JOURNAL OFVIROLOGY, Oct. 2011, p. 10201–10212 Vol. 85, No. 19 0022-538X/11/$12.00 doi:10.1128/JVI.00062-11
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Plasmid Vector-Linked Maturation of Natural Killer (NK) Cells Is
Coupled to Antigen-Dependent NK Cell Activation during
DNA-Based Immunization in Mice
䌤
Ren Zhu,
1,2‡ Maryline Mancini-Bourgine,
1,2Xiao Ming Zhang,
3,4Florence Bayard,
1,2Qiang Deng,
1,2† and Marie-Louise Michel
1,2*
Institut Pasteur, Laboratoire de Pathogene`se des Virus de l’He´patite B, De´partement de Virologie, Paris, France1; INSERM U845,
Centre de Recherche, Croissance et Signalisation, Faculte´ de Me´decine Paris Descartes, Paris, France2; Institut Pasteur,
Unite´ de Recherche de Re´gulation Immunitaire et Vaccinologie, De´partement d’Immunologie, Paris,
France3; and INSERM U883, Institut Pasteur, Paris, France4
Received 11 January 2011/Accepted 7 July 2011
Plasmid DNA vaccines serve in a wide array of applications ranging from prophylactic vaccines to potential therapeutic tools against infectious diseases and cancer. In this study, we analyzed the mechanisms underlying the activation of natural killer (NK) cells and their potential role in adaptive immunity during DNA-based immunization against hepatitis B virus surface antigen in mice. We observed that the mature Mac-1ⴙCD27ⴚ NK cell subset increased in the liver of mice early after DNA injection, whereas the number of the less mature Mac-1ⴙCD27ⴙNK cells in the liver and spleen was significantly reduced. This effect was attributed to bacterial sequences present in the plasmid backbone rather than to the encoded antigen and was not observed in immunized MyD88-deficient mice. The activation of NK cells by plasmid-DNA injection was associated with an increase in their effector functions that depended on the expressed antigen. Maturation of NK cells was abrogated in the absence of T cells, suggesting that cross talk exists between NK cells and antigen-specific T cells. Taken together, our data unravel the mechanics of plasmid vector-induced maturation of NK cells and plasmid-encoded antigen-dependent activation of NK cells required for a crucial role of NK cells in DNA vaccine-induced immunogenicity.
Natural killer (NK) cells are defined by the absence of CD3 and by the expression of CD56 in humans or NK1.1 in mice and are commonly considered the first immunological lympho-cyte defense against pathogens or tumors (9). NK cells have a heterogeneous repertoire, location, function, and developmen-tal origin. They mediate their action by remarkable developmen-talents of cellular cytotoxicity as well as by secretion of a range of pro-inflammatory cytokines/chemokines, such as gamma interferon (IFN-␥), tumor necrosis factor alpha (TNF-␣), macrophage inflammatory protein (MIP), and RANTES. NK cells exert immunoregulatory effects on other immune cells, such as T, B, and dendritic cells (DC), through soluble factors and/or cell-to-cell contact during immune responses. As such, NK cells form the bridge between innate and adaptive immunity (2, 41). They express a wide variety of receptors on their surface, and the balance of activating/inhibitory receptors governs the acti-vation of this innate lymphocyte population (27). Recently, distinct NK cell subsets were characterized in humans and mice (13). In humans, two subsets of NK cells have been identified according to CD56 expression levels. CD56bright
CD16⫺NK cells produce a large quantity of cytokines, such as IFN-␥, but are poorly cytolytic. In contrast, CD56dimNK cells
display potential cytolytic function but produce less cytokines (11), although this dichotomy was recently disputed (16). Based on Mac-1 and CD27 (a TNF receptor family member) expression, similar NK subsets were suggested to be present in mice (20). It has been shown that NK cell maturation in mice is a 4-stage process that starts from a Mac-1⫺CD27⫺stage and progresses: Mac-1⫺ CD27⫹ 3 Mac-1⫹ CD27⫹ 3
Mac-1⫹CD27⫺(10). Mac-1⫹CD27⫹NK cells are consid-ered not only powerful cytotoxic cells but also potent cyto-kine producers, compared to the more mature Mac-1⫹ CD27⫺NK cell subset (19).
NK cells are implicated as playing a crucial role in host resistance to bacteria, parasites, and viruses (6). Chronic in-fection with hepatitis B virus (HBV) is generally considered an immune-mediated disease, and viral persistence probably re-sults from an inefficient immune response characterized by marked dysfunctional TNF-␣ and IFN-␥ production by NK cells (8, 43) and by the hyporesponsiveness of HBV-specific T cells during HBV infection (32). For example, the percentage of NK cells was dramatically reduced in peripheral blood mononuclear cells (PBMCs) from chronically HBV-infected patients (53). In the presence of certain cytokines, such as interleukin-8 (IL-8) and IFN-␣, NK cells can also induce liver inflammation through the TRAIL pathway in chronic HBV-infected patients (14). NK cells also play an important role in the early control of virus replication during HBV infection (15, 17), revealing the functional diversity of NK cells during HBV infection in humans. In addition to antiviral therapies, im-* Corresponding author. Mailing address: Laboratoire de
Patho-gene`se des Virus de l’He´patite B, Institut Pasteur and INSERM U845, 28 rue du Docteur Roux, 75724 Paris CEDEX 15, France. Phone: 33 1 45 68 88 49. Fax: 33 1 40 61 38 41. E-mail: marie-louise [email protected].
† Present address: Unit of Tumor Virology, Institut Pasteur of Shanghai, 200025 Shanghai, China.
‡ Present address: Transgene SA, 67405 Illkirch Graffenstaden, France.
䌤Published ahead of print on 20 July 2011.
10201
on November 7, 2019 by guest
http://jvi.asm.org/
mune-based treatments, such as therapeutic vaccination, could be a way to reactivate efficient immune responses and control viral replication during chronic infection. We previously dem-onstrated that pCMVS2.S, an HBV-based DNA vaccine en-coding small and middle HBV envelope proteins, elicited and reactivated HBV-specific T cell responses in HBV-infected patients (37). This plasmid DNA vaccine also activated innate cells, such as CD56brightNK cells, in chronic HBV patients.
Interestingly, the activation of NK cells correlated with vac-cine-activated T cell responses (48).
Plasmid DNA contains immunostimulatory CpG motifs con-sidered a kind of “built-in adjuvant” for DNA vaccines through recognition of the Toll-like receptor 9 (TLR9) signaling path-way (38). However, it was recently suggested that the double-stranded structure of DNA can stimulate the innate immune system independently of TLRs, by signaling through nonca-nonical IB kinase (IKK) and TANK-binding kinase 1 (TBK1) (23).
Little is known about the molecular mechanisms underlying the activation of NK cells during DNA-based immunization. Therefore, we addressed this issue in an immuno-competent murine model injected with an HBV-based DNA plasmid. We show that DNA immunization promotes NK cell activation and homeostasis in the liver, through TLR/IFN-␣-mediated signal-ing pathways. HBV-plasmid DNA injection enhanced NK cell effector functions, including cellular cytotoxicity and IFN-␥ production, in an antigen-dependent fashion. Our work pro-vides evidence for the involvement of NK cells in DNA-based immunization, and this may further be exploited for therapeu-tic vaccine approaches.
MATERIALS AND METHODS
Mice. Seven- to 14-week-old wild-type, MyD88-deficient (MyD88⫺/⫺),
CD3ε⫺/⫺mutant (44), and IFN-␣ receptor-deficient (IFN-␣R⫺/⫺) C57BL/6
mice were bred and maintained in the Institut Pasteur animal facility under specific-pathogen-free conditions. All animal procedures were performed in compliance with approved protocols and Institut Pasteur guidelines for the proper use and care of laboratory animals.
