50,uliters of virus or subviral component, 35 ,uliters of 0.14 M Tris-hydrochloride, pH 7.5, and 5 /Aliters ofNa'251 in 0.2 M Tris, pH 7.5 (200
,uCi/
sample). Labeling was initiated by addition of 20
,uliters ofchloramine-T (10mg/ml) and was allowed
tocontinue at room temperature for the various times listed in the figure legends. The reaction was termi-nated by addition of 20,liters ofsodium-m-bisulfite (24mg/ml) and 1 ,uliter of 0.01 M potassium iodide. Excess 1251 was removed by overnight dialysis with several changes of buffer (0.01 M Tris-hydrochloride, pH 7.5, 0.001 M ethylenediaminetetraacetic acid [EDTA], 0.1 M NaCl for intact virus, and 0.01 M Tris-hydrochloride, pH 7.5, 0.001 M EDTA, 0.1% SDS for dissociated virus). When polyhedral bodies
wereiodinated, excess 125I was removed by
sedimenta-tionthrough 30% sucrose in an SW65 rotor at 10,000 rev/min for 15 min at 4 C. The pellet was resuspended in 5 ml of buffer (0.1 M Tris-hydrochloride, pH 7.5, 0.001 M EDTA, 0.1 M NaCI) and resedimented in a Sorvallrefrigerated centrifuge at 10,000 rev/min for
10 min. This final pellet of 1251-labeled polyhedral bodieswassolubilized with 1% SDS-8 Murea at100C for 2 minpriortoelectrophoresis.
Polyacrylamide gel elfctrophoresis. Dissociated viral fractions were made 5% in sucrose, 0.005% in
bromophenol blue dye, and analyzed on 5%
poly-acrylamide gels (120 mm inlength) prepared in 0.1 M sodium phosphate buffer, pH 7.2, 10%glycerol, 0.13%
bisacrylamide,0.02 MEDTA,0.1%SDS,0.1%temed
(N, N, N',N'-tetramethylethylenediamine),0.08% per-sulfate. Electrophoresis buffer consisted of 0.1 M sodiumphosphate, pH 7.2, 0.1%SDS, 0.02 M EDTA.
Electrophoresis was usually doneat6mapergelfor
25hr oruntilthedyefrontwas nearthebottomof the
gel.Fordeterminationof1251content,gelswerefrozen
in ethanol-dry ice and fractionated by a mechanical
gel fractionater into 1-mm slices. The gel fractions
weredried and countedinatoluene basescintillation fluid (4 g of2,5-diphenyloxazole and 50mg of
1,4-bis-2-(5-phenyloxazolyl)benzene per liter of toluene)
in an Intertechnique liquid scintillation counter at
full window. Molecularweightdetermination of
poly-peptideswasdonefollowingthegeneral procedureof
Weber and Osborn (35). Polypeptide staining was performed with Coomassie brilliant blue (0.25%)
in amixtureof 50%methanol-10%glacialacetic acid for 3 hr at roomtemperature. Gels were allowed to remain in destaining solution (7.5% glacial acetic
1054 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
acid-5%methanol) for1 hratroomtemperatureprior
to electrophoreticdestaining inthesamesolution.
RESULTS
Reovirus:
efficacy
ofiodination as a means of localizing virus polypeptides in relation to virus surface. Studies with reovirus type III(25)
have shownthat reoviralpolypeptides
canbe isolated from three sources: wholevirions,
viral"top
component"
(RNA-free
viralshells),
and sub-viral core particles obtainedby
enzymatic
re-moval
of
the outercoatpolypeptides.
This samereport has shownthat reovirus
polypeptides
aredistributed in three
classes,
X(containing
twocomponents, X1 and
X2),
,u(containing
twocom-ponents,
1,u
andA2),
and o-(containing
threecomponents,
o,,
O2,anda3).
Becauseit isalready
knownwhichofthereovirustypeIII
polypeptides
composethe externalcoatand which the internal
core proteins
(25,
16), this system seemed toprovide an excellent control to ourgeneral
pro-posal
to examine theposition
ofdiplornavirus
polypeptides
by
using
the chloramine-T iodina-tiontechnique.
Equilibrium sedimentation and electron
micros-copy of reovirus. When a crude
preparation
of reovirus typeIII was sedimented toequilibrium
in preformed
gradients of
CsCl,
thesample
separated
into
threemajor
areas oflight-scatter-ingparticles, adense
major
band,
two less densesatellite bands, and a third series of even less
dense minor bands. These three areas were
col-lected and repurified on separate gradients of
CsCl. Based on buoyant density determinations
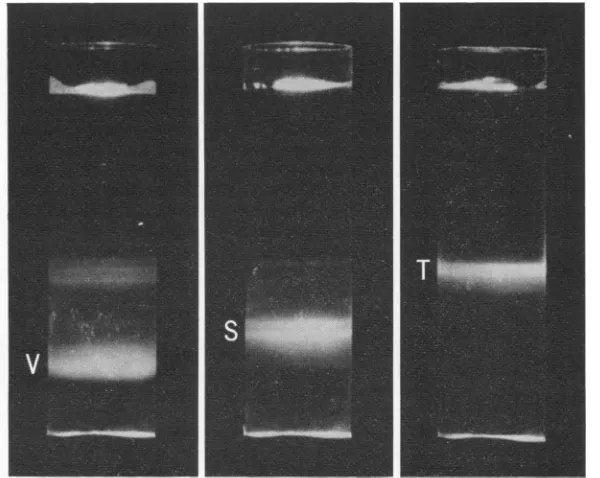
of 1.38, 1.36, and 1.30 g/cm3, the respective bands seen in Fig. 1 were arbitrarily designated
V, S, andTtorepresent virions, satellite virions,
and top component. Representative samples of
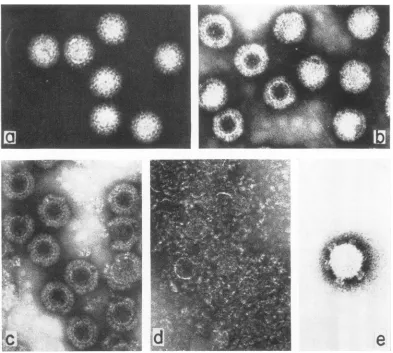
these three preparations were then examined in the electron microscope. Figure 2 shows that,
although both populations Sand V are approxi-mately 75 nm in diameter and morphologically similar icosahedrons, the satellite virions (p =
1.36 g/cm3, Fig. 2a) have uniformly allowed little penetration of stain, whereas the major
virion fraction (p = 1.38 g/cm3, Fig. 2b) shows
heavypenetration by PTA. Top component
ma-terial (p = 1.30 g/cm3, Fig. 2c) showed empty
intactshells which werecompletely permeableto PTA and composed primarily of an inner shell membrane and a relatively intact outer capsid
structure.Upon storage at 4 C, these shells tended to degrade into intact shell membranes and
random aggregates of capsomeres (Fig. 2d),
while both populations S and V appeared to
remainintact.
