JOURNAL OFVIROLOGY, Nov. 1986, p. 564-568 Vol. 60, No.2 0022-538X/86/110564-05$02.00/0
Copyright© 1986,AmericanSocietyforMicrobiology
Ion
Etching of Bacteriophage X: Evidence That the Right End of the
DNA Is
Located
at
the Outside of the
Phage
DNA Mass
JAYC. BROWN* ANDWILLIAM W. NEWCOMB
Departmentof
Microbiology,
Universityof Virginia School of Medicine, Charlottesville, Virginia 22908 Received 2 June1986/Accepted5August1986Bacteriophage A was etched in anAr+ plasmaunder conditions in which thecapsid and some of the DNA were eroded (by sputtering) from the particle surface. Analysis of the DNA remaining in etched phage demonstrated an enrichment insequences derived from the left end and middle of the genome; sequences from theright end wereselectively lost. The results suggest that the DNA in the maturephageisarranged with its leftend toward the center and itsrightend toward the exterior of the overall DNA mass. Since the leftend is the first to enter thephage prohead, theresults are mostcompatible with the view that proheadfilling also proceeds from the center to the exteriorofthecavity.Thesuggestedarrangement of A DNA iscomparable to thatobserved inphageT4andis consistent with thespiral-foldmodel ofpackaged DNA.
Recentplasma etchingstudies with bacteriophageT4have
providednewinformation about thearrangementofDNAin thematurephage head (2). ThelastDNApackagedinto the
proheadwasfoundtobeatthe
periphery
oftheoverallDNA mass,while the first DNApackaged
wasinferredtobeatthe center. Such an arrangement would be expected if DNA were condensed beginning at the center of theprohead
cavity andprogressing towardthe outside.
It is attractive to suppose that DNA will prove to be
packaged in other double-stranded DNAphages (suchasX,
P22, T7,and
4)29)
asit is inT4(4, 14), butthis neednotbe the case. There exist significant differences ingenomic
organization and virusstructureamong thedouble-stranded DNAphages, and these mayunderliecorresponding
differ-encesin theway DNAispackaged into thematurehead. For
instance, whereas T4 and P22 DNAs are packaged by a
"headfull" mechanism,packaging in X, P2, and other phages begins andends atfixedpoints onthegeneticmap(14, 18). Permuted andterminally redundantgenomes are character-istic of some double-stranded DNA phages (e.g., T4 and P22), while in others the DNAis identical in all individual particles(e.g.,X,P2,+29, and others). T4differs fromother
phages in that the firstDNA to enter theproheadappears to
be the firstejected during infection ofsensitive cells (3). It
alsodiffers from A inthat itcontains DNA-binding internal
proteins andpeptides (4).
The experiments described below were undertaken to
study the arrangement ofDNA in bacteriophage X. They were designed to determine whether DNA is arranged at
random in the phage head or whether particular regions of the DNA occur systematically in particular regions of the overall DNAmass. Our experimental strategy involved the use ofAr+ etching to distinguish externally disposed from
internalDNAsequences. Intactphage were etched in an Ar+
plasmaunderconditions designed to produce a more or less
geometrically
uniformerosion of the capsid and some, but not all, of the DNA. The DNA remaining after etching (which was expected to be derived from the center of the overall DNA condensate) was then analyzed to determine whether it corresponded to particular mapped restrictionfragments ofmature XDNA.