DNA immunization.Each mouse was injected on a single occasion with 100g of endotoxin-free plasmid DNA (PlasmidFactory, Germany) that either ex-pressed the HBV small and middle envelope proteins (pCMVS2.S) (39) or was devoid of genes coding for HBV proteins (pCDNA3.1). The DNA was injected intramuscularly (i.m.) in a final volume of 100l directly into mature tibialis anterior muscles without any muscle pretreatment (30).
Formulation of oligodeoxynucleotides and immunization protocol.The CpG oligodeoxynucleotide (ODN) 1826, a class B ODN (5⬘-TCC ATG ACG TTC CTG ACG TT-3⬘) and the negative-control ODN (5⬘-TCC ATG AGC TTC CTG AGC TT-3⬘) were purchased from InvivoGen (San Diego, CA). Ten micrograms of ODN 1826 or control ODN was administered once by i.m. injec-tion into wild-type C57BL/6 mice, as for the DNA immunizainjec-tion.
Cell preparation and flow cytometry analysis.After perfusion with phosphate-buffered saline, the liver was removed and gently pressed through a 100-m cell strainer. Parenchymal cells (pellet) were separated from mononuclear cells (MNCs) by centrifugation at 50⫻gfor 5 min (56). After a single wash, MNCs were isolated using a 35% Percoll solution (Amersham Biosciences), and red blood cells were lysed in an ammonium chloride buffer. Spleen cells were pre-pared as described previously (34). Spleen and liver MNCs were stained with fluorescein isothiocyanate (FITC)-labeled anti-Mac-1 (M1/70; BD Biosciences), phycoerythrin-labeled anti-NK1.1 (clone PK136; BD Biosciences), FITC-labeled anti-NKp46 (BD Biosciences), peridinin chlorophyll protein-labeled anti-CD3 (145-C11; BD Biosciences), and allophycocyanin-labeled anti-CD27 (LG.7F9; eBioscience). Then, membrane labeling and intracellular cytokine staining reac-tions were performed as previously described (35). Apoptotic cells were quan-tified using APC-labeled anti-annexin V (BD Pharmingen) and 7-amino-actino-mycin (7-AAD). At least 5,000 events gated among the population of interest
were acquired on a FACSCalibur or FACSCanto cytometer using CellQuest or Diva software (BD Biosciences) and analyzed with FlowJo software (Tree Star, Inc., Ashland, OR).
In vitroevaluation of NK cell functions.To evaluate IFN-␥production by NK cells, 4⫻106splenocytes were cultured for 20 h with recombinant mouse IL-12
(rIL-12; 25 ng/ml; R&D Systems) or with medium. Then, IFN-␥intracellular cytokine staining was performed as previously described (18).
To evaluatein vitrocytotoxicity, a total of 2⫻106
spleen MNCs labeled with FITC-conjugated anti-CD107a (clone1D4B; BD Biosciences) from DNA-immu-nized or nonimmuDNA-immu-nized mice were incubated for 4 h with YAC-1 target cells (effector/target ratio [E:T], 50:1) or with medium alone. For wild-type mice, this corresponded to a 1:1 ratio of NK cells to target cells. CD107a expression on NK cells was measured as described previously (1).
Depletion of NK cells.To deplete NK cells, mice were injected in the perito-neum with rabbit anti-mouse/rat asialo-ganglioN-tetraoglyceramide (asialo-GM1) polyclonal antibody (Cedarlane, Canada), 2 days before and 0, 5, and 10 days afterpCMVS2.S injection. Alternatively, anti-NK1.1 monocloncal antibody (MoAb; PK136) was injected intravenously (i.v.; 50g in 200l) 1 day before and 5 days afterpCMVS2.S injection (28). Depletion of the NK cell population was confirmed by flow cytometry on peripheral blood, spleen, and liver MNCs. ELISPOT assays for T cells.IFN-␥-producing T cells were quantified among splenocytes byex vivo enzyme-linked immunosorbent spot (ELISPOT) assay after peptide stimulation for 24 h, as previously described (4). The CD8⫹T cell response was tested using the H-2Kb
-restricted V-8-M and I-8-L HBV envelope peptides. The CD4⫹ T cell response was tested using the IAb-restricted
[image:2.585.301.542.68.295.2]RGLYFPAGGSSSG peptide (R-13-G) (40). Briefly, sterile nitrocellulose MSIP 96-well plates (Millipore, Bedford, MA) were prewetted for 1 min with 15l 35% ethanol, washed with water and coated at 4°C overnight with 50l mouse IFN-␥MAb (R4-6A2; 5g/ml; BD Biosciences) in 0.1 M bicarbonate buffer (pH⫽9.6). Freshly isolated splenocytes were incubated with individual peptides FIG. 1.pCMVS2.S DNA injection activates NK cells. MNCs were isolated from livers and spleens ofpCMVS2.S DNA-immunized and nonimmunized wild-type C57BL/6 mice at different time points after injection (0, 12, 24, 36, and 48 h and 7, 10, and 14 days) and analyzed for the percentage of total NK cells among lymphocytes. (A) Kinetics of the percentage of NK cells gated as NK1.1⫹ CD3⫺ cells among lymphocytes in liver (upper panel) and spleen (lower panel), deter-mined by flow cytometry and expressed as means⫾SEM of NK cells from 3 to 28 mice in each group at each time point. The maximal decrease in NK cells was observed at 36 h postinjection and is indicated by arrows.*,P⬍0.05. (B) At 36 h afterpCMVS2.S DNA injection, the expression levels of B220 (upper panel) and CD69 (lower panel) on spleen-derived NK cells were evaluated forpCMVS2.S-immunized or nonimmunized mice and are shown as means⫾SEM of 6 to 12 mice in each group.
on November 7, 2019 by guest
http://jvi.asm.org/
at 2g/ml in complete␣-MEM medium (4). Splenocytes from nonimmunized mice and cells in culture medium alone were used as negative controls to determine background levels. A Bioreader 4000 counter (BioSys) was used to score the number of spots. Each cell population was titrated in triplicate. The response was considered positive if the median number of spot-forming cells (SFC) in triplicate wells was at least twice that observed in control wells con-taining medium, and at least 50 SFC per million splenocytes were detected.
Statistical analysis.Data were expressed as means⫾SEM. Nonparametric unpaired comparisons were performed using the Mann-WhitneyUtest.
Statis-tical analysis was carried out using Graphpad Prism 5 software. Values ofP⬍
0.05 were considered significant.
RESULTS
Injection of DNA encoding small and middle HBV envelope proteins results in activation of NK cells in liver and spleen.
[image:3.585.135.451.64.547.2]We previously showed that i.m. injection ofpCMVS2.S DNA, FIG. 2. DNA injection induces early maturation of NK cells in liver. Wild-type C57BL/6 mice were injected i.m. withpCMVS2.S DNA. (A) Kinetics of the percentage of the Mac-1⫹CD27⫹NK cell subset among total NK cells in liver (left panel) and spleen (right panel), expressed as means⫾SEM of 4 to 28 mice per group.*,P⬍0.05. (B) The 4-stage process of NK cell maturation in mice. (C) The percentages of the different NK cell subsets in liver (upper panels) and spleen (lower panels) frompCMVS2.S-immunized mice 36 h postinjection and from nonimmunized mice were evaluated by flow cytometry. Data are expressed as the means for 9 to 12 mice/group, and the percentages of each subset among NK cells are represented as pie charts. (D) Representative fluorescence-activated cell sorting plots of Mac-1 and CD27 staining, used to define the percentages of NK cell subsets found in livers of nonimmunized andpCMVS2.S DNA-immunized mice.
