JOURNALOFVIROLOGY, Jan.1978,p.215-223 Copyright ©1978 AmericanSociety for Microbiology
Vol.25, No. 1 Printedin U.S.A.
Replication Process of the Parvovirus H-1
X.
Isolation
of a Mutant
Defective
in
Replicative-Form
DNA
Replication
SOLON L. RHODE III
PutnamMemorialHospital Institute for Medical Research, Bennington, Vermont 05201
Received forpublication26 May 1977
A
temperature-sensitive
mutant of H-1, ts14, that ispartially
defective inreplicative-forn
(RF)
DNAsynthesis
hasbeen isolated. ts14 H-1 ischaracterized
byadecrease in
plaque-forming ability
andproduction
ofinfectious virusattherestrictive
temperature of39.50C.
RF DNAsynthesis
ofts14isreduced to 3 to7% of that of
wild-type
H-1ateither the restrictive or thepermissive temperature.A complementation
analysis
ofRF synthesis of ts14 and a viable defective H-1virus,
DI-1,
orwild-type
H-3indicates that the defective RF DNA synthesis ofts14 is
cis-acting.
ts14, unlikewild-type H-1,
causes amultiplicity-dependent
inhibition
ofDI-1orH-3,
butnotLuII,
RF DNAsynthesis.
Mixedinfections ofcells withtwo
parvoviruses
also exhibited a cross-interference for viralprotein
synthesis
thatwasmultiplicity dependent.
ts14inhibitedinfectious virusproduc-tion of H-1 or
H-3,
but notLuIlI.
LuIII- orH-3-pseudotype particles
wereproduced by coinfection with H-1. H-3 and H-1 showed similar interactions with
ts14, and H-3 DNAwas more
homologous
toH-1 thanwasLuIIIby
comparativephysical
mapping
studies. Theresults suggest that ts14 is a mutant with a defectina
regulatory
sequence ofits DNA thatinfluences
RF DNAreplication.
The
parvovirus H-1,
which hasagenome oflinear
single-stranded
DNA, produces
adouble-stranded
replicative-form
(RF)
DNAduring
in-fection (5,
6).
This RF DNAreplicates
andpro-vides the
template
for thesynthesis
ofprogenysingle-stranded
DNAforincorporation
intovir-ions
(6-8). Progeny
DNAsynthesis
requires
oneorboth of the virion
capsid proteins
(71.
It hasbeen shown that RF DNA
replication requires
aviral gene
product
(S.
L.Rhode,
in D. Wardand
P.Tattersall,
ed., Replicationn
of
Mam-malian
Parvoviruses,
inpress).
Toanalyze
thisrequirement,
I have searched fortemperature-sensitive mutants defective in RF DNA
repli-cation. In this
study,
thetemperature-sensitive
mutant ts14 will be shown to be defective in
RF DNA
replication (RF
rep-),
but notcondi-tionally
with the temperature.Plaque-forming
ability
and virusproduction
aretheonly
param-eters ofts14
replication
that have been found tobe heatlabile.Thenature of theRFrep-defectofts14was
explored by
complementation
studies betweents14 and RF
rep'
viruses ofH-1, H-3,
andLuIII. The RF rep-phenotype
was found to becis-acting.
ts14alsoproduced
amultiplicity-depend-ent inhibition of RF DNA syntheses of the
helper
virusesH-1, H-3,andH-T, butnotLuIII.A
multiplicity-dependent
cross-interference forviral
protein (hemagglutinin
[HA])
synthesiswas also defined for H-1, H-3, or LuII. H-3
DNA was
found
to be more homologous thanLuIII DNA to
H-1
by physical
mappingtech-niques.
These studies suggest that ts14 has amutation ina
regulatory region
of itsDNA thatinfluences RF DNA
replication
and that ts14proteins
are notdefective for RF DNAreplica-tion.
MATERIALS AND METHODS
CeUs andvirus strains. The cells used in this study are human newborn kidney cells transformed bysimianvirus 40 (NBcells).The H-1 mutants ts14 through-18 wereisolatedaspreviously described(3, 7) from a virus stock mutagenized with N-methyl- N'-nitro-N-nitrosoguanidine, obtained from Aldrich ChemicalCo., Inc., Metuchen, N.J. (8). Mutageniza-tionwas carriedout by treatinga parasynchronous culture of hamsterembryocellsinfected with H-1at amultiplicityof infection of5with mediumcontaining N-methyl-N'-nitro-N-nitrosoguanidine,25
iLg/ml,
from 12 to 14 hpostinfection.Themutagenwasremoved, and the cultureswerewashed twice withHanks bal-ancedsalts and refed withregularmedium. The cul-tures wereharvestedat56hpostinfection.Theyield ofplaque-formingviruswasreduced 90%bytreatment with thisdosage ofmutagen. The H-1 mutant DI-1 wasisolatedfroimapreparation of defective interfering (DI) viruses obtainedby serial passage at high multi-215on November 10, 2019 by guest
http://jvi.asm.org/
216
plicity with wild-type (wt) H-1 helper virus (manu-script inpreparation).
Virusand HA neutralization. Virus preparations in tissueculture mediumwereadjustedtopH8.5with 0.5NNaOH and frozen and thawed three times. Equal volumes of virusand antiserumweremixed and incu-bated for 1 h at 37°C. After neutralization, twofold dilutions of the sample weremade with phosphate-buffered saline for titration of HA with guinea pig erythrocytes. Other samples were used directly to inoculate cover glasscultures of NB cells for deter-mination ofproductionof viralantigenby immunoflu-orescent (FA) staining. Cells with nucleipositivefor parvovirusantigenswerecounted withanocular grid at X400.
Preparation of radionuclide-labeled viral DNA. Parasynchronous cultures of NB cells were infected with H-1 and incubated inalow-phosphate medium containing [32P]orthophosphate at 20 to 50
,uCi/ml and/or
[3H]thymidine
in the presence of5-fluorodeoxyuridine,0.5,ug/ml, from12 to 16h postin-fection. Viral DNAwasextracted bythe method of Hirt andpurifiedby sedimentation in neutralsucrose
gradientsaspreviouslydescribed(7). Hirtextracts to beanalyzeddirectly bygel electrophoresiswere con-centratedbyprecipitationwith ethanol. The
precipi-tates were collected by centrifugation and dried in vacuo. The DNA was redissolved in 50 mM Tris-hydrochloride (pH7.5)-imMEDTA anddigested for 30min at roomtemperature withpancreatic RNase (treatedfor20minat80°C)at50ug/ml.