*
Corresponding
author.MATERIALSANDMETHODS
Phage growth. AllexperimentswerecarriedoutwithAvir (the gift ofA. Blasband) grown on plate cultures of Esche-richia coli CS-109. PhageDNA wasradioactively labeled
by
including 0.5 mCi of
[3H]thymidine
(62.5Ci/mmol;
NewEngland Nuclear Corp., Boston, Mass.) in 120 ml of top agar. Phage were purified by two cycles of CsCl density gradient ultracentrifugation as described by Maniatis et al. (12). Electron-microscopic examination of
negatively
stained
preparations
revealed that phageparticles
were intact and that96%
or more had full (DNA containing) heads.Ion etching. Ar+ etching was performed in a modified Polaron model E5100 sputter coater. Plasmas were devel-oped ina
cylindrical
chamber14cmhigh
by15cmwide andcontaining
disk-shaped
aluminum electrodes 14 cmindiam-eter and separated by 4 cm. All etching experiments were
carried out at 5 mA in an atmosphere of 100% Ar at 100 mtorr(ca. 13.3 Pa).Phagetobe used forDNAanalysiswere
etchedon aluminumfoil (Reynolds 625) sheets(2by2cm). Phage
(1010)
(containing800 to1,000cpm of3H label) in5,ulof SM buffer (0.1 M NaCl, 8 mM MgSO4, 5 mM Tris hydrochloride[pH7.5]) werespread evenly on a foil square
(where they occupied less than a single phage monolayer)
which was then washed briefly (5 s) in 0.1 M ammonium
acetate and quickly frozen in liquid N2. Phage were then
lyophilized overnight and etched for various times. Direct measurement ofthe DNA remaining after etching was
ac-complished
by countingthe entire foil square in atoluene-based liquid scintillation fluid (Research Products
Interna-tional, Elk Grove Village, Ill.).
DNA extraction, analysis, and
32p
labeling. DNA was extracted from etched phage by incubating foil squares(containing etched phage) for 30 min at 37°C in 1 ml of a
solution containing 0.5 mgofpronase per ml, 0.5% sodium
dodecyl sulfate, and 5 mM EDTA(pH 8.0). This procedure was found to solubilize 85% or more of the phage DNA,
which was then purified by phenol extraction, followed by two cycles of ethanol precipitation. Purified DNA was
analyzed by electrophoresis on2%agarosegels (6 cm long;
Tris-acetate [pH8.0] buffer system), stained with 0.5 ,ug of
ethidium bromideper ml,andphotographed by UV transil-lumination as describedbyManiatis et al. (12).
32p
label was564
on November 10, 2019 by guest
http://jvi.asm.org/
*
k
*~~~~*;!t. 8 < +.> t;
:
Wi
-
*'
#',''
4~~~~~~~~~~~~~~~~~~~~.
c r;.ezKKEi4
*
*~
''s4
...*...
.3
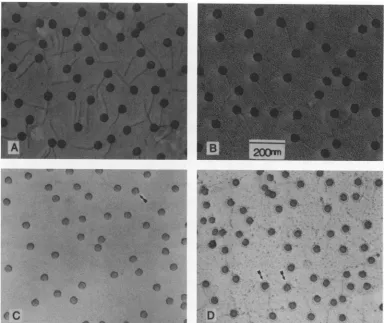
SFIG. 1. Electron-microscopic analysis of Ar+-etched X. Native phage particles were critical point dried and etched on electron-microscopic grids. After being etched, phage were shadowed with Pt-Pd and photographed in a JEOL 100cx electron microscope. Micrographs show unetched phage (A) and phage etched for 10 s (B), 1 min (C), and 3 min (D). Arrows indicate particles showing the characteristic equatorial constriction (see the text).
introduced into purified DNA by nick translation by using the method of Maniatis et al. (13) with [a-32P]dCTP (Amersham Corp., Arlington Heights, Ill.) as the sourceof
radioactive label. All enzymes (DNA polymerase I and
DNase I), buffers, and nonradioactive deoxynucleoside tri-phosphateswereobtained from Bethesda Research Labora-tories,Gaithersburg, Md. DNA substrate (0.2to1.0,ug)was
included ineach50-p1lreaction mixture, and after incubation (for 1 hat 150C), 32P-labeled DNAwasrecovered by three cycles of ethanol precipitation. Underthe conditionsused, intact Xvir DNAwaslabeledtoalevelof108dpm (or more)
per,g of DNA.