VOL. 85, 2011 NK CELL MATURATION AND ACTIVATION BY DNA VACCINE 10203
on November 7, 2019 by guest
http://jvi.asm.org/
a DNA vaccine that encodes the small and middle HBV en-velope proteins, elicited HBV-specific IFN-␥-secreting T cells and anti-HBs antibodies in mice (29, 33) and in chronic HBV patients (36, 37). Herein, we used a single i.m. injection of
pCMVS2.S DNA into C56BL/6 mice to investigate the involve-ment of NK cells in immune responses induced during DNA-based immunization. Injection was into mature muscle, with-out any cardiotoxin pretreatment that could locally attract inflammatory cells and possibly impact on our observations (30). We first analyzed the kinetics of NK cell expansion in C57BL/6 mice spleen and livers following pCMVS2.S DNA injection. NK cells were defined as NK1.1⫹CD3⫺cells in lym-phocyte populations. 36 h afterpCMVS2.S injection, the per-centage of NK cells in spleen and liver was dramatically and significantly reduced (6.44 ⫾ 0.53% (n ⫽ 13) versus8.08 ⫾ 0.30% (n⫽28) for liver,P⬍0.05 and 1.48⫾0.18% (n⫽10)
versus2.28⫾0.14% (n⫽31) for spleen,P⬍0.05) (Fig. 1A).
To confirm this observation, the absolute number of NK cells in spleen and liver was determined and we found this was significantly reduced after pCMVS2.S DNA injection, com-pared to nonimmunized mice (see Fig. 3A). Accordingly, quan-tification of apoptotic cells using Annexin V and 7-AAD la-beling showed that the numbers of early apoptotic cells were significantly higher in the spleens of mice injected with
pCMVS2.S DNA than in the spleens of untreated mice (6.99⫾ 0.14% (n ⫽3) versus3.85⫾ 0.74% (n ⫽3), P ⬍ 0.05). As shown in Fig. 1B, the decrease in NK cells at 36 h in immu-nized mice was associated with a significant increase in the expression of activation markers such as B220 and CD69 on spleen NK cells, suggesting that i.m.pCMVS2.S injection ac-tivates NK cells in peripheral organs.
DNA injection induces early modulation of NK cell subsets.
In the mouse, distinct NK cell subsets with specialized func-tions and repertoires have been identified. NK cell subsets express various cytotoxicity receptors, such as NKp46 (52), and the activating receptors NKG2D (21) or Mac-1 and CD27 (20). We examined whether NK cell subsets could be modulated followingpCMVS2.S DNA injection. As illustrated in Fig. 2A, a single injection ofpCMVS2.S resulted in a significant reduc-tion (P⬍0.005) in the percentage of the Mac-1⫹CD27⫹NK cell subset in both liver (11.06⫾1.51% [n⫽8] versus 19.27⫾ 1.39% [n⫽26]; means⫾standard errors of the means [SEM]) and spleen (15.72⫾1.68% [n⫽4] versus 24.59⫾1.35% [n⫽
[image:4.585.43.285.67.718.2]29]) 36 h posttreatment. In contrast, no change was observed for either NKp46⫹ or NKG2D⫹ NK cell subsets (data not
FIG. 3. CpG sequences contribute to the alteration of the Mac-1⫹ CD27⫹NK cell subset during DNA-based immunization in mice. (A and B) Wild-type C57BL/6 mice were either nonimmunized (n⫽16) or receivedpCDNA3.1 (n⫽14) orpCMVS2.S (n⫽14) DNA injec-tions. At 36 h after injection, the percentages (left panels) and absolute number (right panels) of total NK cells (A) and of the Mac-1⫹CD27⫹ NK cell subset (B) were evaluated in liver (uper panels) and spleen (lower panels). Data are means⫾SEM. (C) Wild-type C57BL/6 mice were injected with 10g control ODN (left panel) or ODN 1826 (right panel). Mac-1 CD27 NK cell subsets were analyzed by flow cytometry 36 h after ODN injection. Representative flow cytometry analysis of Mac-1 and CD27 expression by hepatic NK cells from two independent experiments is shown. The numbers in the quadrants represent the percentages of NK cell subsets.
on November 7, 2019 by guest
http://jvi.asm.org/
shown). Between 7 and 14 days after DNA injection, the per-centages of total NK cells and the Mac-1⫹CD27⫹subset had returned to the basal levels.
NK cell maturation is a 4-stage process that starts at the Mac-1⫺CD27⫺stage and terminates at the Mac-1⫹CD27⫺ stage (Fig. 2B). At 36 h afterpCMVS2.S injection, a reduced percentage of the intermediate Mac-1⫹CD27⫹NK subset was observed in the mouse liver (14.11 ⫾ 1.34%) compared to nonimmunized mice (21.15⫾1.92%;P⬍0.01) (Fig. 2C, upper panel). A representative dot plot of the flow cytometry analysis is shown in Fig. 2D for nonimmunized andp CMVS2.S-immu-nized mice. The decrease was concomitant with a significant increase in the most mature Mac-1⫹CD27⫺ NK cell subset (38.41⫾3.84% [n⫽9] versus 49.59⫾2.20% [n⫽12];P⬍
0.05). The level of this NK cell subpopulation in the liver even exceeded the percentage detected in the spleen (Fig. 2C, lower panel). This suggested that DNA injection induced precocious differentiation and maturation of NK cells in the liver.
Changes in NK cell subsets during DNA-based immuniza-tion are not antigen dependent.To investigate whether the modification in NK cell subsets induced bypCMVS2.S DNA injection was antigen dependent, wild-type C57BL/6 mice were injected with eitherpCMVS2.S orpCDNA3.1, the latter being devoid of genes that code for HBV proteins, or were nonim-munized. The percentage of total NK cells and their subsets, including Mac-1⫹CD27⫹, were evaluated 36 h after injection (Fig. 3). Significant reductions in the percentage of total NK cells (Fig. 3A, left panels) and in the percentage of the Mac-1⫹
CD27⫹NK cell subset (Fig. 3B, left panels) were observed in liver and spleen when mice were injected with either
pCDNA3.1 orpCMVS2.S. Similarly, the absolute numbers of total NK cells and of Mac-1⫹CD27⫹ NK cells were signifi-cantly decreased in the liver and spleen of DNA-injected mice compared to nonimmunized mice (Fig. 3A and B, right pan-els). This indicated that the alteration of NK cells was not antigen dependent but could be due to bacterial sequences present within the plasmid backbone.
