0022-538X/80/08-0555/05$02.00/0
Simple Affinity Procedure for the Purification of Mammalian
Viral Reverse Transcriptases
M. G.SARNGADHARAN,' V. S. KALYANARAMAN,' R. RAHMAN,' ANDR. C. GALLO2*
Department of Cell Biology, LittonBionetics, Inc., Kensington,Maryland 20795,' and Laboratory of Tumor CellBiology,NationalCancer Institute,National Institutes of Health,Bethesda,Maryland202052
Polyguanylic acid was found tobe a potentinhibitor of RNase H associated with mammalian viral reverse transcriptase, indicating a strong interaction
be-tween polyguanylic acid and the reverse transcriptase protein. Based on this observation, wehave developed three simple procedures for the purification of mammalian viralreversetranscriptases. In the first procedure,anucleic acid-free extractofRauschermurine leukemiaviruswasappliedtoa column of phospho-cellulose and thereversetranscriptasewaselutedbyalow concentration (50,M) ofpolyguanylic acid. Polyadenylic acid and polyuridylic acid could not replace
polyguanylic acid forthe elution. Inthe second procedure, a polyuridylic
acid-Sepharose column was substituted for phosphocellulose, and the elution was again achieved by polyguanylic acid. In the third affinity procedure, thereverse transcriptase inanucleicacid-free viralextractwasincubated in the cold with 50 ,uM polyguanylic acid and the complex was adsorbed onto a DEAE-cellulose column. Afterwashing toremoveuncomplexed andweakly complexed proteins, thereversetranscriptasewas eluted inaconcentrated form at0.3 MNaCl with arecoveryofgreaterthan70%.Bypolyacrylamide gelanalysisin thepresenceof sodiumdodecylsulfate, theenzymeappearedtobenearlypure.
DNApolymerasesareknowntohave affinities
toanionic
polymers
(2), such asphosphocellu-loseand
carboxymethylcellulose,
andthereforealso interact with variable affinities withmost
nucleic acids becauseofthepolyanionicnature
of the latter. This is in addition to
possible
specific affinitiesbetween
polymerases
andpar-ticular nucleic acids. Several
chromatographic
procedures described for the purification of
DNApolymerases utilize their
affinity
tophos-phocellulose or to one of several matrix-bound
nucleic acidsto adsorb theseenzymes (7). The
enzymesarerecovered fromthese matrices
by
arelatively
nonspecific
stepofelutionwithasaltsolution. Most of the efforts to
improve
thechromatographic
procedures
for thepurification
of DNA
polymerases
havebeen directedatiden-tifying
affinity
matrices that showadsorption
specificitytowards one
particular
DNApolym-erase as
compared
withanotherormatrices thatshowwidely varying affinities toward different
DNApolymerases.
An alternative approach to enzyme
purifica-tion in
general
istouse aspecific
effector mole-cule with strong affinity to elute the enzyme from anion-exchangeorsimilaradsorption ma-trix. Substrates, inhibitors, and other effectors bind toenzyme molecules andproduce
substan-tial changes in protein conformation
(often
changing proteinadsorption characteristics
to-ward the chromatographic media), resultingin
elution (5, 8). We examined whether such an
approach was possible for the purification of
mammalianviral reversetranscriptases.
Polyguanylic acid[poly(G)] wasshown to be
a potent inhibitor ofthe RNase H
activity
as-sociated with the reversetranscriptase moleculeofRauscher murine leukemia virus (R-MuLV)
and simian sarcoma virus (6). In comparison,
polyadenylic acid [poly(A)], polyuridylic acid
[poly(U)], and polycytidylicacid were only
min-imally active or not at all active against this
enzyme (6). Our interpretation of this finding
hasbeenthatthe reverse transcriptase molecule
hasa very strong and possibly specific interac-tion with poly(G). We report here that this
interactionwith poly(G) has proved to be very
useful in developing simpleaffinity procedures
for thepurification ofmammalian viral reverse
transcriptases.