Southernhybridization. Southern hybridizationswere
per-formed with1.8-p.g samples of unmethylated ADNA(Sigma Chemical Co., St. Louis, Mo.) which were digested to
completion with AccI,
McI,
or NruI (GIBCO/BRL Life Technologies, Inc., Gaithersburg, Md.). Digestion productswere then heated to 65°C for 5 min and separated by electrophoresison0.85%agarosegelswhichwere rununtil the1,444-base-pairAccIfragmenthadmigrated10cm.After
electrophoresis, gels weredenatured in alkali,blottedonto
nitrocellulose sheets(5 by11cm)with lOx SSC(lx SSC is
0.15 M NaCl plus 0.015 M sodium citrate), and dried as
describedbyManiatisetal.(12).Blotswereprehybridizedin
6x SSC-0.5% sodiumdodecylsulfate-5x Denhardt solution
containing100 ,ugof denatured salmonsperm DNA(Sigma)
permlfor 4 hat65°Candhybridized overnightat65°Cin 5
mlof thesamemediumcontaining0.01M EDTA and at least
5 x
105
dpm ofdenatured, 32P-labeled probe DNA (from etchedphage). Afterhybridization,
filterswerewashed for2h at 65°C in several changes of 0.1x
SSC-0.5%
sodium dodecylsulfate, dried,
andradioautographed
with KodakXAR medicalX-rayfilm. Noradioactivity wasdetectedon
filters if denatured A DNA was omittedduring the
blotting
step.Electronmicroscopy.Phagesamplestobe examined in the electron
microscope
were adsorbed to carbon-Formvar-coated electronmicroscope grids,
criticalpoint
dried in a Tousimis samdri 780 criticalpoint
dryer,
and etched as described above on aluminum foil supports. Afterbeing
etched, phage
specimens
wererotary shadowed with Pt-Pd andphotographed
at50,OOOx
in a JEOL-100cx electronmicroscope
aspreviously
described(15).
RESULTS
Ar+ etching.Ar+etchingat5 mAfor1 to5minwasfound
toproducethetypeof uniform and
partial
erosionrequired
for our studies.Electron-microscopic
analysis
(Fig. 1)
ofphage etchedunder these conditions revealed that the head
remained
recognizably
intact as its diameter wasprogres-sively diminished.Asin thecaseof
T4,
erosionappeared
tobe
directionally
uniform(2).
The reduction in diameteramountedto
approximately
16%after 1 min and 30% after 3 min ofetching (Fig.
1C andD,
respectively). Phage
tails werevisibly
damaged
after 10 s ofetching (Fig.
1B)
andon November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.116.500.69.392.2]566 BROWN AND NEWCOMB
100
0
U.c
0
Cu
U
co *0
Cu
c._
c
a)
0.
80
60
40
20
1 2 3 4
Time of Etching (minutes)
[image:3.612.85.276.66.305.2]5
FIG. 2. Effect ofAr+ etchingontheamountofDNApresent in
theX head. PhagewhoseDNAhad beenradioactively labeledby growthin thepresenceof[3H]thymidinewerelyophilizedon
alumi-numfoilsupports, etched for the indicated times, and counted ina
liquid scintillation counter. Points show the average and extreme amounts of radioactivity remaining in at least six identical
determinations.
completely lost after 30to60s(Fig. 1C). Adistinct
constric-tionorcleftwasobserved inasignificant proportion of phage
particles etched for 1minormore(arrows in Fig. 1C and D).
The structural basis for the cleft is notknown, although it resembles asimilarconstriction observed in phageT4(2).
The amount of DNA remaining in etched phage was
determinedwithApreparations whose DNA had been radio-actively labeled by growth in thepresenceof[3H]thymidine.
