Vol.38, No. 2 JOURNAL OFVIROLOGY, May1981,p.430-437
0022-538X/81/050430-08$02.00/0
Genome Structure
of
Avian Sarcoma
Virus Y73 and
Unique
Sequence
Coding for
Polyprotein
p90
MITSUAKI YOSHIDA,`* SADAAKI KAWAI,2 AND KUMAOTOYOSHIMA2
Departmentof Viral Oncology,CancerInstitute, Toshima-ku,' andDepartmentof Oncology,Instituteof
Medical Science, University of Tokyo, Minato-ku,2 Tokyo, Japan
Received 7November1980/Accepted 23 January 1981
Thegenomestructureofanewly isolatedsarcomavirus,Y73,wasstudied. Y73
isadefective,potentsarcomagenic virus and contains 4.8-kilobase (kb) RNAas
itsgenome;incontrast,helper virusassociated with Y73 had 8.5-kb RNA, similar
toother avianleukemia viruses. Fingerprinting analysis of these RNAs
demon-strated that the 4.8-kb RNA containsaspecific RNAsequence of 2.5kb, which
representsthetransforminggene(yas) of Y73.Thisspecificsequence wasmapped
in the middle of the genome and had at both ends 1- to 1.5-kb sequences in
commonwith Y73-associated virus RNA. Thisstructureisverysimilartothose
of avian and mammalian leukemia viruses. Invitro translation of the 4.8-kb RNA
and theimmunospecificity of the products directly demonstrated that polyprotein
p90,containing p19, isaproduct translated from capped 4.8-kb RNA and that the
specific peptide portion iscoded by theyas sequence.Protein 90,whichwasalso
found in cells transformed withY73,wassuggestedtobeatransformingprotein.
Amongavianretroviruses,Rous sarcoma virus
(RSV) has been most thoroughly studied
be-cause of itscompetencyinreplication and
sar-comagenic transformation. The sarcoma gene
(src)in RSV codesaprotein,p6OBrc (4, 21),which
has protein kinase activity
phosphorylating
ty-rosine residues in target proteins (5, 10).
Re-cently, we (27) reported that a newly isolated
avian sarcoma virus, Y73 (11), contains a
sar-comagenic transforminggene (yas) showingno
homology with the src sequence ofRSV. Fur-thermore, aspecific protein, p90, was found in
Y73-transformed cells and shown tohave
pro-tein kinase activity, which phosphorylated
ty-rosine residues in p90itself and in heavy-chain molecules ofimmunoglobulinGcomplexedwith
p90 (13). Fujinami sarcoma virus (7) was also recently reported to have a distinct class of transforminggene (8, 16).
Independent isolates of avian sarcoma viruses
RSVandB77 are competent inreplicationand
havevirtuallythe same genomestructure: 5' gag
polenvsrc 3' (26). On the otherhand,allavian
acuteleukemia viruses are defective in
replica-tion,deleting thepoland env genes and part of
thegag gene, instead of the region containing
the transforming gene in the middle of the
ge-nome(3, 6,9, 18,22).Since Y73 is a sarcomagenic
virus, but is defective in replication (11), it
seemed interesting to determine its genome structure.
Inthis work, wecharacterized the Y73 genome
by RNaseT1 fingerprintinganalysis and in vitro
translationof thegenome RNA. Results showed
that thestructural features of the Y73 genome are
essentially
thesame asthose of genomes ofotheracuteleukemia viruses and that polypro-tein p90 is a gene product of the specific
se-quence(yas),suggestingthatp90is a
transform-ingprotein.
MATERLALS AND METHODS
Cells and viruses. Chicken embryo fibroblasts
wereprepared from C/Oor C/BE chickenembryos
andculturedasdescribed before(28).Y73 viral stock
wasgrownasdescribed previously (13), and
Y73-as-sociated virus(YAV) wasisolatedbytwosuccessive
limiting dilutionsonchickenembryofibroblasts and
confirmedtohavenofocus-formingabilityunder the
standardagarlayer.
Viralpreparations were harvested every 12hand
purified byprecipitationwith ammoniumsulfate and
by successive discontinuous and continuous sucrose
gradientcentrifugations asdescribedpreviously (30).
Labelingof viral RNA with[32P]phosphatewascarried
outexactlyasdescribedpreviously (30).
