0022-538X/92/095190-10$02.00/0
Copyright X 1992, AmericanSocietyforMicrobiology
Second-Site
Suppressor
Mutations Assist in
Studying the
Function
of the 3'
Noncoding Region
of
Turnip
Yellow
Mosaic Virus
RNAt
CHING-HSIU
TSAI',2
ANDTHEO W.DREHER'
2,3*Program inGenetics,1 Centerfor Gene Research andBiotechnology,3 and Departmentof
Agricultural
Chemistry,2
Oregon
StateUniversity,
Corvallis,Oregon
97331-7301
Received 6April1992/Accepted 19 May 1992
The 3' noncoding region of turnip yellow mosaic virus RNA includes an 82-nucleotide-long tRNA-like structure domain and a short upstream region that includes a potential pseudoknot overlapping the coat protein terminationcodon. Genomic RNAs withpointmutations in the 3'noncoding regionthat result in poor
replicationinprotoplastsandnosystemicsymptomsinplantawereinoculated onto Chinesecabbage plantsin
aneffort toobtain second-sitesuppressormutations.Putative second-site suppressor mutationswereidentified by RNaseprotection and sequencing and were then introduced into genomic cDNA clones to permittheir characterization. A C-57--U mutation in the tRNA-like structurewasa strongsuppressorof the C-55---A mutation which prevented both systemic infection and in vitro valylation of the viral RNA. Both of these phenotypeswererescued in the double mutant.AnA-107--C mutationwas astrongsecond-site suppressor
of the U-96-->Gmutation,permittingthe double mutant to establishsystemicinfection. The C-107 andG-96 mutations are locatedon opposite strands ofone helix ofa potential pseudoknot, and the results support a
functional role for thepseudoknotstructure.Amutationnearthe 5' endofthegenome(G+92- A),atposition -3 relativetotheinitiation codon of the essentialopenreadingframe206,wasfound to beageneralpotentiator
of viralreplication,probablyas aresultofenhancedexpressionofopenreadingframe 206. The A+92mutation enhanced thereplicationof mutant TYMC-G96inprotoplasts butwasnota sufficiently potentsuppressorto permit systemic spreadof theA+92/G-96double mutantin plants.
Turnip yellowmosaic virus (TYMV)has apositive-sense single-stranded RNA genome 6.3 kb long. The entire
ge-nomes ofaEuropean isolate (27) andan Australian isolate
(21) have been sequenced, and we have reported the se-quenceofTYMC, the Corvallis strain ofTYMV, which is
clonally propagated in cDNA form in pTYMC (10). In
common with several other plant viruses, including brome mosaic virus (BMV) and tobacco mosaic virus (TMV), the 3'-terminal region of TYMV RNA comprises a tRNA-like
structure (recently reviewed in reference 25). In TYMV RNA,this structure is82nucleotideslong(Fig. 1)andcanbe specifically recognized byseveraltRNA-associating proteins suchasvalyl-tRNA synthetase and (CTP, ATP):tRNA
nu-cleotidyltransferase from wheatgerm, yeast cells, or
Esch-erichiacoli(11, 17, 25) and by elongationfactor EF-Tu from E. coli(18)or EF-lao from wheat germ(19). The tRNA-like structuresarethoughttoplayarole in viral RNAreplication
(12), andfor TYMVthereisastrongcorrelation between the valine acceptance and replication in vivo of RNAs with mutations in the tRNA-like structure (37). The mechanism bywhichtheabilitytobe valylatedcontributestosuccessful replication remainsunclear, however.
Asecondreadilyrecognizable feature of the3' noncoding region ofBMVand TMV RNAs is the presence ofone or
more pseudoknots upstream of the tRNA-like structure.
Three consecutive pseudoknots arepresent at this location in TMV RNA (40), and mutations that delete or disrupt
secondary structural elements of the downstream
pseudo-knot result inaloss of replicationin tobacco plants orcells
* Corresponding author.
tTechnicalreport9862fromtheOregon Agricultural Experiment
Station.
(35). In the 3' noncoding region of BMV RNA3, there are
four consecutivepseudoknots(31),and deletion studieshave indicated the importance of this region for successful viral replication in barley protoplasts (22a). In TYMV RNA, a
potential pseudoknot overlapsthe termination codon ofthe
coat protein gene (32) (Fig. 1), but no experiments have investigated the role of this part of the TYMV genome in virus viability. Pseudoknots have recently become
recog-nized as structural elements involved in the regulation of
geneexpression, suchastranslationalread-through (42) and
frameshifting(2, 4, 8).Itispossible that therecognition and bindingofproteinstopseudoknots is involved in regulatory
events (30, 34, 36), and the interaction of one or more
proteinswith the pseudoknots present in the 3' noncoding regions upstream of the tRNA-like structures ofTMV and BMVRNAsmayparticipate ineventscontrollingviral RNA replication.
Our previous studies have characterized the decreased replication that results from certain substitutions in the 3' tRNA-like structure of TYMV RNA (37). During those studies,itwasnoticedthat inrarecasessystemicsymptoms
appeared (with considerable delay) whenmutants normally incapable of supporting systemic infectionwere inoculated
onto Chinese cabbage plants. Thesymptoms were
consid-ered to derivefrom thereplication ofanovel mutantvirus that hadarisenfromthe inoculum by acquiringone or more
mutationsas aresult ofpolymeraseerrorduring the limited
replication the mutants were capable of supporting. The
RNA-dependent polymerases of viruses are known to be relativelyerror prone (9). In the studiesreported here, our
aim was to explore the ability of such plant inoculation experiments to yield novel mutants with second-site
sup-pressor mutations that might open up new avenues for
5190
on November 9, 2019 by guest
http://jvi.asm.org/
SECOND-SITE SUPPRESSOR MUTATIONS OF TYMV RNA 5191 A.
/CUCU\
A CCCG-CCC UCGdAACCA3'
A i II | II . Ii\I -i
C GGGC GGG-AGCCU
G U -40 \ CA -20
uCA
U G -ro
U A U AUl
A C AAUA-sG
G-C A A C
C-G U A-U
G-C-so U-A
A-U C-G
G-C AUU-G-C UCUUGUC 5'
G -107Stp
U-A C
C-G U-A G-C U-A C-G
,,U
C C-53
C A
C AC
/-57
55A
B.
AU>\
A
A
C1-\
A-U U-A
C-G \_\
AUUG-CUCUUGAAUC
-107Stop
C.
A 0
A C
A-U U-A C-G
AUUG-CUCUUGCAUC
-107
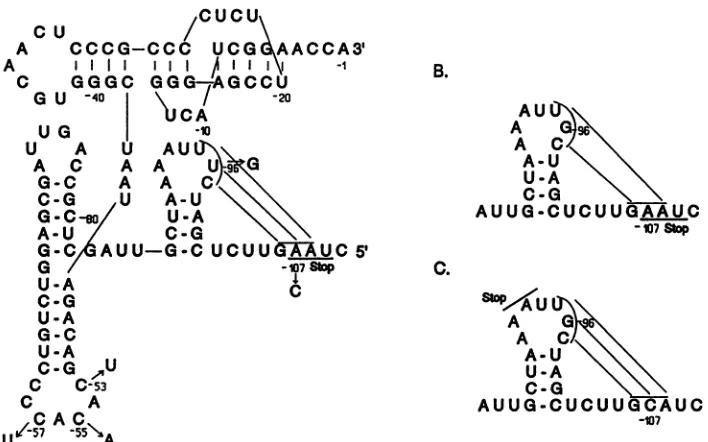
FIG. 1. Secondary structure model of the 3' noncoding region ofTYMV RNA. (A) The L conformation of the tRNA-like structure of wild-type TYMC RNA. A pseudoknot is present in the amino acid acceptor stem near the 3' end. The base pairs contributing to a potential pseudoknotupstreamof the tRNA-like structure are also shown, together with the coat protein termination codon (Stop), which it overlaps.