Gelelectrophoresis.Slabgel electrophoresiswas conducted in an EC470 apparatus (E-C Apparatus Corp.,Philadelphia, Pa.), using gels (0.3 by 12by 16
cm)of1% agarose.Afterelectrophoresis,thegelswere dried invacuoandexposedtoKodakRoyalX-Omat RP/R2for upto2weeksatroomtemperature. Acryl-amide gels with acrylamide concentrations greater than 7.5%werepartially dehydrated byimmersion in 70% ethanol plus30% 0.15 MNaCl-50 mM Tris-hy-drochloride (pH 7.5)for1to2hat4°Cbeforedrying. Thisprocedure resulted insomeshrinkageofthe gel butprevented cracking duringthedryingprocess.
Quantitationof 32Por3H indried agarosegels was done byexcising the DNA-containing region of the gelandincubatingthe slice in 0.15 ml of50mM Tris-hydrochloride (pH 7.5)-0.5 mMMgCl2-2 ,ug of pan-creatic DNase for 12 to 16 h at 37°C. The sample
was then treated with 0.6 ml of NCS (Amer-sham/Searle,Chicago, Ill.)and counted with 10 ml of toluene-basedscintillation fluid inaliquidscintillation spectrometer. Acrylamide gelslices weresolubilized with perchloric acid and hydrogen peroxide (4) and counted withAquasol (NewEngland NuclearCorp., Boston,Mass.).
Restriction endonucleasedigestionof H-1 RF DNA. The bacterial restriction endonucleases used here were purchased from Miles Laboratories, Inc., Elkhart, Ind.Digestionswereperformedaspreviously described (9).
RESULTS
Isolation andcharacterization ofts14. All
H-1isolates
showing
temperaturesensitivity
forplaque formationwere
propagated
totiters of5x 107or higher. Theisolate ts14 had a titer of
less than 5 x
103
PFU/mnl
at the restrictivetemperature of 39.5°C and a titer of 3 x
108
PFU/mlatthepermissivetemperatureof33°C.
Theplaquesatthepermissive temperature
ap-peared
smaller than those ofwtH-1.All ofthemutantsisolatedwerescreened for reduced RF
DNA
replication
atthe restrictive temperatureby
infecting
with amutant one 100-mmdish ofNB
cells
synchronized with methotrexate andlabeling the viral DNA with
[32P]orthophos-phate from 10 to 14 h postinfection at39.5°C.
The viral DNAs were extracted and analyzed
by
electrophoresis
in aslab
gel of 1% agarose,as described in Materials and Methods. One
isolate, ts14, was selected for furtherstudy
be-causeit
synthesized considerably
less32P-labeledRF DNA monomer or dimer than didwtH-1.
ts14isthe
only
mutantisolatedtodate that hasbeen defective in RF DNA
synthesis
at therestrictivetemperatureof39.5°C.
ts14 and wt H-1 DNA syntheses were
com-pared undera
variety
ofcondtionstodetermine
if the
synthesis
ofts14 RF DNA wastempera-ture
labile
and if itwas rapidly inhibited by ashift from the
permissive
tothe restrictivetem-perature. In this
experiment,
viral DNA waslabeled for the indicated times with
[3H]-thy-midine and fractionated in a slab
gel
of 1%agarosein thepresenceofmarkers of
32P-labeled
H-1 DNAs. Themonomerand dimer RF DNA
and viral DNA bands of each
sample
wereex-cised from the dried
gel,
and the3H
wasmea-sured
by
liquid
scintillation spectrometry. Theresults
(Table
1) indicate that ts14 monomerand dimer RF DNA
synthesis
are defectiveatboth39.5and33°C. The
synthesis
of viral DNAwas also
decreased
at both temperatures, butnot as
profoundly
as that of RF DNA(72
to87%
compared
with92to96%).
Since the diminished
synthesis
ofts14 viralDNAwas nottemperature
conditional,
thepro-duction of infectious viruswasexaminedatthe
restrictive andpermissivetemperatures.The
re-sults indicate thatts14 infectious virus
produc-tion at
39.50C
is defective, with yields about10-1
to10-2
of those of ts14 at 33°C or wt at either temperature (Table 2). The production ofts14 at33°Cissurprisingly high, consideringthedefectivenessofts14RF DNAsynthesis at
330C.
The parameters of ts14replication
thatare
thermolabile
areplaque-forming
ability
andinfectiousvirusproduction.I was notsuccessful
indemonstratingcomplementation betweentsl,
ts2,
orts18andts14forinfectiousvirusproduc-tion at
39.50C.
tsl,ts2,
and ts18 are RF DNArep' (7),butnoneoffiveH-1mutantstested to
date have been complemented by each other
even though
they
demonstrated differences inJ.
on November 10, 2019 by guest
http://jvi.asm.org/
phenotypeatthe restrictive temperature. There-fore, negative complementation has not been
proventobesignificant for conditionalmutants
of H-1. Theonlyviruses for which complemen-tation has been demonstratedare DI particles of H-1 (Rhode,inpress).
Complementation
oftsl4and RFrep'
H-1.Aputative missensemutation in a viral
pro-teinmightbe difficult to identify directly, so a
genetic approachwas usedtodetermine ifts14 has amutation in astructural gene foraviral proteinrequired for RF DNA replication. If ts14 produces atrans-acting defective protein, then
coinfectionwithanRF
rep'
helper virusshouldrescue the ts14 RF DNA and greatly increase itsreplicationrate.Alternatively,ats14protein
may beinhibitory to RF DNAreplication and willactin atrans-dominant fashion, inhibiting
the replication of an RF
rep'
DNA. Also, the mutationmaybe inaregulatory region of ts14DNA and, consequently, behaveas a
cis-domi-nant mutation. Complementation experiments with H-1weremadepossible by the isolation of
DI-1, aviable defectiveH-1 virus with normal
levels of RF DNA replication whose RF DNA
can be identified by virtue of an addition of about65 base pairs in the HindII-D region of the H-1 physical map (unpublished results).