Preparation ofviral RNA. Virion RNAwas
ex-tractedby proteinase treatment followed by phenol extraction (30). Virionsweredisrupted by adding0.1 volumeof10%sodiumdodecyl sulfateinthepresence of 100
,Lg
ofproteinaseKper ml. After incubation for30minat37°C,RNAwasextracted three times with
phenol-chloroform (6:4) and then precipitatedwith 2
volumes of ethanol.RNAwasdissolvedin0.1%sodium
dodecyl sulfate and heated at 100°C for 1 min and
then fractionatedon a gradientof15 to30%sucrose
containing50mMTris-hydrochloride (pH 7.5),0.1M
NaCl, and 1 mM EDTA by centrifugationat 45,000
rpm for3h inaBeckmannSW50.1rotor at 15°C. In 430
on November 10, 2019 by guest
http://jvi.asm.org/
GENOME STRUCTURE OF Y73 431
[image:2.500.137.371.411.582.2]someexperiments, RNA containingpolyadenylicacid
[poly(A)] wasseparated by passing the preparation
through acolumnof oligodeoxythymidylic
acid-cellu-lose as described before(31).
Fingerprinting analysis of[32PJRNA.
Two-di-mensional gelelectrophoresis was used for
fingerprint-ing RNase Ti-resistant oligonucleotides. The
proce-dure wasessentiallyas described by Lee and Wimmer
(17).Oligonucleotides separated on gels were extracted
with 0.5 ml of water, lyophilized, and digested with
RNase A (0.1mg/ml). The digests were analyzed by
electrophoresis at pH 3.5 onDEAE-cellulose paper as
described by Barrell (1).
Invitro translation of viral RNA.
mRNA-de-pendentreticulocyte lysate was prepared as described
by Pelham and Jackson (20) and used to translate Y73
RNA asdescribed previously (29). Each reaction
mix-ture(12
pl)
contained 0.2jlg
of viral RNA and 20ACi
of [3S]methionine in addition to the standard
re-agents. Twomicroliters of the reaction mixture was
usedforgel electrophoresisin thebuffer described by Laemnmli (15), and the gels were dried for
autoradiog-raphy orfluorography. Cells were labeled with
[3S]-methionine as described previously (31), and immu-noprecipitation was carried out according to Kessler (14).
RESULTS
RNA of Y73. Since the original Y73 stock
containedalarge excessofYAVofsubgroupA ashelper (11), the transformed cellswerecloned
in softagarmedium. Virusesreleased from the selected producer cellswerelabeled with
[32P]-phosphate, and the RNAwasanalyzed by aga-rose gel electrophoresis. As shown in Fig. la,
a b c
I
8.5kb-28S
-
48kb-0
x
E
a
18S-most of the RNA containing poly(A) migrated
as high-molecular-weight aggregates, probably
50-70S dimers. After heat treatment at 1000C
for 1 min, the aggregates were dissociated into
two RNA species: 4.8-kilobase (kb) and 8.5-kb
RNA components (Fig. lb). The 4.8-kb RNA
extracted from the gel sedimented as 26S on
sucrosegradient centrifugation (Fig.le),and the
RNAwassmaller thanthose ofknown defective
acute leukemia viruses (3, 6, 12, 18). On the
otherhand, 8.5-kb RNA sedimentedas35S,and its mobility in the gel was the same as that of
RNA of transformation-defective RSV. From these results, and from the findings that
non-transformingYAV isolated bylimiting dilution contained only 8.5-kb RNA and that the4.8-kb
componenthasaspecificsequence, asshownin
thispaper, we concluded that the 4.8-kb RNA
species isthe genome ofY73 carrying the
sar-comagenic transforminggene.
Analysis of Y73 RNA byfingerprinting. The relationship between 4.8-kb and 8.5-kb
RNAs wasanalyzed by
fingerprinting
RNaseT1-resistantoligonucleotides. 32P-labeled 4.8-kband
8.5-kbRNAs wereseparated byagarosegel elec-trophoresis andselected forpoly(A)-containing
RNA on oligodeoxythymidylic acid-cellulose.
The fingerprint of the 8.5-kb RNA (Fig. 2D) showed that thehelper 8.5-kb RNAwas asingle
component, and asexpected, the fingerprintof
4.8-kb RNA contained thespecific
oligonucleo-tides thatwere missing in that of 8.5-kb RNA,
f ract'ion no.
FIG. 1. Analysis ofY73 RNA byagarosegelelectrophoresis (a, b, c)andsucrose
gradient
centrifugation
(d, e).[32P]RNAofY73 virionscontainingpoly(A) before (a)andafter (b)heattreatment at1X0°C for1 min
and[32PJRNAoftransformation-defective PragueRSVwere
subjected
toelectrophoresis
in 2% agarosegel
in40mMTris-hydrochloride (pH 8.0),20mM sodium acetate, and1mM EDTA.After
electrophoresis,
RNAswereextractedfromthe bandscorrespondingto4.8-kb and 8.5-kb RNA and
centrifuged through
15to30%sucrosegradientscontaining20mMTris-hydrochloride(pH 7.5),0.1MNaCi, 1mMEDTA,and 0.2% sodium
dodecyl sulfatein anSW50.1 rotorat45,000rpmand15°C
for
180min. (d) 8.5-kbRNA; (e)4.8-kb RNA.Dotted linesare[3H]rRNAaddedasinternal markers..