Thevariousmutationsin the 3' noncodingregion studied in this investigation are indicated. (B) Disruption of the potential pseudoknot in
mutant TYMC-G96RNA. (C) The potential pseudoknot of mutant TYMC-C107/G96 RNA, similar to that of TYMC RNA. The C-107 mutationcauses anextension of the coat protein ORF, to terminate as indicated. Nucleotides are numbered from the 3' end and are preceded byaminussigntodifferentiate from nucleotides near the 5' end (Fig. 2).
studying the role of the 3' noncoding region (including the potential pseudoknot) and tRNA-like structure of TYMV RNA. We show that second-site suppressormutations can
bereadily isolated, and wepresentresults basedon such a
mutation that support a role for apseudoknot immediately upstream of thetRNA-likestructure.
MATERIALSANDMETHODS
Materials. Chinese cabbage (Brassica pekinensis cv.
Wong Bok or SpringA-1) plants were grown in a growth
chamber under16-h day lengthat21°C. The hybrid cultivar Spring A-1wasusedinmostexperiments andwaspreferred
overWong Bok because of its greatergenetic uniformity.
PlasmidpTYMC(41), from whichinfectiousgenomicRNA (TYMC) can be transcribed with T7 RNA polymerase, is shown in Fig. 2. T7 RNA polymerase and reverse
tran-scriptasewerepurchasedfrom LifeSciences,m7GpppGcap
analogwas purchased from NewEngland Biolabs, Inhibit-Ace RNase inhibitorwas purchasedfrom5 Prime-3 Prime,
Inc., T7DNApolymerase (Sequenase)waspurchasedfrom United StatesBiochemical, Thermusaquaticus DNA
poly-merase was purchased from Promega, and restriction
en-zymes were purchased from GIBCO-BRL, Boehringer Mannheim, and NewEnglandBiolabs.Macerase and
Cellu-lysinwerepurchasedfrom Calbiochem.Synthetic deoxyoli-gonucleotides were made by automated phosphoramidite
synthesisandpurifiedon20%polyacrylamide-7Mureagels.
Protoplast andplantinoculations.Younghealthyleaves(3
to 4g)weretaken from 6-week-old Chinesecabbage plants
that had been held in the dark for 3days to deplete starch
grains. Protoplasts were prepared and inoculated (5 jig of
transcript RNA per 4 x 105 cells) as described previously (41)except thatprotoplasts were released after incubation of leaf slices in the hydrolytic enzymes overnight at 25°C. Inoculatedprotoplasts were incubated under constant light at25°C for 48 h priorto harvest.
Three-week-old Chinese cabbage plants with two true leaveswereused forinoculations of whole plants.Each leaf was mechanically inoculated with 10 ,ul of RNA transcript
(0.25
mg/ml)
in 50mMglycine-30mMK2HPO4
(pH 9.2)-1% bentonite-1% celite.Attimes, plantswereinoculatedwith asuspension
oflysed protoplasts
harvested 48h postinocula-tion(5
x104
cells perleaf).
Virion RNA extraction and
characterization.
Virus was isolatedfrom infected plants by polyethylene glycol precip-itationaccording
toLane(23) and quantitated spectrophoto-metrically. Virion RNA wasprepared from virus by double phenol-chloroform extraction,onechloroform-isoamyl alco-holextraction,
and ethanolprecipitation.
To assess thestability
of mutated sequencesduring
replication
inplants,
the
regions
encompassing
the relevant mutations were re-verse transcribed andamplified by polymerase
chain reac-tion(PCR) using
theoligomers
described below. The pres-ence of mutations was determined either directly by sequencing the PCRproducts
(43)
orby
digesting
with diagnosticrestriction enzymes that cleaved thewild-type
but notmutantsequence.Cloning and transcription. In the
preparation
of several mutants, PCR was used to generate smallfragments
for substitution intopTYMC.
Eachfragment
wascompletely
sequenced
prior
tosubcloning,
inorder toverify
the pres-ence ofonly the desired mutations.Sequences
withmuta-tions in the 3'
noncoding region
were PCRamplified
asVOL. 66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.138.492.102.323.2]5192 TSAI AND DREHER
ORF-69 Arg-His ORF-206Val-lie +1460 G--A
!EagI
CCGUACGC
T7promoter ORF-69 --I
1.0 2.0
EcoRl Ncol PstlI Bafurhil
i
+92
G
-A ORF-69 Ser-AsnCP
II
3.0 4.0 5.0 6.0
SstI PstI
smlHInd
II-53 C-->U -55 C--A -57 C--U -96 U->G -107AC
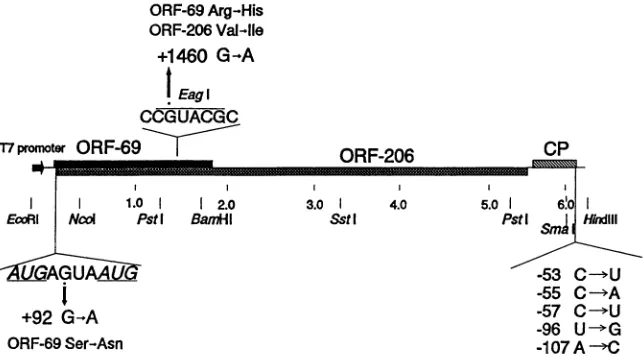
FIG. 2. Diagram of pTYMC, thegenomiccDNAclone from which infectioustranscriptscanbegeneratedwith T7 RNA
polymerase.
The mutations from the 5'region of thegenomethatwerestudied in thisinvestigationareindicated,togetherwiththe effect that eachsubstitution hasonORFs. The mutations in the 3'noncodingregionaresummarized(seeFig.1formoredetail).The three ORFs of TYMC RNA(ORF-69, ORF-206,and thecoatproteinORF[CP])areindicated,andrestriction sites used forcloningareshown.Nucleotides toward the5' endarenumberedconventionallyandmarked +,while thosenearthe 3' endarenumbered from the 3' endand marked -.
260-bpfragments,usingthe5'oligomer d(GGGTCAAAGAT TCGATTC) and the3' oligomer
d(TTCGAGCTCAAGCTT
GGTTCCGATG), which includes aHindIlI
site positioned atthe3' end of thegenomicsequence(39).
Mutant pTYMC-U53 was made in this way from cDNA clonempTY-U53,
which was used for in vitrovalylation studies
(13).
Mutants pTYMC-U57/A55 andpTYMC-C107/G96
were generated frommutantvirion RNAortotal RNA isolated from infected tissue, respectively, by PCRamplification
after reverse transcription primedby
the 3'oligomer
described above (20). The amplified mutant fragments were treated with E. coliKlenowpolymeraseto removenon-base-paired termini,
digestedwithHindlll,
andsubcloned into the SmaI(6062)
andHindIll (3')sites ofpTYMC(Fig. 2).
MutantpTYMC-G96wasgeneratedvia 3'PCR
amplifica-tion as described above. Afteramplification of260-bp frag-ments from wild-type pTYMC and from the fortuitously isolated mutantclonepTYMC-U53/G96, eachwasdigested withAluI (which cleaves 82nucleotides from the 3' end of the genomic sequence). The purified 5' fragment from pTYMC-U53/G96 and 3' fragment from pTYMCwerethen religatedandsubcloned into the SmaI(6062)and HindIII(3') sites ofpTYMC.
Mutant pTYMC+A92was also created via reverse tran-scription and PCRfrommutantvirion RNA. PCR to create pTYMC+A92 useda5' 59-meroligomer correspondingto a 5' EcoRI site and the T7 promoter fused toTYMC nucleo-tides 1 to 35 and the 3' oligomer d(ATGGTAATACATC AGG) to produce a 564-bp fragment. The mutant fragment wassubstituted into the EcoRI (5') andNcoI (210) sites of pTYMC. To make pTYMC+A1460, full-length cDNA was made from mutant virion RNA by using the 3' genomic oligomer, and second-strand DNA was made by priming with the 5' 59-mer. The mutant fragment was substituted into pTYMCby subcloning between thePstI (1309) andBamHI (1755) sites. [PCR analysis for the A+1460 mutation used the 5'oligomer
d(CCTGAGGCAACATTGG)
toprime at nucle-otide 1240 and the 3' oligomer d(AGCATGGACTTCTGT TCG)toprime opposite nucleotide 1573.]The combinatorial mutants G96+A92, pTYMC-G96+A1460, pTYMC-G96/U53+A92, and pTYMC-G96/
U53+A1460 were made by subcloning fragments, using restriction sites shown inFig. 2.
