JOURNALOFVIROLOGY, Nov. 1984,p. 396-402 0022-538X/84/110396-07$02.00/0
CopyrightC 1984, American Society for Microbiology
Vol.52,No. 2
Large
Surface Proteins of Hepatitis
B
Virus Containing
the Pre-s
Sequence
KLAUS H. HEERMANN,' UDO GOLDMANN,1 WOLFGANG
SCHWARTZ,'
TORSTENSEYFFARTH,l
HORSTBAUMGARTEN,2 ANDWOLFRAM H. GERLICHl*
Departmentsof Medical
Microbiologyl
andImmunology,2 University ofGottingen, Gottingen, Federal RepublicofGermany
Received 6 June1984/Accepted 25 July 1984
Thesequenceof hepatitis B virus DNA containsan openreadingframe whichcodesforanot-yet-identified
protein ofatleast 389amino acids. Only the products starting atthe third(GP33/GP36)orthe fourth (P24/
GP27) initiation signal have been characterizedascomponentsofthe viral surface antigen. We foundalarger
protein, P39, and itsglycosylatedform,GP42, in hepatitis B virus particles and viral surface antigenfilaments. Immunological cross-reactions showed thatP39/GP42 is partially homologoustoP24/GP27and GP33/GP36.
The uniqueportion of itssequencebound monoclonalantibodies which had been induced by immunization with hepatitisB virusparticles. Proteolytic cleavage patternsand subtype-specific size differences suggested thatthe sequenceof P39startswiththefirstinitiation signal of theopenreading frame. Its amino-terminalpart(pre-s
coded)isexposed attheviral surfaceand, probably, is highly immunogenic. A model is presented of how the openreadingframe for the viral envelope leadstodefinedamountsof three different proteins.
Atypical consequence ofacute orchronic infection with
hepatitis B virus (HBV) is secretion of the viral surface
antigen (HBsAg) intotheblood of infected persons. Even in the presenceof efficientviral
replication, only
averyminor part ofthetotal HBsAg formsthe viralenvelope.
A largeramountis foundonfilamentsof20-nmdiameter and variable
length. By far themostHBsAgis presenton
small,
noninfec-tious, 20-nm particles (20). These
particles
are used inpurified formas a vaccine
against
HBV.There are two types of chronic carriers of
HBsAg.
A smallerfraction ofthecarriershashigh
titers ofHBV,
ahigh concentration of excess HBsAg, and soluble viral coreprotein (e-antigen)in the blood. Most adult
HBsAg
carriers have, however, very littleor noinfectiousHBVorfilamentsintheblood andonly moderate concentrationsof
HBsAg
20-nm
particles
and antibodiesagainst
eantigen.
Themecha-nism for the
suppression
of viremia in these carriers isunknown. Immune reactions
against
unidentified proteinsmaybe involved (1, 5).
HBsAg from both types of carriers consists ofa
major
protein, P24, and its
glycosylated
form,
GP27(19).
PurifiedHBsAg20-nmparticles from viremic carriers
have,
inaddi-tion,afurther
glycoprotein, GP33,
anditstwofold-glycosyl-ated form, GP36 (24,
25).
The amino-terminalprotein
se-quence ofP24 has been
analyzed (19)
andaligned
with the sequence of cloned HBV-DNA(6a,
17, 29).
Acoding
sequence of 226
triplets
ending
with a stop codon wasidentified forP24 (gene
s).
Gene sbegins only
at the fourthpossiblestartcodonofa
larger
openreading
frame(ORF),
soit ispreceded in phase
by
163or174codons, depending
onthe viral subtype (pre-s
region 28).
Recently,
itwas shown that GP33 consists of the P24 sequence and anamino-terminalpartofca. 55 aminoacids
(13a, 24).
Thefinding
ofanamino-terminal methioninein GP33
(14)
and thetranscrip-tion data(2, 22a)suggest thatthe sequenceof GP33 startsat
the third initiation signal of the
ORF,
which is 55 codons upstream of the fourthsignal (2).