VOL. 38,1981
on November 10, 2019 by guest
http://jvi.asm.org/
432 YOSHIDA, KAWAI, AND TOYOSHIMA
00:.-fE-;S.-Sffi-.
:00'tV,0000000..00.:00077.?
A 00 ;f}0 -u:0- 0-0 SS00: :00 000
:E. :fX
FIG. 2. RNaseT, fingerprinting analysis ofY73 RNA.[32PJRNA of Y73wasfractionatedinto26S(A) and
35S(D)fractions,and eachfractionwastreated with weak alkalitointroduceoneortwo cutsin the RNA
molecule,asdescribedpreviously(27). Then RNA fragments containing poly(A)wereseparatedon
oligodeox-ythymidylicacid-cellulose andfractionatedintosixfractionsbysucrosegradient centrifugation. Eachfraction
wasrecentrifuged, and thepeak fractionwasdigestedwith RNaseT,andfingerprinted by two-dimensional
gelelectrophoresis. (A) 26S RNA;(B) 18-20S RNAfrom26SRNA; (C) 5-14S RNA fromt 26S RNA; (D) 35S
RNA; (E)19-25S RNAfrom35SRNA;(F) 5-14SRNAfrom 35S RNA.
indicating that 4.8-kb RNA contained the
spe-cific nucleotide sequences (Fig. 2A). All ofthe
oligonucleotidespotsfound in the fingerprintof
8.5-kb RNA were detected in that of 4.8-kb
RNA, butmostofthemwereminorcomponents.
These minorspotswereprobablycontamination
ofthe 4.8-kb RNA fraction with fragments of
8.5-kb RNA. To verify this explanation, each
spotwascut outfrom thegelsand quantitated
by counting the radioactivity of32P. Since the
accuratelength of eacholigonucleotidewasnot
known, the ratio of the yield of each
oligonu-cleotide from the4.8-kbRNA fraction to thatof
the corresponding spot from 8.5-kb RNA was
used to estimate the apparent molar
yield
ofeacholigonucleotide, assumingthatpure8.5-kb
RNA
gaveeacholigonucleotide
inamolarratio.Inthe case ofspotsspecificto4.8-kbRNA, the
yield ofoligonucleotides of 8.5-kb RNA which
were similar in size was used to calculate the
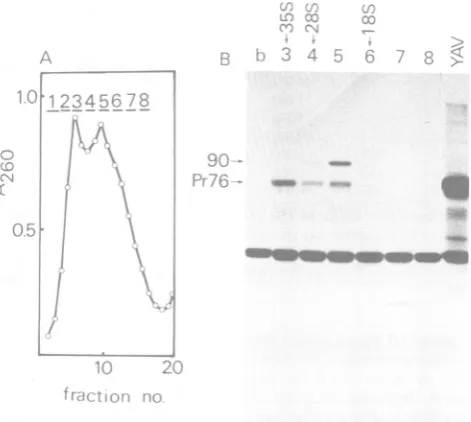
ratio. TheresultsaresummarizedinTable1.
All
spots wereclassifiedinto three groupsby their
yield ratio,thatis,less than0.5, between0.8and
1.2,andmorethan1.4.
Oligonucleotides
showingJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.500.77.438.76.473.2]VOL. 38,1981
TABLE 1. Yi
oligonucleotidesfS
Spots in 26S RNA fraction No. Yield (cpm) 1 775 2 736 3 295 4 985 5 291 6 126 7 978 8 230 9 201 10 745 11 168 12 494 13 779 14 556 15 192 16 139 17 353 18 320 19 185 23a+b 867 24 389 25 131 27 201 28 216 29 806 30 164 31 616 32 142 38 564 39 575 40 101 41 227 42 568 43 607 44 320 45 500 46 164 47 177
GENOME STRUCTURE OF Y73 433
'ields ofRNaseTi-resistant
rom26S and 35S RNAfractions
Spots in 35S RNA
fraction Ratio 26S/
No. Yield 35S (cpm)
(1.03)a (0.98)
51 753 0.39
52 643 1.50
53 779 0.37
(0.18)
54 702 1.39
55 502 0.46
56 491 0.41
57 553 1.35
58 441 0.38
(1.12)
59 562 1.38
(1.19)
60 466 0.41
61 379 0.36
(0.91) (0.82)
62 388 0.47
65 466 1.86
(1.08)
66 358 0.36
68 536 0.37
69 524 0.41
70 542 1.49
71 486 0.34
72 367 1.67
73 486 0.29
80 393 1.43
81 355 1.61
82 234 0.43
(0.97)
(1.06) (1.13) (0.83)
77 383 1.30
78 415 0.39
79 397 0.44
'Theratio ofoligonucleotides specificto the 26S RNA fractionwascalculated by using the yield (counts
per minute; cpm) of oligonucleotides of 358 RNA,
whicharesimilar insizetothespecificone;this ratio
isshown inparentheses.