Plasmid DNAs were prepared from 50-ml bacterial cul-tures, and the mutantsequenceswereconfirmedby double-stranded DNAsequencing (5). Capped genomic transcripts labelled with [a-32P]UTP (0.1 Ci/mmol)werepreparedwith T7 RNA polymerase from DNA templates linearized with HindlIl and analyzed as described previously (41) prior to inoculation.
Analysis of viralproducts byWesternimmunoblottingand Northern (RNA) blotting. The levels of coat protein in harvestedprotoplastswereanalyzed in Westernblots,using horseradishperoxidase-labelled secondary antibodyand the chromogenic substrate 4-chloro-1-naphthol (detection limit of 2 ng ofcoatprotein)asdescribedpreviously (41).Results werequantitated byscanning laserdensitometrywith refer-ence to a dilution series ofvirus (37).
RNAwas extracted from protoplasts, glyoxalated, elec-trophoresed through 1% agarose, and transferred to nylon membranes asdescribed previously (41). The hybridization probewas a 32P-labelledRNAtranscript complementaryto 0.9kb at the 3' end of TYMV RNA(PstI-HindIII fragment; Fig. 2),permittingthedetectionof bothgenomic and subge-nomic RNAs(37). RNAlevelswerequantitated byscanning laserdensitometryorwitha,B-emission radioisotopescanner (Ambis Systems, San Diego, Calif.).
RNase protection assays. Antisense RNA probes (minus strand) wereprepared bytranscription with T7 or T3 RNA polymerase after subcloning appropriate fragments from pTYMC into the vectorpT7/T3a-18,which contains the T3 and T7 promoters (GIBCO-BRL). Probes representing TYMC sequences between the following restriction sites were used: full-length (Hindlll [3']-EcoRI [5']), HindIII (3')-NcoI (210),HindIII(3')-BamHI(1755),HindIII (3')-SstI (3286),BamHI(1755)-NcoI (210), and BamHI (1755)-EcoRI (5'). Virion RNAs (2 ,ug) were mixed with antisense RNA probes(1R,g)anddenaturedat85°C for 5 minin the presence of 40 mM piperazine-N,N'-bis(2-ethanesulfonic acid) (PIPES; pH 6.7), 0.4 M NaCl, 1 mM EDTA, and 80% formamide (in a 30-pI reaction). After hybridization over-night at 55°C, 300
RI
ofRNase solution containing 10 mM%irIF-gavu LTLILIMILIMM
-
J. VIROL.NR)F=onr%
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.151.473.79.258.2]SECOND-SITE SUPPRESSOR MUTATIONS OF TYMV RNA 5193
TABLE 1. Second-sitesuppressionof the G-96mutationa
Mutant Relative coat Appearance of Scored Sequence of
protein systemicsymptomsc progenye
TYMC
(wild type)
1.0 7 5/5 WildtypeTYMC-U53 1.1 7 2/2 U-53
TYMC-G96 0.02-0.3 14-21 14/21 (rf,U-96
TYMC-G96/U53 0.02 21 1/5 (r)9
TYMC-C107/G96 0.45 7 5/5 C-107/G-96
TYMC+A92 1.6 7 3/3 A+92
TYMC-G96+A92 0.21 7(11)h 9/9 A+92/U-96
TYMC-G96/U53+A92 0.02 ND ND ND
TYMC+A1460 0.9 7 1/1 A+ 1460
TYMC-G96+A1460 0.02-0.3 14-21 10/12 A+1460/U-96
TYMC-G96/U53+A1460 0.02 ND ND ND
aAlldatarepresent averages of at least threeexperiments. ND, not determined.
bCoat protein levels determined in Western blot analyses of extracts from Chinese cabbage protoplasts harvested 48 h postinoculation; error, =20%.
c Time of appearance (days postinoculation) of recognizable symptoms in the upper leaves of inoculated Chinese cabbage plants. d Number of Chinese cabbage plants with systemic symptoms/number of plants inoculated.
Sequences at the mutant loci of viral RNA in fully symptomatic systemically infected leaves.
f(r) refers to thedelayed appearance of systemic infection in two plants (genotypes found to be A+92/G-96 andC-107/G-96;see text). The remaining 12 symptomatic plants were infected with the revertant U-96 genotype.
g (r) refers to the delayed appearance of systemic infection in one plant (genotype found to be A+1460/U-53; see text). h Symptoms were observed in eight plants after 7 days and in one plant after 11 days (see text).
Tris-HCl (pH 7.5), 0.3 M NaCl, 5 mM EDTA, RNase A (32
,ug/ml),
and RNase T1(1.6 ,ug/ml)wasadded to the samples, which were incubated at 30°C for 1 h (16, 24, 28). RNases wereremoved bytreatmentwith50 ,ug of proteinase K per ml in the presence of0.5% sodiumdodecyl sulfate (SDS)at 37°C for 30 min. After phenol-chloroform extraction, the samples were ethanol precipitated and then analyzed by electrophoresis on agarose orpolyacrylamide gels.In vitrovalylation of transcripts. Fragments (264 nucleo-tides long) corresponding to the 3' region of TYMC and derivative mutant RNAswere synthesized by transcription fromBstNI-linearized cDNAclones and used in studieson valylation kinetics as described previously
Q13).
Valylation was determined by the incorporation of [ H]valine by a valyl-tRNA synthetase activity present in an extract from wheatgerm(13).Topermitthesynthesisoffull-lengthgenomicRNA tran-scripts capable ofaminoacylation, therestriction overhangs
generated
by HindIII linearization ofpTYMC
and selected mutant derivatives were removed by treatmentwith mung beannuclease (11). The treatedDNAs were extractedwith phenol-chloroformandethanolprecipitatedand then usedas templates for thesynthesis
ofgenomic transcripts by
T7 RNA polymerase (as described above except that the cap analog was omitted). The resultanttranscripts
werevaly-lated to determine the moles of valine bound per mole of mutant RNArelative towild-typeRNA.
RESULTS
Recovery of variants capable of
systemic
infection after inoculation withreplication-deficient
TYMC mutants. Our previous studies on the replication of TYMC RNAs with mutationsin the tRNA-likestructureyielded
twoinstances of phenotypic reversion after the inoculation of Chinese cabbageplants
withprotoplasts
infected withpoorly
repli-catingTYMCmutants(37).
TYMC-A55replicated poorly
in protoplasts andusually
produced
nosystemic
infection in plants, butdelayedsystemic
symptomsappeared
in one of three plants inoculated. In the secondinstance,
delayed
systemic symptoms werereported
in one of fiveplants
inoculated via
protoplasts
with RNAcontaining
a U-53mutation(37). Sincethe viruses responsible for both of these phenotypic reversions appeared to harbor second-site sup-pressormutations,wehave characterized these mutants and furtherexploredthepotentialforobtaining second-site mu-tationscapable ofsuppressingdeleterious mutations in the 3' noncoding region of TYMV RNA.
