0022-538X/07/$08.00⫹0 doi:10.1128/JVI.01778-07
Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Allogeneic Differences in the Dependence on CD4
⫹
T-Cell Help for
Virus-Specific CD8
⫹
T-Cell Differentiation
䌤
Christopher C. Kemball,
1† Eva Szomolanyi-Tsuda,
2and Aron E. Lukacher
1*
Department of Pathology, Emory University School of Medicine, Woodruff Memorial Research Building, Room 7307,
101 Woodruff Circle, Atlanta, Georgia 30322,1and Department of Pathology, University of
Massachusetts Medical School, 55 Lake Avenue North, Worcester, Massachusetts 016552
Received 13 August 2007/Accepted 24 September 2007
CD4ⴙ T-cell help enables antiviral CD8ⴙT cells to differentiate into fully competent memory cells and sustains CD8ⴙT-cell-mediated immunity during persistent virus infection. We recently reported that mice of C57BL/6 and C3H strains differ in their dependence on CD28 and CD40L costimulation for long-term control of infection by polyoma virus, a persistent mouse pathogen. In this study, we asked whether mice of these inbred strains also vary in their requirement for CD4ⴙT-cell help for generating and maintaining polyoma virus-specific CD8ⴙT cells. CD4ⴙ T-cell-depleted C57BL/6 mice mounted a robust antiviral CD8ⴙ T-cell response during acute infection, whereas unhelped CD8ⴙ T-cell effectors in C3H mice were functionally impaired during acute infection and failed to expand upon antigenic challenge during persistent infection. Using (C57BL/6ⴛC3H)F1mice, we found that the dispensability for CD4ⴙT-cell help for the H-2
b
-restricted polyoma virus-specific CD8ⴙT-cell response during acute infection extends to the H-2k
-restricted antiviral CD8ⴙ T cells. Our findings demonstrate that dependence on CD4ⴙT-cell help for antiviral CD8ⴙ T-cell effector differentiation can vary among allogeneic strains of inbred mice.
The differentiation of antiviral CD8⫹effector T cells during acute infection is believed not to require CD4⫹T-cell help, with helper T cells instead being critical for generating and/or maintaining competent memory CD8⫹T cells (5, 7, 8, 23, 41, 43). To combat viral infections that establish persistent infec-tion, an effective immune response may depend critically on CD4⫹T-cell help to sustain CD8⫹T-cell-mediated immunity and counter T-cell exhaustion resulting from repetitive antigen (Ag) encounter. In this connection, helper T-cell-deficient hosts infected by virulent variants of lymphocytic choriomen-ingitis virus suffer profound functional deterioration and dele-tion of antiviral CD8⫹T cells (20, 27, 33, 54). Additionally, CD8⫹T-cell control of murine gammaherpesvirus 68 infection is eventually lost in the absence of CD4⫹helper T cells (11). Thus, multiple variables of virus-host interactions during per-sistent infection (e.g., viral load, activation state of Ag-present-ing cells, and bystander inflammatory mediators) may modu-late the degree to which CD4⫹ T-cell help is needed for antiviral CD8⫹T-cell responses.
Using mouse polyoma virus (PyV), we extended this investiga-tion to the CD4⫹ T-helper-cell dependence of virus-specific CD8⫹ T-cell and humoral responses to a low-level systemic persistent viral infection. PyV, a natural mouse pathogen, es-tablishes silent persistent infection in outbred mice and immu-nocompetent inbred strains of mice (40). Virus-specific CD8⫹
T cells check persistent viral loads and protect against PyV tumorigenesis (19, 29, 35). Using major histocompatibility complex (MHC) class II-deficient C57BL/6 (B6) mice, we cently determined that the expansion, differentiation, and re-call potentials of PyV-specific CD8⫹effector T cells are unaf-fected by the absence of CD4⫹ T-cell help, although their numbers decline dramatically during persistent infection (26). We previously showed that naı¨ve, virus-specific CD8⫹T cells are continuously recruited during the persistent phase of PyV infection (25, 49). In MHC class II-deficient B6 mice, however, the recruitment and/or maintenance of anti-PyV CD8⫹T cells is dramatically impaired (26), suggesting that CD4⫹T-cell help may also be needed to resupply a deteriorating virus-specific CD8⫹T-cell population during persistent infection.
There is also evidence that allogeneic differences affect host requirements for mounting Ag-specific CD8⫹T-cell responses. For example, B6 (H-2b) and C3H (H-2k) mice differ in their
ability to generate allospecific CD8⫹T cells in the absence of CD28 and CD40L costimulation (52). With respect to PyV infection, we recently demonstrated that mice of these two inbred strains vary in their ability to control chronic infection after combined CD28 and CD40L costimulation blockade (24). This unexpected mouse strain difference in antiviral immuno-surveillance led us to ask whether these strains also differ in their dependence on CD4⫹ T-cell help for generating PyV-specific CD8⫹T cells. In this study, we found that CD4⫹T-cell depletion in C3H mice, but not B6 mice, adversely affected the effector differentiation of PyV-specific CD8⫹ T cells during acute infection and impaired their capacity to expand upon Ag-specific challenge during persistent infection. The defective H-2k-restricted PyV-specific CD8⫹T-cell response in CD4⫹ T-cell-depleted C3H mice was overridden in CD4⫹T-cell-depleted (B6⫻C3H)F1(B6C3F1) mice. These findings indicate that host
background affects the dependence on CD4⫹T-cell help for
gen-* Corresponding author. Mailing address: Department of Pathology, Emory University School of Medicine, Woodruff Memorial Research Building, Room 7307, 101 Woodruff Circle, Atlanta, GA 30322. Phone: (404) 727-1896. Fax: (404) 727-5764. E-mail: alukach@emory .edu.
† Present address: Molecular and Integrative Neurosciences De-partment, The Scripps Research Institute, 10550 North Torrey Pines Road, Mail Code SP30-2110, La Jolla, CA 92037.
䌤Published ahead of print on 3 October 2007.
13743
on November 8, 2019 by guest
http://jvi.asm.org/
erating optimal CD8⫹T-cell responses to persistent virus infec-tion.
MATERIALS AND METHODS
Mice.B6 and C3H/HeN female mice were purchased from the Frederick Cancer Research and Development Center of the National Cancer Institute. B6C3F1 mice were purchased from Charles River Laboratories. Mice were housed and bred in accordance with the guidelines of the Institutional Animal Care and Use Committee and the Department of Animal Resources at Emory University.
Viruses and inoculations.Adult female (6 to 12 weeks of age) mice were inoculated subcutaneously (s.c.) in each hind footpad with 2⫻106PFU of PyV
strain A2. Recombinant vaccinia virus (VV) carrying a minigene for the PyV MT389-397 sequence (VV-MT389) was prepared as described previously (18). The mice received 1 ⫻106
PFU of VV-MT389 or VV Western Reserve (VV-WR) intraperitoneally (i.p.).
CD4ⴙT-cell depletion. The mice received 250g of rat anti-mouse CD4 antibody (Ab) (clone GK1.5) or 250g of ChromPure rat immunoglobulin G (IgG) (Jackson ImmunoResearch) i.p. on days⫺3,⫺1, and⫹1 relative to the day of PyV infection. This regimen depleted⬎97% of CD4⫹splenocytes by the time of infection with PyV as assessed by flow cytometry using a CD4 monoclonal Ab (MAb) that does not compete with GK1.5 for binding (clone RM4-5; data not shown). For long-term depletion, mice were also treated with 250g of MAb GK1.5 or rat IgG weekly for 8 weeks, and the absence of CD4⫹T cells in GK1.5-treated mice was confirmed at the endpoint of the experiments by flow cytometric analysis of splenocytes with CD4 MAb RM4-5 (data not shown).