The translationproducts
beginning
atthefirstorsecondstartcodon havenotyetbeen identified. The conservation of the pre-sregion
during
evolution(15)and the invitro
transcription
data(13)
make it*Correspondingauthor.
verylikely that at least one larger surface protein exists. In the search for this hypothetical protein, we analyzed
purified HBV particles and filaments in addition to 20-nm
particles. Immunization of mice with HBV particles led to
monoclonalantibodies,which selectively detectedtwolarge
viral surface proteins. Cross-reactions with P24 and GP33,
partial proteolysispatterns,glycosylation data, andsubtype
heterogeneities suggeststrongly that these twolarge surface
proteins are products of the total ORF and that the monoclo-nalantibodiesaredirected againstepitopes codedby the
pre-s region. We present models of how the six envelope proteins ofHBVarederivedfromonecontinuousORF and of how theirexpression may be regulated. Being
immuno-genic components of the viral envelope, the large surface
proteinsmay be ofgreatmedical importance.
MATERIALS AND METHODS
Purification of HBV and HBsAg particles. Plasma units (250 ml) from one HBV carrier were passed through a
column (10 by 120 cm) of Bio-Gel A5M (Bio-Rad
Labora-tories)with TNE (0.13 M NaCl, 0.01 M Tris-hydrochloride (pH 7.4), 0.001 M disodium EDTA). Samplesofthefractions close to thevoid volume were mixed with 0.3%
,B-mercap-toethanoland 0.5% Nonidet
P-40,
and the HBVcore antigen was determined by an enzyme immune assay (7). Positivefractionswere combined and centrifuged in anangle rotor (6
by90 ml) through a layer of 20% sucrose-TNE for 20 h at 25,000 rpm and 10°C. The pellets were suspended in 0.5 ml of TNE and layered on an S-shaped sucrose gradient with best resolution between 35 and 45% (wt/wt) sucrose. After 20 h at
34,000 rpm in an SW42 rotor, fractions were assayed for
HBsAgand viral core antigen. In electron microscopy, peak
fractions of viral core antigen consisted of HBV particles with less than 10% filaments. Peak fractions of HBsAg
consisted of>95% long filaments. HBsAg 20-nm particles with much GP33/GP36 were purified from Bio-Gel ASM
fractions containing the highest HBsAg activity by banding
in CsCl (9). These preparations were free of HBV or long
filaments.
Antisera. For raising anti-P24, purified HBsAg without
minor proteins (27) was treated for 30 min with 1% dithio-threitol (DTT) and 1% sodium dodecyl sulfate (SDS) and
396
on November 10, 2019 by guest
http://jvi.asm.org/
thendialyzed for2hagainst0.01% DTT-0.01% SDS in 0.13 NaCl. Foranti-GP33, purified HBsAgwith much GP33 but with undetectable GP42 was used without denaturation. Portions (100 pg) of HBsAg subtype ad were mixed with Freund complete adjuvant andinjected intramuscularlyinto guinea pigs. The injections were repeated with incomplete
adjuvant after 4 weeks. Serawere taken 10days later. Monoclonal antibody. Ten micrograms of purified HBV was mixed with complete Freund adjuvant and injected intraperitoneally into BALB/c mice. After 4 weeks the
injections were repeated with incomplete adjuvant, and 10
days later spleens were obtained for fusion with myeloma cell line P3-X63-Ag8.653 as described previously (10). Su-pernatants of
immunoglobulin-producing
hybridomas wereplacedon microtiterplateswhichwerecoatedwith 40 ng of HBV andfilamentsperwell.Bindingofmouse immunoglob-ulinwasdetected bytheaddition ofperoxidase-labeled anti-mouseimmunoglobulin(Dako, P161). Clone A18/7
(IgG,K)
was recloned and injected intoBALB/c mice forgrowth as
ascites tumors. Immunoglobulin G from ascites liquid was
purifiedby precipitation with 18%(wt/vol) sodium sulfate. Gel electrophoresis and immune staining (26). Purified
HBV orHBsAg was denatured by 2.5% SDS and 5% DTT
for 5 min at 100°C. Proteins were separated in 12 or 15%
polyacrylamide-N,N'-methylenebisacrylamide gelswith the
Laemmli buffersystem and 2 to 5 V/cm (23). After
electro-phoresis,theseparated proteinsweretransferredtoaporous
membrane (GVHP; Millipore Corp.) bytransversal
electro-phoresisat40 V in 0.025 M Tris(pH 8.3), 0.192Mglycine, and 15% (vol/vol) methanol. Nonspecific protein binding
was saturated with 20% fetal calf serum-TNE for 1 h.