Iodination and gel electrophoresis ofthe
poly-peptides of reovirus and its subviral particles. Samples of these three populations were then
dissociated with urea-SDS, labeled with 1251,
FIG. 1. Sedimentation ofreovirus subfractions on apreformedgradient ofCsCl centrifugedfor 9 hr in an
SW50.1 rotor at36,000 revlmin. V, Virion (p = 1.38g/cm3;S, satellite virion (p = 1.36g/cm3); T,top com-ponent (p = 1.30g/'cm3).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.96.396.394.633.2]LEWANDOWSKI AND TRAYNOR
FIG. 2. Electron micrographs ofreovirussatellite (a),reovirus virion (b), top component (c), topcomponent stored at4C(d),andreovirus core(e). Particlesshown in a-d were isolatedfrom bands ofCsClequilibrium density gradients as shown in Fig. 1,dialyzedversus0.01Mammoniumacetate, pH 7.0,negatively stained with 1% PTA
atpH7.0,andsprayedwith anebulizeronto a7-hole, platinumSiemensgridwhich had acarbon-coated collodion
film. Particles shown in(e)weretreatedin0.01 Mammoniumacetate,pH 7.0, with 100 ,ug ofa-chymotrypsin/ml
at37Cfor60minprior tonegative staining.a-d, X160,000;e, X260,000.
and
subjected
topolyacrylamide gel
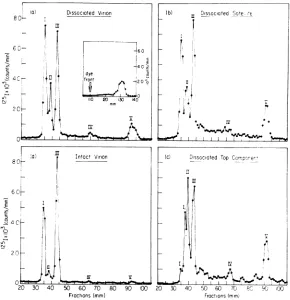
electrophore-sis. Figure 3a shows that the three
major
classes of viralpolypeptides, X,
A,
ando,
and theminorpeptide component VIII aresusceptibleto
iodina-tion. Thedistribution of 1251 label is 21, 38, 36, and 4%,
respectively,
and agreesfavorably
withthe values of24, 40, 32, and4% obtained when
virus is labeled in vivo with either 14C or 3H
amino acids (25). This distribution pattern
re-mainsconstant within thosetime periods tested,
from 1 to 15 min. Though not diagrammed, no
major differences could be detected in the
poly-peptide compositions of virion, satellite, or top
components. This
finding, together
with theinability to detect any gross differences in total
nucleic acid content betweenthetwopopulations
Vand S (Lewandowski and Traynor, unpublished observations),suggeststhat the
increased
buoyant density and susceptibility toPTA uptake of the majority population may be due to looseningratherthanremovalofsurfacepolypeptides.Such
arealignment would explain why populations of particles withthe same
RNA-protein
ratio exhibit differential uptake andbinding
ofsmall molecules likeCsCl(21),and PTA.Previous results suggested to us a potential relationship between reovirus core particles and purified CPV. Reovirus cores and CPV possess
similar segmented dsRNA genomes of
approxi-mately 15 x 106 daltons (14), similar buoyant
densities inCsCl, 1.435 4 0.005 g/cm3 (25, 15), and similar transcriptase activities demonstrable
1056 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
Froctions (mm)
3-0
(c)
6.0 _ 1
to~~~~~~~~~~~~~~~~~~~~~~~I
3) 40 50 60 70 80 90 100 110
Froctions (rr,rr)
in vitro (4, 13, 22, 24). Accordingly, intact
reo-virus was digested with
chymotrypsin
and sedimented toequilibrium
inCsCl. Twonucleo-protein core components resulted, a major one
at thedensity of1.44g/cm3
(Fig. 2e)
andaminorsatellite component at thedensity of 1.42g/cm3.
Littledifference couldbeobservedbetweenthese
two populations by electron
microscopy.
The most striking physical feature of reoviruscores isthe presence of regularly
spaced,
chimney-like
spike projections (Fig. 2e; reference 18) on the 12 fivefold vertices of the icosahedron. The two core populations were disrupted with urea-SDS, labeledwith 1251, andanalyzed for their
polypep-tidecontent by
polyacrylamide
gelelectrophore-sis. The superimposed patterns are presented in
Fig. 3b. In agreement with the literature data
(16, 18), reovirus core consists mainly ofthe X polypeptides, and polypeptide 02, and a minor
amount of ul. Satellite core, however, has
addi-tional labeled material migrating between the ,u
and a- peaks, and as
low-molecular-weight
pep-tide. Satellite core apparently represents an
incompletestageofremovalofthediscrete
degra-dation products X, Y, and Z (37), and small
peptides of the major coat polypeptides IA and
03 thereby
causing
the core particles to exhibita decreased buoyant
density.
Localization ofreovirus polypeptides. We next
investigated whether one could use the 1251 in vitro labeling procedure to
successfully
dis-criminate between surface and internal polypep-tides in the intact virion by the rate of labeling of the tyrosines of each class of polypeptide.Intact reovirus type III was treated with 1251
reagent for 60 sec; the subsequently obtained
electropherograms of the dissociated proteins
are shown in Fig. 3c. Two striking features are
observed: first, the X core polypeptides contain
very little label, about
7%
of total comparedwiththe 21
¼o
of totalwhendissociated
viruswaslabeled with iodine (Fig. 3a), and second, the
external coatpolypeptides ofthe ,u and
a-
classes areappreciably
labeled, and, in particular,o-has a disproportionate amount of label
(58%-)
compared to the amount observed
(36%)
whendissociated virus is iodinated. Thus, limiting the
[image:5.493.44.233.51.570.2]iodination time appears to discriminate between
FIG. 3. a, Polyacrylamide gel electrophoresis of reoviruisdissociatedwith6 -ifurea-0.2%'SDSat45C for15 mini priortoiodinatioln. Two huundred,lAiters of sample wasiodiniatedJor 15 miniat room temperature
asdescribedin Materialsacid Methods. The iodinated
sample was analyzed oln a 5c% polyacrylamide gel.
Migration wasfor6 hrat 3 mAper tube, anid 1-mm
sliceswere exami,iedfortheirradioactivecontent. Di-rection ofmigration isfrom le,ft to riglht. b, Reovirus
coreparticle (A) aiid satellite core (-), dissociated
with5 mi irea-0.2%,G SDSat45 C for45 miii prior to
iodination. lodination was 5miii atroom temperature.
Tlhe iodiniatedsamples wereplacedoii 4%
polyacryla-midegels aiid run,fbr 6hrat3mApertube.c,Reovirus iodinated prior to dissociationi with 6 -i urea-0.2%
SDSat100Cfbr2 min.Onte huintdred ,uliters of'sample
wasiodinated for60sec asdescribedin Materials alid Methods. Electrophoresis was oii 5% polyacrylamide
gels for27hrat7 mApertutbe.
0
c~j
0
re)
c'
50 60
Froctions (mm)
0
LI'
2
on November 10, 2019 by guest
http://jvi.asm.org/
LEWANDOWSKI AND TRAYNOR
polypeptidesin the undenatured virion. Not only
is thetechnique capable of accurately confirming
which classes of viral peptides comprise the
ex-terior coat, but also which particular class (in
this case
a)
is most accessible to iodination andpresumably, therefore, most exterior. These in
vitro dataare in good agreement with the recent
data
showing that,
invivo,
U is the firstprotein
toberemoved during the uncoating process (23)
followed
byg2.
Whenintact reovirus cores wereiodinatedprior to SDS dissociation (diagram not
shown)core polypeptideo inparticular showed a
lower percentage of total label relative to the X
components than was seen when cores were
dis-sociated prior to labeling (Fig. 3b). Thus, in the
intact reovirus core, the tyrosines of the X poly-peptides appear to be more available for
iodina-tion than are those of the ,u and a- core
poly-peptides.