Peptides, cell surface and intracellular staining, and flow cytometric analyses. LT359-368C7Abu, MT245-253, LT638-646, and MT389-397 peptides were syn-thesized and stored as described previously (24, 25). Red blood cell-lysed spleen cells were directly stimulated ex vivo with synthetic peptides (1 or 10M) and stained for surface CD8␣and intracellular gamma interferon (IFN-␥), as de-scribed previously (25). To detect cellular degranulation, 0.5g of CD107a and CD107b Abs (BD Biosciences) was included during peptide stimulation, as described previously (6). The total number of Ag-specific, IFN-␥⫹CD8⫹T cells
was determined by subtracting unstimulated IFN-␥⫹CD8⫹cells from peptide-stimulated IFN-␥⫹CD8⫹cells. In some experiments, mice received
bromode-oxyuridine (BrdU) in the drinking water (0.8 mg/ml) for 8 days, and spleen cells were stained using a BrdU flow kit (BD Biosciences) according to the manufac-turer’s instructions. Allophycocyanin-conjugated Db
LT359-368 or Dk
MT389-397 tetramers were used to stain spleen cells. CD4, CD8␣, NKG2A/C/E (clone 20d5), IFN-␥, CD44, Ly6C, and isotype control Abs were purchased from BD Biosciences. CD27, CD122, CD127, NKG2A (clone 16a11), PD-1, and isotype control Abs were purchased from eBioscience. CD11a, CD62L, granzyme B, and isotype control Abs were purchased from Caltag Laboratories. It merits pointing out that while the clone 20d5 MAb used to detect NKG2A also recognizes the NKG2 isoforms NKG2C and NKG2E, it is the only MAb available to detect NKG2A in C3H mice; moreover, MAb 20d5-sorted Ag-specific CD8⫹T cells predominantly express NKG2A transcripts (22, 34, 53), and in B6 mice, we have found that an allele-specific NKG2A-specific MAb (clone 16a11) (48) and MAb 20d5 stain similar percentages of LT359-specific CD8⫹T cells (C. C. Kemball and A. E. Lukacher, unpublished observations). Samples were acquired on a FACSCalibur (BD Biosciences), and data were analyzed using CellQuest soft-ware (BD Biosciences).
51Cr release assays.RMA/S (H-2b) cells were labeled overnight at room
temperature with 200Ci of Na2 51
CrO4(PerkinElmer) in 200l of Iscove’s
modified Dulbecco’s medium (Invitrogen) containing 10% fetal bovine serum and LT359 peptide (10M final concentration) or medium alone. AG104A (H-2k) target cells were prepared as described previously (29), except that cells
were pulsed with 10M MT389 peptide (or medium alone) overnight and then labeled with Na251CrO4for 90 min at 37°C. Target cells were aliquoted at 5,000
cells/well into U-bottomed 96-well microtiter plates (Costar) and cocultured with red blood cell-lysed spleen cells for 4 h at 37°C, with51Cr detected by a Packard
Cobra 5005 gamma counter. Ag-specific effector:target ratios were calculated based on the frequencies of tetramer⫹CD8⫹T cells. The percent specific lysis was calculated as [(51
Cr release with effector cells⫺spontaneous51
Cr release)/ (total51Cr release with 1% Triton X-100⫺spontaneous51Cr release)]⫻100.
The spontaneous51
Cr release from target cells in all assays was 10 to 15% of the total detergent lysis. The percent specific lysis values are plotted as the means⫾
standard errors of the means of the results for quadruplicate wells.
Plaque assays.PyV plaque assays were performed as described previously (10). The limit of detection of this assay is 1 PFU/mg of spleen. Ovaries from VV-infected mice were snap-frozen in sterilized Kontes tubes, resuspended in
ice-cold 10 mM Tris buffer (pH 8.0), and homogenized using an overhead stirrer (Wheaton) and disposable Teflon pestles (Kontes). Homogenized samples were freeze-thawed three times, sonicated for 2 min, and centrifuged to remove cell debris. Titers of infectious virus were determined by plaque assay of BSC40 cell supernatants. The limit of detection of this assay is 2 PFU/pair of ovaries.
Quantitative PCR for PyV DNA.DNA isolation and TaqMan-based PCR were performed as described previously (25). The PyV DNA quantity is expressed in genome copies per mg of tissue and is calculated based on a standard curve of known PyV genome copy number versus threshold cycle of detection. The de-tection limit with this assay is 10 copies of genomic viral DNA.
ELISA for serum VP1 Ab.PyV major capsid protein VP1-specific enzyme-linked immunosorbent assays (ELISA) were performed as described previously (24). Twofold serial dilutions of a positive-control serum sample harvested from PyV-infected B6 mice on day 21 postinfection (p.i.) were used to obtain a standard reference curve on each 96-well plate, and the VP1-specific IgG con-centrations of the test samples were expressed in arbitrary units based on com-parison with this standard curve.
Statistics.Statistical significance was determined by an unpaired Studentttest, assuming unequal variances. AP value of⬍0.05 was considered statistically significant.
RESULTS
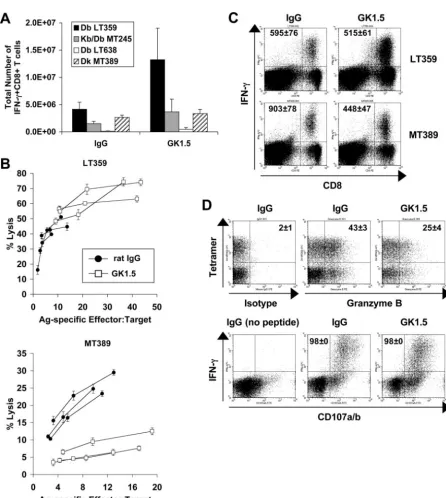
PyV-specific CD8ⴙT-cell responses in CD4ⴙT-cell-depleted B6 and C3H mice. In acutely infected B6 mice, CD4⫹T-cell depletion mediated by the administration of the MAb GK1.5 was associated with a two- to threefold increase in the number of CD8⫹T cells directed toward the dominant LT359 and subdomi-nant MT245 and LT638 PyV epitopes, such that the PyV-specific CD8⫹T-cell immunodominance hierarchy remained unchanged (Fig. 1A). In distinct contrast, CD4⫹T-cell depletion did not alter the number of splenic PyV-specific CD8⫹T cells in acutely in-fected C3H mice (Fig. 1A). Similar results were obtained by enumerating splenic DbLT359 or DkMT389 tetramer⫹CD8⫹T cells directly ex vivo (data not shown). CD4⫹T-cell depletion in naı¨ve C3H and B6 mice induced a “memory-masquerading” phe-notype (i.e., an increased frequency of cells expressing high levels of CD44 and Ly6C) by splenic CD8⫹T cells (37) and a doubling in BrdU uptake by CD8⫹T cells (data not shown). These results suggest that the homeostatic proliferation by CD8⫹T cells com-pensating for the downsized CD4⫹T-cell compartment is insuf-ficient to explain the strain-associated difference in PyV-specific CD8⫹T-cell expansion in CD4⫹T-cell-depleted mice. This con-clusion receives further support from the finding that acutely infected MHC class II⫺/⫺ B6 mice, like anti-CD4-treated B6 mice, mount an elevated PyV-specific CD8⫹T-cell response (26). CD4⫹T-cell depletion also differentially affected PyV-spe-cific CD8⫹T-cell function in B6 and C3H mice during acute infection. In C3H mice, antiviral CD8⫹T cells primed in the absence of CD4⫹ T cells manifested diminished Ag-specific cytotoxic activity; however, unhelped PyV-specific CD8⫹ T cells in B6 mice retained full cytotoxic effector capability (Fig. 1B). The defective cytotoxic T-lymphocyte activity of unhelped CD8⫹T cells in C3H mice was associated with a significant reduction (P⬍0.01) in intracellular granzyme B expression, while Ag-stimulated degranulation, as detected by upregulated surface expression of the lysosome-associated membrane pro-teins CD107a and CD107b, remained intact (Fig. 1D). Addi-tionally, although unhelped PyV-specific CD8⫹T cells in both B6 and C3H mice produced IFN-␥ex vivo upon peptide stim-ulation, the anti-IFN-␥mean fluorescence intensity (MFI) of unhelped DkMT389-specific CD8⫹T cells was twofold lower than in CD4⫹ T-cell-sufficient C3H mice (Fig. 1C). These
on November 8, 2019 by guest
http://jvi.asm.org/
FIG. 1. Effect of CD4⫹T-cell depletion on PyV-specific CD8⫹T-cell responses in acutely infected B6 and C3H mice. (A) Total numbers of splenic IFN-␥⫹CD8⫹T cells specific for DbLT359, Kb/DbMT245, and DbLT638 (B6 mice) and DkMT389 (C3H mice) during acute infection (day
7 to 8 p.i.) in MAb GK1.5- or control rat IgG-treated mice. The values plotted are the averages⫾standard deviations (SD) of the results for three mice per group and are representative of two to three independent experiments. (B) Ex vivo cytotoxicity of DbLT359- and DkMT389-specific CD8⫹ T cells was measured on day 8 and 7 p.i., respectively, in MAb GK1.5- or control rat IgG-treated mice. The Ag-specific effector:target ratio was calculated for each mouse based on the frequency of DbLT359 or DkMT389 tetramer⫹CD8⫹splenocytes. Lysis of unpulsed targets was⬍5% at all effector:target ratios measured (data not shown). Data are representative of three independent experiments. (C) The MFI of intracellular IFN-␥ staining was determined for splenic DbLT359- and DkMT389-specific CD8⫹T cells at day 7 to 8 p.i. in MAb GK1.5- or control rat IgG-treated mice. Values shown on representative dot plots are the average MFI⫾SD for three mice, and data are representative of three independent experiments. (D) The percentages of splenic DkMT389 tetramer⫹CD8⫹T cells expressing intracellular granzyme B were determined (top panels). Values shown on representative dot plots (gated on CD8⫹T cells) are the mean percentages of tetramer⫹cells that are granzyme B⫹(mean⫾ SD of the results for three mice). Dot plot quadrant gates for granzyme B are set from an IgG1 isotype control. The degranulation capacities of peptide-stimulated MT389-specific IFN-␥⫹CD8⫹T cells were assessed by CD107a/b cell surface staining (bottom panels). The values shown on representative dot plots (gated on CD8⫹T cells) are the mean percentages of IFN-␥⫹cells that are CD107a/b⫹(mean⫾SD of the results for three mice). Data are representative of two independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
findings indicate that CD4⫹T-cell help is required for the full differentiation of virus-specific CD8⫹T-cell effectors in acutely infected C3H mice but is dispensable for effector differentia-tion in B6 mice.