As synthetic ODNs containing unmethylated CpG se-quences are known to mimic the immunostimulatory activity of bacterial plasmid sequences (24), wild-type C57BL/6 mice were injected i.m. with 10g of either CpG-ODN 1826 or its control ODN. Similar to DNA immunization, a single injection of ODN 1826 substantially reduced the percentage of NK cells found in the spleen at 36 h postinjection (from 3.108⫾0.26% for untreated mice [n⫽4] to 0.660⫾0.15% for ODN 1826-injected mice [n⫽6];P⬍0.01). In contrast, the control ODN injection did not significantly affect the percentage of splenic NK cells (2.505⫾0.20% [n⫽6]). A significant decrease in the Mac-1⫹CD27⫹NK cell subset was also observed in mice that received ODN 1826 (11.19⫾1.61% [n⫽6]) compared to the control ODN group (17.89⫾1.48% [n ⫽6];P⫽0.013). In addition, a 2-fold increase in the percentage of the mature Mac-1⫹ CD27⫺ NK subset arose in the livers of mice that received ODN 1826 (70.02 ⫾ 4.59% [n ⫽ 6], compared to 33.81⫾2.45% for control ODN [n⫽6]). An absolute number determination confirmed this result (data not shown). A rep-FIG. 4. Absence of phenotypic alteration of NK cells in MyD88⫺/⫺mice. Wild-type C57BL/6 (WT) or MyD88⫺/⫺mice were either
nonim-munized or imnonim-munized by injection withpCMVS2.S DNA. The percentages of total NK cells (A) and of the Mac-1⫹CD27⫹NK subset (B) in liver (left panels) and spleen (right panels) were quantified 36 h postinjection. Data are means⫾SEM of 3 to 4 mice per group. NS, not significant.
VOL. 85, 2011 NK CELL MATURATION AND ACTIVATION BY DNA VACCINE 10205
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.107.475.72.357.2]resentative flow cytometry analysis is shown in Fig. 3C. Glob-ally, our results clearly suggest that the alteration of NK cell subsets is due to bacterial sequences, early after DNA-based immunization.
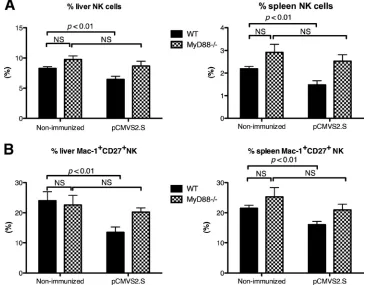
Alteration of NK cells during DNA-based immunization de-pends on the TLR signaling pathway.The sequence of bacte-rial origin in plasmid DNA contains CpG motifs, which are recognized by TLR9 and lead to an adjuvant effect through activation of the innate immune system (46). However, some studies have shown that TLR9 or CpG is not essential for the induction of innate immunity after DNA vaccine injection (3, 49). This prompted us to clarify whether DNA vaccine acti-vated NK cells through the TLR signaling pathway. The phe-notypes of liver- and spleen-derived NK cells were evaluated 36 h afterpCMVS2.S injection into mice deficient for MyD88 protein, which is required for signaling through most TLRs. The percentages of NK cells did not differ significantly between naive MyD88-deficient and wild-type mice (Fig. 4A). The per-centages of total NK cells and of the Mac-1⫹CD27⫹NK cell subset were not significantly reduced in liver and spleen upon
pCMVS2.S immunization of MyD88⫺/⫺mice (Fig. 4A and B,
hatched bars). This contrasted with the situation in wild-type mice, where the percentage of total NK cells and of the Mac-1⫹CD27⫹NK cell subset were significantly reduced (P⬍
0.01) (Fig. 4A and B, black bars). This suggests that the phe-notypic alteration of NK cells observed in wild-type mice early after DNA-based immunization is dependent on the TLR sig-naling pathway.
pCMVS2.S injection enhances cytotoxicity and cytokine pro-duction by NK cells. NK cells mediate numerous functions, including cytotoxicity and cytokine production. We examined whether the injection of plasmid DNA affected NK cell func-tions. To this end, we used an ex vivoassay in which freshly purified splenocytes were stimulated with YAC-1 target cells for 4 h. Cytotoxic granule release was measured, which is propor-tional to surface CD107a exposure. In response to YAC-1 target cells, the percentage of spleen NK cells expressing CD107a was significantly increased when mice were immunized with
pCMVS2.S DNA compared to injection withpCDNA3.1 or to nonimmunized mice (Fig. 5A). It is of note that, in the absence of YAC-1 target cells, the percentage of NK cells positive for CD107a expression was already increased in the spleens of
pCMVS2.S-injected mice compared to nonimmunized mice. As IFN-␥ synthesis by NK cells is usually very low in the absence ofin vivopreactivation by an IFN-␣inducer (51), we quantified IFN-␥synthesis by NK cellsin vitroin response to rIL-12 by using a single-cell analysis after intracellular staining of splenocytes. The percentage of IFN-␥-producing NK cells was increased 2-fold for mice immunized withpCMVS2.S com-pared to mice injected withpCDNA3.1 or to nonimmunized mice (Fig. 5B). Importantly, neither the percentage of NK cells expressing CD107a nor IFN-␥ production by NK cells was enhanced in mice injected withpCDNA3.1 DNA. In contrast,
pCMVS2.S-DNA immunization enhanced NK cell effector functions, suggesting that antigen expression is required for the activation of NK cells early after injection with a DNA plasmid that encodes HBV proteins.
Role of T cells in NK cell maturation and function. The priming of antigen-specific T cells is an early event that occurs in lymph nodes following DNA-based immunization. To assess
the potential role of antigen-specific T cells on NK cells, we injected CD3ε⌬5/⌬5 mutant mice withpCMVS2.S DNA (44). Thirty-six hours after injection, neither the percentage of total NK cells (Fig. 6A) nor the percentage of NK cell subsets (Fig. 6B), either in the spleens or livers of the CD3εmutant mice, was significantly different from the respective percentages found in untreated mice (Fig. 6). This suggests a potential role for antigen-specific T cells in NK cell maturation.
The percentage of NK cells expressing CD107a increased 2-fold inpCMVS2.S DNA-immunized CD3εmutant mice but remained lower than that of immunized C57BL/6 wild-type mice (Fig. 6C and 5A). This might be related to the different numbers of NK cells in the splenocytes that were used to assess cytotoxicity, which were 4-fold higher in the splenocytes from CD3ε⌬5/⌬5 mutant mice than in the splenocytes of wild-type mice (Fig. 6A). Regarding IFN-␥production, the percentage of CD3ε NK cells secreting IFN-␥was very low, even after stimulation with rIL-12. Therefore, in the absence of T cells, NK cell maturation is not achieved and NK cell functions are only slightly enhanced after immunization withpCMVS2.S.
[image:6.585.347.491.69.307.2]Activation of NK cells depends on endogenous IFN-␣during DNA-based immunization.Because induction of type-I IFNs plays an important role in adaptive immune responses during DNA-based vaccination (23), and since it is known that type-I IFNs can influence the function and homeostasis of NK cells (50), we investigated whether activation of NK cells induced by FIG. 5.pCMVS2.S DNA injection enhances the functions of NK cells. Wild-type C57BL/6 mice were injected with eitherpCMVS2.S or pCDNA3.1 DNA or were nonimmunized. (A) The percentages of CD107a expression on spleen-derived NK cells in medium alone or after 4 h of incubation with YAC-1 target cells were measured by flow cytometry 36 h after DNA injection (E:T [NK:YAC] ratio, 1:1). Data are expressed as means⫾SEM for 11 to 23 C57BL/6 mice per group. (B) Percentages of IFN-␥-producing NK cells analyzed by intracyto-plasmic staining of spleen-derived cells incubated in medium alone or with rIL-12 for 20 h. Data are expressed as means⫾SEM for 10 to 19 C57BL/6 mice per group.
on November 7, 2019 by guest
http://jvi.asm.org/
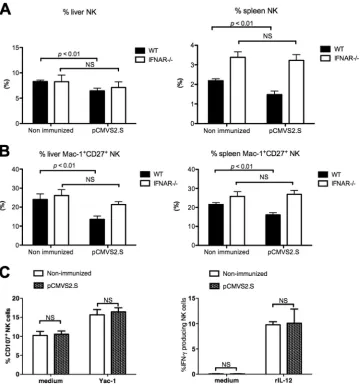
pCMVS2.S was dependent on the type I IFN pathway in mice. IFN-␣R⫺/⫺mice, which lack the type I IFN-mediated signaling
pathway, and wild-type mice were immunized withpCMVS2.S DNA. The absolute numbers of splenic NK cells did not differ significantly between nonimmunized IFN-␣R⫺/⫺ mice and
wild-type mice (data not shown). At 36 h after pCMVS2.S DNA injection, the percentages of NK cells and of the Mac-1⫹ CD27⫹ subset were similar in both liver and spleen from
pCMVS2.S-immunized and nonimmunized IFN-␣R⫺/⫺mice
(Fig. 7A and B). The CD107a expression on NK cells in re-sponse to YAC-1 target cells was unchanged in spleen cells from IFN-␣R⫺/⫺mice, whether they werepCMVS2.S
immu-nized or nonimmuimmu-nized (Fig. 7C, left panel). IFN-␥production by spleen NK cells in response to rIL-12 was also to the same level in immunized and nonimmunized IFN-␣R⫺/⫺mice (Fig.