Theseexperimentsweredoneby comparing the
yields of 32P-labeled RF DNA of ts14 at 370C in thepresenceand absence ofcoinfection with
TABLE 1. Synthesis ofwtandtsl4RFDNAand viral DNAat39.5or330Ca
[3H]thymidineincorporationb Incuba- ts14(%)
teionp
Di- Mono-tsl4/wt
tep mer mer Viral (total
RF RF DNA cpm)
DNA DNA
33°C 4 4 13 2,623/70,275
39.50C 7 6 28 2,970/43,432
33-39.50Cc 5 4 16 5,391/122,207
aReplicate cultures of parasynchronous NB cellswere in-fectedwithts14orwtH-1atamultiplicityofinfection of 25
to50. Viral DNAwaslabeledby incubating the cultures in prewarmed medium containing 5-fluorodeoxyuridine, 0.5
ag/ml,and [3H]thymidine, 5 puCi/ml, 10' M. Thelabeling periodswere:33.C,18 to 20 hpostinfection;39.50C,12 to 14 hpostinfection;and33 to 39.50C, 19 to 20 hpostinfection.
Theincorporationof[3H]thymidineintoH-1 RF DNAand
viral DNAwasquantitatedasdescribed in thetext.
bThevalues shownarethe relativeamountof 3H in each type of DNA for ts14 compared with wt, expressed as a
percentage and the total countsper minute (dimer RF+
monomerRF + viral DNA) for each virus. Monomer RF
DNA represents about 80% of the total, dimer RF DNA
represents about 15 to 18% ofthe total, and viral DNA
represents the remainder.
cIncubatedat330C until18hpostinfection, then shifted
[image:3.505.262.451.62.191.2]to39.50C.
TABLE 2. Production of tsl4 at the restrictive and permissivetemperaturesa
Virus production H-1 Group Incuba- (PFU/ml) type no. tiontemp
1hp.i. 48hp.i.
tsl4 1 330C 4x 106 1 x 10l
39.50C 1x 107 1 X 107 2 39.50C 1 x105 9 x106
wt 1 330C 7 x106 6x10
39.50C
1x107 1X109 2 39.50C 2 x106 3 x105aNB
cell
cultures were inoculated withts14 or wtH-1 at a multiplicity of infection of 5 to 10 and ad-sorbed for 60 min at their respective temperatures. The cultures were washed twice with Hanksbalanced salt solution and incubated at theirrespective temper-atures.Cultures were harvested and virus infectivity was titratedby plaque assay at 33°C at 1 and 48 h postinfection (p.i.).
DI-1. The total incorporation of 32p into RF DNA from each infection was determined by
gel
electrophoresis
or by sucrose gradientcen-trifugation. The yield of 32P-labeled RF DNA
of each virus inamixedinfectionwascalculated,
using the relativeamountof each viral DNA in
the mixture determined by the amounts of its
marker
fragments
(HindII-D or -D'). Arepre-sentative
electropherogram
ofa HindII digestof the RF DNA ofts14,DI-1, andamixture of
thetwois shown in
Fig.
1.Parasynchronous NBcultures were infected with ts14, DI-1, or ts14 and DI-1 at various multiplicities of infection
(Table
3). Itshould be noted that themultiplic-ities of infection ofts14 and DI-1 quoted here
are based on the infectivity titrations of each
virus by plaque assay, and the efficiencies of
thisassayfor thetwovirusesmaydiffer. When
ts14 had a
multiplicity
of infectionadvantage
overDI-1
(experiment
1,Table
3), it inhibitedthe RF DNA
synthesis
of DI-1 andreduced thecombined
yield
of RF DNA for DI-1 andts14by
65%compared
with DI-1 alone. In otherwords,
thets14 RFrep-phenotype
wastrans-acting.
Since amultiplicity-dependent
cross-in-terference for viral
protein synthesis
doesoccurin mixed
infections,
asdiscussed below,experi-ments 2and3 weredone with DI-1atthesame
multiplicity
asts14orwithamultiplicity
advan-tageof3:1overts14.The results showthateven when DI-1 was the majority virus and should
have produced most ofthe viral protein, ts14
RF DNA
synthesis
wasonlymodestly increased. Because ts14 RF DNA incorporated relatively lownumbers of32p
countsper minute and be-cause thisincreasedidnotapproacha wtlevelof
incorporation,
it was notconsideredasasig-nificant complementation of the defective RF
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.53.247.406.514.2]DNAreplication. Thus, the ts14 mutation was
cis-actingatlowmultiplicities of ts14 in comple-mentation with DI-1. DI-1 RF DNAsynthesis
was reduced by the trans-dominant effect of
ts14 ts14
+ +
ts14 DI-I Dl-l
D-i-1. w --D
FIG. 1. Gelelectrophoresis pattem ofHindIIand HindII-plus-HpaII digestion products oftsl4and
DI-1RFDNA. The32P-labeledRFDNAoftsl4-,DI-l-,
and tsl4-plus-DI-1-infected cultures wereprepared
byHirt extraction and sedimentation in a neutral
sucrosegradientaspreviouslydescribed(7). Samples ofeach were digested in 20-IAl volumes for4 h at
37°Caspreviouslydescribed(9).(1)40%ofthetotal
yield ofts14RF DNAdigested byHindII;(2)20%of the totalyield ofts14plusDI-1digested byHindII; (3)asin(2), digested byHindIIandHpaII; (4)10%
oftheyield of DI-1RF DNAdigested by HindII.
ts14inproportiontothe ratio of ts14toDI-1 in the inoculum.