Thereafter, asuitableamountofantibodywas addedfor1 h underagitation. After a thorough washing, 1251_ or
peroxi-dase-labeled second antibody against the first antibody in
20% fetal calf serum was added for 1 h. The presence of
antibody was visualized either by autoradiography or by
enzymatic staining with 0.01% diamino benzidine-0.06% H202-0.05 MTris-hydrochloride (pH 7.4).
RESULTS
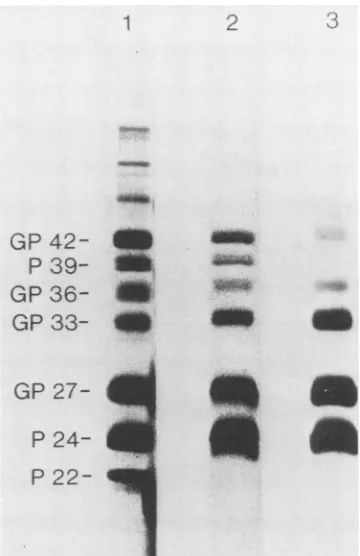
Proteincomposition of HBV particles, filaments, and 20-nm particles. All three morphological forms ofHBsAg consis-tently containedsixprotein bands: P24,GP27, GP33, GP36,
a39-kilodalton (kDa) protein, and a42-kDaprotein.
Experi-mentstobe described later showed that the 42-kDaprotein
was glycosylated, but the 39-kDa protein was not. Thus,
theyarereferredtoasP39 and GP42(Fig. 1). HBV particles
contained,inaddition, the P22 bandof the viral core protein
(7) and several weaker protein bands of>45 kDa (Fig. 1, lane 1) Mock preparations of HBV particles from negative human plasma also showed several high-molecular-weight
bands, but there were no bands atthe position of the viral
proteins.
The relative staining intensities of P24,GP27, GP33, and
GP36 were very similar in the three morphological forms.
Theintensities ofP39 andGP42 differedsignificantly, how-ever, among the three forms. In HBV particles and fila-ments,theGP42 band was more intense than the GP33 band; in 20-nmparticles, the GP42 band was weaker than the GP33 band. P39 and GP42 occurred in all isolates as pairs; GP42
was always more intense than P39. This typical staining
behavior of the three morphological forms was confirmed with samples from six further HBV carriers. The intensity of
the protein bands after silver staining suggests that HBV
particlesmaycontain up to 20 times more P39 and GP42 than do 20-nmparticles fromthe same plasma source. According
to the particle mass of 3 x
106
daltons, 20-nm particles1
GP
42-P
39-GP
36-GP
33-2
:
a.
Em
a.:9
GP
27-P
24-P
22-FIG. 1. Protein compositionof HBV particles (lane 1), HBsAg
filaments (lane 2), and20-nmparticles(lane 3). Theparticles were purified from the plasma ofa chronic HBV carrier (subtype ay). Sampleswith similarproteincontentswereelectrophoresed through 12%polyacrylamide,andtheproteinswerestained with silver(16). Size markerswerelysozyme(14.3kDa),trypsininhibitor(20kDa),
carbonic anhydrase (30kDa), ovalbumin(45 kDa),serumalbumin
(68kDa), and phosphorylase B (94kDa).
consist of ca. 100 protein subunits. The weak staining
suggeststhat only one oroccasionally twoGP42/P39 mole-cules may be present in 20-nmparticles. In contrast, HBV,
with itsfour-times-larger surface, may contain 40to 80 P39
orGP42 molecules per virion.
Monoclonalantibody againstHBV. Five micewere
immu-nized with purified HBV particles, and all developed high
serum
antibody
titers against HBsAg. Several thousandhybridoma cloneswere derived fromthe spleencellsofthe
mice,butonlytwoofthe clonesproducedantibodiesagainst
HBV. Neither antibody bound to mock preparations of
HBV. The antibodies were apparently directed
against
thesame or very
closely
relatedepitopes,
since mixtures of theantibodies did not
produce
strongerbinding
thansingle
antibodies. Oneofthemonoclonal antibodies
(MA18/7)
wasusedfor further
experiments.