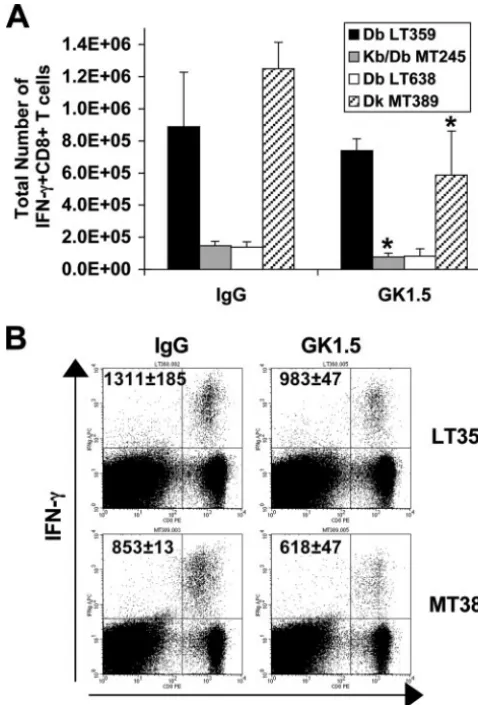
We next asked how depletion of CD4⫹T cells during acute infection impacted the antiviral memory CD8⫹T-cell response in C3H and B6 mice. Transient depletion of CD4⫹T cells in mice at the onset of infection with lymphocyte-tropic and mac-rophage-tropic variants of lymphocytic choriomeningitis virus (e.g., clone 13 or t1b) that establish a chronic infection is associated with high-level virus persistence and virus-specific CD8⫹T-cell exhaustion and deletion (33, 54). Mice received three injections of MAb GK1.5 (or rat IgG as a control) at the time of PyV inoculation, and PyV-specific CD8⫹ T-cell re-sponses were assessed at⬃day 40 p.i.; at this time, staining with CD4-specific but non-GK1.5-competing MAb RM4-5 showed that splenic CD4⫹T cells had reconstituted by 70% or 30% of their levels in rat IgG-treated groups in B6 and C3H mice, respectively (data not shown). Notably, the higher mag-nitude of the acute antiviral CD8⫹T-cell response in CD4⫹ T-cell-depleted B6 mice than in C3H mice (Fig. 1A) did not translate into an enlarged PyV-specific memory CD8⫹T-cell compartment of B6 mice (Fig. 2A). Rather, GK1.5-treated B6 mice had fewer PyV-specific CD8⫹T cells, with those specific for the MT245 epitope being significantly (P⬍0.05) reduced in magnitude compared to the numbers in persistently infected CD4⫹ T-cell-sufficient control mice (Fig. 2A). Similarly, GK1.5-treated C3H mice had significantly (P ⬍ 0.05) fewer PyV-specific memory CD8⫹ T cells than C3H mice treated with control Ab (Fig. 2A). The results of direct ex vivo enu-meration of DbLT359 or DkMT389 tetramer⫹CD8⫹T cells paralleled the numbers obtained by intracellular IFN-␥ stain-ing assays (data not shown). However, while anti-PyV CD8⫹T cells from persistently infected GK1.5-treated B6 and C3H mice produced IFN-␥directly ex vivo, the IFN-␥MFI for mice of both strains was reduced in comparison to that of control rat IgG-treated mice (Fig. 2B). Together, these data demonstrate that CD4⫹T-cell depletion of PyV-infected mice differentially affects the PyV-specific CD8⫹T-cell responses in B6 and C3H mice during acute, but not persistent, infection.
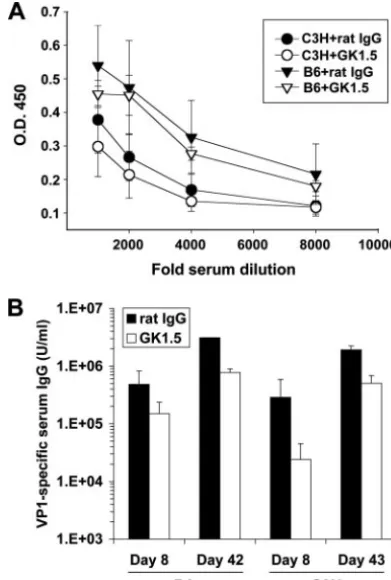
Effect of CD4ⴙ T-cell depletion on PyV-specific humoral responses in B6 and C3H mice.PyV infection elicits a neutral-izing, isotype-switched, T-cell-independent Ab response against the major viral capsid protein VP1 (44, 47). CD40⫺/⫺and T-cell-deficient B6 mice mount lower VP1-specific IgG responses than wild-type B6 mice (45), raising the question of whether PyV infection would elicit a diminished VP1-specific Ab re-sponse in B6 mice depleted of CD4⫹T cells and, given the differences between B6 and C3H mice in their anti-PyV CD8⫹ T-cell responses, whether this would occur to the same degree in both mouse strains. As shown in Fig. 3A, although C3H mice generated a lower VP1-specific IgM response than B6 mice early after infection, CD4⫹T-cell depletion did not alter VP1-specific IgM responses in mice of either strain. VP1-VP1-specific IgG titers, however, were reduced by⬃3-fold and⬃10-fold in GK1.5-treated B6 and C3H mice, respectively, by day 8 p.i. (Fig. 3B). Over time, the VP1-specific IgG titers increased in CD4⫹T-cell-depleted and control mice of both strains, fitting earlier observations that PyV-specific T-cell-independent IgG titers increase in T-cell-deficient mice between days 7 and
21 p.i (46). By day 42 to 43 p.i., the anti-VP1 titers were reduced by only⬃twofold and⬃fourfold in GK1.5-treated B6 and C3H mice, respectively (Fig. 3B). These data suggest that CD4⫹T-cell depletion incurred at the time of virus inoculation had only a modest impact on the anti-PyV humoral response in both B6 and C3H mice.
