Vol. 56, No. 1 JOURNALOF VIROLOGY, OCt. 1985, p.293-297
0022-538X/85/100293-05$02.00/0
Copyright© 1985,American Society for Microbiology
NOTES
Formation of
a
Cruciform Structure
at
the Simian Virus
40
Replication Origin
Abolishes T-Antigen Binding
to
the Origin
In
Vitro
DANIEL G. TENEN,lt* LORAL. HAINES,' ULLA M. HANSEN,' ROBERT G. MARTIN,2
AND DAVID M. LIVINGSTON'
Dana-Farber CancerInstituteand the Harvard MedicalSchool, Boston, Massachusetts 02115,1 andLaboratory of
MolecularBiology, National Institute of Arthritis, Diabetes, andDigestiveandKidney Diseases, Bethesda, Maryland
202052
Received 28 March 1985/Accepted 5 June 1985
HeteroduplexDNAmoleculeswereformedby annealinganintact simianvirusreplicationorigin-containing
fragmenttoamutantderivativelacking the indigenous wild-type 27-base-pair (bp) invertedrepeatwithinthis structureandcontaining a nonhomologous 26-bpinverted repeatsequenceinitsplace. Results ofrestriction
enzymeand Si endonucleasecleavage analyses strongly suggestedthata13-bp stem-loopstructureformedat
thesite ofnonhomologybetween thesetwoDNAs. Thisstructurelies within theboundaryofsimian virus40 T-antigen-bindingsite2, and itspresenceinhibited T-antigen bindingtothatsequencebutnot toanadjacent
higher-affinity binding site (site 1). Therefore, the conformation of sequences within an otherwise intact
T-antigen-binding sitecanhave major effectsupon T-antigen bindingthere.
Theorigin of simian virus40(SV40) DNAreplication, like
other procaryotic and eucaryotic origins, contains a large inverted repeat sequence with the potential for forming a
stem-loop structure. This 27-base-pair (bp) sequence (a
perfect 13-bp invertedrepeatseparated by1bp) lieswithin a
37-bp strongbinding site, site 2, for SV40large Tantigen. Site2constitutesthe coreoftheviralreplicationorigin(1, 3,
7). Since bindingto this site isessential forthe initiation of
viral DNA replication, it was of interest to investigate the
effects ofthe formation ofa stem-loop structure atthis site
on T-antigen binding to the origin region in vitro. In this report, we describe the results of such experiments.
Previously, we described the formation of heteroduplex
molecules between replication origin-containing fragments ofwild-type SV40DNAandthose ofadeletionmutantthat
lacked 18 of the 27bp ofthe site2palindrome attheorigin (8). These heteroduplex molecules contained a stem-loop
structure consisting ofa 7-bp stem and a 3-bp loop on the
wild-type strand. The presence of this hairpin structure on
either the E or the L strand inhibited T-antigen binding to
either strand atsite 2. The presenceof this structureonthe Estrand alsoinhibitedT-antigen bindingtothe Ebutnotthe L strand at site 1, a high-affinity binding site abutting the
early sideofthecoreorigin sequence. Inspeculatingonthe reasons forT-antigen failing to bindto the altered site 2 in
these heteroduplexes, we pointed out that the asymmetric deletion within the 27-bp inverted repeat resulted in an
"abnormal" stem-loop (7-bp stem with a 3-bp loop versus
13-bp stemwith a 1-bp loop). It was therefore stillpossible
thatsuccessfulbindingtosite 2 would result ifastem-loopof the same size containing the same number of internal base
*Correspondingauthor.
tPresentaddress: Beth IsraelHospital,HarvardMedicalSchool,
Boston, MA02215.
pairs were formed at this locus in both DNA strands. The
experiments describedbelow eliminate thispossibility.
A mutant viral genome (pRGM29) containing a
substitu-tion of 26 bp of foreign sequence for the entire 27-bp
palindrome at the origin has recently been described (C. Nobileand R. G. Martin, submittedforpublication). The 26
bp substituted for the wild-type sequence also contained a
13-bp invertedrepeat,butthisrepeat wastotallyunrelated to the wild-type repeat. When heteroduplexes were formed between the deletion-substitution mutant and wild-type
DNA, the resulting hybrids were found to contain a
stem-loopstructureoneach DNA strand within site 2 (Nobile and
Martin, submitted), as shown below. The heteroduplex,
wild-type, andpRGM29 homoduplex DNAs were
individu-ally incubatedwithincreasingamountsofpurifiedTantigen, whichwas isolated fromSV40-infected monkey cells.