Inatwo-site enzymeimmune assaywith MA18/7at thesolidphase
andperoxidase
conju-gated
aslabeledantibody,
HBVparticles
gavethe strongestsignal
of the threemorphological
forms. A2-fold-higher
protein concentration of
purified
filamentsora20-fold-larger
amount of 20-nm
particles
was necessary to bind the same amount oflabeledantibody
to the solid phase(Fig.
2). The HBV particles used for immunization had been ofHBsAg subtype ayw. MA18/7 reacted, however, withsubtype
adwas well.
The part of the viral envelope which bound MA18/7 was
sensitive to proteolysis by trypsin. A limited digestion of
HBsAg 20-nm particlescompletely abolished the bindingof MA18/7. In contrast, the binding ofaconventional anti-P24
antibody
increased slightly after the digestion (Fig. 3A).on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.354.534.75.353.2]398 HEERMANN ET AL.
1-l1 ng/ml
160
idooFIG. 2. Binding of labeled monoclonal antibody MA18/7 to in-creasing amounts of HBV (U), HBsAg filaments (0), or 20-nm particles(A).Wells of microtiterplates were firstcoatedwith 0.5 ng of MA18/7 each and thenincubated with the indicated dilutionsof purified particles for1 h at 37°C. Afterbeing washed, peroxidase-labeled(8)MA18/7 in1% bovineserumalbumin-phosphate-buffered saline was added for 1 h, and after further washings, ortho-phenylenediamine-H202assubstrate wasadded. Thecolored reac-tionproduct wasassayed at493 nm(E493).
Reduction oftheHBsAgparticles didnotchangethebinding of MA18/7, but most of the conventional anti-P24 did not
bind anylongerto theparticles (Fig. 3B).
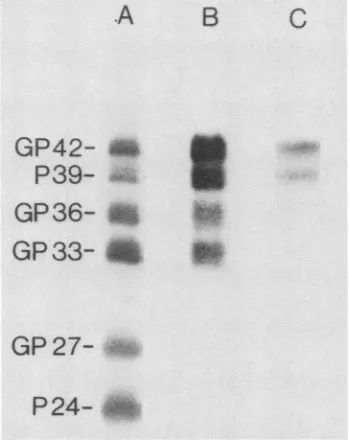
Immune reactivity of isolated HBsAg proteins. When the
immune blot technique was used after gel electrophoresis, MA18/7bound only to P39 and GP42
(Fig.
4, laneC).
Thisfinding showed that P39 and GP42 sharedan
epitope
which was not present onthesmallerHBsAgproteins.Thefinding
also excluded the possibility that the P39 and GP42 bands were mere aggregation artifacts consisting ofP24 orGP27. The epitope of MA18/7 was completely resistant to the
combined effects ofreduction,
detergent,
andheat,
which were employed beforeelectrophoresis.The sizes of P39 and GP42 were consistent with the
assumption that they might be
products
of the whole ORFfor HBsAg.
According
to thathypothesis,
P39 and GP42 would have asequence uniquetothem of108or 119amino acids, and MA18/7 would bind to thisregion.
In addition, theywould have all sequences and the denaturation-resistantepitopes of P24 or GP33. Two conventional
guinea
pigantiserawereusedto testthis
hypothesis,
because monoclo-nal antibodies against denatured P24 or the pre-s part ofGP33 were not available.
An antiserum against P24 and GP27 was raised
by
theinjection ofreduced and denatured
particles
which did nothave detectable
GP33, GP36,
P39,
or GP42. This anti-P24antibody
boundequally
well to all sixHBsAg
proteins,
includingP39 andGP42
(Fig.
4,laneA).
Asecond antiserumwas produced by the
injection
of 20-nmparticles
whichcontained much GP33 and GP36 but no detectable P39 or
GP42 (23). The injected animals
produced
moreantibody
against the pre-s part ofGP33 than
against
denatured P24,although the inoculum contained more P24 than GP33. At
the dilution used for the
experiment
shown inFig.
4,lane B,immunoglobulinG fromthis antiserum bound wellto GP33
and GP36 butweaklytoP24andGP27. P39and GP42were also well immune stained
by
thisantiserum.Insummary, GP42 andP39are
indistinguishable by
immu-nological methods. The two
proteins
have threeantigenic
regions: oneisdefinedbythe
HBV-specific antibody
MA18/7, the second is coded
by
the pre-s part ofGP33,
and thethird iscoded bygenes.