CPV: electronmicroscopyofpolyhedral bodies.
CPV, perhapsthe mostwidespreadviral pathogen
of insects (1), derives itsname from light
micros-copy observations
W*'
that infection is associated/. ' 4'
..
....s
T.w..,. _..t1
b.]ili&
with the formation of unique cytoplasmic
struc-tureswhich are designated "polyhedral inclusion
bodies." Inasmuch as polyhedral bodies of
cer-tainshapes(icosahedral, hexahedral, and
pyrami-dal [10]) can becloned to give riseto polyhedra
of thatparticular shapeinsubsequent infections, polyhedral shapesappear tobecontrolled by the
strain of viruswhichthepolyhedracontain.
Thin-section electron microscopy was
per-formed on hexahedral polyhedral bodies
pro-ducedinsilkworm (Bombyxmori) larvae. Figure
4a showsthe gross structure of nativevirions as theyexistintheocclusion bodypriorto dissocia-tion of the polyhedral matrix. At the edges of
these polyhedra one can observe (arrows, Fig.
4a and 4b) native virions as they are partially
embedded orjust being released from thehighly
crystallized polyhedral protein matrix. The
ex-truding virion appears to have adensecore area
surrounded by an outer shell membrane which
has discernible spikes but no thick outer
capsid
structureanalogoustothereoviruscoat.
Equilibrium sedimentationand electron
micros-./
FIC;. 4.
Tlhin-sectioni
electronmicrograph ofCPVpolyhedralbodiesandvirus. Polyhedralbodieswerepreparedasdescribedin MaterialsandMethodsandfixedin2%glutaraldehyde,postfixedinosmium,embedded inepoxy,
and sectioned with the LKB ultratome. Sections werestained with20%0 utranylacetate in methanolfor20min,
poststainedinleadcitratejor5mini,andexaminedin aSiemensElmiskopIelectronmicroscope.a, X84,000; b,
X390,000.
1058 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
copyofpurified CPV. To observe the fine
struc-ture of CPV,
purified polyhedral bodies
wereexposed to pH
conditions known
tosolubilize
the polyhedral matrix without destroying the
infectivity of the released virions (13). Density
gradient centrifugation of such concentrated
ex-tracts onpreformed
gradients
of CsClproduced
three detectable light-scattering components at
equilibrium. A typical light-scattering pattern (Fig. 5) shows the
major
virus band(V)
atp =1.435 g/cm3, a minor diffuse shoulder area
(S)
at1.425g/cm3,andavery
sharp
minortopband
(T) at p = 1.30
g/cm3.
Previous work
in ourlaboratory (15) has shown that the
major band
(V) iscomposed of
virions
containing
the entiregenome complement
of
10 dsRNA segmentsin
equimolaramounts,whereas the diffuse shoulder
area contains
satellite virions
(S) which
are de-ficient insomeof thesmallerRNAsegments.By analogy toreovirus,
the RNA-free material equilibratingat p = 1.30 g/cm3 was designatedTfortopcomponent.
When
analyzed by
electronmicroscope,
parti-clesfrom themajor virus bandwerebynomeans
homogeneous with respect to their response to PTA
staining.
Thisvariation
inuptake
ofstain
was at times so
striking
thatwe selected severalmicrographs
(Fig.
6) whichhighlight
different structuralfeatures of CPV. Virionsappear to beicosahedrons
with an average diameterof
ap-proximately 65
nm,
acapsomeric-like
surfacestructure,thesuggestion ofanearly six-sided
out-line with 20 capsomeres often visible
along
the circumference, and distinctpyramidal spikes
onthecorners of the
polygon
(Fig.
6aand6b).
These pyramidal spikes which seem to originate deep within the central core area of the virus areparticularly
prominent
in thoseparticles
whichare
heavily internally
stained and aredistinct
from the
chimney-like
spikes
seen with reoviruscores
(Fig. 2e). Though six
spikes
arereadily
discernible,
one couldpredict,
basedon theob-servedstructureofthe
virion,
atotal of either 10,or, more
probably,
12(9). Frequently,
virions (Fig. 6c) seem to be extruding long, tail-likestructures. Due totheir6to 7nm diameter, itis
unlikely
thatthesetailsaresingle RNA duplexes which would measureapproximately
2 nm.More likely, these structures represent duplex
RNA bundles or
nucleoprotein
structuressome-what like thoseof
eucaryotic
chromosomefibers(6).Whenvirus samples aregiven minimum
elec-tron beam exposure (36), one can observe the
capsomeric-like surface structures but not the
spikestructuresingreaterdetail (Fig. 6d). Ingeneral,nodifference was observed between
the structure of virions banding at 1.435
g/cm'
and particles in the less dense, diffuse satellite
FIG. 5. Equilibrium sedimentation of CPV on a preformed gradientofCsCl (p = 1.370 to1.453g/cm3)
for 9 hr at 36,000
rev/min
in an SW50.1 rotor. V,Virion(p = 1.435g/cm3);S,satellitevirion(p = 1.425
g/cm3); T, top component (p = 1.30g/cm3).
zone, except thatsatellitevirionsmorefrequently
show tail-likestructures protruding from virions.
Exposure of CPV to chymotrypsin under the
sameconditionsthatproduce reovirus cores (25)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.493.256.438.74.553.2]LEWANDOWSKI AND TRAYNOR
FIG. 6. Electront micrographs showinig various morphological aspects of CPV isolated from CsCI
denisity
gradientsasshown inFig. 5. a, Virions withnegativestaininig;b,shadowed virioni;c, viriontextrudingnucleic acidcontaining tails; d,negatively stainied virionis givenminiimum electron beamexposure;antde,niegativelystainedtop
component.a, X215,000; b, X160,000; c, X95,000;danide, X320,000.
from whole reovirus showed no gross effect on
either CPV ultrastructure or buoyant density.
Electron microscopeexamination ofthematerial
banding atadensity of 1.30 g/cm3shows empty
virions completely permeable to PITA (Fig. 6e) and, as was true of the RNA-free uppermost
reovirion band, aggregates of partially
dissoci-ated viral shells.
Polypeptide composition of CPV and its
sub-viral forms.The
polypeptide
componentsofCPVwere investigated by the same techniques
em-ployed in model experiments with reovirus. Samples of the three viral components, virions, satellite area, and top component, were first
dissociated with urea-SDS, labeledwith
1251,
andsubjectedtoanalysison
polyacrylamide
gels.The1060 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.493.62.450.79.520.2]results are seenin theelectropherograms of
Fig.
7.
Disrupted
whole virions(Fig. 7a)
were foundtocontain five distinct iodine-labeled
polypeptides
and about 10% of total 1251 migrating as small peptide material in front of the dye marker (insert). The proportionof total radioactivity in
each of the viral
polypeptides
obtained after iodination of dissociated whole virus was: I,36%; II, 16%; III,
34%;
IV, 2%; and V, 10%,respectively.
When dissociated satellite and dissociated top
componentof CPVweresubsequently labeled in
vitro and analyzed on parallel
polyacrylamide
gels, the
polypeptide
patterns shown inFig.