A nucleic acid-free extract of R-MuLV was
prepared from 8 x 1011 virusparticles (2.7 mg)
andappliedto a5-ml column of
phosphocellu-lose. After washing with 0.1 M NaCl until no
moreproteinseluted,thecolumnwas
developed
with 20 ml of 50,uM poly(G) (minimum size,
6S;
Miles Laboratories) in the presence of 0.1 M NaCl plus 0.5 mM
MnCl2.
The eluates were collected in 1-ml fractions andassayed
forre-versetranscriptaseactivitywith
(dT)_15. (A).
as 555on November 10, 2019 by guest
http://jvi.asm.org/
556 NOTES
the
primer-template. Any
residualreversetran-scriptase
activity
on thephosphocellulose
col-umn wasrecoveredbyastepelution with 0.5 M
NaCl[withnoadded
poly(G)].
Figure
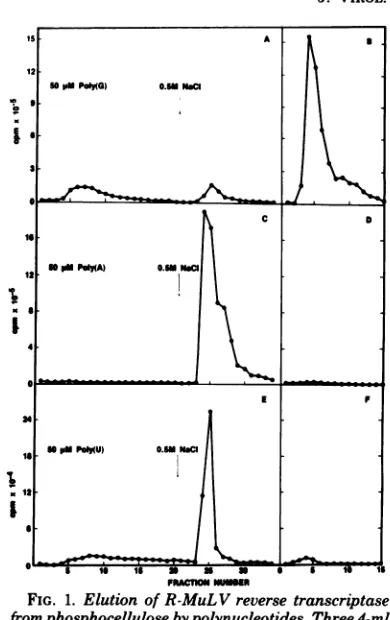
1A showsthat the
poly(G)
eluateyielded only
a small amountofenzymeactivity,
and yetnotmuch of theenzymeactivity
remainedonthephospho-cellulose column to be eluted
by
the high-saltwash.Thereasonfor thisapparentlowrecovery
wasthe fact that
poly(G)
has a strongaffinity
for thereverse
transcriptase
protein and that itinhibits thereverse
transcriptase
activity (9)
asit inhibitsthe RNaseH
activity (6).
Tomeasurethe true reverse
transcriptase
activity
in thepoly(G)
eluates,
thepoly(G)
hadtoberemovedfrom these fractions. The
poly(G)-eluted
frac-tionswere
pooled
andloadedonto a1-mlcolumnof DEAE-cellulose. All the enzyme
activity
bound to the
column,
confirming
that theen-zymethatwaseluted from
phosphocellulose
didnot exist as free enzyme
molecules,
but wascomplexed with
poly(G).
It is known that freereverse
transcriptase
does notbind toDEAE-celluloseatthe salt concentration
(0.1
MNaCl)
which was present in the
poly(G)
eluates(4).
The reverse
transcriptase
wasselectively
re-covered fromDEAE-cellulose
by
abatch elutionwith 0.3 M NaCl which dissolved the
poly(G)-reverse
transcriptase
complex,
butdidnotelute thepoly(G). Figure
1B shows that theenzymeappearedas a
sharp peak
inaconcentrated formrepresenting an increase in
activity
ofseveral-fold compared to the
poly(G)-containing
frac-tions
(Fig. 1A).
Theseseries ofexperiments
dem-onstrated that
poly(G)
elution removedmostofthe reverse
transcriptase
activity
bound to aphosphocellulose column. There was only a
small amountof
activity remaining
onthecol-umn after the
poly(G)
elution that could berecovered
by
ahigh-salt
wash(Fig.
1A).The elution of reverse transcriptase from
phosphocellulose involvedaspecific interaction
between theenzymeandpoly(G), and this
inter-actionwasstrongenoughtooffset theaffinity of
theenzymefor
phosphocellulose.