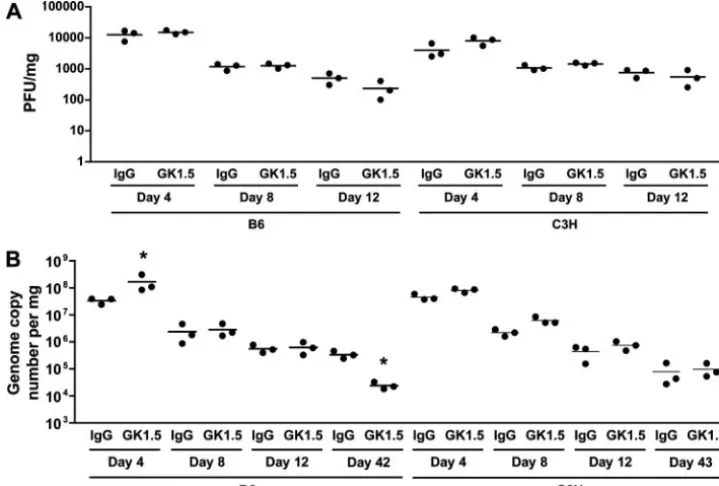
[image:4.594.303.542.70.422.2]CD4ⴙT-cell depletion minimally affects viral clearance.We next asked whether the differential effects of CD4⫹ T-cell depletion on the antiviral CD8⫹ T-cell response in B6 and C3H mice during acute infection were associated with changes in PyV clearance and persistence. During the acute phase of PyV infection, the viral loads (by infectious virus titers) (Fig. 4A), numbers of viral DNA genome copies (Fig. 4B), and
FIG. 2. CD4⫹ T-cell depletion of PyV-infected mice has similar effects on antiviral memory CD8⫹ T-cell responses in B6 and C3H mice. (A) Total numbers of splenic IFN-␥⫹CD8⫹T cells specific for DbLT359, Kb/Db MT245, and DbLT638 (B6 mice) and DkMT389
(C3H mice) during persistent infection (day 42 to 43 p.i.) in mice treated with MAb GK1.5 or control rat IgG at the onset of infection. Values plotted are the averages⫾standard deviations of the results for three mice per group and are representative of two to three inde-pendent experiments (*,P⬍0.05). (B) The MFI of intracellular IFN-␥ staining was determined for splenic DbLT359- and DkMT389-specific
CD8⫹ T cells at day 42 to 43 p.i. in MAb GK1.5- or control rat IgG-treated mice. The values shown on representative dot plots are the average MFI⫾standard deviations of the results for three mice, and the data are representative of three independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
kinetics of viral clearance were similar between GK1.5-treated and control IgG-treated mice of both strains. Importantly, GK1.5-treated B6 and C3H mice both controlled persistent viral infection as well as CD4⫹T-cell-sufficient mice (Fig. 4B). Notably, the PyV DNA in GK1.5-treated B6 mice at day 42 p.i. was reduced by⬃1 log compared to the level in control IgG-treated mice, which may be attributed to the higher-magnitude effector PyV-specific CD8⫹T-cell response during acute infec-tion lowering the set point for virus persistence. By this time p.i., infectious PyV cannot be detected by plaque assays (28). Overall, CD4⫹T-cell depletion did not substantially alter the kinetics of viral clearance or the level of PyV persistence.
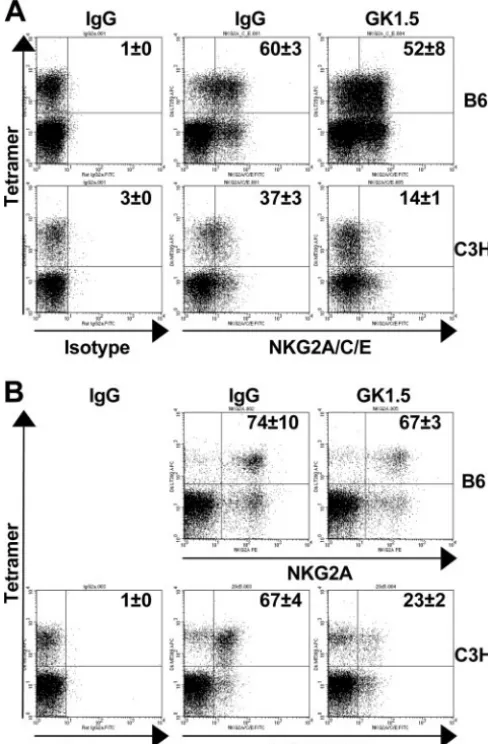
Phenotypes of unhelped PyV-specific CD8ⴙ T cells in B6 and C3H mice.Irrespective of the availability of CD4⫹T-cell help, DbLT359 and DkMT389 tetramer⫹CD8⫹T cells were CD11ahighCD62LlowCD122lowCD127high/lowin both acutely
and persistently infected mice (data not shown). Surface stain-ing with Abs to CD94/NKG2A, however, revealed marked CD4⫹T-cell-dependent differences between antiviral CD8⫹T cells in B6 and C3H mice. In both acutely and persistently infected C3H mice, substantially fewer DkMT389 tetramer⫹ CD8⫹T cells expressed CD94/NKG2A in the CD4⫹
T-cell-depleted animals than in the control Ab-treated group (Fig. 5A and B). In contrast, a similar percentage of DbLT359
tet-ramer⫹CD8⫹T cells expressed CD94/NKG2A at both stages of infection in B6 mice regardless of the presence or absence of CD4⫹T cells (Fig. 5A and B). These data suggest that the lack of CD4⫹ T cells at the onset of the antiviral CD8⫹ T-cell response impedes the differentiation of PyV-specific CD8⫹T cells in C3H, but not in B6, mice.
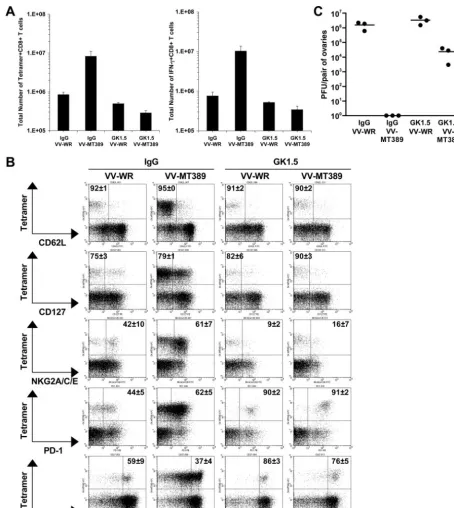
Defective recall by unhelped PyV-specific CD8ⴙT cells in persistently infected C3H mice.Starting at the time of virus inoculation, PyV-infected C3H mice received MAb GK1.5 or rat IgG weekly for 8 weeks and were then challenged by a recombinant VV encoding the MT389-397 PyV epitope (VV-MT389) or VV-WR as a control. At day 4 postchallenge, the total number of splenic MT389-specific CD8⫹T cells in mice receiving VV-WR closely approximated that in unchallenged PyV-infected animals at a similar time point p.i. (compare Fig. 6A and 2A). In comparison to the number in mice receiving the VV-WR virus, the number of DkMT389 tetramer⫹CD8⫹ T cells expanded 10-fold in rat IgG-treated C3H mice infected by VV-MT389 (Fig. 6A). Similar results were obtained by enumerating MT389-specific IFN-␥⫹CD8⫹T cells, indicating that the expanded populations of virus-specific T cells in rat IgG-treated C3H mice were functional (Fig. 6A). In contrast, in comparison to the number in mice receiving VV-WR, DkMT389 tetramer⫹ CD8⫹T cells failed to expand upon VV-MT389 challenge in CD4⫹ T-cell-depleted C3H mice (Fig. 6A).