aratio of 0.8 to 1.2 were all specific to 4.8-kb
RNA, and all otheroligonucleotides showinga
ratio of less than 0.5 or more than 1.4 were
confirmed by RNase A digestion to have the
samenucleotidecompositionasthe
correspond-ingspots obtained from 8.5-kb RNA (data not
shown). From these findings it was concluded
that 22 oligonucleotides produced ataratio of
morethan 1.4(10 spots)andbetween0.8 and 1.2
(12 spots) originatedfrom 4.8-kb RNA andthat
the restofthe minor spotsproducedataratio
of less than 0.5 represented 8.5-kb RNA
frag-mentscontaminatingthe 4.8-kb RNAfraction.
Spot
no.6 showedavery lowyieldalthough
itwas
specific
tothe 4.8-kbRNAfraction. Thisspot
might
beexplainedbyamicroheterogeneity
in the sequence of 4.8-kb RNA,but thereason
isunknown.
Thus,
spotno.6wasremovedfrom thefollowing
considerations.Analysis
ofthe nucleotide composition of 22oligonucleotides
originating
from 4.8-kb RNAsuggested
that 11oligonucleotides
werespecific
and
produced
inaratio of 0.8to1.2andthat 10oligonucleotides were shared with YAV 8.5-kb
RNAandthuswereproducedat aratio ofmore
than 1.4
(Table 2). Assuming
a random distri-butionoftheoligonucleotides
alongthe genome,the results suggest that about half the 4.8-kb
sequenceis
specific
to4.8-kbRNA.For determination ofthe location of the
oli-gonucleotides
specificto4.8-kbRNA within thegenome, RNA
fragments
containingpoly(A)
were fractionated and purified bysucrose
gra-dient centrifugation, followed by
recentrifuga-tion of eachfractionandfingerprinting (Fig. 2B
and
C).
Althoughthe 4.8-kb RNA fraction wascontaminated with fragments of 8.5-kb
RNA,
only
22spots, whichwere concludedtobepro-duced from 4.8-kb RNA (Table 2), were
fol-lowed,
and the otherspotswereignored.Thus,
atentativeoligonucleotidemap wasconstructed
(Fig. 3).
YAV 8.5-kb RNAwasanalyzedsimilarly
(Fig.
2E and F), and the two oligonucleotide [image:4.500.54.245.98.497.2]mapswerecompared. Tenoligonucleotideswere commontoboth
RNAs,
and 4 ofthese 10wereTABLE 2. Qualitativeanalysis ofRNaseT,
oligonucleotides of Y73 26S RNA and YA V 35S RNA
Sptn.Spot
no.ofY73
oSf
YA RNase A digestion products 26S RNA RNA1 U,C,G
2 U,C, AC, G
4 52 U,C, AC, AU, AAG 6 U,C, AC, AU, AAAU,G
7 54 U,C,AC,AU,AAC,AAG
10 57 U, C, AAG, AAAU 12 U, C,AC, AG, AAAU 13 59 U,C,AU, AG,AAC, AAAC
14 U,C,AC,AU,AAC,G
17 U, C, AC,AU,AAC, AAU, G 18 C, AC, AU, AAC, AAU, AAAC, G 23 a + b 65? U,C, AC, AU, AG, AAC, AAAG
24 C,AC, AG, AAAC
29 70 U,C,AC,AU, G 31 72 U,C,AC,AU,AAC,G 38 80 U, C,AU,AAU,G 39 81 U, C,AU, G
41 U, C,AU, AG
42 U, C, AC,AU, AAU, AAAN,a G 43 U, C,AC,AU, AG,AAC,AAU
44 U, C,AC, AU, AG 45 77 C, AU, AAU,AAAC, G
aN, Not identified.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.500.258.450.409.665.2]434 YOSHIDA, KAWAI, AND TOYOSHIMA
mapped within about 1kb adjacent topoly(A)
atthe 3'end;the other6 were detected
only
inthe
poly(A)-containing
RNA fragments largerthan20SfortheY73and 30S for
YAV,
andwerethusmappedwithin about 1 to 1.5kb in the 5'
regionsof the genomes. The
remaining
oligonu-cleotides specificto4.8-kbRNAweremapped
inthe middleportionof the 4.8-kb genome RNA.
These results suggest that the 4.8-kb specific
RNAsequence iscomposedofa continuous
se-quence of about 2.5 kb of RNA and is flanked at
the two endswith 1- to 1.5-kb RNAsequences
commonwith those ofhelperYAV RNA.