Proteolytic cleavage between the pre-s and gene s coded
sequences. Evidence on the
primary
structure of P39 andGP42 was obtained by digestion with the glutamic acid-specific protease from Staphylococcus aureus V8. Gene s has only two glutamic acids at positions 2 and 164; the pre-s sequence does not have any glutamic acid. Previously, we have shown that GP33 or GP36 is slowly cleaved by V8 protease into P24 or GP27 and an 11-kDa fragment which consistsof the 55 pre-s-derivedamino acids and of N-linked
glycan (24). If P39 and GP42 contain the whole pre-s sequencelinkedtothegene s sequence, V8 protease should generate a fragment of164 or 175 amino acids which binds
MA18/7. Figure 5, lanes B, shows that such a fragment of 18 kDa was generated. Since both GP42 and P39 were cleaved by the protease, the production of only one fragment react-ing with MA18/7 showed that P39 and GP42 were completely identicalin their pre-s part.
Subtype heterogeneity of the pre-s sequence. All known
DNA sequences of different HBsAg subtypes specify a constant size of gene s. The pre-s sequences of the two
sequencedaywsubtypeshave 163codons(6a, 17). The adw2
subtype is largelya homologof subtype ayw, but it has an
additional 11 codons at the 5' end of the ORF (29). In
agreementwith the DNA sequence data, P39 and GP42 from four adwsubtypeisolates were consistently larger than those from two ayw subtypes (Fig. 6, lanes B). The difference was ca. 1.0 to 1.5 kDa.
Asubtype-independent microheterogeneity of the HBsAg proteins has been described previously. This heterogeneity resides inthe gene s part (23) and is also visible in P24/GP27 ofthe isolates shown here. A smaller GP33 or GP36 from ayw subtypes was not noted (Fig. 6, panel A), and so the observed subtype-specific size heterogeneity is not due to the sequences present in the gene s part. The subtype differences between adw and ayw suggest that the sequence of P39 andGP42 starts with the very first codons of the ORF for HBsAg.
Glycosylation of the HBsAg proteins.Asensitive,
glycopro-1'
[image:3.612.60.299.75.207.2]6o
* b1i0
- +FIG. 3. Effects of trypsin (A) and DTT (B) on the bindingof MA18/7(0) and anti-P24(0). PurifiedHBsAg20-nmparticles (0.8 ,ugin100,ul) with relatively much GP42wereadsorbedtomicrotiter
wells(NuncII) for 4 hat20°C. The wellswerewashed,and 100p.lof
differenttrypsin dilutionsorof0.1 M DTTwasadded for 30minat
37°C.ReducedSHgroups wereblocked with 0.1 M iodacetamide for 16 h at4°C. Wellswere washed, anda 1:8 dilution ofhybridoma
supernatant MA18/7 or a 1:8,000 dilution of anti-P24 guinea pig
serumin1% bovineserumalbumin-TNEwasadded for 1 hat37°C.
Afterathorough washing, bindingof the antibodieswasquantitated by the addition of peroxidase-labeled second antibody (1:1,000 Dako) and by the measurement of bound peroxidase at 493 nm
(E493)-E493 7
/-,/
A A B
jig/ml
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.356.526.452.618.2]PRE-s PROTEINS OF HEPATITIS B VIRUS 399
-A
B
C
GP42-
P39-
GP36-GP33- A
P
GP 27-
AP24-
A
FIG. 4. Immune staining ofHBVsurface proteinswithanti-P24 (laneA),anti-GP33 (lane B), and MA18/7(laneC). PurifiedHBsAg
filaments(ayw) weredenatured,and theproteinswereseparated by
electrophoresisandtransferredtoporousmembranes. LaneAwas stained with a 1:50 dilution of anti-P24 serum, lane B wasstained with a 1:400dilution of anti-GP33 serum, andlane Cwas stained with a 1:10 dilutionofhybridomasupernatant.
tein-specific stain (3) showed that GP42 was glycosylated,
but P39 was completely glycoside free (Fig. 5, lane C). In
agreement with the conclusion drawn before, the 18-kDa
fragment of the glu-specific cleavage also did not contain glycan. Thus,theglycanof GP42 is boundtothegenespart, mostprobablyatthesamesiteasin GP27 (18)orGP36(24).