7band7c wereobserved. Attentionmight be directed
to the occurrence of
polypeptide
breakdown asindicated by the presence of
high
background labelmigrating
between components III and V(Fig. 7b). This breakdown phenomenon is also
apparentin the analysis ofUdissociated top
com-ponent
(Fig. 7c).
Becausethe sizedistribution of
6
0-C E
>:e
0
n.-.
20
50 60 70 Froctions (mm)
specificbreakdownproducts of the
larger
species
might well overlap several
of
thenativespecies
in such a multicomponent system, it isgenerally
rather difficult to relate breakdown products to
a particular native component. The
exception,
however, seems to be breakdown
product
ILwhich, because of its size, can only be derived
from component I. The release of thedsRNA of
CPV, resulting first in a satellite area deficient inaportion ofthe RNA (15) and subsequently in
the fraction
called
top componentwhich,
by virtue of its low buoyantdensity,
isapparently
devoid ofanyof theRNA genome, seems
there-fore to be accompanied by the breakdown
particularly of the largest
polypeptide
species,
component I.
Localization of CPV polypeptides. The rate at
which thepolypeptidesofintact CPV virions are
iodinatedwasdetermined in the same manner as
with reovirus. The electrophoretic pattern of
labeled polypeptides obtained when
CsCl-puri-Froctions(mm)
FIG. 7. Electropherograms ofiodinated polypeptides of CPV virion (V), satellitevirion (S),andtopcomponent
(T) isolatedfrompreformnedgradients ofCsCl(Fig. 5).Samplesin a to c were dialyzedagainst 0.1 M
Tris-hydro-chloride(pH7.5)-0.001 M EDTA-0.1mNaCl,dissociated with 1% SDS, andiodinatedfor 60 sec asdescribed in
Materials and Methods.Thesampleindwasiodinatedfor 60 sec prior todissociation. Electrophoresis was on 5%
polyacrylamide gelsfor 25 hr at 6 mA per tube. The insert in a was included to demonstrate thepresence of small
peptide material whichmigrates ahead of the dye marker.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.493.98.388.307.607.2]LEWANDOWSKI AND TRAYNOR
fiedwhole viruswasiodinatedfor a briefperiod
prior to dissociation is shown in Fig. 7d.
Com-ponent III has a higher portion (56%), whereas
components II, IV, and V have lower portions (5%, 1%, and 2%, respectively) oftotal label
than when whole virus is dissociated prior to
iodination
(Fig. 7a). Component I(34%)
is slightly lower. ComponentsI and III maythere-forerepresent outer,and componentsII, IV,and
V, inner polypeptides of the CPV particle. It
must be understood, however, that the specific
location ofthe tyrosineresiduesin theindividual
viral
polypeptides isnot known. Thus, thepossi-bility
that tyrosine groups of the surface poly-peptidesareburied or those ofinternalpolypep-tidesare exposed cannot yetbeeliminated inany
interpretation
by using this technique of 1251labeling
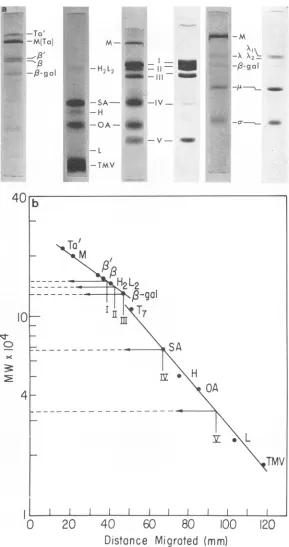
ofintact virusor subviral particles. Molecular weight determinations of CPVpolypeptides. Molecular
weights
of the poly-peptides from CPV were determined bycom-parison of their electrophoretic mobilitieson SDS
gels with those of proteins of known molecular
weights (35); polypeptides usedasstandardswere
tektin a' and a (220,000 and 200,000); myosin (200,000); Escherichia coli RNApolymerase sub-units
,3'
and ( (163,000 and 155,000); gammaglobulin H2L2 (150,000); ,B-galactosidase (130,000); bovine serum albumin (67,000);
gamma globulin H chain
(50,000);
ovalbumin(45,000); gamma
globulin
Lchain(22,000);
andtobacco
mosaic virus(TMV)
coatprotein
(17,500).
Figure 8a illustrates the comparative migrations of CPV and markers on 5%poly-acrylamide gels. When plotted as in
Fig.
8b,
these data show that, in the gel system
used,
migration
islinear within the220,000 to130,000
range and again within the 110,000 to
18,000
molecular
weight
ranges. The molecularweights
of CPV polypeptides I through V, determined
from the plots of
Fig.
8b,
are151,000,
142,000,
130,000, 67,000, and 33,000,
respectively.
Break-down product 1L seems to have resulted fromthe
cleavage
ofafragment(s)
of5,000
to6,000
molecular weight from
polypeptide
I.An incidental
point
arising
from thesede-terminations of molecular weights concerns the
Xi
andX2 polypeptides
of reovirus type III.Literature values place these two
species
at155,000 and 140,000, a difference of
15,000
(25). Wehave, however,
consistently
observed adifference ofonly
approximately
6,000(Fig.
8a).
Recently ithas beenshown that
electrophoretic
mobilitiesin SDSgelscanbeaffectednot
only
by
changes in molecular sizebut also
by
changes
in charge and molecular conformation(29)
caused bythechemicalmodification ofpolypeptides.
Todetermine whetherthe iodination
procedure
alterstheapparentmolecular weights of the viral
poly-peptides, iodinated
CPV polypeptides weresub-jected
to co-electrophoresis in the same gelwith unlabeled carrier polypeptides (figure notshown). Examination of theindividual slices for both dye stain and 1251 content showed that the iodinatedpolypeptides migrated,
ontheaverage, 5% moreslowly
than nontreated proteins.Consequently,
whenever molecular weight values were deter-mined in subsequent studies by using
iodinated
proteins versus non-iodinated markers, a 5% correction factorwasapplied.
Once themolecular weights of the CPV
poly-peptideweredetermined, it was of interest tosee
if,
as appears to be the case for reovirus (37),a clear correspondence exists between the size
of the dsRNA segments and thesize ofthe
dis-crete viral polypeptides. The genome of CPV
consists of 10 discrete dsRNA segmentsranging
in size from the largest, estimated at 2.7 X 106 daltons, to the smallest, at 0.37 X 106 daltons (14). It would appear (Table 1) that viral poly-peptides I through V could be coded for by dsRNA segments I, II,III, VI, andVIII, respec-tively.Thisleaves, however, five genome segments
either without a corresponding viral structural
protein or coding for proteins which might be
presentin the virus inamounts sufficiently small
to be undetectable by either dye retention or
1251I labeling.
Analysis of the polyhedral proteins of CPV.
We have already noted that the shape of the polyhedral body seems to be determined by the
strainof virus which itcontains. Itseemed
possi-ble, therefore, that the highly organized poly-hedral matrix (Fig. 4b) is composed of proteins which are also direct gene products ofthe viral
genome. To investigate this possibility,
poly-hedral bodieswerepurifiedon astep-gradient of
sucrose (13) andiodinated. The bulk ofthe
poly-hedral body appears, by microscopy, to be
ma-terial other than
virions-proper
(Fig. 4a). Henceit seemed reasonableto assume that the pattern
of in vitrolabeling would reflect the highcontent
of the polyhedral
polypeptide(s).
lodinated polyhedral bodies were separated from free,nonreacted1251byseveralcyclesofsedimentation
through step-gradients of sucrose and were
solubilized with urea-SDS. The labeled protein
was then analyzed by
polyacrylamide
gel elec-trophoresis in the presence of unlabeled viralmarkerpolypeptides. Theresults are seenin Fig.