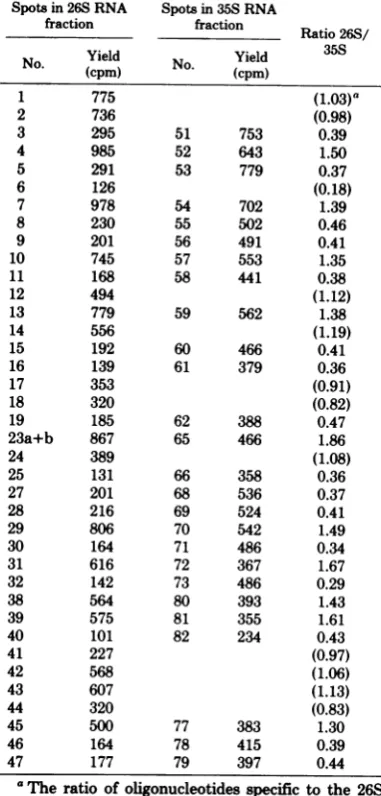
Invitro translationof Y73RNA. To
study
YAV / O X 0t1-0 J o 1f CNi
YAV ( Ln UINr), U" 0 00 i.0 D P.,1 r-...N- to LODto
the gene
products
codedby
the specificse-quence, poly(A)-containing RNA isolatedfrom
Y73 virions was fractionated by size (Fig.
4A)
andtranslated in an mRNA-dependent
reticu-locyte lysate (20). Polyacrylamide gel analysis
(Fig. 4B)
of thetranslationproducts showedthatPr76wasthe mainproduct fromRNAfractions
larger
than 28S, as found in previousexperi-ments on translation of avian erythroblastosis
virus RNA (29). This result was also expected,
becausethehigher-molecular-weightfraction of
RNA contained mainly helper YAV RNA in
which the gag gene is located near the 5'
ter-minus and theprotein-synthesizingsystemfrom
L1 1. _-. He_CANDoiox .1. U i
S
s
to r-Is
P. toWs
to A)>
5' I I I I C0Ci
8.5kb 8 7 6 5 4 3 2 1 0
4.8kb
Y73
CaPE2ZE=
I I 71;7yrovKzj :!(..oow-.o.oo~~~~~~~~~~~~~cNc%JXN-o0.0-.0~~~~~~~~~~~~~-r!.X- C4__ ____
( 4-tn X--_-CN M - xW "- -); a
.5
o90 Protein r
9 + p
-pl9+? specific polypeptide
FIG. 3. Physicalmapsofoligonucleotides and polypeptide codedby Y73.Spotnumberswithasingleline
areoligonucleotidesshared with4.8-kband8.5-kbRNAs, and those withadoublelinearespecificto4.8-kb
RNA. The ordersof theoligonucleotideswithin the respectiveparenthesesareuncertain.
A
(I) /) (I)
04
CI N oo
B b 3 4 5 b 7 8
2<
90-
Pr76-fraction no.
FIG. 4. Polyacrylamidegelelectrophoresisof thetranslationproducts of Y73 RNA and helper YA V RNA.
Y73 RNAcontaining poly(A) was fractionatedon asucrose gradient(A) asdescribedinFig. 1, and about 0.2
pgofRNAineachfractionwastranslatedin anmRNA-dependentrabbitreticulocytelysate, containing20
pCiof[35S]methionine, andanalyzedon a 7 to
15%o
polyacrylamide gradientgel(B). Track b had noaddedRNA;othertracks arelabeledwithnumberscorrespondingtothefractionsofRNAfromthesucrose gradient
in (A). TrackYA V is 0.5
pg
ofheat-treated70S RNAof helper YA V.OKIOP"& I
J. VIROL.
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.500.64.451.230.341.2] [image:5.500.141.377.386.597.2]VOL. 38, 1981
eucaryotic cells translates only one gene near
the 5'end of polycistronic viral mRNA (19, 21,
25).
IncontrasttoPr76,a productwithamolecular weight of about 90,000 (p90) was synthesized mostefficientlyfrom slightlysmallerRNAthan
28S (Fig. 4b). Since fractions of similar sized
RNAofYAV didnot support detectable
synthe-sis ofp90, the results indicatethatp90wascoded
by the 4.8-kb RNA of Y73. The effect of
7-methyl-GTP, a cap analog, onthe synthesis of
p90wastestedtoknowwhetherp90was
synthe-sizedby intact cappedRNA orfragmentedRNA
(2). As seen in Fig.5, thesyntheses ofp90and
Pr76were bothstrongly inhibited by the
addi-tion of7-methyl-GTP. On the other hand, the
amount of smaller minorproducts was not
re-ducedby addition of 7-methyl-GTP.These
find-ings strongly support the idea thatp90 is
syn-thesizedby intact 4.8-kb RNAwhich has a cap
structure atthe5'terminusandalsothatsmaller
productsaresynthesizedbyfragmentedRNA.