Thesubstitu-tion of eitherpoly(A) orpoly(U) forpoly(G)as an eluant did not elute reverse transcriptase
fromphosphocellulose;
furthermore,
noactivitywasfound in theeluatesevenafterthe
polynu-cleotides wereremoved by subsequent
DEAE-cellulose
chromatography(Fig.1C,D, E,and F). Allenzymeactivity could be recovered, however,by subsequent elutionwith 0.5 MNaCl(Fig.1C
andE),orby50,Mpoly(G) (datanotshown).
Since reverse transcriptases bind to
matrix-attached nucleic acids in general, and in fact,
poly(U)-Sepharoseis acommonlyusedaffinity
adsorbent for reversetranscriptase,theinability
ofpoly(U) and poly(A) to elute
phosphocellu-v
I
I
I
FRACTnONVUSER
FIG. 1. Elution of R-MuLV reverse transcriptase fromphosphocellulose bypolynucleotides. Three 4-ml samplesof nucleic acid-freeextractofR-MuL V, pre-paredasdescribed elsewhere (M. Robert-Guroff, V. S.Kalyanaraman, and M.G.Sarngadharan, Int. J. Cancer,inpress) from8x1011 virusparticles(2.7mg ofprotein),weredialyzedagainst50mM Tris-hydro-chloride, pH 8, containing1 mMdithiothreitol, 20% glycerol,0.02mMphenylmethylsulfonylfluoride, and 0.05% Triton X-100(buffer A) and applied to three 5-ml columns ofphosphocellulose equilibrated with bufferA.Afterthecolumnswerewashed withbuffer
A containing0.1M NaCl until no additional UV-absorbing materialwaseluted (25 to 30ml), they were developed with20mlofa 50
,uM
solution ofpoly(G) (A),poly(A) (C), or poly(U) (E) in buffer A containing0.1 MNaCl and0.5mMMnCl2. Subsequently, the columns were washedfurther with 15 ml of 0.5 M NaClin bufferA to recover anyremaining reverse transcriptase activity. Aliquots of10ulfrom the1-ml fractions collected were assayed for reverse
transcrip-taseactivity (Robert-Guroff et al.,inpress). The
frac-tions frompolynucleotide elutions were separately pooledandapplied to1-micolumns of DEAE-cellu-lose equilibrated with Tris-hydrochloride, pH 7.9, containing 1 mM dithiothreitol, 20% glycerol, 0.02 mMphenylmethylsulfonyl fluoride, and 0.05% Triton
X-100 (buffer B), and any enzyme in the pools re-coveredfreeof thenucleotides in a concentrated form by elution with0.3 MNaCl in buffer B (B, D, and F). Theprimer-template used for reverse transcriptase assaywas(dT)_15.(A)n in all cases, except in fractions containing poly(U), in which case (dG)
(QC),
wastheprimer-template.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.504.261.456.61.371.2]lose-bound reverse transcriptase indicated that
the affinity ofreverse transcriptase forpoly(A)
and poly(U) was weaker than its affinity for
phosphocellulose, and much weaker than the
affinity for poly(G). One would predict,
there-fore,that poly(G)should elute reverse transcrip-tase bound topoly(U)-Sepharose.To verifythis
prediction, a nucleic acid-free extract of
R-MuLV wasappliedtoa 5-ml column of
poly(U)-Sepharose (Pharmacia Fine Chemicals, Inc.).
After the column was washed with buffer
sup-plemented with0.2 MNaCl until no more
pro-teins eluted, it was developed with 50 yM
poly(G) in the wash buffer. Fractions of 1 ml
werecollected, andthe reverse transcriptase
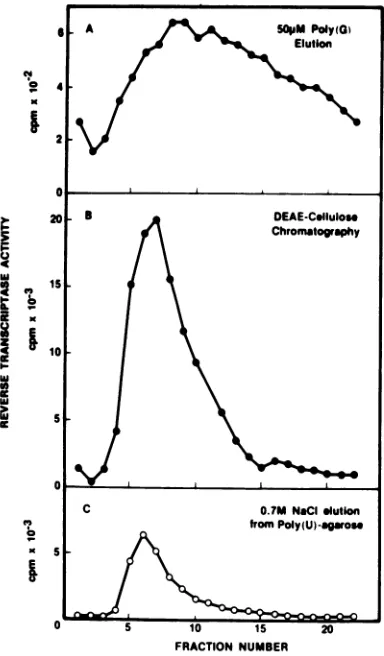
ac-tivitiesweredetermined(Fig. 2A). Atremendous
increase in enzyme activity was observed once
again when poly(G) was removed from the
eluted reverse transcriptase fractions by
chro-matographyon aDEAE-cellulose column(Fig.