9.
The labelingpattern would suggest that poly-hedral protein consists
primarily
of two major polypeptides with approximate molecular weights of29,500 and 19,500. There are,however,
minoramounts of labeled
protein
in discretepositions,
1062 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
M- e0mom
-H2L2EliliS-I
-H
-2L2
_-J4_ -I __ft
-M
-x
-(3- gal
.*4W -M-
_.
-SA-
41
-IV-OA
- -4
41 -QA-
_.J
., V
-L
I'l-T_
-TM V
20
40
60
80
10
Distance
Migrated
(mm)
FIG. 8. Molecularweightdeterminations oj CPVpolypeptidesbasedonelectrophoreticmigrationofassorted
markerspecies.a,Samplesweresolubilizedwith1%SDSandsubjectedtoelectrophoresison5%
polyacrylamide-0.1% SDSgelsat8 mApergel for17.5hr. Abbreviations: Ta' andTa, tektin a' anda;M, myosin;B' andB, E. coli RNApolymerase subunits B' and B; H2L2, gamma globulin H2L2; f8-gal, j3-galactosidase; SA, bovine
serum albumin; H, heavychain ofgammaglobulin; OA, ovalbumin; L, light chain ofgamma globulin; TMV,
TMVcoatprotein; I-V, CPVpolypeptides;X, IA, a, reoviruspolypeptides. Migration isfromtop to bottom, b,
Plotofmolecularweightversus distancemigrated.
1063 _4-w -Ta'
_swE -M(Ta)
-/3-9al
---crC a
40
10
0 >X
4
II
0
b
To'
/3;
m
9L
----_
-H2L
I I
3-gal
'III
F~~~
OA
K
I I I I I I I
)O
120
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.493.97.386.55.602.2]LEWANDOWSKI AND TRAYNOR
TABLE 1. Correspondence of'geniome segmenit size of cytoplasmic polyhedrosis virus to the size
of viral antd polyhedral polypeptides
CPV genome com-ponent no. I III IV v VI VII VIII Ix x Molecular weight Observed dsRNA segment' (X 106) 2.70 2.44d 2.15 1.85 1.14 0.83 0.62 0.54 0.37 Estimated ssRNAb transcript (X 106) 1.35 .22d 1.07 0.92 0.57 0.41 0.31 0.27 0.18 Estimated gene productc 150,000 136,000d 119,500 102,800 64,000 46,000 34,000 30,000 20,500 Polv-peptide observed in the virion 151,000 142,000 130,000 67,000 33,000 Poly-peptide observed in poly-hedral matrix 122,000e lOG,00(' 48,OO0e 29, 500 19,500
aCalculated from Lewandowski and Leppla (14) byusing reovirusRNAsegmentsasmolecularweightimiarkers.
bssRNA,Single-strandedRNA.
cEstimated molecular weightof gene pr-oduct = mol wt
ssRNA message/[mol wt nucleotide/(3 nucleotides'aa)] X mol wtaa. Mol wt niucleotide, 3 X 102; inol wt aa, 102;aa,
aminoacid.
dAverage valuesforcomponents anid 111 which migrate
closely.
eMinorpeaksindicatedbyarrowsinFig.9.
a fl
I lI12
CPVI: m I
ICe r!~~~~~~~~~~
1~~~~
Cr--05-1 ft _A(" dr) r,n ar) '7n O -- ,A ^IA
JIJF4u bs60 70 80 9C
Fractions (mm)
FIG. 9. Electropherogram of polypeptides from
iodinated,dissociated polyhedralbodiesofCPV.
Poly-hedral bodieswereputrifiedonzsucrosegradientts as
de-scribedinMaterialsanldMethods,
iodinated.for
15miii, anidsoltibilized with1%Z7O
SDS-8 Ml uireaat 100 C for 2mini. This labeled sample was mixed with unlabeled
marker CPV polypeptides anid sutblected to electro-phoresis oni 5c polyacrylamide-0.JO SDS gels at 7 mApergel,fbr21.5 hr. Migrationi isfrom lefttoright.
Arrowsindicate thepositionsofthreeminorpolypeptide species which, inadditionztothetwomajorpeaks,
corre-spontdinsizetoexpected viralgenieproduicts (seeTable 1).
someof which overlapthemarkerviral structural
polypeptides and several of which do not.
Com-parison with unlabeled CPV structural protein
markers and application of the 5 correction
factor associated with the iodination procedure
provide the molecular weight estimates seen in
the lastcolumn of Table1;themolecularweights
of the two major and three of the minor
poly-hedral polypeptides suggest that they might be
direct gene products of the five remaining
cis-tronsofthegenomeof CPV. As thepolypeptides
of the polyhedral matrix are being laid down around individual virusparticlesinthecytoplasm
ofthehostcell (27), it ispossiblethat othergene
products which, although perhaps not part of
the final virion superstructure but which were
functioning during virus replicationand
matura-tion, might easilybecome entrappedinthe
grow-ing polyhedral structure. This may explain the
presenceofthe minorpeaksinFig. 9.
WTV: Equilibrium sedimentafion and electron
microscopyof WTV.Arelationship between
reo-virusand WTV hasbeenproposedbasedon
elec-tron microscopy data showing somewhat similar
external morphologies andonthecommon
pres-enceofdsRNA as the geneticmaterial (2, 8, 28,
30).When sucrose-purifiedWTVwassedimented
to equilibrium in preformed gradients of CsCl,
thepreparation separated into threemajor classes
of light-scattering particles: two lower bands
which made up the bulk of the preparation, a
diffuse middle area, and an upper area of
obvi-ouslyless dense material. Immediate
resedimenta-tion of these three areasresulted in the fractions
seeninFig. 10.Thetwolower bands had buoyant
densities of 1.445 and 1.425 g/cm3, whereas the
middle diffuse area and the upper bands equili-brated at 1.39 to 1.41 and 1.30 g/cm3,
respec-tively. Dueprimarilyto previousreportsthat the
buoyant density of WTV ranges from 1.395 to
1.410 (D. R. Black, Ph.D. thesis, Univ. of
Cali-fornia, Berkeley, Calif., 1970; reference 7), the
middle diffuse area was arbitrarily designated V
(for virion). Becausethebuoyantdensities of the
lower bands approximately resembled those of
intact CPV and reovirus cores, these fractions
weretentatively designatedC(for cores),whereas
the top band was designated T (for top
compo-nent). The presence of upper bands (T) in the
centergradientwhich receivedonlythelower ma-terial(C) suggestedthatTcould ariseas a
break-downproductofC.