The in vitro product p90 was found to be
immunoprecipitated with serum against
dis-ruptedvirions ofB77 (Fig. 5).Sincethis antise-rummainly containedantibody to gag proteins,
p90 was suggested to contain amino acid
se-quences related togagproteins. Thisanti-B77
antiserum precipitated p90 from an extract of
cellstransformed with Y73,and, as seen in Fig.
5, no detectabledifference wasobserved inthe
molecular sizes of thein vivo and in vitro
prod-ucts. This stronglysuggeststhatthese twop90
proteinsareidentical. Whenanotherantiserum that had noantibody activity against p19 was
used, Pr76 andp27 wereprecipitated from
trans-formed
cells,
butp90
was not(Fig. 5). This result demonstrated thatp90
contained the amino acidsequence ofp19, butnotthat ofp27.However,
this
experiment
does not exclude the possiblepresenceofapartialsequenceofp27 in the
p90
molecule.
DISCUSSION
In this paper, we characterized the genome
structureof
Y73,
anewly
isolatedaviansarcomavirus
which, like RSV, induces fibrosarcomainchickens (11). The
transforming
gene(yas)
of Y73waspreviously
characterizedas a newtypeof sarcomagene with no
homology
withsrcofRSVor
fps
ofFujinamisarcomavirus (27). Inthepresentstudy, the yassequence, whichwas
characterized by
specific
oligonucleotides,
wasmapped in the middle
portion
of the genome, with portions of 1 to 1.5 kb common to the 3'and 5' ends ofhelperYAV RNAatthetwoends.
Thisstructural feature of the genome makes it
very similar tothe genomes of avianacute
leu-GENOME STRUCTURE OF Y73 435
Bh
[image:6.500.260.450.79.347.2]-p9O
~Pr76
FIG. 5. Comparison of in vitro and in vivo
trans-lationproductp90. (A)Effect of 7-methyl-GTP on in
vitrosynthesisofp90; 35S RNA (a, b)and26S RNA
(c, d) were translatedas described inFig. 4 in the
absence (a, c)orpresence(b, d) of 200 mM
7-methyl-GTP. The translationproducts in the absence of
7-methyl-GTP were immunoprecipitated with rabbit
antiserumagainst disruptedvirions,andprecipitates
were analyzedon a9%gel; (e) and
(t)
areimmuno-precipitated translation productsfrom 35S RNA and
26SRNA,respectively. (B)Extractsofcellsinfected
with YA V(g)orwith Y73(h,i)weremixed with
anti-gagserum, which hadanti-p19activity (g, h) or no
anti-p19activity (i), and theprecipitateswere
ana-lyzedon thegel.
kemiaviruses such asavian
erythroblastosis
vi-rus(3) or MC29 (18),tothose ofFujinami
sar-comavirus (8, 16), andtothose of mammalian
sarcoma-acute leukemia viruses
(23, 24).
Be-causeof theclose
similarity
of thegenomestruc-ture of Y73 to that of avian acute leukemia viruses,wehavetested its
leukemogenic
capac-ity in chickens. One-week-old chickenswere
in-oculated
intravenously
with Y73(1.5x104focus-forming units) or with avian
erythroblastosis
virus pseudotyped with Rous-associated virus
type 1 (104 focus-forming
units)
as control. Allchickensinoculated with avian
erythroblastosis
virusdeveloped erythroblastosiswithin 8
days;
however, no sign of leukosis was detected in
chickensinoculatedwith Y73
by
examinationofA
on November 10, 2019 by guest
http://jvi.asm.org/
436 YOSHIDA, KAWAI, AND TOYOSHIMA
peripheral blood at 3- to 5-day intervals up to 1
month.Most of these birds developed
subcuta-neous sarcoma at the site of intravenous
inoc-ulation within 2 to 3 weeks. Eight surviving
chickens were autopsied at 1 month; six had
sarcomas in one or multiple sites or internal
organs suchasspleen, lung, liver, ormuscle, in
additionto subcutaneous tumors.These
obser-vations unequivocally confirmed Y73 as a
defec-tive sarcoma virus. Thus, the
replication-com-petentavian sarcoma virusesRSV and B77 are
exceptionalin genome structure, and generally
the features of sarcoma-acute leukemia viral
genomes are defective, with the transforming
genelocatedin the middle of the genome.