Suchphage were etched on aluminum foil targets, and the
amountof DNA remainingwas determined by counting the
entire foil square. The results (Fig. 2) demonstrated that
DNAwas lost (by sputtering) rapidly during the first 1to 2
minofetching andmoreslowly thereafter. The resultsarein
qualitative agreement with the rate of erosion expected on
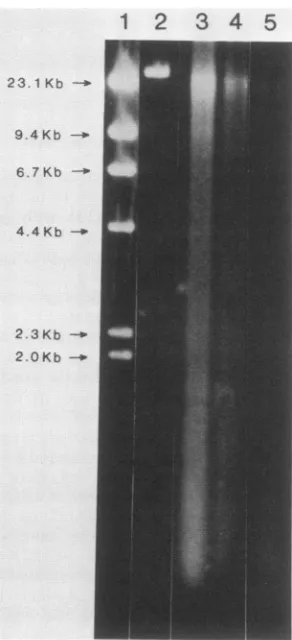
thebasisof the morphological observations shown in Fig. 1. Thephysicalstateof DNA isolated from etched phagewas
analyzed by agarose gel electrophoresis. Short periods of
etching that resulted in little DNA loss were nevertheless
foundtoproduce significant damagetoXDNA.After30sof
etching, for instance (Fig. 3, lane 3), no full-length DNA
molecules remained. Fragmentswerefound ina verybroad
distribution from approximately 20,000 to 150 base pairs, with evidence of concentrationsattheupperand lower ends
ofthedistribution. Longer periods of etching(Fig. 3, lanes 4 and5) producedanetloss ofDNAandadecrease inaverage
fragment length. No evidence of a discrete or preferred
fragment sizewasobserved afteranyof the etching periods
examined. Etching for periods of approximately 5 min or
longer resulted in the loss ofall detectable DNA.
DNA analysis by Southern hybridization. The genomic origin of DNA remaining in etched phage was analyzed by
Southernhybridization.DNAwasfirstisolated from etched
phage (see Materials and Methods)and labeled in vitro with
32pbynicktranslation(13). Labeled DNAwasthen usedto
probeSouthernblots ofA restrictiondigests.It wasexpected
that regions ofthe genome lost duringthe etching process would not be represented in the 32P-labeled probe and therefore would not label corresponding restriction frag-ments in theblot.
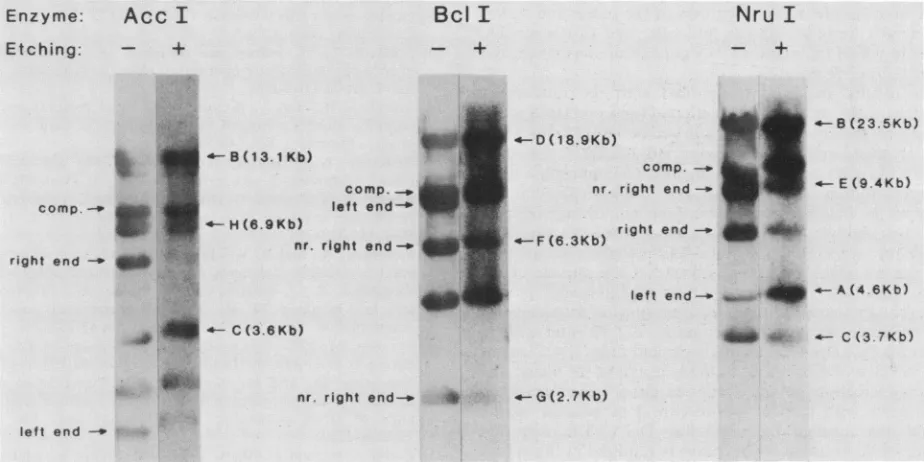
Southernhybridization was carried out with whole A DNA
digested separatelywithAccI,
BcI,
andNruI. Theproducts ofrestriction enzyme digestion (Fig. 4) were separated by agarosegel electrophoresisand blottedontonitrocelluloseas described in Materials and Methods. Hybridizations were performedat65°Cwith32P-labeledprobeDNA derived fromphage etched for various times and (as a control) from
unetched phage. The results (Fig. 5) demonstrated that
etchingwas correlated with a nonuniform labeling of
frag-ments in all three restriction
digests.