As shown in Fig. 6B, CD4⫹ T-cell depletion also led to alterations in the expression of particular inhibitory receptors and markers of differentiation by PyV-specific CD8⫹T cells in persistently infected C3H mice. Regardless of whether the challenge infection was by VV-WR or VV-MT389, DkMT389
tetramer⫹CD8⫹T cells were predominantly CD62Llowand
CD127low. Interestingly, GK1.5 treatment was associated with
a dramatic reduction in CD94/NKG2A expression and a marked increase in PD-1 expression by MT389-specific CD8⫹ T cells. Of note, VV-MT389 infection in CD4⫹T-cell-sufficient C3H mice resulted in a pronounced loss in CD27 surface expression by DkMT389 tetramer⫹CD8⫹T cells, indicative of effector differentiation (2, 17, 32). In contrast, most of the MT389-specific CD8⫹ T cells in CD4⫹T-cell-depleted C3H mice remained CD27higheven after VV-MT389 challenge
in-fection.
We next asked if virus-specific memory CD8⫹T cells gen-erated and maintained in the presence or absence of CD4⫹T cells confer protection from VV-MT389 challenge infection. As shown in Fig. 6C, VV-MT389 was cleared from the ovaries of IgG-treated mice by day 4 postchallenge. Although the ovaries of GK1.5-treated, VV-MT389-challenged mice har-bored infectious virus, the titer was nearly 100-fold lower than in VV-WR-infected mice. That these unhelped, unexpanded, antiviral CD8⫹T cells were functionally intact was indicated by their ability to produce IFN-␥after MT389 peptide stimulation (Fig. 6A). Thus, while the lower number of unhelped virus-specific CD8⫹T cells likely accounted for the incomplete VV-MT389 clearance, these cells retained antiviral effector func-tion in vivo.
Unhelped B6C3F1 mice generate H-2k
[image:5.594.64.260.71.361.2]-restricted PyV-specific CD8ⴙT-cell effectors.Using acutely infected B6C3F1 mice, we
FIG. 3. CD4⫹T-cell depletion of PyV-infected mice diminishes the PyV-specific IgG response in both B6 and C3H mice. (A) VP1-specific serum IgM in MAb GK1.5- or control rat IgG-treated B6 and C3H mice on day 4 p.i., as determined by ELISA. Values plotted are the average optical densities⫾standard errors for three mice per group. O.D. 450, optical density at 450 nm. (B) VP1-specific serum IgG titers during acute (day 7 to 8 p.i.) and persistent (day 42 to 43 p.i.) phases of infection in MAb GK1.5- or control rat IgG-treated B6 and C3H mice, expressed in arbitrary units. The values plotted are the averages ⫾standard deviations of the results for three mice per group.
on November 8, 2019 by guest
http://jvi.asm.org/
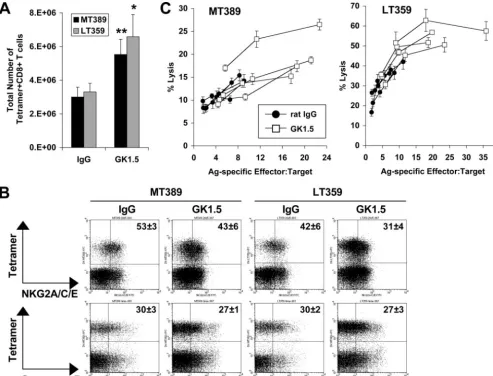
concurrently compared the functions and phenotypes of the DkMT389 and DbLT359 CD8⫹T-cell responses in the absence of CD4⫹ T-cell help. As shown in Fig. 7A, GK1.5-treated B6C3F1 mice mounted significantly higher-magnitude T-cell responses of both specificities (which were codominant) than their control Ab-treated counterparts. Similar sizeable propor-tions of unhelped and helped DkMT389 and DbLT359
tet-ramer⫹CD8⫹T cells expressed CD94/NKG2A and granzyme B (Fig. 7B). Both unhelped DkMT389 and DbLT359 CD8⫹T cells in B6C3F1 mice were also functionally intact, based on ex vivo Ag-specific cytotoxicity (Fig. 7C) and high intracellular anti-IFN-␥MFI after peptide stimulation (data not shown). Thus, the functionally defective and phenotypically less-differ-entiated PyV-specific CD8⫹T-cell response in CD4⫹ T-cell-depleted C3H mice was corrected when crossed with B6 mice.
DISCUSSION
In this study, we describe an unexpected allogeneic differ-ence in the contribution of CD4⫹T-cell help to the quality of the PyV-specific CD8⫹ T-cell response. CD4⫹ T cells were found to be dispensable for the priming and expansion of antiviral CD8⫹T cells during acute infection in both B6 and C3H mice, but only in B6 mice did PyV-specific CD8⫹T cells fully differentiate to effectors. Unhelped PyV-specific CD8⫹T cells in C3H mice exhibited defective Ag-specific recall and alterations in phenotype compared to the Ag-specific recall and phenotype in CD4⫹ T-cell-sufficient C3H mice. These allogeneic differences in antiviral CD8⫹T-cell phenotype and function could not be attributed to differences in the abilities of B6 and C3H mice to control PyV infection. Moreover, CD4⫹ T-cell-depleted B6 and C3H mice mounted similar Ab
re-sponses to PyV. Using CD4⫹T-cell-depleted B6C3F1 mice, we further determined that the B6 background could compensate for the defect in effector differentiation of H-2k-restricted
PyV-specific CD8⫹T cells. This study highlights a generally under-appreciated consideration of allogeneic differences in the re-quirements for generating optimal CD8⫹T-cell responses to viral infection.
In CD4⫹T-cell-deficient C3H mice, we observed a dissoci-ation between cytokine and cytotoxic activities by PyV-specific effector CD8⫹ T cells. Because these unhelped PyV-specific CD8⫹T cells produced IFN-␥and degranulated upon cognate epitope stimulation, the absence of CD4⫹T-cell help did not incur a global defect in T-cell-receptor activation. This disso-ciation between effector functions is reminiscent of our previ-ous finding that antiviral CD8⫹T cells in acutely infected C3H mice express the inhibitory CD94/NKG2A receptor, which dampens cytotoxic activity without affecting IFN-␥ functional-ity (36). In addition, CD8⫹T-cell multifunctionality may be associated with the T-cell differentiation state or the duration of Ag encounter (42, 51). Because antiviral CD8⫹T cells retain full cytotoxic effector potential in CD4⫹T-cell-sufficient C3H mice, we had proposed that the CD94/NKG2A receptor oper-ated to restrain excessive cytotoxic T-lymphocyte-medioper-ated cytopathology as virus infection levels declined during acute infection. However, CD4⫹T-cell depletion in C3H mice de-creased the proportion of PyV-specific CD8⫹T cells express-ing this inhibitory receptor. Because these T cells express lower levels of granzyme B, it is tempting to speculate that CD94/ NKG2A-mediated inhibition is superfluous in a situation where T cells are intrinsically deficient in cytotoxic effector capability. CD94/NKG2A has also been included among the surface phenotypic markers used to gauge CD8⫹T-cell
differ-FIG. 4. Comparison of viral clearance in CD4⫹T-cell-depleted B6 and C3H mice. (A) The viral titers in the spleens of B6 and C3H mice given MAb GK1.5 or control rat IgG were determined at the indicated day p.i. (B) The numbers of PyV genomes in the spleens were quantitated by TaqMan real-time PCR at the indicated day p.i. Each point represents an individual mouse, and the bars indicate the means (*,P⬍0.05).
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.594.111.471.71.314.2]entiation status (30, 34). It merits pointing out, though, that while they are IFN-␥effector competent, unhelped antiviral CD8⫹T cells produce a reduced level of IFN-␥following Ag stimulation. From these observations, we favor the interpreta-tion that CD4⫹ T-cell insufficiency in C3H mice creates a priming environment that provides suboptimal signals for full T-cell effector differentiation.