2B). After thepoly(G) elution, the residual
re-verse transcriptase activity on poly(U)-Sepha-rose wasrecovered byasubsequent0.7 MNaCl
wash (Fig. 2C). A comparison of the results
showninFig.2B andConce again indicatesthe
effectiveness of
poly(G)
to elute matrix-boundreversetranscriptase.
In theseinstances, thereversetranscriptaseis
adsorbed
(along
with otherproteins)onto asolidmatrix[phosphocelluloseorpoly(U)-Sepharose]
and elutedwithadilute solution ofpoly(G).Two
levelsofspecificityareinvolvedinthese
proce-dures, the first at the adsorption step and the
second attheelution step, and therefore these
procedures haveadvantagesoverthose
employ-inga
nonspecific
salt elutionsteptorelease theenzymefrom these adsorptionmatrices. Useof
substrates, inhibitors, and other effectors to
elute enzymes from
adsorption
matrices isknown
generally
toresultinsubstantialenzymepurification (5, 8)because of the
high
specificity
involved.
Thebasis of theprocedures described above
was the formation of a
high
affinity complex
between reverse
transcriptase
andpoly(G)
whosecharacteristicswere
significantly
different from theproperties
of the free enzyme. Thecomplex
hadaloweraffinity
tophosphocellulose
and
poly(U)-Sepharose
andahigher affinity
toDEAE-cellulose than didthefreeenzyme.Since
poly(G) didnotelute otherproteinstoany
sig-nificantdegreefrom
phosphocellulose,
asjudged
from the protein
profile
when asample
wasanalyzed by
sodiumdodecyl
sulfate-polyacryl-amide
gel
electrophoresis (data
notshown),
itwas reasonabletoassume that the property of
forming this
complex
withpoly(G)
wassome-what specific to reverse
transcriptase.
On thebasis of this
rationale,
weattempted
thefollow-24'
'
21
0
20' DEAE-C.lluloo
Chromatography
Is
C15
00
w
C O.7MNaCIglutton
from
Poly(U)-gaos
5 10 15 20
FRACTION NUMBER
FIG. 2. Elution of R-MuLVreverse transcriptase bypoly(G)fromapoly(U)-Sepharose column. A
nu-cleicacid-free extractfrom 7x 1011particlesof R-MuLV(2.5 mgofprotein)wasdialyzed againstbuffer C(bufferBplus1mMMnCI)andappliedto a5-ml
columnofpoly(U)-Sepharoseequilibratedwithbuffer C.After washing with 30mlofbuffer Ccontaining
0.2MNaCl,thecolumnwasdevelopedwith30mlof
50 LMpoly(G) inbufferCcontaining0.2MNaCl(A),
and the eluates were collected into 1-mlfractions. Thesefractions werepooledand chromatographed
onDEAE-cellulose (B) to removethepoly(G) as de-scribed in Fig. 1. After the poly(G) elution, the poly(U)-Sepharose column was washed with 0.7M
NaCl in buffer Cto remove the residualactivity of reversetranscriptase(C).
ing
simplified
scheme topurifyreversetranscrip-tase from R-MuLV. Anucleic acid-free extract of R-MuLV(10mg of
protein)
wasincubated in ice for 5 min with 50,M poly(G) and 0.5 mMMnCl2
and then applied to a 1-ml column ofDEAE-cellulose.