RF DNA synthesis in heterotypic infec-tions oftsl4 H-1 with H-3 or LuLI. In the
complementation tests with ts14 and DI-1, it
was not possible to determine if DI-1 proteins were synthesized, since ts14 and DI-1 proteins
areantigenically similar. Therefore,
complemen-tation experiments were carried out with ts14 and other nondefectiveparvoviruses capable of infecting NB cells. In preliminary experiments,
ts14 H-1 caused a reduction in total RF DNA
synthesis on coinfectionwith wtH-1, H-3, and
H-T,aspreviously described for coinfection with
DI-1. In contrast,LuIII RF DNA synthesiswas
not inhibited. Therefore, H-3 and LuIII were
selected for further study. First, itwas necessary
toobtain markers for H-3andLuHI RF DNAs,
so that synthesis of their respective RF DNAs
could be measured in the presence ofH-1 RF
DNA. Cleavage maps of H-3 and LuIII were
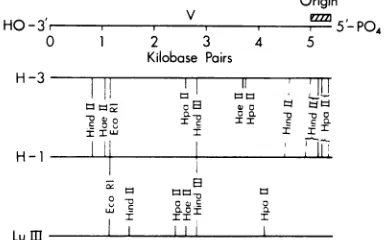
determined for the restriction endonucleases EcoRI,HindIII, HindII,HaeIII, and HpaII(Fig. 2). The assignment ofthe origin of replication and the 5' terminushasbeenmade only for H-1; H-3 and LuIIIare oriented in the figure on
the basis ofhomologous cleavage sites and the variations instructureofthemolecular endsas
describedfor H-1(9).TheRFDNAsofts14 H-1, H-3, or LuIII alone or of H-3 or LuIII in
mixed infection with ts14 were labeled with
[32P]orthophosphate and prepared for digestion with HaeH as in Table 3. Replicate cultures wereharvested for HA neutralization assaysto
measure the effect of mixed infection on viral
protein synthesis. Cover glass cultureswerealso
infected with the virus mixtures in the same
manner and stained for the antigens of both viruses oreach virusseparately by the indirect
FAstaining method. The FA staining in the H-3 experiment showed that 70% of the cells (300
TABLE 3. InhibitionofDI-1RF DNA synthesis bycoinfection withtsl4a
MOjb RF DNAyieldc
Exptno.
DI-1 ts14 ts14 DI-1 tsl4/DI-1 Total(cpm/107cells)
1 10 20 0.9 0.1 1.6 115,540
2 5 5 0.8 0.4 0.4 205,065
3 15 5 1.5 0.8 0.2 144,683
aTheyields ofts14 RFDNA orDI-1 RF DNAafter double infection at 37°C with ts14 and DI-1 were
calculated from thecountsper minuteof3P recovered in theirrespective marker fragments, HinduI Dfor ts14andHindII D' forDI-1,asillustrated inFig.1. The valueswereadjusted for relative fragment sizes and quantities appliedtothegel. The total RF DNAyieldsweremeasured by eithergel electrophoresis in0.8% agarose gels or Cerenkov counting ofthe preparative neutral sucrose gradients. The specific activities of
[32P]orthophosphate
in eachexperimentarenot thesame.b
MOI,
Multiplicity of infection(PFU/cell).
cValues shownare:theyield of each virus RFDNAinthe
mixed
infectionnormalizedto theyield of thatvirus alone(ts14, DI-1),the ratioof ts14 RFDNA tothat of DI-1 in the mixture (ts14/DI-1), andthetotal countsperminute inmonomerRF DNAper107cells.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.66.259.123.374.2]counted) were
positive
foreither H-1 orH-3or both (incubated with anti-H-1 and anti-H-3), with 78% positive for H-1 only and 62% positivefor H-3
alone.
Theresult
forthe
LuIIIexperi-ment was 75% positive for H-1 or LuIII, 71%
for H-1, and 51%positive for LuIII. Therefore,
the majority of the infected cells produced
an-tigens of both viruses in the mixed infections.
HO- 3'r-0
V
1 2 3
Kilobase Pairs
c 0
I i
zo II
Origin
4 5
11
t
0 aII
ILI
0 10a
O 00.
Q- B
l, m0 c().cS
Ix
I II IFIG. 2. Comparison ofthephysical mapsof H-1,
H-3,andLuIII.ThecleavagemapsofH-3 andLuIII
to the bacterial restriction endonucleases endo R EcoRi, endo R HaeII, endo R HindII, endo R HindIII, and endo R HpaII were determined with native RFDNA asforH-1 (9). Cleavagesites thatarehomologousbetweenH-1 and H-3orLuIII
are indicated by the long solid lines, and nearly homologous cleavagesitesaremarkedbythe dashed lines.CleavagesitesonH-3orLuIIIwithout similar sites on H-1 aremarked by short solid lines. The clusterof HpaIIcleavagesites atthe rightendof H-3,indicatedbythe braces,isverysimilar, butnot identical, tothatattherightendofH-1. Thereare no additional cleavagesitesfor HindIIwithin the brackets markedHindII. TheorientationoftheH-3 andLuIII cleavagemapswithrespect tothe5'-PO4 terminusofthe viral strand andorigins ofreplication
werenotdeterminedandare assignedhere strictly
on the basis oftheirhomology toH-1 (13, 14). The totallength ofH-I has beenadjusted upwardto5,400 basepairs, based on more recent estimates ofthe
sizeof
OMK74
DNA(11).The yields of RF DNA for each virus were
determined, using
their distinct HaeII-Afrag-ments
(Table
4). Coinfection
with ts14H-1
re-duced
H-3 RF DNAsynthesis
about 50%
anddid notinhibit LuIII RF DNA
synthesis.
Thelevel ofts14RFDNA
synthesis
was notchanged
by H-3.On the other
hand,
inthe culturecoin-fected with
LuIII,
ts14 RFDNAsynthesis
wasmarkedly reduced,
whereasLuIII RFDNAsyn-thesis was
unchanged.
It will be shown in thenext section that in the mixed infections with
ts14 at alower
multiplicity
than thehelper virus,
as used
here,
themajority
ofthe HAssynthe-sized were H-3 or
LuIII,
not H-1. When ts14and
H-3wereused inamixed infection with amultiplicity
of 3:1 in favor of ts14, themajor
portion of the HAwasH-1 and the
yield
of"P-labeled RF DNA of H-3 was reduced at least
70%(datanot
shown).