That CD4⫹T-cell deficiency negatively impacts PyV-specific CD8⫹T-cell differentiation in C3H mice is also supported by the failure of CD27 to downregulate following Ag-specific challenge infection. For example, human cytomegalovirus-spe-cific CD8⫹T cells decrease their surface expression of CD27 upon virus reactivation, which is dependent on the interaction of CD27 with its ligand CD70 (21). Recently, Marzo et al.
reported that CD27 surface expression by virus-specific mem-ory CD8⫹ T cells is associated with diminished granzyme B expression and reduced cytotoxic effector capability, and they proposed that CD27 and granzyme B, in addition to CD62L, should be used to define functionally and anatomically distinct subpopulations of memory CD8⫹T cells (32). Although tran-sient depletion of CD4⫹T cells at the onset of acute infection in B6 and C3H mice failed to reveal an allogeneic difference in the functional competence of memory PyV-specific CD8⫹T cells (based on IFN-␥production; Fig. 2), continuous CD4⫹ T-cell depletion in C3H mice showed that unhelped memory antiviral CD8⫹T cells were markedly impaired in their recall response (Fig. 6A). In sharp contrast, we previously demon-strated that the memory PyV-specific CD8⫹T cells in MHC class II-deficient B6 mice are fully capable of responding to Ag challenge (26). A lack of CD4⫹T cells in C3H mice appears to deprive the predominantly CD62Llow PyV-specific memory
CD8⫹T cells of the plasticity needed to respond to Ag chal-lenge.
We previously reported that DkMT389-specific CD8⫹ T cells primed in the face of CD28 and CD40L costimulation blockade do not upregulate CD94/NKG2A (24). CD4⫹T-cell depletion was also associated with reduced CD94/NKG2A ex-pression by memory MT389-specific CD8⫹T cells (Fig. 5B), and few memory MT389-specific CD8⫹ T cells expressed CD94/NKG2A in persistently infected CD4⫹T-cell-deficient mice (Fig. 6B). Byers et al. previously reported that CD94/ NKG2A-expressing memory MT389-specific CD8⫹ T cells preferentially proliferate during persistent infection and fol-lowing Ag rechallenge, whereas memory MT389-specific CD8⫹T cells that do not express CD94/NKG2A proliferate poorly (9). Our present finding that unhelped memory MT389-specific CD8⫹T cells (which do not express CD94/NKG2A) do not expand following Ag challenge provides additional support for the idea that CD94/NKG2A identifies a subpopulation of proliferation-competent antiviral memory CD8⫹T cells.
[image:7.594.42.286.72.444.2]We and others have shown that inflammation is detrimental to the differentiation of Ag-specific effector and memory CD8⫹T cells (1, 3, 4). Recently, Badovinac and Harty sug-gested that the degree of inflammation elicited by microbial infection is directly proportional to the duration and magni-tude of the effector phase of the CD8⫹T-cell response (3). In this context, elevated inflammation incurred by higher levels of infection would tip the balance from memory to effector T-cell differentiation in order to favor pathogen clearance at the expense of durable immunity. In support of this possibility, influenza virus-specific CD8⫹T cells appear to reach a more focused stage of effector differentiation in the infected lung than in the spleen, which does not support influenza virus infection (31). Perhaps CD4⫹T cells promote virus-specific CD8⫹effector differentiation by increasing or altering the pro-file of proinflammatory mediators secreted by innate immune cells (12, 13, 38). In the absence of CD4⫹T cells, the character of virus-associated inflammation may change, with a conse-quent shift away from effector differentiation. In this context, the different outcomes of CD4⫹T-cell depletion for anti-PyV CD8⫹T-cell effector status between B6 and C3H mice might reflect allogeneic differences in the profiles of CD4⫹ T-cell-induced inflammatory mediators that may provide signal 3 for CD8⫹T-cell differentiation (14–16).
FIG. 5. CD4⫹T-cell depletion differentially affects the expression of CD94/NKG2A by PyV-specific CD8⫹T cells in B6 and C3H mice. CD94/NKG2A expression on splenic DbLT359 and DkMT389
tet-ramer⫹CD8⫹T cells in MAb GK1.5- or control rat IgG-treated mice was evaluated during the acute (A) (day 7 to 8 p.i.) and persistent (B) (day 42 to 43 p.i.) phases of infection. Dot plot quadrant gates are set from IgG2a isotype controls. The values shown on the representa-tive dot plots (gated on CD8⫹T cells) are the mean percentages of tetramer⫹ cells that are CD94/NKG2A⫹ (means ⫾standard devia-tions of the results for three mice), and the data are representative of two independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
FIG. 6. PyV-specific memory CD8⫹ T cells in CD4⫹ T-cell-depleted C3H mice fail to expand and are phenotypically altered following Ag-specific rechallenge. Persistently PyV-infected C3H mice (treated weekly with MAb GK1.5 or control rat IgG from the onset of PyV infection) were rechallenged on day 54 p.i. with a recombinant VV encoding the MT389-397 epitope (VV-MT389) or with the control, VV-WR, and T-cell responses were analyzed 4 days later. (A) Total numbers of splenic DkMT389 tetramer⫹CD8⫹T cells and MT389-specific IFN-␥⫹CD8⫹T cells. The values plotted are the means⫾standard deviations of the results for three mice per group. (B) Percentages of DkMT389 tetramer⫹CD8⫹ T cells in the spleen expressing CD62L, CD127, CD94/NKG2A, PD-1, or CD27. The values shown on the representative dot plots (gated on CD8⫹ T cells) are the mean percentages of tetramer⫹cells that are CD62Llow, CD127low, NKG2A⫹, PD-1⫹, or CD27high(means⫾standard deviations
of the results for three mice). Dot plot quadrant gates for NKG2A and PD-1 are set from IgG2a and IgG2b isotype controls, respectively. (C) VV titers in ovaries of MAb GK1.5- or rat IgG-treated C3H mice at day 4 after VV-MT389 or VV-WR challenge infection. Each point represents an individual mouse, and the bars indicate the means. No VV was detected in the ovaries of IgG-treated, VV-MT389-rechallenged mice. All data are representative of two independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
Interestingly, the CD4⫹T-cell dependence for PyV-specific CD8⫹T-cell effector differentiation in C3H mice was overrid-den when these mice were crossed with B6 mice. In sharp contrast to PyV-infected C3H mice, helped and unhelped DkMT389-specific effector CD8⫹T cells generated in B6C3F1 mice were phenotypically and functionally indistinguishable. It is also notable that the LT359-specific and MT389-specific CD8⫹ T-cell responses, which are immunodominant in the respective B6 and C3H mouse strains, are codominant re-sponses in the F1mice. By simultaneously analyzing D
kMT389
and DbLT359 CD8⫹ T-cell responses in these F
1 mice, we
ruled out Ag specificity as an explanation for the defect in effector differentiation in CD4⫹T-cell-depleted C3H mice. In addition, B6C3F1 mice depleted of CD4⫹T cells at the time of
PyV inoculation mounted higher-magnitude responses of both MT389-specific and LT359-specific CD8⫹T cells than CD4⫹ T-cell sufficient mice; of these two specificities, only the LT359-specific CD8⫹ T-cell response is elevated in CD4⫹ T-cell-depleted B6 mice (a result recapitulated in MHC class II-deficient B6 mice [26]). Using peptide-loaded MHC class II-deficient den-dritic cells, Frelinger and colleagues demonstrated that the induc-tion of Ag-specific CD8⫹T cells, if present in sufficient magni-tude, can bypass the need for CD4⫹T-cell help (50). Because the H-2b-restricted and H-2k-restricted PyV-specific CD8⫹T-cell responses in B6C3F1 mice are approximately the summation of the anti-PyV CD8⫹ T-cell responses in the parental mice (compare Fig. 1A and 7A), CD8⫹ T-cell self-help offers a plausible explanation for the CD4⫹T-cell-independent
differ-FIG. 7. The B6 phenotype is dominant to the unhelped PyV-specific CD8⫹T-cell phenotype of C3H mice. (A) Total numbers of splenic DkMT389 and DbLT359 tetramer⫹CD8⫹T cells at day 8 p.i. in MAb GK1.5- or control rat IgG-treated B6C3F1 mice. Values plotted are the averages⫾standard deviations of the results for four mice per group (*,P⬍0.05;**,P⬍0.01). (B) The percentages of splenic DkMT389 or
DbLT359 tetramer⫹CD8⫹T cells that expressed CD94/NKG2A or granzyme B were determined by flow cytometry. The values shown on the representative dot plots (gated on CD8⫹T cells) are the mean percentages of tetramer⫹cells that are NKG2A⫹or granzyme B⫹(mean⫾standard deviation of the results for four mice). Dot plot quadrant gates for NKG2A and granzyme B are set from IgG2a and IgG1 isotype controls, respectively. (C) Ex vivo cytotoxicity of DkMT389- and DbLT359-specific CD8⫹T cells was measured at day 8 p.i. in MAb GK1.5- or control rat IgG-treated mice. The Ag-specific effector:target ratio was calculated for each mouse based on the frequency of DkMT389 or DbLT359 tetramer⫹ CD8⫹splenocytes. Lysis of unpulsed AG104A or RMA/S target cells was⬍5% and⬍15%, respectively, at all effector:target ratios measured (data not shown). All data are representative of two independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:9.594.45.539.70.446.2]entiation of DkMT389 CD8⫹T cells in these F
1 mice.