The plant with symptomsoriginating from a TYMC-A55 RNA inoculum developed systemic symptoms after 3 to 4 weeks,whereas symptoms appeared 7 days after inoculation with wild-type TYMC. Virion RNA was prepared and re-versetranscribedbyusingaprimer hybridizingtothe 3' end. The 3' region was amplified by PCR to yield a 260-bp fragment, whose sequencerevealed a
C-57---U
mutation in the wobble position of the anticodon in addition to the original A-55 mutation (Fig. 1). The U-57 mutation thus appearedtosuppressthe poorreplication phenotype result-ingfrom the A-55 mutation.Theplantwithsymptomsreported to have originated from TYMC-U53 RNAinoculumdevelopedsystemicsymptoms3 to 4 weeks after inoculation. As described in an authors' correction
(38),
we subsequently becameawareofan inad-vertent U-96---G mutation in the 3' noncoding regionthat had been introduced via PCR into our pTYMC-U53 clone. Thesystemic symptoms were thus derived from a TYMC-G96/U53 RNAinoculum,
which inmost casesdoesnotgive
riseto systemicinfection (Table 1). The sequence ofthe 3' regionof the virion RNA,obtained after PCR
amplification
asdescribedabove,revealed the presence ofonly
theU-53 mutation. RNaseprotection
experiments using
antisense RNAprobes indicated the presence ofanovel mutationin thisvirion RNA(Fig. 3)
thatwasshownby sequencing
oftheappropriate
PCR-amplified fragment
to be a G+ 1460->A substitution. As shown in Table 1, TYMC-U53replicates
likewild-type TYMC in both Chinese
cabbage
protoplasts
and plants. It thus
appeared
possible
that the A+1460 mutationwasabletopartially
suppresstheG-96mutation,
although the latter had reverted to wild typeprior
to full symptom development.Twofurthermutations with
potential
second-site suppres-sor activity were isolated after inoculation ofplants
with protoplasts infected with TYMC-G96 RNA. Fourteen of twenty-one plantsdeveloped
systemic
symptoms 14 to 21 VOL.66, 1992on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.61.560.89.219.2]WT R
1 2 3 4 5
--2036
--1635
--1018
--516
344 298
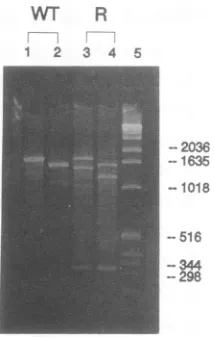
FIG. 3. Localization of the A+ 1460 mutationbyRNase protec-tion analysis. Virion RNAs from plants infected with wild-type
TYMC (lanes 1 and 2) or a genetic variant arising from the
TYMC-G96/U53 inoculum (lanes 3 and 4) were annealed to the
indicated antisense RNA probes and digested with a mixture of
RNases. The productswereelectrophoresed on a 1% agarose gel
and stained with ethidium bromide. Lanes: 1, wild-type RNA
probed with the EcoRI (5')-BamHI (1755) probe (1.75 kb); 2,
wild-type RNAprobed with the NcoI (210)-BamHI (1755) probe (1.55 kb); 3,mutantRNAprobedwith the EcoRI(5')-BamHI (1755) probe; 4, mutant RNAprobedwith the NcoI (210)-BamHI (1755) probe; 5,DNAmarkers(1 kb; GIBCO-BRL),withsizesindicated in
base pairs. The cleavage pattern ofthe mutant RNA indicates a
mutation about 310 nucleotidesupstreamof the3' endoftheprobe, i.e., at about nucleotide 1445; sequence analysis revealed the
A+1460 mutation.
days after inoculation,whileno symptomsdeveloped in the remaining seven plants (Table 1). Total RNAs were
ex-tracted from the symptomatic tissue of the 14 plants and
reverse transcribed with theTYMC-specific 3' primer, and the 3' 260-bp fragments were PCR amplified as described above.Digestion of the amplified fragmentswith the restric-tion enzyme DraI, which cleaves the wild-type T-96TTAAA-91 but not the mutant G-96TTAAA-91
sequence, followed by sequencing of Dral-resistant
frag-ments,showed that the G-96 mutationwasretained in 2 of
theplants but had reverted in the remaining 12. The virion
RNAfromoneoftheplantsthat retained the G-96 mutation contained an A-107--C mutation. Virion RNAs from the
other plant were subjected to RNase protection analysis followed by PCR amplification andsequencing, resulting in thedetection of a G+92->Amutation(the wild-type G+92
sequencewaspresentalso). Thenovel mutations from these
two plants appeared to be suppressors of the G-96
muta-tion.
The U-57 mutationsuppressesthedefectivevalylationand replication caused by the C-55-->A mutation. The U-57/ A-55 double mutation was transferred into pTYMC as
described above toyield pTYMC-U57/A55. The replication of TYMC-U57/A55 in Chinese cabbage protoplasts and plants was studied by inoculation with capped transcripts
made from HindIII-linearized plasmid DNA with T7 RNA polymerase andcomparedwiththat ofwild-typeTYMC and of theparentmutantsTYMC-U57andTYMC-A55(Table 2). Protoplasts inoculatedwithTYMC-U57/A55 RNA
accumu-latedcoatprotein (detectedonWesternblots)toalevel 85% of that of protoplasts inoculated with wild-type TYMC RNA; accumulations of genomic and subgenomic RNAs, determined from Northern blots,were 65 and45%,
respec-tively, of thewild-type levels(Table 2). Inoculationofplants resulted in systemicsymptoms indistinguishable from those produced by the wild type, and virions could be isolated from infected tissue inayield similar tothat fromwild-type infections(Table 2). Comparison of these results with those describing the replication of TYMC-A55 RNA (Table 2) clearly shows that the U-57 mutation isapotentsuppressor
of thedeleterious effect of theA-55 mutationwithregardto
replication.
Table 2 also summarizes our studies reported elsewhere
(13) onthevalylation properties ofa3' fragment of TYMV
RNAthat carries the UAA anticodon (U-57/A-55 double mutation). TY-U57/A55 RNAcould bevalylatedto comple-tion by wheat germ valyl-tRNA synthetase and had a
VmaxlKm (ameasure of theefficiency ofvalylation) of 0.075 relativetowild-type RNA. Bycontrast,TY-ASS RNA could beonly 13% valylated and hadarelative VmaxlKm of 0.0046
(13) (Table 2). With regardtovalylation, the U-57 mutation also stronglysuppresses the effectof the A-55 mutation.
Replication of TYMC-G96 and TYMC-G96/U53 in
proto-plasts. To characterize the replication properties of these
two mutants that gave rise to the suspected second-site
TABLE 2. U-57suppression of the A-55 phenotype'
Invitrovalylation properties Replication in Chinese cabbage relative to wt (genomic RNA as inocula)
Mutant (264-nt-long 3' RNAs)" Protoplasts harvested 48 hpi Plants
mol ofvalmol Relative Coat Genomic Subgenomic Relative Systemicsymptoms
ofRNAC
Vm.,,/Km
proteind RNAe RNAC virionyieldf scoregwt 1.0 1.0 1.0 1.0 1.0 1.0 9/9
TY-ASS 0.13 0.0046 0.02& Dh 1(r)/3h
TY-U57 1.0 1.1 105h°.9 0.8h 1.8h 5/5h
TY-U57/A55 1.0 0.075 0.85 0.65 0.45 1.0 3/3
aAll data are averages ofthreeor moreexperiments.nt,nucleotide;hpi, hourspostinoculation;wt, wild type; D,double-stranded genomicRNA detected by
invivo labelling(37) in place ofquantitationofsingle-strandedgenomicRNA,the very low level of which forthismutant resulted ininterferenceby inoculum
RNA;-, nonedetected;(r), reversion of phenotype resultingfrom acquisitionofsecond-site suppressor mutation.
bValylation by wheatgerm valyl-tRNAsynthetase, summarized from reference 13; error forVmax/K,,,determination is
"20%.
cExtent ofvalylationafter
60
minat 0.6 ,uM RNA(3'-CCA
species).dCoat protein levels determinedinWestern blots; error,
"20%.
e RNAlevelsdetermined inNorthern
blots; error,"20%.
fVirionyields determinedafterpurificationfrom systemically infectedleaves. g Numberofplants with systemic symptoms/numberof plants inoculated.