Thus,
ts14inhibited H-3RF DNA
synthesis
inamultiplicity-dependent
manner, as found
previously
with DI-1 H-1.With wt H-1 and H-3 at
multiplicities
of 10PFU/cell, the
bulk
of theHAwasH-1(c.f.
Table5, experiment 3) and the RFDNA
synthesis
ofeach viruswas notreduced.
ts14 inhibtion of HIA
synthesis
of otherparvoviruses.
Theparvoviruses H-1, H-3,
andLuIII have similar hostrangesin that
they
allinfect many of the same human and hamster
cell
lines (1, 14).They
also showatleast somesequence
homology
as determinedby physical
mapping
with bacterial restrictionendonucle-ases
(Fig. 2).
Itwill be shown here thatcoinfec-tion ofH-3-orLuIII- infected NB cultures with
ts14 H-1 exhibits a cross-interference for the
synthesis
of their HAs(capsid proteins).
The inhibition of DI-1 H-1 or H-3 RF DNA
synthesis by
ts14 H-1 could result from theinhibition of
synthesis
of aputative
RFrep'
gene
product.
ForLuIII,
whose RF DNAsyn-thesis
was notrepressed by
ts14,
itshould bedetermined
whetherLuIII
inhibitedts14protein
TABLE4. Effect of
tsi4
or wtH-i
coinfection on the RF DNA synthesis of H-3 andLuIIIa
MOIb RF DNAyielda
Exptno. Viruses H-3or
H-1i LIII H-1 H-3orLuIII Total cpm
1 ts14+H-3 6 12 1.07 0.47 384,717
2 ts14+LuIII 6 12 0.04 1.23 143,921
3 wt +H-3 10 10 1.34 1.15 1,585,713
aThe yields of32P-labeledH-1 and H-3 orLuIII RF DNA after single or mixed infection at 37°C were
calculated by determining the relativeamountof eachvirus in themixture and measuring the total yield of RF DNAof each infectionby gel electrophoresis. Intheseexperiments, the HaeIIA fragments of each virus wereusedasmarkerfragmentsand thecountsperminute recoveredwerecorrected for differences in size of therespectiveHaeIIAfragments. Experiments1and2weredoneatthesametime.
bMOI, Multiplicity of infection
(PFU/cell).
cValues shownaretheyield of each virusRF DNAin the mixed infection normalized to the yield of that
virusalone and total counts perminute in monomer RF DNA. H-1
Lu
mE
219
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.54.246.160.280.2] [image:5.505.51.448.514.584.2]220
synthesis. For H-3 and LuIII, whichare
antigen-ically distinct from H-1, these possibilities can
be testedby measuringtheeffect ofts14 on
H-3 or LuIII HA synthesis. This was done by
infecting cultures with ts14 H-1, H-3, or LuIII
alone orincombination (and similarly with wt
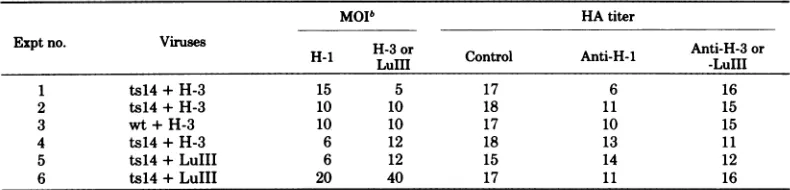
H-1 and H-3) and comparing the yield of HA after neutralization with the various antisera (Table 5). Itwasfound that ts14 inhibited H-3
orLuIIIHAsynthesis and thatwtH-1inhibited
H-3 protein synthesis as well. This inhibition
wasmostmarked whents14wasusedatahigher
multiplicitythan that of H-3. FAstaining of NB
cellsinfectedwith ts14and H-3 (as inTable 5, experiment 1) revealed about 60% of the cells
tobepositive for H-3 intranuclear antigen,
sug-gesting thatmostcellsmayhaveproduced H-3
protein in reduced amounts. By giving H-3 a
multiplicity advantageof2:1 (experiment 4, Ta-ble 5), theproportionofH-3 HAwasincreased
to an amount greater than that of H-1. This
was the same experiment used for analysis of H-3RF DNAsynthesis in thepresence ofts14
in Table 4. Thus, it isunlikely that inhibition of H-3proteinsynthesis accountedforthe inhi-bition of H-3 RF DNA synthesis caused by coinfectionwithts14. The failure ofts14to in-hibitLuIII RF DNAsynthesiswas not due to LuIII inhibition ofts14 protein synthesis since LuIII RF DNAwas not found to be inhibited forexperiment5 (c.f. experiment 2, Table 4) or
forexperiment6 (comparativedata notshown) of Table5,wherets14proteinwaspredominant.
Since the hemagglutinating units after each neutralizationwere not additive tothecontrol,
perhaps dueto phenotypic mixing ofcoat
pro-teins, thesedatacannot beinterpretedinstrictly quantitative terms. They do indicate that H-1,
H-3,andLuIIIcompeteforsomelimitingfactor
concerned with viralprotein synthesisandthat H-1 tends to dominate these mixed infections, especially whenit hasanadvantage in
multiplic-ity.
Effect of ts14 H-1onproduction of
infec-tious virus inmixed infections with other parvoviruses. The effect of coinfection with ts14 H-1 on the production ofinfectious virus
ofwt H-1, H-3,andLuIIIwas examined.Since
ts14producesnoplaquesat39.5°C,theproduct of mixed infectionscanbe plaquedat39.5°Cto
determine theyield ofwtvirus. The results of these experiments are shown in Table 6. ts14 inhibited wt H-1, but not LuIII. The increase in titer for LuIII (as determined by PFU at
39.5°C) wassurprising, since there appearedto
benoLuIII HAproducedinthesecultures
(Ta-ble 7).The possibility of phenotypic mixing in whichLuIIIDNAisencapsidated with H-1
pro-tein (completely or in part) was tested by a
neutralization assay. Particles containing LuIII
DNA were assayed by FA staining for LuIII
antigen after the infectious particleswere
neu-tralizedbytreatment with H-1 antiserum. The LuIII from the mixed infection of ts14 and LuIII produced 7.3 1.2 (mean of 10 microscopic fields at x200 95% confidence limit) LuIlI FApositive cells, and, after neutralization with H-1 antiserum, this dropped to 0.5 ± 0.3. The
control standard LuIII gave 19.3 ± 2.3 LuIII FA-positive cells and 16.3 ± 1.3 after neutrali-zation with anti-H-i. Thus, a large portion of
the virus (from the mixed infection) that
pro-duced LuIII FA antigen could be neutralized with H-1 antiserum. This implies that LuIII DNA was encapsidated wholly or partly with
H-1protein.