The amino-terminal (24) mannose-rich glycan (22, 23) of
GP33 or GP36 is apparently nonexistent in the products of
the whole ORF.
The nature of the glycan in GP42 was studied further by digestion with endoglycosidase F. This enzyme removes
asparagine-linked glycans from their protein part (4). The
electrophoretic mobility of
glycoproteins
is shiftedby
3 kDa for everyglycanremoved. Figure 5,lanesD,show that GP42decreased after digestion and P39 increased. GP36
disap-pearedcompletely, GP33 decreasedstrongly,andanintense
P30 band appeared. This
finding
supports the conclusions thatGP42 contains oneasparagine-linked glycan group and that this group is the only difference from P39. The results shown in Fig. 5, lanes D, directly confirm our previousreport(24)thatGP33 hasoneglycangroupandGP36hastwo
glycans.
DISCUSSION
Theprotein composition of HBsAghasbeenthe
subject
ofnumerous studies. Due to the variability of the HBsAg proteins (23), discrepant results onthe minor
proteins
werereported. Theelectrophoretic components largerthan GP42 are now understood asdimers (11, 23), and inthis
study
nosuchproteinscould be
reliably
identified asHBsAg
orHBV components. Feitelson et al. found the minor HBsAgpro-teinsp43,p35,andp32,whichprobably correspondtoGP42, GP36, and GP33. These
proteins
shared manytryptic
pep-tides with P24 or GP27, but they also had unique peptides(6). Sanchezetal. demonstrated serological cross-reactions
between theminorproteins (p27, p31, p35,andp40)andthe
major proteins (p22 and p25). They suggested "repeating antigenic determinants" in all HBsAg proteins (21). These findings are consistent with our data. Stibbe and Gerlich (24) showed that limited proteolysis of GP33orGP36generated
the same fragments as proteolysis of P24 or GP27, but the
twolarger proteins hadanamino-terminal extension of 50 to 60aminoacids,with threeproteolytic cleavagesites predict-ed by the pre-s(2) sequence (see Fig. 7 for definition).
Machidaetal. (14) alsofoundtwominorglycoproteins, p31
and p35, to be coterminal with P24. They showedbyamino acid analysis that these proteins contained a cyanogen bromide fragment which was coded by thepre-s(2) region.
Thefragment carries the HBV-associated receptor for
cross-linked human albumin (13a). Neurath et al. synthesized a
peptide
containing the 26amino-terminal aminoacids of the pre-s(2) region. Antibodies against this peptide bound to GP33 and GP36 (16a).The major mRNA of HBV in infected liver starts (2, 22a)
closelyupstreamof the thirdinitiation codon oftheORF(17, 29), and soit is very likelythatGP33 and GP36 also start at
A
B
1 2
*iv-i
42-_
39--
M--
___
~~~-
- 42 -___--39
10
36- AN
-33_-__w
-42 -39 -36 -33 -30 -27 -24 27
24
- 18 >
FIG. 5. Generation ofan 18-kDa pre-s-coded protein fragment by V8 protease(lanesB), concanavalin A binding of HBV surface glycoproteins(lane C), and removal of glycan by endoglycosidase F (lanes D). (A) Silver stain. (B) HBsAg filaments (adw2) were digested with6 ,ugof V8 protease (Bio-Rad) in 25,ulof0.1% SDS-1% DTT-0.1 MTris-hydrochloride (pH 7.4) for 72 h at 37°C. Each 24h,newprotease was added. The digested proteins were separated by SDS-gel electrophoresis, transferred to a membrane, reacted with MA18/7 and 125I-labeled anti-mouse immunoglobulin G, and autoradiographed. Lane
Bi,
Control incubation without protease; lane B2, protease digest. (C) Before treatment with MA18/7, the membrane withthe separated proteins was incubated with 100u.gof concanavalinAperml and10 mgofbovine serum albumin per ml in 0.13 M NaCI. After 30 min at 37°C, the washed membrane was agitated with 30 ,ug of horseradish peroxidase (type VI; Sigma Chemical Co.)perml in 0.13 M NaCl. After a further 30min and subsequent washings, the bound peroxidase was detected as de-scribed in thetext. LaneCshows the staining of lane B2. Note the absence of stainingin P39 and the 18-kDa fragment (-). (D) In a parallel experiment, filamentsweredigestedwith5,u1
of endoglyco-sidaseF(Bio-Rad) in35 ,ul of 0.1 M sodium phosphate (pH6.1)-S50mM disodium EDTA-0.1% SDS-1% Nonidet P-40-1% mercap-toethanol for16h at 37°C. The proteins were separated, transferred
to amembrane, and immune stainedwith anti-P24 serum. LaneDl, Control incubation; lane D2, endoglycosidase F digest. Note the increase ofP39and theformation ofP30 in the digest.