For instance, in the case ofthe AccI digest, fragments J (5.6kilobases [kb]), H(6.9 kb),and probably E (11.8 kb) werelabeled to alesser extentby theprobe preparedfrom etchedphagethanbythe
control probe. Other AccI fragments were labeled to a similarextent
by
thetwoprobes. Likewise, fragmentsF(6.3 kb) and G (2.7kb) of the Bclldigest
andfragmentsF(6.7kb),E (9.4 kb), and C (3.7 kb) of the NruI digest were more weaklylabeled bytheetchedprobe than
by
theunetchedone (Fig.5).As acontrol experiment, blots that had been
hybridized
with[32P]DNA
prepared from etchedphage
werereprobed
with
[32P]DNA
derived from unetchedphage.
Radioauto-1 2 3 4 5
23.1Kb -0
9.4 Kb
-6.7Kb
-4.4Kb -.
2.3Kb
-2.0 Kb-b
FIG. 3. Agarosegel electrophoresisof DNA isolated from
Ar+-etchedX.DNAwaspreparedfrom 2 x 1010 phage unetched (lane 2) oretched for30s(lane 3), 1min (lane4),or3min (lane 5). Lane 1 showsDNAsize markers obtainedfromaHindIII digest ofXDNA (GIBCO/BRL).
I I
II I I I~~~~~,
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.365.511.358.678.2]ARRANGEMENT OF DNA IN PHAGE A 567
Acc I
639 499
2191 13,070 3574 11,831 1444 6954 2720 5580
II
II
lll1l- l l
*
lrignhd
A B C D E FG H I J
BclI
517 1576
8844 s 4459 18,909 4623 6330 2684
1560
rI
lI
I
I
III
A B C D E F G Hi
Nru
I
704
4590 23,460 3653 9401 6694
A
I
C D EA B C D E F
FIG. 4. Restrictionmapsof DNAforAccI, Bc(I,and NruI.Mapsshowdigestion sites expectedonthe basis of the publishedsequence for XcIindlts857Sam7 DNA (16). Numbers indicatethe sizes of therestrictionfragments in base pairs.
graphs showed that the pattern of labeling observed after reprobing matched that seen when comparable blots were
hybridized with thecontrol probe only (datanotshown). DISCUSSION
Inspection of therestrictionmapsshown in Fig. 4reveals
acommon feature of DNA sequences underrepresented in
etchedphage. Allarederived from theright endorfrom the
right end to approximately the middle of mature DNA. This resultsuggeststhat there existsatleastsomedegreeof
order in the wayDNA isarranged in the phage head. More particularly, compared to the remainder of the DNA, the
Enzyme:
Acc
I
Etching: - +
Bcl
I
+
DNAoriginating from the right end of thegenome is found
systematically nearerthe surface of the overall DNAmass.
Thesuggestedarrangementof A DNA is therefore
compara-bletothatobserved(2) in phage T4 because in bothcasesthe
last DNApackaged into the prohead in vivo (the right end in the case ofA [7, 8, 17, 19]) is found atthe outside of the
matureDNA condensate.
The Southernhybridization results shown in Fig. 5
illus-tratethepoint thatwerarely observed restriction fragments, even those derived from the extreme right end of the
genome, that were completely unlabeled by DNA from
etchedphage. Reduced, butnotnegligible, labeling of
right-Nru I
+
6-B(23. 5Kb)
;Kb)
comp. _*
nr. right end- 4-E(9.4Kb)
) right end- _ -4-0D (18.
i * - B(1(I3.1 Kb)
comp. _ left
end-w
;_
~~H
(6.9Kb)nr. right
end-right end _M
-.Sll 4-C (3.6Kb)
--F(6.3 X
left end-p
t
_*.A(4A 6Kb)* -C(3.7Kb)
nr. right end-. * -G(2.7Kb) left end -_
FIG. 5. RadioautographofSouthernhybridization experiments performedwithArestrictiondigestsand DNAspreparedfrometchedand
unetched phage. Probe DNAs were prepared by isolatingDNAfrometched (AccI,5 min; BcllandNruI, 1 min) orunetchedphageand
labelingit with32Pbynick translation. Theywereused tohybridizeblots ofADNA that had beendigestedwith the indicatedrestriction
endonucleases.Hybridizationandradioautographywereperformedasdescribed in Materials andMethods.Selected restrictionfragmentsare
indicatedbyletterandsize(in kilobases). At the left of each blotareshown thepositionsof the twoendfragmentsand also thecomposite
(comp.) fragment produced when the left andrightends arejoinedat their cohesive sites; nr., near. Other restriction fragmentscan be
identified from themaps shown inFig.4.