In mixed infection with H-3 at 39.5°C, ts14 inhibited H-3production. However, the
replica-TABLE5. Effectofmixedinfectiononthesynthesis ofts14H-i,wtH-i,H-3, and LuIIIHAsa
MOIb HA titer
Exptno. Viruses H-3or Anti-H-3or
H-1i -30r Control Anti-H-I -LuIII
1 ts14+H-3 15 5 17 6 16
2 ts14+H-3 10 10 18 11 15
3 wt+H-3 10 10 17 10 15
4 ts14+H-3 6 12 18 13 11
5 ts14+LuIII 6 12 15 14 12
6 ts14+LuIII 20 40 17 11 16
aNB culturesweresynchronizedwith methotrexateand infected withts14H-1, wtH-1, H-3,and
LuIlI
or combinations of H-1 and H-3orLuIIIat37°C.Thecultureswereharvestedat18hpostinfection andfrozen and thawedthreetimes.Lysates (0.1ml)weretreatedwith0.1mlofspecificantisera for1hat37°Cand then dilutedto0.4ml withbuffer,andtheHAwastitrated withtwofold dilutions. TheH-1, H-3, andLuIIIantisera neutralized allof theHAproduced bytheirrespectiveviruses insingle infectionstotitersofless than4 and showednocross-reactivity.Controlsweretitrateddirectly,withouttreatmentwithserum.Similarresultswere obtainedwhen controlsweretitrated aftertreatmentwith nonimmuneserum.bMOI,Multiplicityofinfection
(PFU/cell).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.65.460.488.583.2]TABLE 6. Effect of coinfection withtsl4 on production of infectious virus by wt H-i-, H-3-, orLulII-infected NB cellsa
Virus yield(PFU/ml)
Expt Virus MOIb 39.50C 330C
lhp.i. 48hp.i. lhp.i. 48hp.i.
1 (asyn) wt 4 2x104 3x107 6x 105 4x 108
ts14 20 0 0 2x106 1X 107
wt+ts14 4+20 2x 104 9X 104 2X106 1X107
2 (syn) LuIII 4 8x 104 9X106 lx 6 6x107
ts14 20 0 0 7x106 4x107
LuIII+ts14 4+20 3x 104 5 x 6 3 x106 5x107
3 (asyn) H-3 4 2 x 104 7 x 105 8 x 105 1X 107
ts14 20 0 0 1x106 3 x106
H-3+ts14 4+20 3 x104 3x 104 3 x106 3x106
4 (syn) H-3 10 4 x104 2 x106 1 x 6 2 x107
ts14 10 0 0 2 x 105 4 x106
H-3+ts14 10+10 6x104 1x 105 9x 105 2x 107
aNB
cells
wereinfected withwtH-1,LuIII,orts14and with each virusplusts14 at39.5°Cforexperiments1through3andat37°Cforexperiment4.Yields of infectious virusweredetermined byplaqueassayat 1 and 48h postinfection (p.i.).Theplaque assays were done atboth 39.5 and 33°C. Since ts14does not produce plaques at39.5°C, wt H-1, LuIIl,and H-3can betitrated in the presence ofts14. Inexperiment 2, the NB cellsweresynchronized(syn) with methotrexate (0.5
jig/ml)
at37°C.Themethotrexate-treated cultureswere infected with LuIIIormockinfectedat37°Cin the presence of thedrug12hafterinceptionof thetreatment. Theywereinfected withts14 ormock infected4hlater and shiftedto39.5°C, with the methotrexate replaced bythymidine(10-' M). For the other viruses (experiments 1, 2, and 4) the infectionsweredone simultaneously withoutmethotrexatesynchronization (asyn)orwith methotrexatesynchronization.bThemultiplicity of infection (MOI, PFU/cell) is based on the quantity of virus in the inoculum, and adsorptionisapproximately50to70%.
cValues shownarePFU permilliliterasassayedat 39.5or33°C.
TABLE 7. Effect of mixedinfectiononthe synthesis of ts14 and LuIII
HAG
HAtiter"Virus MOIb Control
anti-H-i antiLullI
1hp.i. 48 hp.i.
ts14 20 6 17 <4 17
LuIII 4 0 15 16 12
ts14+LuIII 20+4 7 16 <4 16
aNBcultureswereinfected with ts14, LuIII,orts14plusLuIIIasinTable6.Thecultureswereharvested andsubjected tofreezingandthawingthree times. Samples (0.1 ml) at48hpostinfection (p.i.) weretreated with0.1ml ofspecificantisera for1hat37°Cand then dilutedto 0.4ml with buffer, and the HAwastitrated. Controlsweretitrateddirectly,withouttreatmentwithserum.
IMOI,
multiplicity
of infection(PFU/cell).cValues shownarethe
log2
HA titers.tionof H-3 anditsinfectivity titrationby plaque
assay appear to be more sensitive to the high
temperature than do those of H-1. Therefore, thisexperimentwasrepeatedat37°C and with synchronized NBcells.Evenunderthesemore
favorableconditions, theproduction of H-3was
reduced by about 90% by coinfection with ts14 H-1. Since H-3 RF DNA synthesis was also
reduced by ts14 H-1, this resultwas not
unex-pected.
The H-3 virus
produced
in mixed infectionwithts14
H-1
inexperiment
4,
Table6,
orwith wtH-1atthesamemultiplicity
ofinfectionwastested for
phenotypic mixing
of H-3 DNA withH-1
capsid protein
asabove forLuIII.