Bind-ing by the latter to the origin region of each DNA was
measured bythe DNasefootprint technique(4, 8, 9). The mutantand wild-type plasmids both contain pBR322
sequences
extending
from the SphI site clockwise to theHindIII site and SV40 sequences running clockwisefrom the
HindIll site at nucleotide 5172 to the SV40 SphI site at nucleotide 129. One mutant plasmid (pRGM28) had
sus-tained a 27-bp deletion from nucleotide 5230to nucleotide 13, and anothermutant plasmid (pRGM29) had sustained a
26-bp insertion atthe site of this deletion. The insert
con-tainedtwo Sall and a single EcoRI restriction site (Fig. 1).
These restriction site sequences were oriented sothatthey constituted a
26-bp
inverted repeatstructure(Fig.
1). Eachrecombinant plasmid was digested with MboII, which cleavesatpBR322 nucleotide4346,alocation 46 nucleotides downstream of the above-noted SV40 HindlIl site. The MboII-cleaved DNAwasthen 5' end labeled withT4
poly-nucleotide kinase after
digestion
of the 3'overhanging
se-quencewith T4 DNApolymerase.
Aftersubsequent
cleav-ageofeachplasmid
withMspI,a250-bp fragment containing
293on November 10, 2019 by guest
http://jvi.asm.org/
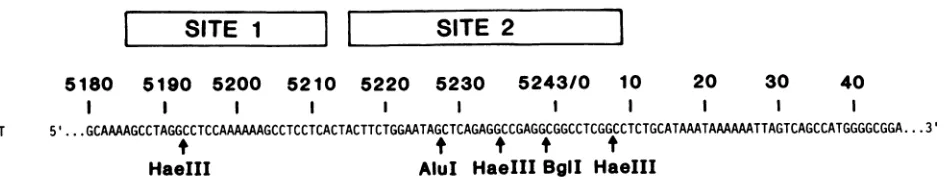
SITE
1
11
SITE 2
5180 5190 5200 5210 5220 5230 5243/0 10 20 30 40
WT 5' ...GCAAGCCTAGGCCTCCAAMAAGCCTCCTCACTACTTCTGGAATAGCTCAGAGGCCGAGGCGGCCTCGGCCTCTGCATAAATAAAAMTTAGTCAGCCATGGGGCGGA...3'
t
t
t
t
t
HaeIII AluI HaeIII BgII HaeIII
pRGM29 5' ...GCAM...3'GGTCGACCCCGMTTCGGGGTCGACC_...
Wild Tvnp
ICG GC
GC
AT
GC
CG
C G
G C
G C
A T
"I
.u
iy.,I
GA
5'. ..CTAGGCCTCCAAAAAAGCCTCCTCACTACTTCTGGAATAGCTC
3' ...GATCCGGAGGTTTTTTCGGAGGAGTGATGAAGACCTTATCGAc
C
pRGM29 A
C
T
GCATAAATAAAAAATTAGTCAGCCATGGGGCGGA.. .3'
GGTATTTATTTTTTAATCAGTCGGTACCCCGCCT
... 5'G
T
G C C G T A
G C G C
G C
G C C G A T A T
[image:2.612.89.561.70.160.2]FIG. 1. Possible cruciformstructure attheSV40originofreplication generated byaheteroduplexbetweenwild-typeDNAand themutant pRGM29. Thetop line shows thewild-type SV40 sequence from nucleotide 5180to46(usingthe SV numberingsystem[10]. Above this sequence are T-antigen-binding sites 1 and 2, as previously defined by DNase footprinting (9). Beloware the locations ofcleavage by
restriction endonucleasesHaeIII, AluI,andBglIinwild-typehomoduplexDNA.Thesecond line denotesthesequence ofthemutantplasmid pRGM29, which is identicalto that ofthewild type exceptthatwild-type bases 5230through 13were deleted and 26bases of restriction enzymelinker sequenceweresubstituted,asindicated.Atthebottom of thefigureisacruciformheteroduplexmolecule that could formas aresultofmixing wild-type andpRGM29 origin fragments(Nobile andMartin, submitted). Thearrowsdenotetwoofthefourpentanucleotide
sequencesfelttobeimportanttoT-antigenbinding atsites 1 and2(2,5).