left end VOL.60, 1986
OK
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.118.493.68.264.2] [image:4.612.76.538.429.660.2]568 BROWN AND NEWCOMB
endfragmentswas the rule. Forinstance, fragment Ffrom the NruI digest, fragment G from the BclI digest, and fragment H from the AccI digest are all derived from the
right endor nearthe right end ofthe genome andwere all labeledtoareduced, but notnegligible,extent. Webelieve thatthis is duetothe fact that thephageheadwasin contact
with its aluminum foil support during the etching process.
Contact is expectedtoshieldasmallpatch of externalDNA
from erosion so some should be represented in DNA
pre-pared from etched phage. This DNA would result in a
reduced but still detectable level of hybridization to
corre-sponding restriction fragments.
The results presented here raise a significant expectation
about thewayDNA is condensed into the A proheadinvivo. Since the left end of the genome is the first to enter the prohead (7, 8, 17, 19) and since it is also found protectedat
the center of the mature DNA mass, one expects that the prohead is most likely to fill beginning at the center and progressing toward the outside of the cavity. The last DNA packaged (the right end in thecaseofX) would then be found attheoutside of the DNA condensate. This expected polar-ity of packaging (i.e., inside to outside) is consistent with electron-microscopicimages of partially filled heads.Such structures, observed in infected cellsby thin sectioning (11)
orosmoticlysis (20), show DNA condensedatthecenterof incompletely filled A particles. Similar images are found in
T4-infectedE. coli (21)and in Salmonella typhimuriumcells infected with phage P22 (10).
The results reported above have important implications for thewayDNAisarranged in thematurephage head. We consider that there exist three general models forthe
orga-nization of phage DNA: (i) the concentric shell, solenoid,or
spool model in which DNA makes toroidalwindings about
anaxis thatmaybecoincident with (5, 9)orperpendicularto
(6) the long axis of the phage; (ii) the spiral-fold (2) and related (P. Serwer, J. Mol. Biol., in press) models in which DNArunsparalleltothe longaxisof thephage and makes
tight (180°) turns at regular intervals; and (iii) a random
model in which thematureDNA condensate hasnoregular
organizational features.
Our results are least compatible with the random or
"random-stuff" model. The location of right-end DNAatthe outside of the condensate would not be expected if DNA
were arranged entirelyatrandom; left-end DNA would be equally likely to occurat the outside. The concentric shell andspiral-fold models,however,arebothbasically
compat-iblewith thedata presented here. OnecanarrangeDNA in
eithertoroidalwindings (as in the concentric shell model)or
infolds orloops (as in the spiral-fold model) in sucha way
that the lastDNA packaged is found atthe outside of the overall structure.Inthe caseof the concentric shell model,
however, this arrangement creates special problems for DNA ejection by phages, such as X, T7, Ti, and 4)29, in
which the last DNA packaged is ejected first(1, 4). Unless thetoroidal axisis thesame asthelong axis of thephage, the
observed ejection polarity requires either rotation of the DNA mass with respect to the capsid or passage of the
exiting end through the remaining DNA. The fact that neither of these unlikelyprocessesisrequired in the
spiral-fold model provides a basis for favoring it at the present
time.
ACKNOWLEDGMENTS
We thank W. Studier for animportant suggestion about experi-mentaldesign, M. Smith and members of his research group for help with themolecularbiology, J. Boring for advice about ion etching methodology, and L. Black for a careful reading of themanuscript. This work was supported by Public Health Service grant GM34036 from the National Institutes of Health.