Thecolumnwaswashedexten-sivelywith0.1MNaClto removeunbound and
weaklyboundmaterials. Under these
conditions,
all thereversetranscriptasemolecules existedas
acomplexwithpoly(G)andtherefore remained
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.257.449.65.393.2]558 NOTES
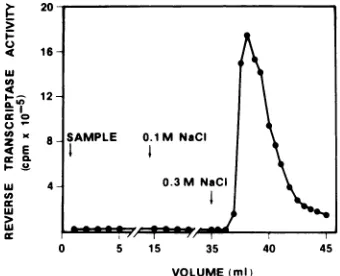
boundtotheDEAE-cellulose column. When the NaCl concentration in the bufferwasraised to 0.3 M, the complex betweenreverse
transcrip-tase and poly(G) was broken and the reverse
transcriptase emerged from the column as a sharp activitypeak, leaving poly(G)still bound tothe column (Fig. 3). The recovery of enzyme
activity fromthe columnwasgreater than 70%.
When
'25I-labeled
envelope glycoprotein(gp7O)
orthemajorstructuralprotein (p30)of R-MuLV
wasmixedwithpoly(G)andappliedto a
DEAE-cellulosecolumn under the conditions described
above,therewas noretention of the radiolabeled
protein on the column, indicating that these
proteins did not form stable
complexes
withpoly(G). Further, when
'25I-labeled
gp7O ofR-MuLVwasmixed with anucleic acid-free virus extract and the mixture was incubated with
poly(G) and
chromatographed
onDEAE-cellu-lose, thelabeled gp7Owasrecovered in the
un-retardedflow-throughfractions and thereverse
transcriptasewasadsorbedtothe column (data
notshown).
The simple method outlined above for the
purificationofreverse
transcriptase
isessentially
asingle-step
procedure,
and therefore is fast andconvenient. It was of interesttodetermine the
purityof the enzyme obtained
by
thisprocedure.
Forthis purpose, the
purified
reversetranscrip-tasefrom theDEAE-cellulosestepwas
radiola-u I-4
U)
4
I-U) U
z 4E
uL
LU LU
cc
FIG. 3. Chroma of poly(G) and R DEAE-cellulose.A NaClfrom 3.5x10 threefoldwithbuff
with50 4Mpoly(G, wasthenapplied t< -equilibrated with
with buffer B con transcriptasewas taining 0.3M Na( lected and sample scriptase activity plate.
beled with 125I usingthe chloramine-T method (1), and the labeled protein was analyzed by
electrophoresis on a polyacrylamide gel in the presence of sodium dodecyl sulfate. The radio-activityprofile obtained on thegelis shown in
Fig. 4. The enzyme moleculemigrated withan apparent molecularweight of about 70,000 and
was substantially free ofother contaminating
proteins.
Theprocedure describedabove,therefore, in-volves the selectivecomplexingofreverse
tran-scriptase in a crude viral extract with poly(G)
and capturing thecomplex, and only the
com-plex,onDEAE-cellulose (Fig. 3). Since thefree
viral proteins, includingfree reverse transcrip-tase, donotbindtoDEAE-celluloseatthesalt
concentrations employed, the retention of
re-verse transcriptase is solely dependent on the
E
GELSLICENUMBER
E 0.1 M NaCI FIG. 4. Electrophoreticprofile ofiodinatedreverse transcriptase on asodiumdodecyl sulfate-polyacryl-amide
gel.