7C, right panel). Therefore, in contrast to the NK cells of immunized C57BL/6 wild-type mice, plasmid immunization of IFN-␣R-deficient mice did not modify the total percentage, the subset percentages, or the functions of NK cells found in
the spleen. Our results support the notion that the endogenous IFN-␣pathway is implicated in the maturation and functional activation of NK cells early after HBV DNA immunization.
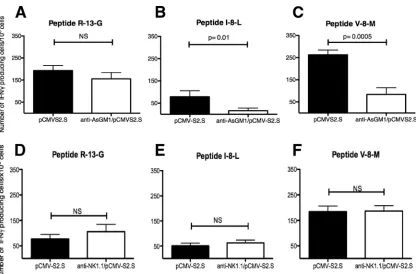
NK cell depletion with anti-asialo-GM1 but notanti-NK1.1 decreases the number of HBV-specific CD8ⴙT cells in mice 14 days after immunization.We examined whether NK cells play a direct role in the induction of adaptive immunity during
[image:7.585.136.453.69.449.2]pCMVS2.S DNA-based immunization. To this end, mice were depleted of NK cellsin vivoby anti-asialo-GM1 (Fig. 8A) or anti-NK1.1 (Fig. 8B) antibody injection before and after im-munization withpCMVS2.S. Anti-asialo-GM1 resulted in pletion of approximately 92% of NK cells. No significant de-crease of NKT cells in liver (12.20⫾2.44% [n ⫽10] versus 16.95 ⫾ 3.04% [n ⫽ 9]) or spleen (1.29 ⫾ 0.07% [n ⫽ 10] versus 1.52 ⫾ 0.11% [n ⫽ 9]) was detected 14 days after anti-asialo-GM1 treatment (Fig. 8A). Anti-NK1.1 injection de-pleted 91.4% of the NK cells in the spleen 14 days after im-munization (Fig. 8B). Anti-CD1d tetramer staining showed that 34% of iNKT cells were depleted from the spleen after FIG. 6. Activation of NK cells induced bypCMVS2.S immunization depends on T cells. At 36 h afterpCMVS2.S injection into wild-type C57BL/6 mice or CD3ε⫺/⫺mice, the percentages of total NK cells (A) and of the Mac-1⫹CD27⫹NK cell subset (B) in the livers (left panels) and
spleens (right panels) of nonimmunized CD3ε⫺/⫺and C57BL/6 mice (n⫽4 each) andpCMVS2.S-immunized CD3ε⫺/⫺and C57BL/6 mice (n⫽
5 each) were determined. (C) Percentages of NK cells from the spleens ofpCMVS2.S-immunized or nonimmunized CD3ε⫺/⫺mice, which
expressed CD107a (left panel, E:T ratio [NK:YAC], 4:1) or produced IFN-␥in response to rIL-12 (right panel) are shown. Data are means⫾SEM.
VOL. 85, 2011 NK CELL MATURATION AND ACTIVATION BY DNA VACCINE 10207
on November 7, 2019 by guest
http://jvi.asm.org/
anti-NK1.1 injection (data not shown). CD4⫹and CD8⫹T cell responses were evaluated 2 weeks after immunization inex vivo
IFN-␥ELISPOT assays. The CD4⫹T cell response was tested using the IAb-restricted HBV-derived R-13-G peptide. In
re-sponse to this peptide, the number of IFN-␥-producing cells in mice did not change significantly upon NK depletion (Fig. 9A and D). The CD8⫹ T cell response was assessed using the H-2Kb-restricted V-8-M and I-8-L peptides, which are derived,
respectively, from endogenous and exogenous processing of the HBV envelope proteins (47). The number of cells that produce IFN-␥in response toex vivostimulation with the two peptides was significantly lower in mice depleted of NK cells when we used an anti-asialo-GM1 antibody than in nonde-pleted mice (Fig. 9B and C). In contrast, no reductions in CD8⫹ T cell responses were observed following anti-NK1.1 antibody treatment (Fig. 9E and F). Unexpectedly, anti-asialo-GM1 but not anti-NK1.1 treatment markedly reduced the number of antigen-specific CD8⫹T cells in the spleen, whereas
both anti-asialo-GM1 and anti-NK1.1 antibodies efficiently de-pleted NK cells.
DISCUSSION
[image:8.585.112.472.69.455.2]It is widely accepted that once plasmid DNA is adminis-tered, the encoded antigen is expressed in the host cell and then captured, processed, and presented by antigen-presenting cells, such as DC, which elicit humoral and cellular responses. Plasmid DNA-based vaccines therefore constitute an efficient tool for the prevention of, or therapeutic intervention against, pathogens and tumors. However, the molecular and cellular mechanisms underlying DNA-based vaccine immunogenicity are not yet well understood. In particular, it still remains un-clear how cellular innate immunity is involved and how this triggers the adaptive immune response and increases immuno-genicity during DNA-based immunization. Our study provides evidence that a DNA-based vaccine activates NK cells and FIG. 7. Activation of NK cells induced bypCMVS2.S immunization depends on endogenous IFN-␣. At 36 h afterpCMVS2.S injection into C57BL/6 or IFN-␣R⫺/⫺mice, the percentages of total NK (A) and of the Mac-1⫹CD27⫹NK cell subset (B) in liver (left panels) and spleen (right
panels) from nonimmunized orpCMVS2.S-immunized IFN-␣R⫺/⫺mice were determined. (C) Percentages of NK cells expressing CD107a (left
panel) and producing IFN-␥in response to rIL-12 (right panel) from the spleen ofpCMVS2.S-immunized (hatched bars) or nonimmunized (empty bars) IFN-␣R⫺/⫺. Data are means⫾SEM for 6 to 7 IFN-␣R⫺/⫺mice per group and 10 C57BL/6 mice per group.