hData fromreference37.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.133.240.77.246.2] [image:5.612.65.565.565.658.2]SECOND-SITE SUPPRESSOR MUTATIONS OF TYMV RNA 5195
G-96
wr U-53 A+92 G-96 G-96 U-53 C-107G-96
U-53 A+92 G-96 A+92
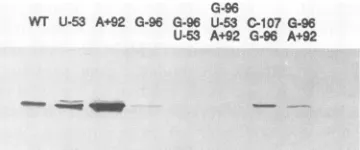
FIG. 4. Coat protein accumulations in Chinese cabbage proto-plastsinoculated with wild-type (wt) and mutant TYMC transcripts. Representativeexperiments that have contributed to the data shown in Table 1 are shown. Protoplasts were harvested 48 h after inoculation with the indicated derivatives of TYMC RNA. Extracts were separated on a 14% polyacrylamide-SDS gel, blotted, and probed with anti-TYMV antiserum. The blot was developed by using horseradishperoxidase-linked second antibodies and 5-chloro-1-naphthol color reagent.
suppressor mutations described above, capped genomic transcripts were prepared from G96 and pTYMC-G96/U53 and inoculated onto Chinese cabbage protoplasts. These clones were verified to contain only the indicated mutations by sequencing the mutant SmaI (6062)-HindIII (3') fragments (Fig. 2) that had been introduced into pTYMC (MaterialsandMethods). Chinese cabbageprotoplasts inoc-ulated with TYMC-G96 RNA accumulated highly variable amounts of coat protein, ranging between 0.02 and 0.3 relative to the wild-type level (Fig. 4; Table 1).In the course of these studies, we inoculated 10 separate batches of protoplasts (principally cultivar Spring A-1) with TYMC-G96 RNA, in some cases makingtriplicate inoculationswith the same preparation of transcript RNA. The results ap-pearedto vary, dependingon the batch ofprotoplasts rather than on other variables such as the transcript preparationor the particular inoculation. In previous experiments (37, 41), we have neverobserved suchvariability.
Since virusamplified in isolated protoplasts is not able to spread and infectothercells (spread in plants being via the plasmodesmatal connections between cells), it is unlikely that a spontaneous mutant capable of efficient replication could give rise to the higher levels ofreplication observed with some TYMC-G96 inoculations. Such mutations are relatively rare events and would need to occur indepen-dently in many cells, since theWesternsignals in protoplast inoculation experiments are the result ofviralreplication in at least 80% of the cells. Nevertheless, we PCR amplified andsequenced the 3' region of RNAsextracted from proto-plast batches that supported relatively high levels of coat protein synthesis and found the G-96 mutation (and no wild-typesequence) to bepresent inall cases. The frequent reversion toU-96 oninoculationofthe infected protoplasts toplants (Table 1) isconsistent with the absenceof second-site suppressor mutations in the RNAs replicated in the protoplasts. Taken together, these data lead us toconclude that the replication levels determined from Western blots refer to the replication of TYMC-G96 and not of some altered genotype. With regard to the variable levels of replication in protoplasts, we suggest that the deleterious effect of the G-96 mutation is sensitive to variations in the status of the host cells that wecurrently donotunderstand. Protoplasts inoculated with TYMC-G96/U53 RNA accu-mulated coat protein to levels 0.02 of the wild-type level (Fig. 4; Table 1), as previously reported (reference
37,
in which the mutant is incorrectly identified asTYMC-U53).
Unlike the results with TYMC-G96 inoculations, the
yields
ofcoatprotein were consistent between experiments. Table 1 andFig. 4 also show the replication propertiesof TYMC-U53, which hasaphenotypeinplantssimilartothewild-type phenotype.
The C-107 mutation is a potent second-site suppressorof the G-96 mutation. TheC-107/G-96doublemutation was introduced into pTYMC to yield pTYMC-C107/G96. The absence of other mutations in the 3' noncodingregion was verified by sequencing. The ability of genomic transcripts from this clone to support replication in protoplasts and systemic infection of plants was then studied andcompared with that of wild-type TYMC and mutantTYMC-G96 tran-scripts. Protoplasts inoculated with TYMC-C107/G96RNA consistently accumulatedcoatprotein to levels 45% of those of TYMC-inoculated protoplasts (Fig. 4; Table 1). When plants were inoculated with protoplasts infected with TYMC-C107/G96, normal systemic symptoms developed with no delay relative to infection with the wild type. The double mutation sequence has been stablymaintained after two serial passages through B. pekinensis cv. Spring A-1 plants. By contrast, systemic TYMC-G96 infection was never observed, and TYMC-G96 replication in protoplasts was variable but never as efficient as that ofTYMC-C107/ G96. The C-107 mutation is thus an efficient suppressorof the G-96 mutation.
The U-96---G mutation is positioned to disrupt base pairing that is partofa potentialpseudoknot overlappingthe UAA codon that terminates the coat protein open reading frame (ORF) (Fig. 1 and 2). The spontaneously recovered A-107--C mutation rescues the abilityforthe pseudoknot to form in TYMC-G96 RNA, by replacing the A-108 AG-106/-97CUU-95 stem of the wild-type RNA with
A-108CG-106/-97CGU-95
in the TYMC-C107/G96 dou-ble mutant (Fig. 1). In addition to its positioning within the potential pseudoknot, the C-107 mutation falls within the natural coatprotein terminationcodon U-105AA-107.The substitution ofUAAwith UAC is expected toresult in the addition of a five-amino-acid extension (Tyr-Val-Leu-Asp-Arg) to the 140-residue wild-type coat protein, with termi-nation occurringat theU-94AA-92 codon (Fig. 1). Prelim-inary results suggestthat the C-terminal extensionresults in less stable virions (36a).The rescued replication ofthe double mutant relative to TYMC-G96suggests afunctionforthepseudoknot. Wehave previouslyreported that sequences betweennucleotides -82 and -159 are involved in obtaining optimalvalylation rates (11), although this result has been
disputed
(26).
To deter-mine whether altered valylatability, which is crucial for replication (37), might explain thereplication properties
of mutants with substitutions affecting thepotential
pseudo-knot, thevalylationof these mutant RNAswasstudied. The kinetics ofvalylationbywheatgermvalyl-tRNA synthetase wasstudied in detail on mutantRNAscontaining
the 3' 258 nucleotides of viral RNA as describedpreviously (13).
The results presented in Table 3 show that the G-96 mutation (mutants TY-G96 andTY-G96/U53)
hadverylittle effecton valylation kinetics,less thandid the U-53 mutation(mutant
TY-U53), which had no detectable effect on theviralphe-notype in vivo.
Uncapped genomic RNAs with 3'-CCA termini
compati-ble with valylation were
prepared
for these mutants to determine whether the mutations in thepotential pseudoknot
could affectvalylation in the context of the 6.3-kb
genomic
RNA. A possible role for the
pseudoknots
upstreamof the tRNA-like structures inTYMV,
TMV,
and BMV RNAs might be as a spacerarm to ensurethespacial
separation
of VOL. 66,1992on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.91.271.76.151.2]TABLE 3. In vitrovalylationpropertiesof 3'genomic fragmentsa
Mutant mol ofofRNAb
Val/mol
ApparentKm.(nM) RelativeVmax RelativeVma.xlK.TY(wild type) 1.0 31 1.0 1.0
TY-G96 0.98 72 1.3 0.54
TY-U53C 1.1 183 1.9 0.32
TY-G96/U53 1.0 212 2.4 0.35
aValylation kineticswere determined by usingwheatgerm valyl-tRNA
synthetase and264-nucleotide-longtranscriptsthatinclude the 3'noncoding
region of TYMV RNA. RNA concentrations refer to the proportion of
3'-CCA-terminating transcriptspresent(13).ErrorsforKm and Vmaxare20%. wt, wild type.
b Extentofvalylation after 60 minat0.6,uMRNA.
cData taken from reference 13.
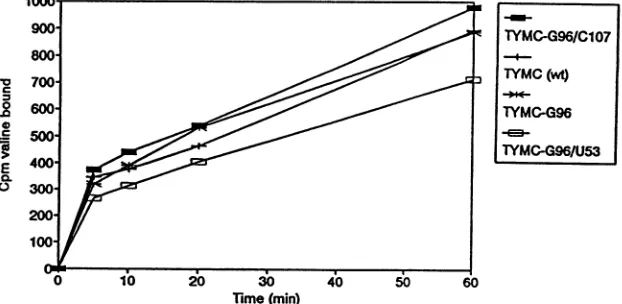
the3'tRNA-like domainfrom theremaininggenomic RNA, ensuringaccessibility to the relevantaminoacyl-tRNA syn-thetase and other proteins needed for replication. Only minor differences in the in vitro valylation bywheat germ valyl-tRNA synthetase were observed between wild-type TYMC, TYMC-G96, TYMC-C107/G96, and TYMC-G96/ U53 (Fig. 5). Thus, altered valylation does not explain the replication behavior ofmutants containing the C- 107 and G-96 mutations.