Asample of
the reverse transcriptase, 0.3 M NaCI purifiedasdescribedinFig. 3, was labeled with125iby themethodof Greenwoodetal. (1). A 100-,ulportion
of
thereaction mixture contained0.5to2,ugof
the 5 15 35 40 45 enzyme,50 mM sodiumphosphate (pH7.5),30,ug ofchloramine-T,250mM
NaCl,
0.1%TritonX-100,andVOLUME(MI) 0.5mCiofNa'25. After1 min at room temperature, ,tography of a preformedcomplex the reaction was terminated by the addition of 50 ,ig
t-MuLV reverse transcriptase on of sodium metabisulfite and 20
til
of5MNaCl. The Inucleic acid-free extract in 0.3 M iodinatedprotein was thenseparated from unincor-12particlesofR-MuLVwas diluted porated 1251 on a BioGelP-10column equilibrated 'er B and incubated in ice for 5min with1MNaCl, 10 mM sodiumphosphate buffer (pH )and 0.5 mMMnCl2.Thecomplex 7.5), 10%, glycerol, and 0.2 mM phenylmethylsulfonyloa1-mlcolumn ofDEAE-cellulose fluoride. The labeled protein was subjected to
electro-bufferB. After extensive washing phoresis on aI
010
polyacrylamide gel in the presence itaining 0.1 MNaCl, the reverse of sodium dodecyl sulfate bythe method ofLaemmlieluted with 10 ml of buffer B con- (3). The gel was divided into1-mmslices, and their Cl. Fractions of 0.65 ml were col- radioactivity was determined in a gamma counter. es were assayed for reverse tran- The standard molecular weight markers run in par-with (dT)-15 '(A), as primer-tem- ellel gels were: P, phosphorylase; B, bovine serum
albumin; 0,ovalbumin; and C, chrymotrypsinogen.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.275.445.258.444.2] [image:4.504.74.244.383.521.2]complex formation. For thisreason, the size of
the DEAE-cellulose colunm is
independent
oftheamount of viral
proteins
in the extract,butisdeterminedbythe amountof
poly(G)
present.Routinely,aslittleas 1ml ofDEAE-cellulose is
enoughtoretain50mlof50,uMpoly(G),making
thisprocedure
extremely
attractiveforachieving
atremendous concentrationofreverse
transcrip-taseduring this step. Unlike mostother
proce-dures,thisprocedure
effectively
eliminates theneedtohandlereversetranscriptaseatlow
pro-tein
concentrations,
because theonly
stepsin-volved are (i) the
preparation
ofnucleicacid-freeextractsfrom virus concentrates, whichdoes
not expose reverse transcriptasetolowprotein
concentrations,
and(ii)
thechromatography
of thecomplex
of reversetranscriptase-poly(G),
which yields the freeenzymeinaconcentrated
form.
LITERATURE CITED
1. Greenwood,F.C.,W. M.Hunter,and J. S. Glover. 1963. The preparation of"1'I-labeledgrowthhormone
ofhigh specific activity. Biochem. J. 89:114-123. 2. Kacian, D. I., K. F. Watson, A. Burny, and S.
Spie-gelman.1971.Purificationof the DNA polymerase of avian myeloblastosis virus. Biochim. Biophys. Acta 246:365-383.
3. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
4. Lewis, B.J., J. W.Abrell, R. G. Smith, and R. C. Gallo. 1974. DNA polymerases in human lymphoblastic cellsinfected with simian sarcoma virus. Biochim. Bio-phys. Acta349:148-160.
5. Pogell, B. M., and M. G.Srngadrharan.1971. Specific elution with substrate. MethodsEnzymol. 22:379-385. 6. Sarngadharan, M. G., V. S. Kalyanaraman, and R.
C.Gabo. 1978. Inhibition by RNA of RNase H activity associated with reverse transcriptase inRauscher mu-rineleukemia virus cores. J.Virol. 27:568-575. 7. Sarngadharan, M. G., M.Robert-Guroff, and R.C.
Gallo.1978.DNApolymerasesof normal and neoplas-ticmammalian cells. Biochim.Biophys. Acta 516:419-487.
8. Sarngadharan,M. G.,A. Watanabe, and B. M. Pogell. 1970.Purification of rabbit liver fructose 1,6-diphospha-taseby substrate elution. J. Biol. Chem. 245:1926-1929. 9. Waters, L C., and W.-K. Yang. 1974. Comparative biochemical propertiesof RNA-directed DNA polym-erases fromRauscher murineleukemia virus and avian myeloblastosisvirus. Cancer Res. 34:2585-2593.