the SV40 origin sequences and labeled onthe early side of
theoriginwaspurifiedbyelectrophoresis through
polyacryl-amide gels and identified by autoradiography. These
wild-type and deletion-substitution mutant DNA segments were
usedin subsequent experiments.
Thefirst questionwas whether Tantigen canbindto site
2from thedeletion-substitution mutant (pRGM29), as mea-suredbyDNasefootprint analysis. Previously, it was shown that an 18-bp deletion at this locus decreased, but did not
abolish, binding to this site (8). In the pRGM29 site 2 sequence,allfouroftheGAGGC pentanucleotide
T-antigen-bindingsequenceswithin the 27-bp inverted repeat (2; Fig. 1) had been deleted and substituted by linker DNA, and no
bindingtosite2 wasobserved at the highest T-antigen input, inkeeping with the results of others (2; Fig. 2). By contrast,
bindingto the wild-type sequence was clearly noted. More-over,T-antigenbinding to site 1 proceeded normally in both the5'-end-labeled mutantand wild-type DNAs (Fig. 2) and in their 3'-end-labeled equivalents (data not shown).
Al-though there was a suggestion of a small difference in the
amount ofT antigen needed to protect wild-type/wild-type
versuswild-type/mutant site1 sequences(see Fig.4),itwas notreproducible in repeated titrations.
Next, 250-bp heteroduplex DNA molecules were formed in vitro between the above-noted wild-type and deletion-substitutionmutant250-bpMboIIlMspI fragments,andtheir
structure was analyzed. Specifically, end-labeled wild-type
orpRGM29 DNA fragments were mixed withan excess of
thecorresponding,unlabeled mutantorwild-type fragment.
After denaturation and reannealing, each reaction mixture
was electrophoresed through a nondenaturing
polyacryl-amidegel. Repeatedly,wild-type/mutantheteroduplex
struc-tures migrated more slowly than did homoduplexes of the
same length, as was noted previously for DNA molecules
containinganotherstem-loopstructureinthis region (8) and for these molecules (Nobileand Martin, submitted).
There-fore, it was possible to purify the desired heteroduplex molecules away from the parental homoduplex molecules
(datanotshown). Analysisof the products ofSi
endonucle-ase cleavage ofsuch gel-purified heteroduplexes showed a
single Si-sensitivesite oneach strand resultingfrom cleav-age of1 to 2 bp centered about nucleotide 5243/0, i.e., the
I
on November 10, 2019 by guest
http://jvi.asm.org/
NOTES 295
WT
29
7r
-r
---
-
A
A
B
C
D
E
F
G
T(pg)
0
1.2 7.2 7.2 3.6 1.2
0
"'pplnqI
# __~~NA.W
_WT
*/29
A B
29*
/WT
C
D E
A
G
G
-20
p .- 1
I
5243/0
.--
-.5238
n-..--A_
,,w,lA. o.
5227
5221
_mp
-
5204
_
am
-5191
-. ...
18onee-
10
[image:3.612.55.290.70.504.2]. _
FIG. 2. SV40 T-antigen footprint protection of wild-type and pRGM29 homoduplex DNAs. The 250-bp wild-type (WT) and 249-bp mutant pRGM29 (29) MboIIlMspI fragments, each 5' end labeledatthe MboIIsiteatpBR322nucleotide 4346,weresubjected
toDNase footprint analysisafter the addition of T antigen purified fromlytically infected monkey (CV-1P) cells and preheatedat30°C for 5 min, as previously described (9). The amount of T antigen added isindicatedatthetopof each lane, and theDNA
concentra-tion ineach reaction mixture wasapproximately 3 x 10-10 M. A similar concentration was employed in all experiments. Bands
correspondingtobinding sites 1 and 2 (9)areindicatedby brackets
along the left side of theautoradiograph.