Mutation A+92 as a potential second-site suppressor of mutation G-96. Mutants TYMC+A92, TYMC-G96+A92, andTYMC-G96/U53+A92 wereconstructedandverifiedby sequencingto contain only the indicated mutations. Proto-plasts inoculated with TYMC+A92 RNAaccumulated coat proteintolevels1.6 times that present inwild-typeinfections (Fig. 4; Table 1), and inoculation of plants resulted in systemic infections indistinguishablefromwildtype but with higher virion yields (1.8 relative to the wild type). The mutantsequence was stablein plants. The G+92--A muta-tion lies between the tandem initiation codonsat the 5' end of theTYMV RNA (Fig. 2) (10) and results in a
Ser-*Asn
substitutionof thesecond codonofORF-69, which encodes aproteinthat is necessary for cell-to-cell movement in plants butnotforreplicationinprotoplasts(1).The A+92 mutation also places a more optimal nucleotide in the -3 position relative to the AUG codon that initiates ORF-206 (3, 22). Theimprovedreplication of TYMC+A92 relative to the wild type might thus be due to more efficient translation of ORF-206, which encodes proteins essential for replicationc
3
0E
a.
0
(41), although no clear enhancement of translation was observedin vitrobyusingareticulocyte lysate (notshown). Protoplasts inoculated withTYMC-G96+A92and TYMC-G96/U53+A92 RNAs accumulated coat
protein
to levels 0.21 and 0.02, respectively, relative to thewild-type level (Fig. 4; Table 1). The variable replication characteristic of TYMC-G96was notobserved. Both mutationswere shown bysequence analysis after PCRamplification
toberetained in the protoplasts inoculated with TYMC-G96+A92(not
shown). When protoplasts infected with TYMC-G96+A92 were inoculated onto plants, systemic symptoms appeared after 7daysineight plantsand after 11daysin theremaining plant inoculated (Table 1). RNA from the systemically infected tissues was assayed for the retention ofthe G-96 mutation by PCR-mediated amplification of 3' sequences followedby DraI digestionof the amplified fragment. Inall but theplantwithdelayedsymptoms, therewasareversion ofG-96tothe wild type U-96sequence(Table1),
while in each offiveplants tested,the A+92 mutationwasretained. Evenin theplant with delayedsymptoms, G-96waspresent onlyinearlysystemicinfection, being replaced bythe U-96 revertant by the time symptoms had fully developed. The replication of RNA with theG-96 mutation in early symp-tomatic leaves remote from the inoculated leaf suggests that the defectresultingfrom thismutation affectssomeaspectof viral infectionother than movement in the plant.The results presented above indicate that the A+92 mu-tation can act as apotentiator ofviralreplication,both in the wildtype and in theTYMC-G96mutant,perhapsby virtue of increased translational expression of ORF-206. The A+92-mediatedsuppressionofthe poorreplication phenotypethat results from the G-96 mutation is notsufficient, however,to supportsystemic spreadinplants,which occurredonlyafter reversion of the G-96 mutation. Reversion did nevertheless occur more frequentlyand more rapidly in thepresence of the A+92 mutation (compare TYMC-G96 and TYMC-G96+A92 in Table 1).
Mutation A+1460 as a potential second-site suppressor of mutation G-96. Mutants TYMC+A1460, TYMC-G96+ A1460, and TYMC-G96/U53+A1460were constructed and verified to contain only the indicated mutations. Protoplasts inoculated with TYMC+A1460 RNA accumulated coat pro-tein to levels of 0.9 relative to the wild-type level, and the TYMC+A1460 inoculum supported normal systemic symp-tom formation in planta (Table 1). The G+1460- A
muta-TYMC-G96/C107
--4-TYMC(wt) TYMC-G96 7YMC-G96/U53
rime(min)
FIG. 5. Comparative in vitro valylation of wild-type (wt) and mutant genomic RNAs. The full-length genomic RNAs indicated (0.2 ,uM) werevalylated (shownas counts per minute of[3H]valinebound per aliquot) with wheat germ valyl-tRNA synthetase.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.160.471.562.714.2]SECOND-SITE SUPPRESSOR MUTATIONS OF TYMV RNA 5197
tion, which results in an Arg--His substitution in ORF-69 andaVal-Ile substitution in ORF-206, thus had no notice-ablephenotype.
Protoplasts inoculated with TYMC-G96+A1460 and
TYMC-G96/U53+A1460
RNA accumulatedcoat protein to levels of0.02 to 0.3 and 0.02 relative to the wild-type level, respectively (Table 1). The variability of viral replication after inoculation with TYMC-G96+A1460 was like that following inoculation with TYMC-G96 RNA. Plants inocu-lated with lysates of protoplasts infected with TYMC-G96+A1460 developed systemic symptoms in 10 of the 12 plants inoculated, with symptoms appearing 14 to 21 days after inoculation. Dral digestion of the 3' 260-bp fragments amplified from total RNA extracted from the symptomatic leaves after reverse transcription and PCR showed that the G-96mutation hadreverted to the wild-type U-96 in all 10 plants. ThepresenceoftheA+ 1460mutationwasstudiedin thesystemic tissueby reverse transcription and PCR ampli-fication, followed by digestion with the EagI restriction enzyme, which cleaves the wild-type C1459GTACG1464, but not the mutant C1459ATACG1464 sequence. In all five progeny RNAs tested, the PCR products were EagI resis-tant, indicating the retention of the A+1460 mutation. Be-cause thereplication behaviors ofG96 and TYMC-G96+A1460 aresimilar,andsimilarlyvariable, it is clear that the A+ 1460mutationhas nodetectableactivity in modifying the phenotype of the G-96 mutation and is not a useful second-site suppressor.DISCUSSION
Inanerror-prone viralreplication system, a mutant with a poor replication rate has the potential to give rise to fitter, more successfully replicating variants that will rapidly out-grow and replace the original mutant inoculum. The new variant genomes will occasionally harbor second-site sup-pressor mutations that can help shed lighton the defect of the original mutation, often in unexpected ways. We have explored this approach for studying the role of the 3' noncoding region of TYMV RNA, using mutant RNAs TYMC-A55, TYMC-G96, andTYMC-G96/U53as the origi-nalinocula. TheseRNAsreplicate detectablyinprotoplasts, so that there is
significant
potential for the generation of mutations by polymerase error. They do not, however, supportsystemic
infectioninplants(Tables1and2),
sothat a more successful revertant or second-site suppressor mu-tantthatoutgrows the inoculumtoproduce systemic symp-toms is easily detected. We have investigated four novel mutationsgenerated
inthisway. Twoarepotentsecond-site suppressors thatprovide
newinsights,
discussed below. The third mutation(A+92)
is a weak second-site suppressor of the G-96 mutation that probably functionsnonspecifically
byenhancing
theexpression
of theessential ORF-206. The fourth mutation(A+1460)
does notdetectably
suppress the G-96mutation(from
which inoculum itwasrecovered)and appears to be a silent mutation under ourexperimental
conditions. Thus, either it was recovered entirely by acci-dent,orperhapsthemutation didact to
weakly
suppress the effects of the G-96 mutation in theparticular
conditions present in the inoculated plant(in
a way thatwe have notbeen able to
reproduce).
In any case, theinability
of the A+1460 mutation to prevent the reversion of thecoupled
G-96 mutation shows that it is not a useful second-site suppressor.