The H-3 virus(i.e.,
H-3DNA)
of both viruspreparations
was
partially
neutralizedby
anti-H-iantiserum,
andpure H-3wasnot. The number of H-3
FA-positive
nucleipermicroscopic
fieldforts14 H-1plus
H-3 virus was 10.6 + 1.1(mean
of 10221
on November 10, 2019 by guest
http://jvi.asm.org/
222
RHODEareas +95%confidence
limit)
withnonimmune serum and 2.9 ± 1.5 after anti-H-1. The same figuresfor thewtH-1plus H-3were 15.1 ± 2.2for nonimmuneserumand4.2 ± 1.2for
anti-H-1. Therefore, H-3 virus
pseudotypes
with H-1antigen in
their capsids
were formed in mixedinfectionwith eitherwt orts14H-1.
DISCUSSION
Current
knowledge
of the structure andrep-lication of
parvoviruses
suggests thatthey
aremonocistronic
and thatthis cistronproduces
thecoat
proteins
(10). Studies ofcellsinfected withH-1 orminute virus of mice have detected the
synthesis
ofonly
twopolypeptides
induced byviralinfection, the
capsid proteins
VP1and VP2'(termed
A and B in minute virus of mice[2,
15]). Recent studies of the
peptide
maps of theAand B
proteins
of minutevirus of mice indicatethat these
polypeptides
arecoded forbyacom-monregion of the minute virus of micegenome
(16). These findings
imply
that thecomplex
processes of viral DNA synthesis have a high
degree of
dependency
onhostcellproteins.
Thesynthesis
of viral DNAcanbe divided intothreeoperational
stages:(i)
synthesis
of theparental
RF DNA,
(ii) replication
of RF DNA, and(iii)
synthesis
ofsingle-stranded
virionDNA.Anal-ysis
ofinfectionby
atemperature-sensitive
mu-tantof
H-1,
tsl,has shown thatsingle-stranded
virion DNA
synthesis requires
a functioncon-trolled
by
one orbothcapsid proteins (7).
Stud-ies of(DI) viruses of H-1 have established that
aviral
protein
isrequired
for RF DNAreplica-tion
(Rhodes,
inpress),
andtemperature-sensi-tivemutants ofH-1 defective in RF DNA
rep-lication,
i.e.,
RFrep-,arebeing sought.
Inthisreport, Ihave described the isolation and
char-acterization of ts14, the
only
mutantwithdefec-tive RF DNA
replication
of 20 H-1 mutantscharacterized
todate.ts14 was isolated as a
temperature-sensitive
mutant witha thermolabileability
toproduce
plaquesatthe restrictivetemperatureof
39.5°C.
Virus
production
was also diminished whenin-fectionwascarriedout attherestrictive temper-ature. An
analysis
of ts14 RF DNAsynthesis
revealedit tobedefectiveatboth the restrictive
and the permissive temperatures. The
uptake
of
[3H]thymidine
into ts14 RF DNA was 3 to7% ofthat ofwt H-1.
Thus,
the RF rep-phe-notype ofts14 was nottemperature
dependent
undertheseconditions.
Anumberofmechanismscanbe
proposed
for the action of the RF rep- mutation. A choice among themwassought by
conducting
comple-mentation tests for RF DNA
replication
be-tween ts14 and RF
rep'
virusesH-1, H-3,
orLuIII. It has been shown previously that H-1
expresses a
trans-acting
RF rep gene product required for RF DNA replication in NB cells(Rhode, in press). Thiswasestablished by
dem-onstrating that nonviable H-1 DI particles
re-quire coinfection with an RF
rep' helper
virusto replicate their RF DNA. H-3
complemented
the H-1 DI
particles
for thisfunction, but LuIIIdidnot.
Theresults obtained inthecomplementation
tests with ts14and H-1 DI-1 or H-3 indicated
that the RF rep- phenotype of ts14 was
cis-acting andwasnotrescuedby coinfection with
these RF
rep'
viruses. This result for thecom-plementation
tests with H-3 was validated byconfirming that H-3 proteins weresynthesized
in the double-infected
cells
by FAstaining andbyanalysis of H-3 HA synthesis. Thiswas not
possible
for the mixed infections with H-1 DI-1,sincets14andDI-1
antigens
areindistinguisha-ble.Since the H-1 RFrep geneproduct is
trans-acting (Rhode, in press), these findings
imply
that the RFrep-phenotype ofts14is not caused
byamutation in the viral protein required for
RF DNAreplication. Insupportofthis
interpre-tation,ts14 proteinswerefoundtocomplement
the RF rep- DI
particles
described above forRF DNA synthesis as well as wt H-1 (Rhode,
in press). The most likely explanation for the
ts14 mutation
affecting
RF DNAreplication
isthat it is a change ina regulatory sequence of
ts14DNA.
Itisnotcertainwhether the RFrep-mutation
isthesamemutation thatprevents ts14plaque
formation and reduces infectious virus
produc-tion at 39.5°C. Three revertants of ts14 were
isolated
through
theirability
toformplaquesat39.5°C, and all three had lost theRF rep-
phe-notype. Since H-1 has high particle/infectivity
ratios
(unpublished
results), it is possible thatthemethod of selection forrevertants salvaged
some
"subliminal"
contaminants intheoriginalts14 preparation. This seems unlikely, because
each of the three revertants differed from wt
H-1 in either
plaque
morphology or plaquingefficiency
at the restrictive temperature.Be-causethe revertants werenot wt virusand be-cause noneof themwereRFrep-butnot tem-perature sensitive,thequestionofwhetherts14 has one or more than one mutation has not
beenanswered.
It was observed in mixed infections of ts14
with DI-1, H-3, or LuIII that the RF DNA
synthesis of H-1 or H-3, but not LuIII, was
inhibited
by
ts14 in amultiplicity-dependent
manner. wt H-1 did not inhibit H-3 RF DNAsynthesisunder thesameconditions.Since both
H-3 and ts14
produce
RFrep'
proteins (Rhode, in press;unpublished results),
this inhibitionon November 10, 2019 by guest
http://jvi.asm.org/
maybedirectlymediatedbyts14 DNA andnot
by theproteinsitcodes for.Themechanism by
which thisoccursisunknown.