on November 7, 2019 by guest
http://jvi.asm.org/
induces early maturation of NK cells in the liver of immuno-competent mice. This phenotypic alteration is accompanied by an enhancement of NK cell functions, including cellular cyto-toxicity and IFN-␥secretion which is linked to expression of antigen by DNA vaccine. However, activation of NK cell func-tions depended on IFN-␣production and was most effective in the presence of antigen and T cells, suggesting a positive feed-back role for antigen-specific T cells on NK cells. Shortly after plasmid injection, the absolute number of NK cells was signif-icantly reduced in the mouse liver and spleen, supporting the hypothesis that NK cell activation is triggered during DNA-based immunization. Quantification of apoptotic cells using annexin V and 7-AAD labeling showed a significant increase in early apoptotic cells in the spleens of pCMVS2.S DNA-in-jected mice compared to those in the spleens of untreated mice. Therefore, NK cell death and/or recirculation to reach the injection site or the lymph nodes probably accounted for the decrease in NK cells. A considerable increase in CD69 and B220 expression in NK cells confirmed their activation state upon plasmid injection. Activation is not induced directly through activating receptors, since NKp46/NKG2D expression on NK cells early after DNA injection was unchanged (data not shown). We even observed phenotypic alteration of NK cells in mice injected with pCDNA3.1, a plasmid devoid of antigen-encoding sequences. This indicates that activation of
NK cells is not antigen dependent but is linked to bacterial sequences present on the plasmid backbone. It was recently shown that bacterial DNA within cells is sensed through dif-ferent pathways, such as TLR, DLM-1/ZBP1 (later renamed DAI), and TBK1 (23). In our study, a single injection of CpG-ODN 1826, but not of a control CpG-ODN, elicited a remarkable reduction in NK cells in the spleen and a less pronounced reduction in the liver. Nevertheless, these effects were accom-panied by a greatly increased percentage of the more mature Mac-1⫹CD27⫺NK cell subset in the liver. CpG-ODN 1826, which is known to target TLR9, therefore mimicked the effects of a DNA based-vaccine and revealed that bacterial sequences within a DNA plasmid target cellular innate immunity medi-ated by NK cells early after injection. To clarify the molecular and/or cellular mechanisms underlying the activation of NK cells by bacterial sequences, we used MyD88⫺/⫺mice, which
lack most TLR signaling pathways. In our hands, the pheno-typic alteration of NK cells specifically induced bypCMVS2.S DNA-based immunization did not occur in MyD88⫺/⫺mice,
[image:9.585.104.484.69.363.2]revealing the importance of the MyD88 signaling pathway for plasmid-induced NK cell maturation.pCMVS2.S DNA injec-tion into mice led to precocious maturainjec-tion of the Mac-1⫹ CD27⫺NK cell subset in liver that even exceeded the level observed in spleen. This might allow the mature NK cell subset to exert effector functions. Indeed, we demonstrated an early FIG. 8. Anti-asialo-GM1 and anti-NK1.1 treatments induce efficient depletion of NK cells in mice. Mice were treated by intraperitoneal injection of anti-asialo-GM1 antibody (A) or i.v. injection of anti-NK1.1 antibody (B). NK cell depletion was verified by flow cytometry at the time of immunization in peripheral blood and 14 days after immunization in the liver and spleen. Representative fluorescence-activated cell sorting dot plots are shown for each group (n⫽8 to 10). Values indicated in the squares are the percentages of NK cells (CD3⫺NK1.1⫹) or NKT cells (CD3⫹ NK1.1⫹) among lymphocytes after anti-asialo-GM1-mediated depletion and the percentages of NK cells (CD3⫺ NKp46⫹) after anti-NK1.1-mediated depletion.
VOL. 85, 2011 NK CELL MATURATION AND ACTIVATION BY DNA VACCINE 10209
on November 7, 2019 by guest
http://jvi.asm.org/
and considerable enhancement of NK cell functions in mice immunized withpCMVS2.S DNA, including cellular cytotox-icity and IFN-␥secretion. It was suggested that the less mature Mac-1⫹CD27⫹NK subset is more potent for cytotoxicity and cytokine production than the more mature Mac-1⫹CD27⫺NK subset (20). In our study, inpCMVS2.S-injected mice, com-pared to nonimmunized mice, a significant increase in CD107a expression was observed in the two spleen NK cell subsets in the presence of YAC-1 target cellsin vitro. CD107a expression was higher on Mac-1⫹ CD27⫹ NK cells than on the more mature Mac-1⫹CD27⫺NK cells (data not shown). In response to rIL-12in vitro, a downregulation of CD27 expression by NK cells was observed (data not shown). These features made it difficult to determine whether or not both effector functions were performed by the same NK cell subset. This is consistent with the results of De Colvenaer et al., who reported that continuous CD27 triggeringin vivoresulted in strongly down-modulated CD27 expression on NK cells and in a severe re-duction in NK cell numbers (12). Our results that showed downregulation of CD27 expression on NK cells and a de-crease in NK cell percentages after DNA-based immunization indirectly confirm their activation state.
It is generally acknowledged that NK cells interact with other components of the immune system, such as DC. The NK-DC synapse can trigger activation of both NK and DC and, in turn, enable NK cells to exert their helper functions, such as maturation, activation, or death of DC (31). This suggests that
NK cells may indirectly modulate adaptive immune responses. Recently, Rottembourg et al. provided evidence that CpG/ TLR9 signaling is involved in activation of DCs after a single plasmid DNA injection in mice (45). Yochida et al. showed impaired DC functions in NK-depleted mice immunized with a protein-based hepatitis B vaccine that contains HBsAg (54). Hence, it is reasonable to believe that NK-DC cross talk is involved in pCMVS2.S DNA immunization. We found that enhancement of NK cell function induced bypCMVS2.S DNA immunization was dependent on specific antigen rather than on bacterial sequences (Fig. 5), indirectly supporting NK-DC cross talk. Moreover, we observed thatpCMVS2.S immuniza-tion-induced early liver NK cell maturation is associated with a significant increase in myeloid DC in liver (data not shown). This might facilitate NK-DC communication and accelerate the establishment of an adaptive immune response.
[image:10.585.84.500.65.339.2]DCs are also involved in innate immune responses through the production of antimicrobial cytokines, such as type I IFNs, especially those secreted by plasmacytoid DC. Type I IFN exposure promotes NK cytotoxicity (7) and IFN-␥release (22). Recently, Ishii et al. showed that type I IFN signaling was essential for DNA-induced antigen-specific T cell responses (23). In our study, the absence of NK cell phenotypic and functional alterations early afterpCMVS2.S immunization in IFN-␣R-deficient mice confirmed the crucial role of the type I IFN pathway inpCMVS2.S-induced NK cell activation. A role for newly primed antigen-specific T cells on NK cell activation FIG. 9. Number of HBV-specific CD8⫹T cells in the spleens ofpCMVS2.S DNA-immunized mice after NK cell depletion. Wild-type C57BL/6 mice were depleted, or not, for NK cellsin vivoby using anti-asialo-GM1 anitbody (A, B, and C) or anti-NK1.1 antibody (D, E, and F) injection before and afterpCMVS2.S DNA injection. Cellular responses were analyzed 14 days post-pCMVS2.S injection. IFN-␥-producing cells were quantified by ELISPOT assay on spleen cells stimulatedex vivowith HBV envelope-derived R-13-G (A and D), I-8-L (B and E), or V-8-M (C and F) peptides. Results are expressed as means⫾SEM of the number of IFN-␥-producing cells/106splenocytes from anti-asialo-GM1-depleted, anti-NK1.1-depleted (white bars), or nondepleted (black bars) mice (8 to 13 mice per group).
on November 7, 2019 by guest
http://jvi.asm.org/
and maturation cannot be excluded. Bihl et al. recently dem-onstrated that antigen-specific CD4⫹T cells, by secreting IL-2, are required for NK cell activationin vivoduringLeishmania
majorinfection (5). Our experiments with DNA immunization
in CD3εmutant mice and in Rag-2⫺/⫺mice (data not shown),
which are devoid of T cells, support the hypothesis that anti-gen-specific T cells are not required for early maturation of NK cells but may rather act in synergy with DC to induce the functional maturation of NK cells in lymph nodes. Finally, we addressed the role of NK cells on adaptive T cell responses. Recent data showed that NK cells are capable of initiating robust antigen-specific T cell responses (26). Anti-asialo-GM1 is widely used to deplete NK cells and was shown to be more effective than anti-NK1.1 antibody (26). In our hands, NK cell depletion using anti-asialo-GM1 or anti-NK1.1 antibodies yielded conflicting results regarding CD8⫹ T cell responses. The CD8⫹T cell response was dampened by NK cell depletion with anti-asialo-GM1 but not with anti-NK1.1 antibody. Nota-bly, asialo-GM1 expression is not strictly confined to NK cells and some NKT cell subsets, and it has been detected on a subpopulation of CD8⫹T cells in C57BL/6 mice (25) and on basophils (42). Therefore, we cannot exclude the possibility that the observed decrease in antigen-specific T cells after anti-asialo-GM1 treatment might be related to a partial deple-tion of asialo-GM1⫹CD8⫹T cells rather than to a direct effect of NK cells on T cell priming.