center of the axis of symmetry of each inverted repeat
sequence (Fig. 1 and 3). No such cleavage was observed
whenthe heteroduplexes were incubated in the absence of the enzyme. Moreover, no Si nuclease cleavage of the
wild-type ormutant homoduplex DNA was observed (data not shown). Similar resultswereobtained with these mole-culesbyNobile and Martin(submitted). Given theabnormal
electrophoreticbehaviorof theheteroduplex molecules,the inverted repeat nature of the wild-type and inserted linker
FIG. 3. Electrophoretic analysisofthe productsof Si nuclease cleavage of heteroduplex molecules. Heteroduplexes formed
be-tween 250-bp 5'-end-labeled (*) MboIIlMspI wild-type (WT) and unlabeled pRGM29 (29) fragments (WT*/29, lanes A and B) and 5'-end labeled pRGM29 and unlabeled wild-type fragments (29*/WT, lanes C and D)werepurifiedbynondenaturing
polyacryl-amide gel electrophoresis and subjected to partial Si nuclease cleavage as previously described (8). Markers included a partial
DNase digest ofa wild-type homoduplex fragment (lane E) and
products of Maxam-Gilbert chemical sequencing reactions (6) of 250-bpwild-type homoduplexMboIIlMspIfragments 5' endlabeled
at the MboII site (lanes G and G + A). The arrow on the left
indicates aband(s) at nucleotide 5243/0 produced by Si nuclease cleavage. The numbersonthe right referto specific nucleotides in the SV40sequence(10).
sequences, and the identification of a single Si nuclease-sensitive siteinananalogous placeoneachstrand,itseemed
likelythat themajorityofheteroduplexmoleculescontained the cruciformstructureshown inFig. 1. Inkeepingwiththis possibility, two restriction enzymes (BglI and HaeIII),
which normally cleave at sites in the wild-type sequences
thatconstitute partof the proposedheteroduplex cruciform
structure, failed to cleave the wild-type strand in the
--ffi~~~~-a
RW-1-1--E
r-K
n
4 ,~1~*: VOL.56, 1985
on November 10, 2019 by guest
http://jvi.asm.org/
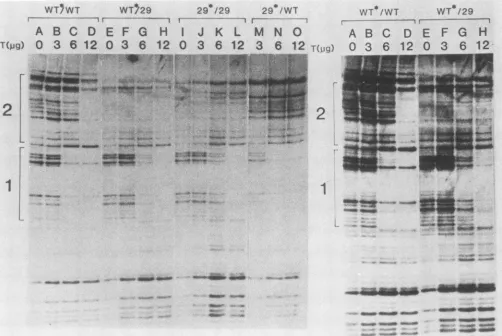
[image:3.612.311.554.71.485.2]296 NOTES
WT7WT
WT%29
29 /29 29 /WT--~~-- r -- ...--- --..
A
B
C
D
E
F G
H
I
J
K
L
M N
O0
3 6
12
0
3
6
12
0
3 6
12
3
6
12
7p
qj __::_:
T( 9)
r
'
2
WT
*/WT
WT /29A
B
C
D
E F
G
H
0 3
6
12 0 3
6
12
1
.. _._f
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~-
Wn:-S g- "mow'4'
-0
FIG. 4. T-antigen footprintprotection ofhomoduplex wild-type (WT*/WT), homoduplexmutant(29*/29),andtwoheteroduplex (WT*/29 and29*/WT) 250-bpMboIIlMspIDNAfragments,5'end labeledattheMboIIsite(*).Leftpanel,DNAwas5' end labeledattheMboIIsite
asindicated in the text and subjected to DNasefootprint analysis after addition of the indicated amountof purified T-antigen protein. Fragments analyzed included WT*/WT and 29*/29 homoduplexes (lanes A to D and I to L, respectively), a heteroduplex between
5'-end-labeled wild-type and unlabeled pRGM29 DNA fragments (WT*/29, lanes E to H), and a heteroduplex between 5'-end-labeled pRGM29 and wild-type DNA fragments (29*/WT, lanes M to 0). Bands corresponding to nucleotides within binding sites 1 and 2, respectively,areindicated bybracketsalongtheleft border. Right panel,Anoverexposureof lanes Athrough Hof theexperiment shown in theleft panel,demonstratingthe bandspresentin site 2 of thewild-typehomoduplexDNA and ofWT*/29DNA. Densitometerscanning of these bands in the WT*/29 heteroduplex lanes revealed nodecrease in intensity of bands within site 2 on T-antigen addition when
normalizedfor the totalintegralof theareasunder thepeaksineach lane. Densitometrywasperformedwith aJoyce-LoeblChromoscan3
densitometer.