Invitrotranslation of 4.8-kb RNAhasdirectly
shownthat apolyproteinof90,000daltons (p90)
containing p19 butnotp27 is aspecific
transla-tion product. The p90 was translated from a
gene near the 5'end of theintact genomeRNA
containing thecap structure. The presenceofan
approximately 1.5-kb sequence homologouswith the 5' region ofYAV RNA suggested thatp19 was the N-terminal polypeptide componentin
p90,because evenifalong untranslated region
suchas 0.5kbwerepresentin the5' region, the
RNA sequence coding for p19, an N-terminal
peptide in Pr76, could be included in the 5' region homologous with YAV RNA. Thus, a
polypeptide of about 71,000 daltons should be
codedbyaspecific nucleotidesequenceofabout
2.5kb andtherefore would beaspecific
polypep-tide.Thisconclusionwassupported by the
find-ings that anti-gagserum that had no anti-p19
activity did notprecipitate p90 andthatrabbit
antiserumagainst
p6Oarc
codedby Schmidt-Rup-pin strain RSV didnotcross-reactwithp90(13).Sincemostofthespecificsequence in the
4.8-kbgenomewouldbeusedforp90, p90seems to
bethe only polypeptide coded by the Y73
ge-nome. This possibility is consistent with the
finding that 4.8-kb RNA is the only species
detectable with specific complementary DNA
(cDNAya,)
incellstransformed with Y73. Theseconsiderations strongly suggest that p90 is
in-volved inthe mechanism ofsarcomagenic
trans-formation of infected cells. This prediction is
supported by the finding thatp90 detected in
cells
transformed
with Y73 had protein kinaseactivity for phosphorylation of tyrosineresidues
in
p90
itselfand inheavy-chain immunoglobulinGcomplexedwithp90(13). However, wedid not
exclude the possibleformation of smaller
prod-uctscodedby the Y73 and undetectable by our
methods.
ACKNOWLEDGMENTS
WethankR.Ishizaki forproviding rabbit antiserum, and
Y.Hirayama for excellent technical assistance.
This work was partly supported by a Grant-in-Aid for
CancerResearch from theMinistry of Education, Sciences andCulture of Japan.
LITERATURE CITED
1.Barrell, B. G. 1971. Fractionation and sequence analysis ofradioactive nucleotides. Proc. Nucleic Acid Res. 2: 751-795.
2. Beemon,K., and T.Hunter. 1977.In vitrotranslation yieldsa possible RSV-src geneproduct. Proc. Natl. Acad. Sci.U.S.A. 74:3302-3306.
3. Bister, K., and P. H.Duesberg. 1979. Structureand specific sequences of avianerythroblastosisvirusRNA: evidence for multiple classes of transforming genes among avian tumor viruses. Proc. Natl. Acad. Sci. U.S.A. 76:5023-5027.
4. Brugge, J.S., and R.L.Erikson.1977.Identification of
atransformation-specific antigeninducedbyanavian sarcoma virus.Nature(London)269:346-348. 5. Collet, M.S.,and R.L.Erikson. 1978. Protein kinase
activity associated with the aviansarcomavirus src
geneproduct. Proc. Natl. Acad. Sci. U.S.A. 75:2021-2024.
6. Duesberg,P.H.,K.Bister,and P. K.Vogt.1977.The RNA of avianacuteleukemia virus MC29. Proc. Natl. Acad.Sci. U.S.A.74:4320-4324.
7. Fujinami,A.,and K. Inamoto.1914.UeberGeschwulste
bei japanischenHaushuhnerninsbesondere uber einen transplantablenTumor.Z.Krebsforsch.14:94-119.
8. Hanafusa, T., L.-H. Wang, S. M. Anderson, R. E. Karess, W. S. Hayward, and H. Hanafusa. 1980. Characterization of thetransforminggeneofFujinami sarcoma virus. Proc.Natl. Acad.Sci. U.S.A.
77:3009-3013.
9. Hayman,M.J.,B.Royer-Pokora,and T. Graf. 1979. Defectiveness of avianerythroblastosisvirus:synthesis of a 75Kgag-related protein.Virology92:31-45.
10. Hunter,T.,and B. M. Sefton.1980.Thetransforming gene product of Rous sarcoma virus phosphorylates tyrosine.Proc.Natl.Acad.Sci. U.S.A.77:1311-1315.
11. Itohara, S.,K.Hirata,M.Inoue, M.Hatsuoka,and K.Sato.1978.Isolation ofsarcomavirusfrom sponta-neous chickentumor.Gann 69:825-830.
12.Kamahora,T.,H.Sugiyama,A.Nomoto,M.Yoshida, and K.Toyoshima.1979.RNAspecificfor the trans-forming component of avian erythroblastosis virus
strain R.Virology 96:291-294.
13. Kawai,S.,M.Yoshida,K.Segawa,H.Sugiyama,R.
Ishizaki,and K.Toyoshima.1980.Characterization
ofY73,anewly isolatedaviansarcomavirus:aunique transforminggeneand itsproduct,a phosphopolypro-tein withproteinkinaseactivity.Proc.Natl.Acad.Sci. U.S.A. 77:6199-6203.