Electron microscopy of the original
sucrose-purified preparation and of the three fractions
resulting from CsCl resedimentation is shown
in Fig. 11. Figure lla shows a typical
sucrose-1064
J. VIROL.100 110 120
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.493.62.256.91.278.2]FIG. 10. Equilibrium sedimentation of wound tumor virion and subviral components. Sucrose-purified WTV
hadpreviously been sedimented to equilibrium in preformed CsCI gradients (p = 1.453 to 1.30g/cm3),anldthree
majorclasses ofliglht-scatterinzgparticlesresulted: two lower bandsequilibratinigatp = 1.445anidp = 1.425g/
cm1, termedfractiontC,amiddlediffusearea with a density from 1.39 to 1.41g/cm3, termnedfractioni V,anidan
upper area equilibratinigat p = 1.30g/cm3, termedfractionz T. Immediate resedimenztationz ofthese threeareas
resultedinthefractionis seent above. Allpreformedgradienlts were 3 ml in volunmewitha2-mlsampleoverlayanzd
wereruniat40,000
revlmiii
Jbr12hrinanSW50.1 rotor.purified preparation containing mainly intact virions
approximately
66nm in diameter. Figure1lb was selected to show some disrupting and
apparently empty virions as well as bundles of strands, possibly analogous to those seen with
CPV. The virions are hexagonal, possessing an
innercore area35to40nmin diameter(Fig. 1 c)
surroundedbyan outercapsid membrane. Figure
lid,
e, and f contain V, C, and T components,respectively. It is apparent that, compared to
reovirus or CPV, prior exposure of WTV to
cesium chloride has a strong degradative effect
when virus is subsequently dialyzed to remove
salts and observed by PTA negative staining.
Whereas both theVand Cfractionsseemtohave
retained aninternalcore structure, both are now
relativelypermeable to PTA anddisplay little of the highly organized capsidstructure seen inthe
preparation not exposed to CsCl. It is not yet
clear whetherthelossofcapsid material occurred
duringCsCI sedimentation or during the
subse-quent dialysis or PTA staining steps. Top
com-ponent (Fig. 1
If)
is composed mainly of emptyshells, completely permeable to PTA and devoid
of much of itscapsid structure and none of the
virus-free strands seen in the V and C
prepara-tions. Exposure ofWTV to chymotrypsin failed
to produce a particle with spike structures
ana-logous to thoseonreoviruscores orCPV.
Polypeptide composition of WTV and its
sub-viral forms. Samples of the sucrose preparation
and fractions C, V, and T from CsCI werethen
dissociated with SDS, labeled with 1251, and
ana-lyzedon polyacrylamide gels. The resulting
elec-tropherograms appear in the upper
portions
ofthefour panels of
Fig.
12. Since the -16 x 106 daltons of viral genome comprise 22% of totalvirionmass (12), we cancalculatethattotal
pro-tein mass is about60 x 106 daltons per virion.
With the sucrose gradient-purified mixture, the
1251 content of this protein mass is distributed
between two major polypeptide peaks (II and
IV), two small peiaks(IandIII), andseveralother
peaks not much greater than the background
(arrows, Fig. 12a). With the V fraction from
CsCl,the numberandheightof thesebackground
peaks were considerably increased, but again
peaks It and IV predominated (Fig. 12c).
De-spitethe appearanceinFig. lIeofalossofsome
of theoutercapsomerestructure, thepolypeptide
content offraction
ClI
(p = 1.425 g/cm3) shownin Fig. 12b is very similar to that of
sucrose-purifiedWTV seeninFig. 12a.Thoughnotshown,
thepolypeptide pattern offractionCI (p = 1.445
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.493.106.388.71.309.2]FIG. 1 1. Electron micrographsof WTV,preparedandstained as in Fig. 2. a andb, WTV purifiedon sucrose density gradientsbythe Kimuramodification as described in Materials and Methods;c, WTV partiallydisrupting
torevealan innercore-like area.Rods of potato virus X were added toenhance drop spreading.
d-f;
Viralcom-poieietsfromCsCIdensity gradienitcentrifuigationt (Fig. 10);d, componeiit V;e,
componentt
C;f;
componentt T.a-f; X160,000.
1066
on November 10, 2019 by guest
http://jvi.asm.org/
[image:14.493.61.448.46.649.2]I CPv IH1 & 7
3t ~~~~~~~(a)
- 2- I E
E2
o r 4
- 1!
'
.p
'3
2k
c
c
[image:15.493.107.395.67.421.2]Frcctions(mm)
FIG. 12. Electropherogram ofiodinated WTVpolypeptides. Upperportions of each part show viralfractionls
dissociatedwith1% SDSfor2minat 100Cpriortoiodinationfor10min.Lowerportionsofeachpanelshow
thesameviralfractions iodiniatedintactfor30secpriortodissociation. A,Sucrose-purifiedWTV. Unlabeled CPV
wasaddedtothissampleto serve asmarker,and thestainledpatternisseen atthe topofthefigure.Arrowsintdicate
the migration positions which wouldcorrespondtothose ofexpectedviral geneproducts (see Table2).b, WTV
"C,," bandisolatedfrom CsCIat theequilibriumpositionofp = 1.425g/cm3. c, WTV "V"areaisolatedfrom
CsCIattheequilibriumpositionofp = 1.410to1.390g/cm3.d, WTV"T"band isolatedfromCsCIatthe
equili-briumposition ofp = 1.30g/cm3.
g/cm3) was also much like that of fraction
CII.
This suggests that the apparent loss of
capso-meres in all the subfractions of
CsCl-exposed
WTV
(Fig. 12b-d)
maynothave occurredduring
sedimentation itselfbut rather as a result of
di-alysis against
low salt buffer or PTAstaining
priorto
microscopy
oftheCsCI-exposed
sample. Analysis of the polypeptide constituency ofWTV top component (T) is seen in the upper
portion of
Fig.
12d. In this case, no distinctpeaks arediscernibleatpositionsIandIII. Since
the bulk of the top component appears to be
composed of empty shells, the polypeptide
pat-ternin
Fig.
12dwould suggestthatcomponentsIIand IV
probably comprise
the membrane andcapsid polypeptides of these nucleic acid-free shells.
Location and molecular weight determinations
of WTVpolypeptides. Toinvestigate the locations ofthe viral polypeptides, samples ofthe sucrose
and CsClfractions ofWTV werelabeledprior to
dissociation and analyzed in parallel. Thelower
halves of Fig. 12a-d show that, inall cases, the
polypeptidemost susceptible to iodination in the
intact structures was species II. This tentatively
suggests thatpolypeptide II comprises the outer
capsomeric layer of WTV and its CsCl
subfrac-tions. Of the non-SDS-treated samples, the
tyro-sineresidues of polypeptide IV were available for
iodination only in top component (T), and then
(b) 3_
E
-2
44L
.0 l~
--~I
U-I
xcL\
i3-1
0 0 %
Au
-.41
.e. -?'if T
.
1, InEZ
I I
on November 10, 2019 by guest
http://jvi.asm.org/
LEWANDOWSKI AND TRAYNOR
TABLE 2. Corresponidelice ofgeniome se
oJ wolutnd tumor viruis to the size oj
associatedpolypeptides W\TV genome component no.
II
11 III IV-V VI-VII VIII IX X-XI XII XIII Molecular weight Ob-served dsRNA seg-menta (X 106) 2.80 2.30 2.08 1.72 1.08 1.01 0.77 0.56 0.54 0.33 Estimated ssRNAb transcript 1.40 1.15 1.04 0.86 0.54 0.50 0.38 0.28 0.27 0.16 Estimated gene product" 155,000 128,000 116,000 96,000 60,000 55,500 42,000 31,000 30 ,000 14,500,,Calculated from Lewandowski ar
(14) by using reovirusRNAsegmentsas
weight markers.
ssRNA,Single-strandedRNA.