heteroduplex molecules (data not shown). BglI resistance
was also observed by Nobile and Martin (submitted). By
contrast,anenzyme(AluI) that normally recognizesa
neigh-boring sequence believed to lie outside of the sequences constituting the cruciform structure cleaved the
hetero-duplex molecules efficiently (data not shown). In short, a
specific sequencethat abuts the proposed hairpin structure wasnotsufficiently perturbed in the heteroduplextobecome immune to restriction enzyme cleavage. Analogous results
were obtained previously with the hairpin arising from the formationofheteroduplexes between wild-type origin DNA
and a restriction origin fragment bearing an 18-bp deletion
withinsite 2(8).
When T antigen was incubated with the above-noted
heteroduplex molecules, 5' end labeled on either the
wild-typeormutantstrand,nobindingtosite2could be detected,
eventhough binding to site 1 was as efficient asbinding to site 1 inwild-type homoduplex DNA (Fig. 4). In particular,
nobindingwasobservedtowild-type site2sequenceswhen
theywerepresentintheabove-noted hairpin structure(Fig.
1), evenafter adding enough T antigentobind nearly all of the available site 2 sequences in wild-type homoduplex
molecules (Fig. 4). Results of two autoradiographic expo-sures are presented to indicate the lack of reduction in intensity of the detectable site 2 bands in the heteroduplex
DNAs. Asanadditionaltestof thiscontention,no
T-antigen-dependent reduction in intensity could be detected in densi-tometric analyses of the 5'-end-labeledwild-type/mutant site 2 bands in this experiment. Included among the
heteroduplex site 2 digestion products were certain rela-tively intense bands that were identical to those of the wild-type/wild-type digest. In thecaseofhomoduplexDNA, thesebandsweremarkedly suppressed bythe addition of12
,g of T antigen. Theywereunaffected, however, whenthe same amount of protein was added to wild-type/mutant
DNA. Similar results were obtained with the same DNA
fragments 3' end labeled on the other strand (data not shown). Moreover,asnotedearlier, with these DNAs there
was noapparent inhibition of site 1 binding.
Taken together, the data presented here show that it is
T(pg)
2
1
1
1..
J. VIROL.
,"-40-.-W
!.:;,:;4t:01 " .4,
_kmpa
--r -0.,
amw - Oww.