With respect to the therapeutic use of an HBV DNA vaccine in patients with chronic hepatitis B, it is obvious that the immunogenicity of apCMVS2.S DNA vaccine could be differ-ent in immuno-competdiffer-ent mice than in chronic HBV-infected patients, who display peripheral NK and T cells with impaired functions (32, 43). The timing of analysis is also important, and indeed our previous results showed an increase in the
CD56brightNK cell subset when measured 1 month after DNA
injection into patients (48). Considered to be abundant cy-tokine producers but weakly cytotoxic, human CD56bright
NK cells are functionally different from the murine Mac-1⫹ CD27⫹NK subset, which combines the two activities. How-ever, it was recently demonstrated that mature CD56dimNK
cells, rather than their CD56brightNK precursor subset, are
the major cytokine producers upon target recognition, illus-trating the complexity of different phenotypes in NK cell subsets (16, 55). This could explain the differences between results observed in chronic HBV-infected patients and in mice.
In conclusion, our study provides new insights into a major implication of NK cells in DNA-based immunization. On the one hand, bacterial sequences present in plasmid DNA vac-cines induce activation and maturation of NK cells through the TLR pathway. On the other hand, the antigen encoded by a DNA vaccine might stimulate DC to produce type I IFNs, thus activating antigen-specific T cells that may further increase the function of NK cells. All these events would contribute to a better development of innate and adaptive immune responses.
ACKNOWLEDGMENTS
This work was supported by fellowships from the French Consulate in Shanghai (to R.Z.), from Tsi Jung Memorial Fund (to R.Z.), from Agence Nationale de Recherche sur Sida et les Hepatitis Virales (to
R.Z.), and from the Association pour la Recherche sur le Cancer (to F.B.).
We are grateful to Daniel Scott-Algara, Claude Roth, and Christian A. J. Vosshenrich (all from the Institut Pasteur) for helpful discussions and critical comments. We thank Vincent Deubel for his constant support.
REFERENCES
1.Alter, G., J. M. Malenfant, and M. Altfeld.2004. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Meth-ods294:15–22.
2.Andoniou, C. E., J. D. Coudert, and M. A. Degli-Esposti.2008. Killers and beyond: NK-cell-mediated control of immune responses. Eur. J. Immunol. 38:2938–2942.
3.Babiuk, S., et al.2004. TLR9-/- and TLR9⫹/⫹mice display similar immune responses to a DNA vaccine. Immunology113:114–120.
4.Bayard, F., S. Malmassari, Q. Deng, Y.-C. Lone, and M.-L. Michel.2010. Hepatitis B virus (HBV)-derived DRB1*0101-restricted CD4 T-cell epitopes help in the development of HBV-specific CD8⫹T cells in vivo. Vaccine 28:3818–3826.
5.Bihl, F., et al.2010. Primed antigen-specific CD4⫹T cells are required for NK cell activation in vivo upon Leishmania major infection. J. Immunol. 185:2174–2181.
6.Biron, C. A.2010. Expansion, maintenance, and memory in NK and T cells during viral infections: responding to pressures for defense and regulation. PLoS Pathog.6:e1000816.
7.Biron, C. A., K. B. Nguyen, G. C. Pien, L. P. Cousens, and T. P. Salazar-Mather.1999. Natural killer cells in antiviral defense: function and regula-tion by innate cytokines. Annu. Rev. Immunol.17:189–220.
8.Bonorino, P., et al.2009. Fine characterization of intrahepatic NK cells expressing natural killer receptors in chronic hepatitis B and C. J. Hepatol. 51:458–467.
9.Cerwenka, A., and L. L. Lanier.2001. Natural killer cells, viruses and cancer. Nat. Rev. Immunol.1:41–49.
10.Chiossone, L., et al.2009. Maturation of mouse NK cells is a 4-stage devel-opmental program. Blood113:5488–5496.
11.Cooper, M. A., T. A. Fehniger, and M. A. Caligiuri.2001. The biology of human natural killer-cell subsets. Trends Immunol.22:633–640.
12.De Colvenaer, V., et al.2010. Continuous CD27 triggering in vivo strongly reduces NK cell numbers. Eur. J. Immunol.40:1107–1117.
13.Di Santo, J. P.2008. Functionally distinct NK-cell subsets: developmental origins and biological implications. Eur. J. Immunol.38:2948–2951. 14.Dunn, C., et al.2007. Cytokines induced during chronic hepatitis B virus
infection promote a pathway for NK cell-mediated liver damage. J. Exp. Med.204:667–680.
15.Dunn, C., et al. 2009. Temporal analysis of early immune responses in patients with acute hepatitis B virus infection. Gastroenterology137:1289– 1300.
16.Fauriat, C., E. O. Long, H. G. Ljunggren, and Y. T. Bryceson.2010. Regu-lation of human NK-cell cytokine and chemokine production by target cell recognition. Blood115:2167–2176.
17.Fisicaro, P., et al. 2009. Early kinetics of innate and adaptive immune responses during hepatitis B virus infection. Gut58:974–982.
18.Haddad, E. A., L. K. Senger, and F. Takei.2009. An accessory role for B cells in the IL-12-induced activation of resting mouse NK cells. J. Immunol. 183:3608–3615.
19.Hayakawa, Y., N. D. Huntington, S. L. Nutt, and M. J. Smyth.2006. Func-tional subsets of mouse natural killer cells. Immunol. Rev.214:47–55. 20.Hayakawa, Y., and M. J. Smyth.2006. CD27 dissects mature NK cells into
two subsets with distinct responsiveness and migratory capacity. J. Immunol. 176:1517–1524.
21.Hayakawa, Y., and M. J. Smyth.2006. NKG2D and cytotoxic effector func-tion in tumor immune surveillance. Semin. Immunol.18:176–185. 22.Hunter, C. A., K. E. Gabriel, T. Radzanowski, L. E. Neyer, and J. S.
Rem-ington.1997. Type I interferons enhance production of IFN-gamma by NK cells. Immunol. Lett.59:1–5.
23.Ishii, K. J., et al.2008. TANK-binding kinase-1 delineates innate and adap-tive immune responses to DNA vaccines. Nature451:725–729.
24.Klinman, D. M.2006. Adjuvant activity of CpG oligodeoxynucleotides. Int. Rev. Immunol.25:135–154.
25.Kosaka, A., et al.2007. AsialoGM1⫹CD8⫹central memory-type T cells in unimmunized mice as novel immunomodulator of IFN-␥-dependent type 1 immunity. Int. Immunol.19:249–256.