VOL. 52,1984
c
D
1 2
I
.Im
O'
A
ifk- 00
.Awbkl
41mw
A"
mm IRW
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.80.255.65.285.2] [image:4.612.305.548.282.490.2]400 HEERMANN ET AL.
l 2 3 4 5 6
(- fv&4* R;--42
*ift'" Oa0080 A -39
27_-inI
P
24-A
B'.
FIG. 6. Different sizes of GP42 and P39 in HBsAg samples of different subtypes. Proteins ofpurified20-nmparticleswere separat-ed induplicate bySDS-gelelectrophoresis. Panel A of thegelwas stained with silver. Panel B was transferred to a membrane and immune stained with MA18/7.Lanes1 and3,Subtypeay;lanes2,4, 5, and 6, subtype adw. Note the smallerdistance between GP33/ GP36 and P39/GP42 inpanelAand the fastermigrationof P39/GP42 inpanelB withtheaysubtypes.
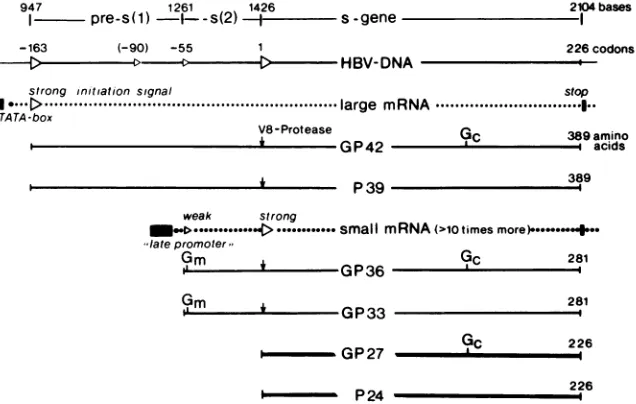
this translation signal (Fig. 7). The first AUG codon of this mRNA,however,lacks thetypicalflankingbases ofastrong
initiation codon (12),soproteinsynthesiswillprobablymore
often begin at the start of P24 or GP27 with its typical
initiation codon. By such a mechanism the two coterminal
proteins with different amino termini may be translated in
defined proportions from one mRNA. All results of this
study confirm the hypothesis that P39 and GP42 are the
translation products of the whole ORF. Thus, atleastsmall amounts of a mRNAstartingupstream of the first initiation codon of theORFmustbepresentinHBV-producingcells,
although it has not yet been found in
HBsAg-positive
liversamples (2). Such a mRNA was, however, presentin COS
cellstransformed withtheappropriate DNAfragments(13). Sincethefirst initiation signal isstrong, thesmaller HBsAg
proteins
are probably not derived from this larger mRNA. The second AUG in the ORF ofsome HBV isolates is notconserved andisprobablynotaninitiationsignal.Thelarge
mRNA has a promoter with a TATA box (13), but the
promoter of the small mRNA is more like the late simian
virus40promoter(2). Thus, expressionofP39/GP42maybe
completely independent
ofexpression ofthesmallerHBsAgproteins.
Since designations based on electrophoretic sizes are
ambiguous
formany reasons, itmaybe preferabletodistin-guishthe threetranslation products ofthe ORF forHBsAg
as
large, middle,
and small(ormajor) surface proteins. ThesequenceoftheORFmaybedivided into three
independent-lyexpressed parts:pre-s(1),presentonlyin the largesurface
protein;
pre-s(2),presentalsoin themiddle protein;andgene s, as suggested previously (28). The expression of threedifferent envelope proteins by the variableuse ofinitiation
codons inoneORFcertainlysavesmanyhundreds of
coding
capacity triplets
and regulating signals. The geneticorgani-zation of thisORFis another example-in additiontotheuse
ofoverlapping ORFs-thatthe HBVgenomeis
evolutionari-ly
selectedto minimum size.The glycosylation patterns of the three surface proteins
show interesting differences. Approximately 40% of the smallproteinisglycosylatedatthe carboxy-terminal
binding
site for N-linked glycans. As suggested by the staining intensity oftheprotein bands,thesamesiteis used less often in the middle protein, but probably moreoften inthe large
protein. The single binding siteof the middle protein in the
pre-s(2)region is alwaysoccupied by amannose-richglycan,
andnoglycoside-free middle proteinP30isfound in serum.