14.Kessler, S. W.1975.Rapidisolation ofantigensfromcells with aStaphylococcus proteinA-antibody adsorption: parameters of the interactionofantibody-antigen
com-plexeswithproteinA. J.Immunol. 115:1617-1624.
15.Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly oftheheadofbacteriophageT4.
Nature(London) 227:680-685.
16.Lee, W.-H.,K.Bister, A.Pawson,T.Robins, C. Mos-covici,andP. H.Duesberg. 1980.Fujinamisarcoma
virus: an avian RNA tumorviruswith aunique trans-forming gene. Proc. Natl. Acad. Sci. U.S.A. 77:2018-2022.
17.Lee,Y.F., andE.Wimmer.1976.Fingerprinting high
molecularweightRNAbytwodimensionalgel electro-phoresis: applicationtopolio virus RNA. Nucleic Acids
Res. 3:1647-1658.
18. Mellon,P., A. Pawson, K. Bister, G. S. Martin, and P.
H.Duesberg.1978.Specific RNA sequences and gene
products ofMC29avian acute leukemia virus. Proc. Natl. Acad. Sci.U.S.A.75:5874-5878.
19.Pawson,T., R.Harvey, and A. E. Smith. 1977.The J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
GENOME STRUCTURE OF Y73 437
sizes of RSV mRNA active in cell free translation. Nature (London) 268:416-420.
20. Pelham,H.R.B., and R. J. Jackson.1976.Anefficient mRNA-dependenttranslation system from reticulocyte lysates. Eur. J. Biochem. 67:247-256.
21. Purchio, A.F., E.Erikson, and R. L Erikson. 1977. Translation of 35S and subgenomic regions of avian sarcomavirusRNA. Proc.Natl. Acad. Sci. U.S.A. 74:
4661-4665.
22. Rettenmier, C. W., S. M. Anderson, M. W. Riemen, and H. Hanafusa. 1979. gag-relatedpolypeptides
en-codedbyreplication-defectiveavianoncoviruses. J.
Vi-rol. 32:749-761.
23. Reynolds, F. H., Jr., T. L. Sacks, D. N. Deobagkar, and J. R. Stephenson. 1978. Cells nonproductively transformed byAbelson murine leukemia virus express
ahighmolecular weight polyprotein containing struc-tural and nonstrucstruc-tural components. Proc. Natl. Acad. Sci. U.S.A. 75:3974-3978.
24. Shields, A., S. Goff, M. Paskind, G. Otto, and D. Baltimore. 1979.Structure of Abelson murine leuke-mia virus genome.Cell18:955-962.
25. VonderHelm, K., and P. H. Duesberg. 1975. Trans-lation ofRSV RNA in a cell free system from ascites Kregs II cells. Proc. Natl. Acad. Sci. U.S.A. 72:614-618. 26. Wang, L-H., P. Duesberg, K. Beemon, and P. K.
Vogt. 1975.Mapping RNase Tl-resistant oligonucleo-tides of avian tumor virus RNA: sarcoma-specific oli-gonucleotides are near the poly(A) end and oligonucle-otides commontosarcomaand transformation-defec-tive viruses are atthe poly(A) end. J. Virol. 16:1051-1070.
27. Yoshida, M., S. Kawai, and K. Toyoshima. 1980. Un-infected aviancellscontainstructurally unrelated pro-genitors of aviansarcomagenes. Nature(London) 287: 653-654.
28. Yoshida, M., M. Owada, and K. Toyoshima. 1975. Strainspecificity of changesinadenylate cyclase activ-ity incellstransformed by avian sarcoma viruses. Vi-rology 63:68-76.
29. Yoshida, M., and K. Toyoshima.1980.In vitro
trans-lation of avianerythroblastosis virus RNA: identifica-tion of twomajorpolypeptides. Virology 100:484-487. 30. Yoshida, M., M.Yamashita, and A. Nomoto. 1979.
Transformation-defective mutants of Rous sarcoma vi-ruswithlonger sizesof genome RNA and their highly frequentoccurrences. J. Virol.30:453-461.
31. Yoshida, M., and H. Yoshikura.1980.Analysis of spleen focus-formingvirus-specific sequencescoding for spleen focus-formingvirus-specificglycoprotein witha molec-ularweightof 55,000 daltons(gp55). J. Virol.
33:587-596. VOL. 38, 1981
![FIG. 1.Dottedwereinsucrosedodecyland(d, 40 e). Analysis of Y73 RNA by agarose gel electrophoresis (a, b, c) and sucrose gradient centrifugation [32P]RNA of Y73 virions containing poly(A) before (a) and after (b) heat treatment at 1X0°C for 1 min [32PJRNA o](https://thumb-us.123doks.com/thumbv2/123dok_us/1479390.100577/2.500.137.371.411.582/dottedwereinsucrosedodecyland-analysis-agarose-electrophoresis-gradient-centrifugation-containing-treatment.webp)