See footnote b in Table 1.
only partially (Fig. 12d). The interpr
the data obtained with WTV seems t
cult,since the virus isunstabletoCsCl.
Molecular weight determinations
labeled polypeptideswere made by inc
labeled CPVinseveral gels. The migral
stained CPV markers is diagrammati
sentedatthetopof Fig. 12a. Basedon
the following values were assigned tc
distinct peaks of WTV polypeptide
156,000; II, 122,000;III, 63,000;and I
These molecular weight estimates are
in Table 2, which shows that a reaso'
respondence exists between the size
polypeptides presentinthevirion and
several of the estimated gene produ
latter values are derived from data
(
suggest that thegenomeofWTV iscoi13dsRNA segnents whichareelectrop]
separable into 10 distinct classes. One
matethat viral polypeptides Ithrough
be coded for by dsRNA segments,
VI(VII), and IX. The predicted elect
positions of the remaining estimated g
uctshave beendesignated by arrowsir It is not yet possible, however, to de
the minor peaks of polypeptide labi
around thesepositions correspondtoc
geneproducts.
DISCUSSION
When the genome of a
dsRNA-virion isintroducedintoahostcell,th
?gnmit
size tion which the viral RNA contains cannot bef virus- translated directly by the ribosomes;
transcrip-tion of single-stranded messages is first required. The RNA extracted from dsRNA viruses does
not appear to beinfectious. Transcription of the
input genome therefore does not appear to be
Polypeptide
accomplished by
any enzymepre-existing
in theobserved inthe virion uninfected cell, but rather by the RNA-dependentRNA polymerase introduced along with the re-spective templates. It would seem to follow that
156 ooo translation of these messenger RNA species
I then provides the remaining virus proteins
re-122,000 quired for successful multiplication of the virion.
Viral genome-specified polypeptides fall into
63,000 two categories, those which compose the
super-structure of the virion and nonstructural
polypep-44,000X° tides which function in virus-specified
biosyn-thesis. In the case of the dsRNA-containing vi-ruses, these two categories appear to overlap.
Whereas each reovirion has a
chymotrypsin-id Leppla
digestible
outer coat structurecomposed mainly
molecular of twoproteins,A2and the outermostcomponent
0J3, with CPV this role of protective outer coat
seems tobe borne on a collective basis by the
poly-hedralmatrix, also composed mainly of two pro-teins. The polypeptides most susceptible to
*etation of iodination with the intact viral or subviral forms o be diffi- are,2and 13forintactreovirus,theX components
for reovirus cores, components I and III for
of these CPV, and class II polypeptide for WTV. Unlike
ludingun- intactreovirus, purified WTV andCPV showno
tion of the detectable response to exposure to chymotryp-ically pre- sin.
thisstudy, All attempts in this laboratory to further
de-the four grade either reovirus core or purified CPV into
label: I, smaller enzyme-active particles have succeeded
[V, 44,000. only in eliminating polymerase
activity,
mostrecorded probably by dissociating the enzyme-template
nable cor- complex. Whatever treatment,chemicalor
physi-of those cal, is used toseparate RNAfrom the viral
pro-the size of teins appears also to destroy
biological
activity.
cts. These Furthermore, wehave had to date no success in
14) which obtaining template-free viral proteins in any form
mposed
of which,whencombined withexogenousviralRNAhoretically
in the proper substrate-salt milieu (4, 13, 22),- can esti- initiates transcription. Based on these facts, the
IIV
might grossstructureofreovirus cores, CPV,andWTV;rophoretic
arecompared
diagrammatically
in the uppergene prod-
portion ofFig.
13. Because WTV is ratherun-nFig. 12a. stable toCsCl,we canonly estimate the minimal
termine if structure of
transcription
which would beana-iel in and logous to reovirus cores and purified CPV. We
Jirect viral have assigned this role to fraction C from CsCl
(p = 1.425 to 1.445 g/cm') which, despite the
harsh effects of high salt, retains transcriptase
containing activity
(Lewandowski,
unpublished observation).einforma- One obvious
question
to arise from thiscom-1068 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
WTV tive aggregate equivalent to procaryotic and eu-caryotic host polymerase.
VIRAL AND TRANSCRIPTASE POLYPEPTIDES
E COLITRANSCRIPTASE
WOUND TUMOR VIRUS -+ t
+--BSUBTILIS TRANSCRIPTASE +--t --- t
CYTOPLASMICPOLYHEDROSISVIRUS --RATLIVER 0bTRANSCRIPTASE
20 15 C G5 02
[image:17.493.47.239.57.235.2]1oltons*I10
FIG. 13. Top, Diagrammatic presentatioli of the
structuresofreoviruscore, CPV,and WTV, represent-inzg what iskntown todateoftheminimal structure of diplornzavirus transcription. Bottom, Comparisoln ofthe
molecular weights ofselectprocaryotic andeucaryotic DNA-dependentRNApolymerase polypeptideswith the viralpolypeptidesassociated with thesuggestedminimal viral RNA-dependenztRNApolymeraseactivities.
parison concernswhich polypeptides among the
total protein mass of the minimal
enzyme-tem-plate complexareresponsiblefor the actual
proc-ess of transcription. Based primarily on the
striking similarity between the polypeptide
com-positionsofE. colideoxyribonucleicacid
(DNA)-dependent RNApolymerase and reovirus cores,
it has beensuggested that the reovirus cores are
made up entirely of transcriptase (11). Similar
comparisons can be made between the
polypep-tides of CPVandWTV,on onehand,and certain
procaryoticandeucaryotic DNA-dependentRNA
polymerases (5, 17, 34), onthe other (lower
por-tion of Fig. 13). The general pattern of these
compositional similarities suggests to us that
these viral and hostenzyme activitiesmay prove
tobeanalogous, perhaps differing chieflyintheir
preference of double-stranded template, either
DNA or RNA. Such a hypothesis raises the
in-teresting possibility of an evolutionary
relation-shipbetweendsRNA-containingviruses andthose
portions of procaryotic and eucaryotic genomes
codingforpolymerase activities. It must be
cau-tioned, however, that in allcases, including
reo-virus, inlieu ofa directreconstitution assay asis
available for the host polymerases, there is no
direct evidence showing which of the viral
poly-peptides designated in Fig. 13 is responsible for
the viral transcriptase activity. One attractive
possibility is, for example, that each of the
capsomeric-like structures seen onthe surface of
reovirus cores, CPV and WTV, in addition to
representing a major entity of virus
superstruc-ture, is also a multicomponent, biologically
ac-ACKNOWLEDGMENTS
WeareindebtedtoC. A.Knight for his cooperation during the
courseof thisresearch,toL.M.Black for thegenerousprovision of woundtumorvirus-inducedroot tumortissue, and toR. C. Williams, K. E. Richards, and J. Toby for their services in the electron microscopy studies.
This investigation was supported by Public Health Service grants AI 00634 from the National Institute of Allergy and In-fectious Diseases and CA 13169-01 from the National Cancer Institute, bycontract 71-2173 within the Special Virus Cancer Program of the National Cancer Institute, and by traininggrant GM 01389 from the National Institute ofGeneral Medical
Sci-ences.