.,Al
t*W--*NM4winia
"i
40AWAM 1,
4'.. Aw-.Ow
co-Illi-10*W*-,*Pl
.,-.I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.67.569.70.406.2]NOTES 297
possible to generate DNA molecules which, in their native configuration, appear to contain stem-loop structures on
both strands at the viral replication origin. In the case
described here, the data suggest that two 13- by 13-bp hairpins were generated by annealing a molecule bearinga
wild-type inverted repeat at the SV40 origin of replication to another that had sustained a complete deletion of this structure and reinsertion of an artificial structure with the same dyad symmetry. When T antigen and this DNA mole-cule were mixed, binding was observed atsite 1, butnone
was detected within site 2, including to sequences within eitherhairpin structure. By contrast, in wild-type homodu-plex DNA, T antigen is known to bind to a sequence (site 2A)composed of 16 nucleotides flanking the early side of the 27-bp inverted repeat structure describedhere plus approx-imately 9 of 13 bp within thewild-type hairpin noted above (8). These 9 bp containone copy of the pentamer GAGGC believed to play an important role in promoting specific
binding of T antigen to sequences in this region (2, 5). Therefore,failuretobindto the same sequence now,inpart, present as astable hairpin structure could be interpreted in at least three ways, none of which are mutally exclusive. First, if theprotein is capable of binding to site 1 and to the 16nucleotides thatconstitute the segmentof site 2A
imme-diately adjacenttotheabove-noted wild-typestemandloop
in normal duplex DNA, then failure to observe such an
interaction in the case noted here may have resulted from
steric interference with a specific protein-DNA binding
event in this region. A second interpretation is that the
protein cannotbind stably to hairpin structures even when theycontainwild-type duplex sequences that normally
con-stitute a high-affinity T-antigen binding site. Third, the presenceofthe26-bpmutant insertionmay preventbinding
of T antigen to the heteroduplex site 2 sequence. From
earlier work, it is clear that binding to site 2 can be negatively affected by stem-loop formation within it even when the deletion mutant strand of the heteroduplex can
accommodate T-antigen binding when present in the
homoduplex configuration (8). Therefore, data consistent with at least one of these possibilities exist. Clearly, simple
formation of a symmetric heteroduplex hairpin structure, one strand of which is wild type, is not enough to
accom-modate specific stable T-antigen binding. Whatever the
detailedbasis for the failure of T-antigen bindinginthis case,
these data again suggestthatthe conformation ofsequences at andnearthe SV40 replication origincan have significant
effects upon the ability of Tantigen to bind there. Hence, if such structures form in vivo, they may have important regulatory effects on the initiation of autonomous viral genome replication or the regulation of the early and late viraltranscription units orboth.
Wesincerely appreciate the helpof Margaret Bradley in purifica-tionofTantigen and inotherdiscussions, and the assistance ofAnn Desaiinthepreparation ofthemanusript.
D.G.T. is the JamesM. FaulknerFellow of the Medical Founda-tion, Boston, Mass. This work was supported by
Public
Health ServicegrantCA15751 fromtheNational Institutesof Health.LITERATURE CITED
1. Bergsma, D.,D.Olive, S. Hartzell, and U. Subramanian. 1982. Territorial limits and functionalanatomyof thesimianvirus40 replication origin. Proc.Natl. Acad. Sci. USA 79:381-385. 2. DeLucia, A. L., B. A. Lewton, R.Tjian,and P.Tegtmeyer. 1983.
Topography of simian virus 40 A protein-DNA complexes: arrangementsofpentanucleotide interaction sitesattheoriginof replication. J. Virol.46:143-150.
3. DiMaio, D., and D. Nathans. 1980. Cold-sensitive regulatory mutantsofSV40. J. Mol. Biol. 140:129-142.
4. Galas,D.J., andA.Schmitz. 1978. DNAfootprinting: asimple methodfor detection ofprotein-DNA binding specificity. Nu-cleic AcidsRes. 5:3157-3170.
5. Lewton, B.A., A. L. DeLucia, and P.Tegtmeyer. 1984. Binding of simian virus40 AproteintoDNAwithdeletionsattheorigin ofreplication. J.Virol. 49:9-13.
6. Maxam, A. M., and W. Gilbert. 1980. Sequencing end-labeled DNA withbase-specific chemical cleavages. Methods Enzymol. 65:499-580.
7. Myers, R., S. Rio,A.Robbins, and R. Tjian. 1981. SV40gene expression is modulated by the cooperative binding ofTantigen toDNA. Cell 25:373-384.
8. Tenen,D.G.,D. M.Livingston, S.-S. Wang,andR.G. Martin. 1983.Effect ofastem-loopstructurewithin the SV40 replication origin upon SV40Tantigen bindingtoorigin regionsequences. Cell 34:629-639.
9. Tenen,D.G.,T.Taylor,L.Haines,M. K.Bradley, R. Martin, and D. M. Livingston. 1983. Binding of SV40 large Tantigen fromvirus-infectedmonkeycellstowild-typeandmutant viral replicationorigins. J. Mol. Biol. 168:791-808.
10. Tooze, J.1981. DNA tumorviruses:molecularbiology of tumor viruses, 2nd ed., part 2/revised. Cold Spring Harbor Labora-tory, ColdSpring Harbor,N.Y.
VOL. 56, 1985