Com-pared to dendritic cell-based immunization, though, viral in-fection elicits a strong host inflammatory response. Thus, the inflammatory mediators induced by PyV infection in the CD4⫹ T-cell-depleted B6C3F1 mice may be more “B6-like” than “C3H-like” and provide a microenvironment conducive to ef-fector differentiation in the absence of CD4⫹T-cell help. Us-ing recombinant inbred mice, Williams et al. showed that mul-tiple genetic loci determined the differences between B6 and C3H mice in susceptibility to CD40L and CD28 T-cell costimu-lation blockade for skin allograft survival (52). The identi-ty(ies) of the genetic locus(i) in B6 and C3H mice that confers CD4⫹T-cell-independent PyV-specific CD8⫹T-cell responses remains to be determined.
We are just beginning to appreciate that the contribution of CD4⫹ T-cell help to the quality of the antiviral CD8⫹ T-cell response varies among different inbred strains of mice. Ramsburg et al. recently demonstrated that recall responses by vesicular stomatitis virus-specific memory CD8⫹ T cells are impaired in CD4⫺/⫺ BALB/c mice, but not in CD4⫺/⫺ or MHC class II-deficient C57BL/6 mice (39). Our findings are in line with this allogeneic variability and further demonstrate that mice of differ-ent inbred strains can differ in their dependence on CD4⫹T cells in driving optimal antiviral CD8⫹T-cell differentiation and effec-tor function.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grants R01CA71971 and R01CA100644 (to A.E.L.) and R01CA66644 (to E.S.-T.) from the National Cancer Institute.
We thank Annette Hadley for expert technical assistance. We also thank Christopher Pack and Devon Livingston-Rosanoff for reviewing the manuscript.
REFERENCES
1.Andrews, N. P., C. D. Pack, V. Vezys, G. N. Barber, and A. E. Lukacher.2007. Early virus-associated bystander events affect the fitness of the CD8 T cell response to persistent virus infection. J. Immunol.178:7267–7275. 2.Baars, P. A., S. Sierro, R. Arens, K. Tesselaar, B. Hooibrink, P. Klenerman,
and R. A. van Lier.2005. Properties of murine CD8⫹CD27⫺T cells. Eur. J. Immunol.35:3131–3141.
3.Badovinac, V. P., and J. T. Harty.2007. Manipulating the rate of memory CD8⫹T cell generation after acute infection. J. Immunol.179:53–63. 4.Badovinac, V. P., K. A. Messingham, A. Jabbari, J. S. Haring, and J. T.
Harty.2005. Accelerated CD8⫹T-cell memory and prime-boost response after dendritic-cell vaccination. Nat. Med.11:748–756.
5.Belz, G. T., D. Wodarz, G. Diaz, M. A. Nowak, and P. C. Doherty.2002. Compromised influenza virus-specific CD8⫹-T-cell memory in CD4⫹ -T-cell-deficient mice. J. Virol.76:12388–12393.
6.Betts, M. R., J. M. Brenchley, D. A. Price, S. C. De Rosa, D. C. Douek, M. Roederer, and R. A. Koup.2003. Sensitive and viable identification of anti-gen-specific CD8⫹T cells by a flow cytometric assay for degranulation. J. Immunol. Methods281:65–78.
7.Bevan, M. J.2004. Helping the CD8⫹T-cell response. Nat. Rev. Immunol. 4:595–602.
8.Bourgeois, C., B. Rocha, and C. Tanchot.2002. A role for CD40 expression on CD8⫹T cells in the generation of CD8⫹T cell memory. Science297: 2060–2063.
9.Byers, A. M., N. P. Andrews, and A. E. Lukacher.2006. CD94/NKG2A expression is associated with proliferative potential of CD8 T cells during persistent polyoma virus infection. J. Immunol.176:6121–6129.
10.Byers, A. M., A. Hadley, and A. E. Lukacher. 2007. Protection against polyoma virus-induced tumors is perforin-independent. Virology358:485– 492.
11.Cardin, R. D., J. W. Brooks, S. R. Sarawar, and P. C. Doherty.1996. Progressive loss of CD8⫹T cell-mediated control of a␥-herpesvirus in the absence of CD4⫹T cells. J. Exp. Med.184:863–871.
12.Castellino, F., and R. N. Germain.2007. Chemokine-guided CD4⫹T cell help enhances generation of IL-6R␣highIL-7R␣high prememory CD8⫹T
cells. J. Immunol.178:778–787.
13.Castellino, F., and R. N. Germain.2006. Cooperation between CD4⫹and CD8⫹T cells: when, where, and how. Annu. Rev. Immunol.24:519–540. 14.Curtsinger, J. M., C. M. Johnson, and M. F. Mescher.2003. CD8 T cell
clonal expansion and development of effector function require prolonged exposure to antigen, costimulation, and signal 3 cytokine. J. Immunol.171: 5165–5171.
15.Curtsinger, J. M., D. C. Lins, C. M. Johnson, and M. F. Mescher.2005. Signal 3 tolerant CD8 T cells degranulate in response to antigen but lack granzyme B to mediate cytolysis. J. Immunol.175:4392–4399.
16.Curtsinger, J. M., D. C. Lins, and M. F. Mescher.2003. Signal 3 determines tolerance versus full activation of naive CD8 T cells: dissociating prolifera-tion and development of effector funcprolifera-tion. J. Exp. Med.197:1141–1151. 17.de Bree, G. J., E. M. van Leeuwen, T. A. Out, H. M. Jansen, R. E. Jonkers,
and R. A. van Lier.2005. Selective accumulation of differentiated CD8⫹T cells specific for respiratory viruses in the human lung. J. Exp. Med.202: 1433–1442.
18.Deng, Y., J. W. Yewdell, L. C. Eisenlohr, and J. R. Bennink.1997. MHC affinity, peptide liberation, T cell repertoire, and immunodominance all contribute to the paucity of MHC class I-restricted peptides recognized by antiviral CTL. J. Immunol.158:1507–1515.
19.Drake, D. R., III, and A. E. Lukacher.1998.2-Microglobulin knockout mice
are highly susceptible to polyoma virus tumorigenesis. Virology252:275–284. 20.Fuller, M. J., A. Khanolkar, A. E. Tebo, and A. J. Zajac.2004. Maintenance, loss, and resurgence of T cell responses during acute, protracted, and chronic viral infections. J. Immunol.172:4204–4214.
21.Gamadia, L. E., E. M. van Leeuwen, E. B. Remmerswaal, S. L. Yong, S. Surachno, P. M. Wertheim-van Dillen, I. J. Ten Berge, and R. A. Van Lier. 2004. The size and phenotype of virus-specific T cell populations is deter-mined by repetitive antigenic stimulation and environmental cytokines. J. Immunol.172:6107–6114.