Anticodon recognition by valyl-tRNA synthetase and the
TYMV
replication system.Therecovery of theU-57muta-tion as asuppressor of the poor replication phenotype that results from the A-55 mutation has provided an important strengthening of our previously determined correlation be-tween valylation and viral replication (37). The two muta-tions were combined in the genomic mutant TYMC-U57/ A55, which replicates systemically togenerate virion yields similar to those of the wild type, and in a short transcript representingthe 3' 258nucleotidesof the viral RNA that can be conveniently used for in vitro valylation studies with wheat germvalyl-tRNA synthetase (13). As discussed more fully by Dreher et al. (13), the short
TY-U57/A55
mutant, with a UAA anticodon, is unexpectedly well valylated, as judged from the valylation activities of the single mutants TY-U57 and TY-A55. We havepostulated from the antico-operativitythat existsbetween the twomutationsthat valyl-tRNA synthetase is sensitive to the conformation of the phosphate backbone as well as functional groups on the bases of the anticodon (13). This is quite unlike the recogni-tion of thetRNAG'ln
anticodon by E. coli glutaminyl-tRNA synthetase, shown by X-ray crystallography to involve highly specificcontactsto thebase of each anticodonnucle-otide (33) in such a way that the anticooperativity between twomutations such as we have observed could not exist.
The unexpectedly efficient valylation observed for the U-57/A-55 double mutant has thus suggested a novel recognitionof the RNA by thevalyl-tRNA synthetase.More pertinent to our viral studies, however, is the correlation betweenthe viralreplicationandvalylation for TYMV RNA with thedeleterious A-55 mutation and TYMV RNA with the rescued
U-57/A-55
double mutation. This result strongly suggests that an interaction with valyl-tRNA syn-thetase,either as a catalyst that adds valine to the 3' end and then plays no further role or as a less transient ligand and subunitofthereplication complex, is crucial for the replica-tion of TYMV RNA. This interpretation is to differentiate from the theoretical but unlikely possibility that one of the viral proteins essential forreplication has an RNA binding domain much like that of valyl-tRNA synthetase, adomain that might have been recruited from the host gene and used inminus-strandpromoter binding. In fact, no homologies to aminoacyl-tRNA synthetases are known inTYMV-encoded ORFs, and it seemsinconceivable that a viralprotein inter-actingwith the tRNA-like structurein an essential function could duplicate the properties observed for the A-55 and U-57/A-55 mutations as well as the several anticodon mutations previously studied (37).Evidence for theexistenceof apseudoknot upstreamof the tRNA-likestructure. The G-96mutationisclearly detrimen-tal to the successful replication of TYMV. The mutation disrupts one of the stems of a potential pseudoknot just upstream of the tRNA-like structure (Fig. 1). A stable, strongsecond-site suppressor mutation that overcomes the effect of the G-96 mutation andpermits the virus to infect plants systemically is the C-107 mutation. The C-107/ G-96 doublemutant RNAwould beexpected to containa pseudoknot similar to that of the wild-type RNA, with a
C.
G rather than aU.
Abase pair. Although further mu-tants will need to be analyzed, it appears likely that the suggestedpseudoknotexists in TYMC RNA and is involved in some step that is important for the successfulreplication of the virus. It is unlikely that the extension of the coat proteinORFthat results from the C-107 mutationaccounts for the suppressor phenotype, since thelarger coatprotein
appears to form virions thatareunstable
during
normal virus isolation (36a). The extension to the coatprotein
ORF should thus be considered adetrimentalmutation,and it mayVOL. 66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
be responsible for the modestly suboptimal levels of coat
protein detected in protoplasts infected with TYMC-C107/ G96.
Nuclease cleavage and chemical accessibility studies are
consistent with the presence of the suggested pseudoknot,
although they are also consistent with adifferent
conforma-tion(14, 32). The short helical stems impartonly amarginal
stability, such that thepseudoknotmayneedtobestabilized by the binding of some factor in vivo. Analysis of the sequencesof othertymoviral RNAs shows thepotential for two pseudoknots upstream of the tRNA-like structure in ononis yellow mosaic virus RNA (6), but no potential
pseudoknots have been located (30a) in the RNAs of kennedya yellow mosaic virus (7) oreggplant mosaic virus
(29). Clearly, the relevance of the putative pseudoknot in TYMVRNArequires further experimentalsupport. In TMV RNA, the existence of three pseudoknots upstream of the tRNA-like structure is supported by nuclease mapping
ex-periments (15, 41), andanimportant function in someaspect
ofthereplication cycle has been demonstrated (35). Functional interaction between the G-96 and U-53
muta-tions. The U-53 mutation appears to be a neutral mutation
withrespect to replication in protoplasts and plants (Table 1). In combination with G-96, however, the U-53 mutation has an influence on the viral phenotype. In comparisons of
TYMC-G96 with TYMC-G96/U53 and TYMC-G96+A1460 with TYMC-G96/U53+A1460, the presence of U-53
re-sulted in a loss of the highly variable replication in
proto-plasts, with replication occurring at the lower levels ob-served in the absence of U-53. This consistently lower replication (by virtue of the lower opportunity for
polymer-aseerrors)presumably explains the observation that
TYMC-G96/U53 inoculumwas more stable in plants, with a lower
rate of reversion to U-96 than for TYMC-G96 inoculum (Table 1). Inacomparison of G96+A92 with
TYMC-G96/U53+A92, the U-53 mutation reversed thesuppressor
effect of the A+92 mutation, since TYMC-G96/U53+A92 replicated to only 0.02 times the level of the wild type in protoplasts, asjudged bycoatprotein accumulation.
No secondary structural interaction between G-96 and U-53 appears feasible (Fig. 1), but if such an interaction
wereto exist, it would presumably interfere with the valy-lation of the viral RNA and decrease the efficiency of replication. The genetic interaction between these two
mu-tant nucleotidesissufficiently strong toovercomethe effect
oftheA+92mutation in stabilizingahigher replication. The
molecular explanation for these effects will require further studies that may uncover unknown functional propertiesof
the3' noncoding region ofTYMV RNA. Theemergence of
this and the other questions discussed in thisreportfrom a
search for second-site suppressor mutations illustrates the
usefulness ofthis approach in studying the roles of the 3'
regions of viral RNAs.
ACKNOWLEDGMENT
This workwassupported byNSFgrantDMB-9019174. REFERENCES
1. Bozarth, C.S., J. J. Weiland,and T. W.Dreher. 1992.
Expres-sionof ORF-69 ofturnip yellow mosaicvirus is necessaryfor
viralspread in plants. Virology187:124-130.
2. Brierley, I., P. Digard, and S. C.Inglis. 1989. Characterization
ofan efficient coronavirus ribosomal frameshifting signal:
re-quirement foranRNApseudoknot. Cell57:537-547.
3. Cavener, D. R.,and S. C. Ray. 1991. Eukaryoticstartandstop
translation sites. Nucleic Acids Res. 19:3185-3192.
4. Chamorro, M., N. Parkin, and H. E. Varmus. An RNA pseudoknot andanoptimalheptamericshift sitearerequiredfor highly efficient ribosomal frameshifting on aretroviral
messen-gerRNA. Proc. Natl. Acad. Sci. USA 89:713-717.
5. Chen, E.Y.,and P. Seeburg. 1985. Supercoilsequencing: afast andsimple method forsequencing plasmid DNA. DNA 4:165-170.
6. Ding, S.-W.,P.Keese,andA. Gibbs. 1989. Nucleotide sequence of the ononis yellowmosaic tymovirus genome. Virology 172:
555-563.
7. Ding, S.-W., P. Keese, and A. Gibbs. 1990. The nucleotide sequence of the genomic RNA of kennedya yellow mosaic tymovirus-Jervis Bay isolate: relationships with potex- and carlaviruses. J. Gen. Virol. 71:925-931.
8. Dinman, J. D., T. Icho, and R. B. Wickner. 1991. A -1 ribosomal frameshift in adouble-stranded RNAvirus of yeast forms a gag-pol fusion protein. Proc. Natl. Acad. Sci. USA 88:174-178.
9. Domingo, E., and J. J. Holland. 1988. High error rates, popula-tion equilibrium and evolution ofRNA replication systems, p.