26.Krebs, P., et al.2009. NK cell-mediated killing of target cells triggers robust antigen-specific T cell-mediated and humoral responses. Blood113:6593– 6602.
27.Lanier, L. L.1998. NK cell receptors. Annu. Rev. Immunol.16:359–393. 28.Lapaque, N., T. Walzer, S. Me´resse, E. Vivier, and J. Trowsdale.2009.
Interactions between human NK cells and macrophages in response to Sal-monella infection. J. Immunol.182:4339–4348.
29.Loirat, D., F. A. Lemonnier, and M.-L. Michel.2000. Multiepitopic
HLA-VOL. 85, 2011 NK CELL MATURATION AND ACTIVATION BY DNA VACCINE 10211
on November 7, 2019 by guest
http://jvi.asm.org/
A*0201-restricted immune response against hepatitis B surface antigen after DNA-Based immunization. J. Immunol.165:4748–4755.
30.Loirat, D., et al.1999. Muscle-specific expression of hepatitis B surface antigen: no effect on DNA-raised immune responses. Virology260:74–83. 31.Lunemann, A., J. D. Lunemann, and C. Munz.2009. Regulatory NK-cell
functions in inflammation and autoimmunity. Mol. Med.15:352–358. 32.Maini, M., K., and A. Schurich.2010. The molecular basis of the failed
immune response in chronic HBV: therapeutic implications. J. Hepatol. 52:616–619.
33.Malanchere-Bres, E., et al.2001. CpG oligodeoxynucleotides with hepatitis B surface antigen (HBsAg) for vaccination in HBsAg-transgenic mice. J. Virol.75:6482–6491.
34.Mancini, M., M. Hadchouel, P. Tiollais, and M. L. Michel.1998. Regulation of hepatitis B virus mRNA expression in a hepatitis B surface antigen transgenic mouse model by IFN-gamma-secreting T cells after DNA-based immunization. J. Immunol.161:5564–5570.
35.Mancini-Bourgine, M., et al.2007. Hepatitis B virus splice-generated protein induces T-cell responses in HLA-transgenic mice and hepatitis B virus-infected patients. J. Virol.81:4963–4972.
36.Mancini-Bourgine, M., H. Fontaine, C. Brechot, S. Pol, and M. L. Michel. 2006. Immunogenicity of a hepatitis B DNA vaccine administered to chronic HBV carriers. Vaccine24:4482–4489.
37.Mancini-Bourgine, M., et al.2004. Induction or expansion of T-cell re-sponses by a hepatitis B DNA vaccine administered to chronic HBV carriers. Hepatology40:874–882.
38.McCluskie, M. J., and A. M. Krieg.2006. Enhancement of infectious disease vaccines through TLR9-dependent recognition of CpG DNA. Curr. Top. Microbiol. Immunol.311:155–178.
39.Michel, M. L., et al.1995. DNA-mediated immunization to the hepatitis B surface antigen in mice: aspects of the humoral response mimic hepatitis B viral infection in humans. Proc. Natl. Acad. Sci. U. S. A.92:5307–5311. 40.Milich, D., et al.1990. Importance of subtype in the immune response to the
pre-S(2) region of the hepatitis B surface antigen. I. T cell fine specificity. J. Immunol.144:3535–3543.
41.Moretta, A., E. Marcenaro, S. Parolini, G. Ferlazzo, and L. Moretta.2008. NK cells at the interface between innate and adaptive immunity. Cell Death Differ.15:226–233.
42.Nishikado, H., K. Mukai, Y. Kawano, Y. Minegishi, and H. Karasuyama. 2011. NK cell-depleting anti-asialo GM1 antibody exhibits a lethal off-target effect on basophils in vivo. J. Immunol.186:5766–5771.
43.Oliviero, B., et al.2009. Natural killer cell functional dichotomy in chronic
hepatitis B and chronic hepatitis C virus infections. Gastroenterology137: 1151–1160.
44.Renard, V., et al.1995. Normal development and function of natural killer cells in CD3 epsilon delta 5/delta 5 mutant mice. Proc. Natl. Acad. Sci. U. S. A.92:7545–7549.
45.Rottembourg, D., et al.2010. Essential role for TLR9 in prime but not prime-boost plasmid DNA vaccination to activate dendritic cells and protect from lethal viral infection. J. Immunol.184:7100–7107.
46.Sato, Y., et al.1996. Immunostimulatory DNA sequences necessary for effective intradermal gene immunization. Science273:352–354.
47.Schirmbeck, R., W. Bo¨hm, N. Fissolo, K. Melber, and J. Reimann.2003. Different immunogenicity of H-2Kb-restricted epitopes in natural variants of the hepatitis B surface antigen. Eur. J. Immunol.33:2429–2438.
48.Scott-Algara, D., M. Mancini-Bourgine, H. Fontaine, S. Pol, and M.-L. Michel.2010. Changes to the natural killer cell repertoire after therapeutic hepatitis B DNA vaccination. PLoS One5:e8761.
49.Spies, B., et al.2003. Vaccination with plasmid DNA activates dendritic cells via Toll-like receptor 9 (TLR9) but functions in TLR9-deficient mice. J. Immunol.171:5908–5912.
50.Swann, J. B., et al.2007. Type I IFN contributes to NK cell homeostasis, activation, and antitumor function. J. Immunol.178:7540–7549.
51.Vahlne, G., S. Becker, P. Brodin, and M. H. Johansson.2008. IFN-gamma production and degranulation are differentially regulated in response to stimulation in murine natural killer cells. Scand. J. Immunol.67:1–11. 52.Walzer, T., et al.2007. Identification, activation, and selective in vivo
abla-tion of mouse NK cells via NKp46. Proc. Natl. Acad. Sci. U. S. A.104:3384– 3389.
53.Yan, M. X., H. T. Mao, Q. Liu, W. Q. Wang, and Y. Q. Li.2006. Elevated levels of serum soluble E-selectin in patients with chronic hepatitis B: cor-relation with T lymphocyte subsets, NK cells and liver inflammation. Hepa-tol. Res.35:111–117.
54.Yoshida, O., et al.2008. Impaired dendritic cell functions because of deple-tion of natural killer cells disrupt antigen-specific immune responses in mice: restoration of adaptive immunity in natural killer-depleted mice by antigen-pulsed dendritic cell. Clin. Exp. Immunol.152:174–181.
55.Yu, J., et al.2010. CD94 surface density identifies a functional intermediary between the CD56bright
and CD56dim
human NK-cell subsets. Blood115: 274–281.
56.Zhu, R., et al.2007. The pro-Th1 cytokine IL-12 enhances IL-4 production by invariant NKT cells: relevance for T cell-mediated hepatitis. J. Immunol. 178:5435–5442.



![FIG. 5. pcells. Wild-type C57BL/6 mice were injected with eitherpCD107a expression on spleen-derived NK cells in medium alone orafter 4 h of incubation with YAC-1 target cells were measured by flowcytometry 36 h after DNA injection (E:T [NK:YAC] ratio, 1:1)](https://thumb-us.123doks.com/thumbv2/123dok_us/155902.33046/6.585.347.491.69.307/injected-eitherpcd-expression-derived-incubation-measured-owcytometry-injection.webp)
![FIG. 6. Activation of NK cells induced by pexpressed CD107a (left panel, E:T ratio [NK:YAC], 4:1) or produced IFN-C57BL/6 mice or CD35 each) were determined](https://thumb-us.123doks.com/thumbv2/123dok_us/155902.33046/7.585.136.453.69.449/activation-cells-induced-pexpressed-panel-ratio-produced-determined.webp)