LITERATURE CITED
1.Aruga, H., and Y. Tanada (ed.) 1971. The cytoplasmic-poly-hedrosis virus of the silkworm, p. 209. Univ. of Tokyo Press,Tokyo.
2. Bils, R. F., and C. E. Hall. 1962. Electron microscopy of wound-tumor virus. Virology 17:123-130.
3.Black, D. R., and C. A. Knight. 1970. Ribonucleic acid
trans-scriptase activity in purified woundtumorvirus. J. Virol. 6:194-198.
4. Borsa, J., and A. F.Graham. 1968. Reovirus: RNA
polymer-aseactivity in purified virions. Biochem. Biophys. Res.
Com-mun.33:895-901.
5. Burgess, R. R.1971.RNApolymerase. Annu.Rev. Biochem. 40:71 1-740.
6. DuPraw, E. J. 1970. Ultrastructure ofinterphase nuclei, p.
172. In DNA and chromosomes. Holt, Rinehart and Winston, Inc., NewYork.
7. Gomatos, P. J., and 1. Tamm. 1963.Thesecondarystructure ofreovirus RNA. Proc. Nat. Acad. Sci. U.S.A. 49:707-714. 8. Gomatos, P. J., and I. Tamm. 1963. Animal and plant
viruses with double-helical RNA. Proc. Nat. Acad. Sci. U.S.A. 50:878-885.
9. Hosaka, Y., and K. Aizawa. 1964. The finestructureof the cytoplasmic-polyhedrosis virus of the silkworm, Bombyx mori(Linnaeus). J.InsectPathol. 6:53-77.
10. Hukuhara, T. 1971. Variations in cytoplasmic-polyhedrosis virus, p. 61-78. In H. Aruga and Y. Tanada, (ed.), The cytoplasmic-polyhedrosis virus of the silkworm. Univ. of Tokyo Press, Tokyo.
11. Joklik, W. K. 1970. The molecular biology of reovirus.J.
Cell.Physiol.76:289-302.
12. Kalmakoff, J., L. J.Lewandowski, and D. R. Black. 1969. Comparison of theribonucleic acid subunits of reovirus, cytoplasmic polyhedrosis virus, and wound tumor virus. J. Virol. 4:851-856.
13. Lewandowski, L. J., J. Kalmakoff, and Y. Tanada. 1969. Characterization ofaribonucleic acid polymerase activity associated withpurified cytoplasmic polyhedrosis virus of thesilkworm, Bombyx mori. J. Virol. 4:857-865.
14. Lewandowski, L. J., and S. H. Leppla. 1972. Comparison of the 3'termini of discretesegmentsof thedouble-stranded RNA genomes of cytoplasmic polyhedrosis virus (CPV), wound tumor virus (WTV) and reovirus. J. Virol. 10: 965-968.
15. Lewandowski, L. J.,and S. Millward. 1971. Characterization ofthegenomeofcytoplasmic polyhedrosis virus. J. Virol. 7:434-437.
16. Loh, P. C., andA. J. Shatkin. 1968. Structural proteins of reovirus. J. Virol. 2:1353-1359.
17. Losick, R., R. G. Shorenstein, and A. L.Sonenshein. 1970. Structural alteration of RNA polymerase during sporula-tion.Nature(London) 227:910-913.
18. Luftig, R. B., S. S. Kilham, A. J. Hay, H. J. Zweerink, and W. K.Joklik. 1972. Anultrastructural studyofvirions and
coresofreovirustype3. Virology 48:170-181.
19. McConahey, P.J., andF.J.Dixon. 1966.Amethod oftracc
REOVIRUS CORE CPV
_ ++ ___ +
on November 10, 2019 by guest
http://jvi.asm.org/
LEWANDOWSKI AND TRAYNOR
iodination of proteins for immunologic studies.Int.Arch. Allergy29:185-189.
20. Millward, S.,and A. F.Graham. 1970. Structuralstudieson
reovirus: discontinuities in thegenome. Proc.Nat. Acad. Sci. U.S.A. 65:422-429.
21. Rohrmann, G. F., and R. G. Krueger. 1970. Physical, bio-chemical, and immunologicalproperties of coliphage MS-2 particles. J. Virol. 6:269-279.
22. Shatkin,A. J.,andJ.D.Sipe.1968. RNApolymeraseactivity in purified reoviruses. Proc. Nat. Acad. Sci. U.S.A. 61: 1462-1469.
23. Silverstein, S. C., C. Astell, D. H. Levin, M. Schonberg, and G. Acs. 1972. The mechainisms of reovirus uncoating and
geneactivationin vivo.Virology47:797-806.
24. Skehel, J. J., and W. K. Joklik. 1969. Studiesonthe invitro transcription of reovirus RNAcatalyzed by reoviruscores.
Virology 39:822-831.
25. Smith, R. E., H. J. Zweerink, and W. K. Joklik. 1969. Poly-peptidecomponents of virions, top componentandcores ofreovirustype 3. Virology39:791-810.
26. Stanley, P., and E. A. Haslam. 1971. The polypeptides of in-fluenza virus. V. Localization ofpolypeptides in the virion by iodination techniques. Virology 46:764-773.
27. Stoltz, D. B., and W. L. Hilsenhoff. 1969. Electron micro-scopic observations on the maturation ofa cytoplasmic
polyhedrosis virus. J. Invert. Pathol. 14:39-48.
28. Streissle, G., and R. R. Granados. 1968. The finestructureof wound tumor virus and reovirus. Arch. Gesamte Virus-forsch. 25:369-372.
29. Tung, J-S., and C. A. Knight. 1972. Relativeimportanceof
somefactors affecting the electrophoretic migration of
pro-teins in sodium dodecylsulfate-polyacrylamide gels. Anal. Biochem.48:153-163.
30. Vasquez, C., and P. Tournier. 1962. Themorphology of
reo-virus. Virology 17:503-510.
31. Verwoerd, D. 1970. Diplornaviruses: a newly recognized
groupofdouble-stranded RNA viruses. Progr. Med. Virol. 12:192-210.
32. Vinograd, J.,and J. E. Hearst. 1962. Equilibrium sedimenta-tion of macromolecules andviruses ina density gradient. Fortschr. Chem. Org. Naturst. 20:372-422.
33. Watanabe, Y. S., S. Millward, and A. F. Graham. 1968. Regulationof transcription of the reovirusgenome.J. Mol. Biol. 36:107-123.
34. Weaver, R. F., S. P. Blatti, and W. J. Rutter. 1971. Molecular structuresofDNA-dependentRNApolymerase (I1) from calfthymus andratliver. Proc.Nat. Acad. Sci.U.S.A. 68: 2994-2999.
35. Weber, K., and M. Osborn. 1969.Thereliability of molecular weight determinations by dodecyl sulfate-polyacrylamide gelelectrophoresis. J. Biol. Chem. 244:4406-4412. 36. Williams, R. C., and H. W. Fisher. 1970. Electron
micros-copyoftobacco mosaic virus under conditions of minimal beamexposure.J. Mol. Biol. 52:121-123.
37. Zweerink, H. J., M. J. McDowell, and W. K. Joklik. 1971. Essential and nonessential noncapsid reovirus proteins. Virology45:716-723.
1070 J. VIROL.