3-36. InE. Domingo, J. J. Holland, and P. Ahlquist (ed.), RNA genetics, vol. III. CRC Press, Boca Raton, Fla.
10. Dreher, T. W., and K. L. Bransom. 1992. Genomic RNA sequenceof turnip yellow mosaicvirus isolate TYMC,a
cDNA-based clone with verified infectivity. Plant Mol. Biol.
18:403-406.
11. Dreher, T. W., C. Florentz, and R. Giege. 1988. Valylation of
tRNA-like transcripts from cloned cDNA of turnip yellow
mosaicvirus RNA demonstrate that the L-shaped region at the 3' end of theviralRNAis not sufficient for optimal aminoacy-lation. Biochimie 70:1719-1727.
12. Dreher,T. W., and T.C. Hall. 1988. RNA replication in brome
mosaicvirus andrelatedviruses, p. 91-113. In E. Domingo, J. J. Holland, and P. Ahlquist (ed.), RNA genetics, vol. I. CRC
Press, BocaRaton, Fla.
13. Dreher, T. W.,C.-H. Tsai, C. Florentz, and R.Giege.
Biochem-istry, in press.
14. Florentz, C., J. P. Briand, P. Romby, L. Hirth, J. P. Ebel, and R.Giege. 1982. ThetRNA-likestructure of turnip yellow mosaic
virus RNA: organization of the last 159 nucleotides from the
3'-OHterminus. EMBOJ. 1:269-276.
15. Garcia-Arenal, F.1988. Sequence and structure at the genome
3' end of the U2-strain of tobacco mosaic virus, a histidine
acceptingtobamovirus.Virology 167:201-206.
16. Grange, D. K., G. S. Gottesman, M. B. Lewis, and J. C. Marini. 1990.Detection of pointmutationsintype I collagen by RNase
digestion ofRNA/RNA hybrids. Nucleic Acids Res.
18:4227-4236.
17. Joshi,S., F. Chapeville, and A. L. Haenni. 1982. Length require-mentsfor tRNA-specific enzymes andcleavagespecificity at the
3' end of turnip yellow mosaic virus RNA. NucleicAcidsRes.
10:1947-1962.
18. Joshi, R. L., H. Faulhammer, F. Chapeville, M. Sprinzl, and A. L. Haenni. 1984. Aminoacyl RNA domain of turnip yellow mosaic virus Val-RNA interacting with elongation factorTu.
Nucleic Acids Res.12:7467-7478.
19. Joshi, R. L., J. M.Ravel, and A. L. Haenni.1986.Interactionof
turnip yellow mosaic virus Val-RNA with eukaryoticelongation factorEF-la. EMBO J.5:1143-1147.
20. Kawasaki, E. S. 1990. Amplification of RNA, p.21-27. In M.A.
Innis, D. H.Gelfand, J. J. Sninsky,and T. J.White(ed.), PCR
protocols: a guide to methods and applications. Academic
Press, Inc., San Diego, Calif.
21. Keese, P., A. MacKenzie, and A. Gibbs. 1989. Nucleotide sequence of the genome of anAustralian isolate of turnip yellow mosaictymovirus. Virology172:536-546.
22. Kozak, M. 1987. At least six nucleotides preceding theAUG initiator codon enhance translation in mammalian cells. J. Mol. Biol.196:947-950.
22a.Lahser, F., and T. C. Hall. Personal communication.
23. Lane, L. 1986. Propagation and purification of RNA plant viruses. Methods Enzymol. 118:687-696.
24. Lopez-Galindez,C., J. A.Lopez, J. A. Melero, L. D. L. Fuente,
on November 9, 2019 by guest
http://jvi.asm.org/
SECOND-SITE SUPPRESSOR MUTATIONS OF TYMV RNA 5199 C. Martinez, J.Ortin, andM.Perucho.1988.Analysis of genetic
variability and mapping of point mutations ininfluenza virus by the RNaseAmismatch cleavage method. Proc. Natl. Acad. Sci. USA 85:3522-3526.
25. Mans, R. M. W., C. W. A.Plei, and L. Bosch. 1991. tRNA-like structures: structure, function and evolutionary significance. Eur. J. Biochem. 201:303-324.
26. Mans, R. M. W., P. W. G. Verlaan, C. W. A. Plei, and L. Bosch. 1990. Aminoacylation of 3' terminal tRNA-like frag-ments of turnip yellow mosaic virus RNA: the influence of 5' nonviral sequences. Biochim. Biophys. Acta 1050:186-192. 27. Morch, M.-D.,J.-C. Boyer, andA. L.Haenni. 1988.
Overlap-ping open reading frames revealed by complete nucleotide sequencing ofturnip yellow mosaic virus genomic RNA. Nu-cleic Acids Res. 15:6157-6173.
28. Myers, R. M., Z. Larin, andT. Maniatis. 1985. Detection of single base substitutions by ribonuclease cleavage at mis-matchesin RNA:DNAduplexes. Science 230:1242-1246. 29. Osorio-Keese,M.E.,P.Keese,and A.Gibbs. 1989. Nucleotide
sequenceof the genome ofeggplant mosaictymovirus. Virology 172:547-554.
30. PleU, C.W. A. 1990. Pseudoknots: a new motif in the RNA
game.Trends Biochem.Sci. 15:143-147. 30a.Plej,C. W.A.Personalcommunication.
31. Plet,C.W.A., J.P.Abrahams, A.van Belkum,K.Rietveld, and L. Bosch. 1987. The spatial folding ofthe 3' noncoding region of aminoacylatable plant viral RNAs, p. 299-316. In
M. A.Brinton and R. R. Rueckert (ed.), Positive strandRNA
viruses. Alan R.Liss, Inc., New York.
32. Rietveld, K. 1984. Ph.D. thesis, p. 118. State University of Leiden, Leiden, The Netherlands.
33. Rould,M.A., J. J. Perona, and T. A.Steitz. 1991. Structural basis of anticodon looprecognition by glutaminyl-tRNA
syn-thetase. Nature (London) 352:213-218.
34. Schimmel, P. 1989. RNA pseudoknots that interact with com-ponentsof the translation apparatus. Cell 58:9-12.
35. Takamatsu, N.,Y. Watanabe, T. Meshi, and Y. Okada. 1990. Mutational analysis of the pseudoknot region in the 3' noncod-ing region oftobacco mosaic virus RNA. J. Virol.64:3686-3693.
36. Tang, C. K., and D. E. Draper. 1989. Unusual mRNA pseudoknot structure is recognized by a protein translational repressor.Cell57:531-536.
36a.Tsai, C.-H.Unpublished data.
37. Tsai, C.-H.,andT. W. Dreher. 1991.Turnip yellow mosaic virus RNAswith anticodon loop substitutions that result in decreased valylation fail to replicate efficiently. J. Virol. 65:3060-3067. 38. Tsai, C.-H., and T. W. Dreher. 1992. Author's correction. J.
Virol. 66:3977.
39. Tsai, C.-H., andT. W. Dreher. In vitrotranscription of RNAs with defined 3' termini from PCR-generated templates. Bio-Techniques, in press.
40. vanBelkum, A., J.P.Abrahams, C.W. A.Plei,and L. Bosch.
1985. Fivepseudoknots are presentatthe 204 nucleotideslong 3' noncoding region of tobacco mosaic virus RNA. Nucleic AcidsRes.13:7673-7686.
41. Weiland, J. J., andT. W.Dreher.1989. Infectious TYMVRNA
from cloned cDNA:effectsin vitroandin vivoofpoint substi-tutions in the initiationcodons oftwoextensively overlapping ORFs. Nucleic Acids Res.17:4675-4687.
42. Wills,N.M.,R. F.Gesteland,andJ.F.Atkins. 1991.Evidence that a downstream pseudoknot is required for translational read-through of the Moloney murine leukemia virus gag stop codon. Proc. Natl. Acad. Sci. USA 88:6991-6995.
43. Winship, P. P. 1989. Animprovedmethod fordirectly sequenc-ingPCR-amplified material usingdimethyl sulfoxide. Nucleic Acids Res. 17:1266.
VOL. 66, 1992