LITERATURECITED
1. Bjornsti, M.-A., B. E. Reilly, and D. L. Anderson. 1983. Mor-phogenesis ofbacteriophage 4)29 of Bacillus subtilis: oriented andquantized in vitro packaging of DNA protein gp3. J.Virol. 45:383-396.
2. Black, L., W. Newcomb, J. Boring, and J. Brown. 1985. Ion etching of bacteriophage T4: support for aspiral-fold model of packaged DNA. Proc. Natl. Acad. Sci. USA 82:7960-7964. 3. Black, L.,and D. Silverman. 1978.Modelfor DNApackaging
intobacteriophageT4heads. J. Virol. 28:643-655.
4. Earnshaw, W., and S. Casjens. 1980. DNA packaging by the double-stranded DNAbacteriophages. Cell 21:319-331. 5. Earnshaw, W., and S. Harrison. 1977. DNA arrangement in
isometricphageheads. Nature(London) 268:598-602.
6. Earnshaw, W., J. King, S.Harrison, andF.Eiserling.1978. The structuralorganization of DNA packaged within the heads of T4 wild-type,isometric and giant bacteriophages. Cell 14:559-568. 7. Emmons, S. 1974. Bacteriophage lambda derivatives carrying twocopiesof the cohesive end site. J. Mol. Biol. 83:511-525. 8. Feiss, M., and A. Bublitz. 1975. Polarized packing oflambda
chromosomes. J. Mol.Biol. 94:583-594.
9. Harrison, S. 1983.PackagingDNAintobacteriophage heads:a
model. J.Mol. Biol. 171:577-580.
10. Lenk, E., S. Casjens,J. Weeks,andJ. King.1975. Intracellular visualization of precursorcapsids inphage P22mutantinfected cells.Virology 68:182-199.
11. Lickfeld, K.,B. Menge,B.Hohn,and T. Hohn. 1976. Morpho-genesis ofbacteriophagelambda: electronmicroscopy ofthin sections. J. Mol. Biol. 103:299-318.
12. Maniatis, T.,E. F.Fritsch,andJ. Sambrook. 1982. Molecular cloning:alaboratory manual. ColdSpringHarborLaboratory, ColdSpring Harbor,N.Y.
13. Maniatis, T., A. Jeffrey, and D. Kleid. 1975. Nucleotide se-quenceof therightward operator of phage X.Proc.Natl. Acad. Sci.USA 72:1184-1188.
14. Murialdo, H., and A. Becker. 1978. Head morphogenesis of complex double-stranded deoxyribonucleic acid bacterio-phages. Microbiol. Rev. 42:529-576.
15. Newcomb, W., J. Boring, andJ. Brown. 1984. Ion etching of humanadenovirus2: structureof the core. J. Virol. 51:52-56. 16. Sanger, F.,A.Coulson,G.Hong,D.Hill,andG. Petersen.1982.
Nucleotide sequence ofbacteriophage lambda DNA. J. Mol. Biol. 162:729-773.
17. Sternberg, N.,and R. Weisberg. 1975. Packagingofprophage andhostDNAby coliphage X. Nature (London)256:97-103. 18. Streisinger, G., J. Emnich, and M. Stahl. 1967. Chromosome
structure in phage T4. III. Terminal redundancy and length determination. Proc. Natl. Acad. Sci. USA 57:292-295. 19. Syvanen, M. 1975. Processing ofbacteriophage lambda DNA
duringitsassembly into heads. J. Mol. Biol. 91:165-174. 20. Yamagishi, H., and M. Okamoto. 1978. Visualization of the
intracellulardevelopment of bacteriophage X with special
refer-ence to DNA packaging. Proc. Natl. Acad. Sci. USA 75:3206-3210.
21. Zachary, A., and L. Black. 1981. DNA ligase is required for encapsidation of bacteriophage T4 DNA. J. Mol. Biol. 149:641-658.
J. VIROL.