Another feature of heterotypic infections be-tween wt H-1 or ts14 and H-3 or LuIII was a
multiplicity-dependent cross-interference for
HA synthesis. It is likely that LuIII inhibited
ts14RF DNAsynthesis (Table 4) by inhibiting
ts14 RF rep protein synthesis, since its own
proteins do not support H-1 (ts14) RF DNA
synthesis (Rhode, in press). Asimilar
cross-in-terference for the production of infectious
vi-rus wasobtained inheterotypicinfectionsexcept
that the magnitude of the inhibition was less
severethanthatfor HAsynthesis. Thisgreater
survivalof H-3orLuIIIinfectiousvirus
produc-tion during coinfection with H-1 was found to
be due, in part, to the production of H-3 or
LuIIIpseudotype particles. These virions have
H-3orLuIII DNAina capsid that contains
H-1protein.
The complex interrelationships amongthese
parvovirusesshouldbereflectedin the
similari-ties and differences in the base sequences of
theirrespectivegenomes.Thephysicalmapsof
H-1,H-3,andLulIRFDNAsfor the restriction
endonucleases Hindll, Hpall,
HindIII,
HaeII,andEcoRI were compared. Using the method
described by Upholt (17), it can be estimated
from these data that H-1 and H-3 have about
95% homologyand thatH-1 andLuIIIare only
about80%homologous.These estimatesdo not
consideradditionsordeletions,and the
65-base-pair differenceinsize of theH-1 and H-3HindII
fragments at the 95% position (0 is at the left
endinFig. 2) maybeanaddition similarto the
oneinH-1 DI-1.The degreeofDNAsequence
homology between H-1 and H-3 or LuIII as
determinedbythesemappingstudiescorrelates
with the interaction of theseviruses at thelevel of RF DNA replication. But a more detailed
knowledge of the nucleotide sequences in the
regionsencompassingthe replication originsof
these viruses and at their promoters for
tran-,scription
is needed to better understand theinteractionsexhibitedbythem.
ACKNOWLEDGMENTS
This researchwassupported by theU.S. Public Health
ServicegrantCA07826-12from theNational Cancer Institute.
The parvovirus antisera used in this studywerekindly provided byHeleneToolan. Igratefully acknowledgeJessica
Bratton for expert technical assistanceandVirginia Haas and JaneenPrattforsecretarial duties.
LITERATURE CITED
1. Hallauer, C., G.Siegl, and G. Kronauer. 1972. Parvo-viruses as contaminants ofpermanent humancelllines. Arch.Gesamte Virusforsch.38:366-382.
2. Kongsvik,J.R., J. F. Gierthy, and S. L. Rhode mH. 1974.Replicationprocess of the parvovirus1. IV. H-1-specificproteins synthesized insynchronized human NBkidneycells.J.Virol.14:1600-1603.
3. Ledinko,N. 1967.Plaque assay of the effects of cytosine arabinosideand5-iodo-2'-deoxyuridine on the synthesis of H-1 virusparticles. Nature (London) 214:1346-1347. 4. Mahin, D. T., and R. T. Lofberg. 1966. A simplified method of sample preparation for determination of tritium"Cor3Sinblood or tissue by liquidscintillation counting. Anal. Biochem. 16:500-509.
5. Rhode,S.L.,EI. 1974.Replication process of the par-vovirus H-1. II. Isolation and characterization ofH-1 replicativeformDNA. J.Virol. 13:400-410.
6. Rhode.S.L., Im. 1974.Replication process of the par-vovirus H-1. III. Factors affectingH-1 RF DNA syn-thesis. J.Virol. 14:791-801.
7. Rhode, S.L.,m. 1976.Replicationprocess of the par-vovirusH-1. V. Isolation and characterization of
tem-perature-sensitive H-1 mutants defective in progeny DNAsynthesis.J. Virol. 17:659-667.
8. Rhode, S. L.,IH. 1977.Replication process of the par-vovirus H-1. VI. Characterization of a replication ter-minusof H-1replicative-form DNA. J. Virol. 21:694-712. 9. Rhode,S. L.,m. 1977.Replication process of the par-vovirus H-1. IX.Physical mapping studies of the H-1 genome. J. Virol. 22:446-458.
10. Rose, J. 1974. Parvovirus reproduction, p. 1-61. In H. Fraenkel-Conratand R. R.Wagner(ed.), Comprehen-sive virology, vol. 3. Plenum PublishingCorp., New York.
11. Sanger,F.,G. M.Air, B. G. Barrell, N. L. Brown, A. R. Coulson,J.C. Fiddes, C.A.HutchisonIH, P. M. Slocombe, and M. Smith. 1977. Nucleotide se-quence ofbacteriophage4X174DNA.Nature (London) 265:687-695.
12. Siegl, G. 1976. The parvoviruses. In S. Gard and C. Hallauer(ed.),Virology monographs.Springer-Verlag NewYork, Inc.,NewYork.
13. Singer, L. I., and S. L. Rhode MI. 1977. Replication process of theparvovirus H-1. VII. Electron microscopy ofreplicative-formDNAsynthesis. J. Virol. 21:713-723.
14.Singer, L. I., and S. L. Rhode HI. 1977. Replication process of theparvovirusH-1. VIII. Partial denatura-tionmappingandlocalization of the replication origin ofH-1replicative-formDNAwith electron microscopy. J. Virol. 21:724-731.
13.Tattersall, P.,P. J.Cawte,A. J. Shatkin, and D. C. Ward. 1976.Three structuralpolypeptides coded for byminutevirusofmice,aparvovirus.J.Virol.20:273-289.
16. Tattersall, P.,A. J.Shatkin, and D.C.Ward. 1977. Sequence homology between the structural polypep-tides of minute virus of mice. J. Mol. Biol.111:375-394.
17.Upholt,W. B.1977.Estimation of DNAsequence diver-gence fromcomparisonof restrictionendonuclease di-gests.NucleicAcidsRes.4:1257-1265.
VOL. 25, 1978