22.Gunturi, A., R. E. Berg, E. Crossley, S. Murray, and J. Forman.2005. The role of TCR stimulation and TGF-in controlling the expression of CD94/ NKG2A receptors on CD8 T cells. Eur. J. Immunol.35:766–775. 23.Janssen, E. M., E. E. Lemmens, T. Wolfe, U. Christen, M. G. von Herrath,
and S. P. Schoenberger.2003. CD4⫹T cells are required for secondary expansion and memory in CD8⫹T lymphocytes. Nature421:852–856. 24.Kemball, C. C., E. D. Lee, E. Szomolanyi-Tsuda, T. C. Pearson, C. P. Larsen,
and A. E. Lukacher.2006. Costimulation requirements for antiviral CD8⫹T cells differ for acute and persistent phases of polyoma virus infection. J. Im-munol.176:1814–1824.
25.Kemball, C. C., E. D. Lee, V. Vezys, T. C. Pearson, C. P. Larsen, and A. E. Lukacher.2005. Late priming and variability of epitope-specific CD8⫹T cell responses during a persistent virus infection. J. Immunol.174:7950–7960. 26.Kemball, C. C., C. D. Pack, H. M. Guay, Z. N. Li, D. A. Steinhauer, E.
Szomolanyi-Tsuda, and A. E. Lukacher.2007. The antiviral CD8⫹T cell response is differentially dependent on CD4⫹T cell help over the course of persistent infection. J. Immunol.179:1113–1121.
27.Kristensen, N. N., J. P. Christensen, and A. R. Thomsen.2002. High num-bers of IL-2-producing CD8⫹T cells during viral infection: correlation with stable memory development. J. Gen. Virol.83:2123–2133.
28.Lukacher, A. E., J. M. Moser, A. Hadley, and J. D. Altman.1999. Visual-ization of polyoma virus-specific CD8⫹T cells in vivo during infection and tumor rejection. J. Immunol.163:3369–3378.
29.Lukacher, A. E., and C. S. Wilson. 1998. Resistance to polyoma virus-induced tumors correlates with CTL recognition of an immunodominant H-2Dk-restricted epitope in the middle T protein. J. Immunol.160:1724–
1734.
30.Maris, C. H., J. D. Miller, J. D. Altman, and J. Jacob.2003. A transgenic mouse model genetically tags all activated CD8 T cells. J. Immunol.171: 2393–2401.
31.Marshall, D. R., E. Olivas, S. Andreansky, N. L. La Gruta, G. A. Neale, A. Gutierrez, D. G. Wichlan, S. Wingo, C. Cheng, P. C. Doherty, and S. J. Turner.2005. Effector CD8⫹T cells recovered from an influenza pneumo-nia differentiate to a state of focused gene expression. Proc. Natl. Acad. Sci. USA102:6074–6079.
32.Marzo, A. L., H. Yagita, and L. Lefrancois.2007. Cutting edge: migration to nonlymphoid tissues results in functional conversion of central to effector memory CD8 T cells. J. Immunol.179:36–40.
33.Matloubian, M., R. J. Concepcion, and R. Ahmed.1994. CD4⫹T cells are required to sustain CD8⫹cytotoxic T-cell responses during chronic viral infection. J. Virol.68:8056–8063.
34.Miller, J. D., M. Peters, A. E. Oran, G. W. Beresford, L. Harrington, J. M. Boss, and J. D. Altman.2002. CD94/NKG2 expression does not inhibit cytotoxic function of lymphocytic choriomeningitis virus-specific CD8⫹T cells. J. Immunol.169:693–701.
35.Moser, J. M., J. D. Altman, and A. E. Lukacher.2001. Antiviral CD8⫹T cell responses in neonatal mice: susceptibility to polyoma virus-induced tumors is associated with lack of cytotoxic function by viral antigen-specific T cells. J. Exp. Med.193:595–606.
36.Moser, J. M., J. Gibbs, P. E. Jensen, and A. E. Lukacher.2002. CD94-NKG2A receptors regulate antiviral CD8⫹T cell responses. Nat. Immunol. 3:189–195.
on November 8, 2019 by guest
http://jvi.asm.org/
37.Murali-Krishna, K., and R. Ahmed.2000. Cutting edge: naive T cells mas-querading as memory cells. J. Immunol.165:1733–1737.
38.Pardoll, D. M., and S. L. Topalian.1998. The role of CD4⫹T cell responses in antitumor immunity. Curr. Opin. Immunol.10:588–594.
39.Ramsburg, E. A., J. M. Publicover, D. Coppock, and J. K. Rose.2007. Requirement for CD4 T cell help in maintenance of memory CD8 T cell responses is epitope dependent. J. Immunol.178:6350–6358.
40.Rowe, W. P. 1961. The epidemiology of mouse polyoma virus infection. Bacteriol. Rev.25:18–31.
41.Shedlock, D. J., and H. Shen.2003. Requirement for CD4 T cell help in generating functional CD8 T cell memory. Science300:337–339. 42.Slifka, M. K., and J. L. Whitton.2000. Activated and memory CD8⫹T cells
can be distinguished by their cytokine profiles and phenotypic markers. J. Immunol.164:208–216.
43.Sun, J. C., and M. J. Bevan.2003. Defective CD8 T cell memory following acute infection without CD4 T cell help. Science300:339–342.
44.Szomolanyi-Tsuda, E., J. D. Brien, J. E. Dorgan, R. L. Garcea, R. T. Wood-land, and R. M. Welsh.2001. Antiviral T-cell-independent type 2 antibody responses induced in vivo in the absence of T and NK cells. Virology280: 160–168.
45.Szomolanyi-Tsuda, E., J. D. Brien, J. E. Dorgan, R. M. Welsh, and R. L. Garcea.2000. The role of CD40-CD154 interaction in antiviral T cell-inde-pendent IgG responses. J. Immunol.164:5877–5882.
46.Szomolanyi-Tsuda, E., Q. P. Le, R. L. Garcea, and R. M. Welsh.1998. T-cell-independent immunoglobulin G responses in vivo are elicited by live-virus infection but not by immunization with viral proteins or live-virus-like particles. J. Virol.72:6665–6670.
47.Szomolanyi-Tsuda, E., and R. M. Welsh.1996. T cell-independent antibody-mediated clearance of polyoma virus in T cell-deficient mice. J. Exp. Med. 183:403–411.
48.Vance, R. E., A. M. Jamieson, D. Cado, and D. H. Raulet.2002. Implications of CD94 deficiency and monoallelic NKG2A expression for natural killer cell development and repertoire formation. Proc. Natl. Acad. Sci. USA99:868– 873.
49.Vezys, V., D. Masopust, C. C. Kemball, D. L. Barber, L. A. O’Mara, C. P. Larsen, T. C. Pearson, R. Ahmed, and A. E. Lukacher.2006. Continuous recruitment of naive T cells contributes to heterogeneity of antiviral CD8 T cells during persistent infection. J. Exp. Med.203:2263–2269.
50.Wang, B., C. C. Norbury, R. Greenwood, J. R. Bennink, J. W. Yewdell, and J. A. Frelinger.2001. Multiple paths for activation of naive CD8⫹T cells: CD4-independent help. J. Immunol.167:1283–1289.
51.Welsh, R. M.2001. Assessing CD8 T cell number and dysfunction in the presence of antigen. J. Exp. Med.193:F19–F22.
52.Williams, M. A., J. Trambley, J. Ha, A. B. Adams, M. M. Durham, P. Rees, S. R. Cowan, T. C. Pearson, and C. P. Larsen.2000. Genetic characterization of strain differences in the ability to mediate CD40/CD28-independent re-jection of skin allografts. J. Immunol.165:6849–6857.
53.Wojtasiak, M., C. M. Jones, L. C. Sullivan, A. C. Winterhalter, F. R. Carbone, and A. G. Brooks.2004. Persistent expression of CD94/NKG2 receptors by virus-specific CD8 T cells is initiated by TCR-mediated signals. Int. Immunol.16:1333–1341. 54.Zajac, A. J., J. N. Blattman, K. Murali-Krishna, D. J. Sourdive, M. Suresh,
J. D. Altman, and R. Ahmed.1998. Viral immune evasion due to persistence of activated T cells without effector function. J. Exp. Med.188:2205–2213.