The same site, however, is not used at all in the large
protein. This observation demonstratesthe stronginfluence of an amino-terminal sequence on the processing of the
following
sequences.Ourfindings with sevendifferent isolates suggestthat the
large surfaceproteinsareessentialcomponents of
complete
-HBV-DNA
2104bases -I
226codons
strong initiation signal stop
I
....D...large
mRNA... FTA TA-box
V8-Protease
P39
Gc 389 amino
acids
389
weak strong
.-t.... ...>...small mRNA(>1otimes
more)..-*+--late promoter,
04
9~~~~~~~c
281GP36 c
GP33 281
GP27
P24
YC
226226 l
FIG. 7. Regulated expression ofsurfaceproteinsfrom HBV DNA andstructureof theHBsAg proteinsinrelationtotheircodingORF.
TheDNAsequenceof the ORF (top line) is from Paseketal.(17). The positionof the four5'-proximalinitiation sites(strong [>]andweak
[>]) for translationand the numbering of codons in relationtogenesareshown in the second line. The secondsignalisnotconserved inother HBVDNAs(6a, 29).Mappingof thetwopostulatedmRNAsisexplainedin thetext.Thebindingsitesof N-linked mannose-richglycans (Gm)
andcomplex glycans(Gj)weremappedpreviously (24).Theexactterminalmappositionsof P39/GP42 in theHBsAg particlesorHBVarenot
yetknown, but theyareprobably identicaltoorclosetotheproposedsites.
1 2 3 4 5 6
GP42- _..
P
39-GP36- _
GP 33- -_ __
GP
947 1261 1426
I pre-s(1)
1--s(2)
ss-gene,-163 (-90) -55
J. VIROL.
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.62.300.76.243.2] [image:5.612.152.470.480.682.2]HBV orof filaments, butnotofHBsAg20-nmparticles. The binding ofMA187 to native HBV or HBsAg in an enzyme immune assay shows that at least parts of the pre-s(1)
sequencearelocatedatthe surface. Therapidinactivationof
the MA18/7 epitope by trypsin is consistent with its exposed position, and it is also inagreement with the large numberof basic amino acids in the pre-s(1) region. Previously, it was shown that the pre-s(2) region ofGP33/GP36 is also at the
outer side of HBsAgparticles (24, 25), and the data on the albumin receptor coded by the pre-s(2) region confirm this
observation (13a).
As seen in the immune blots, both theepitope ofMA18/7 and at least one major epitope of pre-s(2) are resistant to
denaturation by heat, SDS, and DTT. These epitopes are apparently more immunogenic than are the epitopes of
denatured P24. It is likely that the denaturation-resistent epitopes are defined by the primary sequence, irrespective
of the protein conformation. This is insharp contrast tothe
antigenicityofP24, which is highly dependentonthe confor-mation specified by its synthesis in eucaryotic cells. In the
search for immunogenic and protective poly- or oligopep-tides as analternate vaccine against HBV, sequences of the pre-s region may be of interest. Recently, Neurath et al. (16a) showed that the 26 amino-terminal amino acids in the pre-s(2) region act as ahighly efficient immunogen. Testing
cellular(5) and humoral(1)immunity in hepatitisB
convales-cents, itwasnoted that HBV hadanantigenic component(s)
which was absent or rare in HBsAg 20-nm particles. Most
interesting, the immune reaction against the HBV-specific antigenwas loworabsent in viremic HBsAg carriers, butit
waspresentin nonviremic carriers. Accordingtoourresults,
the pre-s(1) part ofthe large surface protein could be that
antigenic component.
ACKNOWLEDGMENTS
We thank R. Thomssen and 0. Gotzeforgenerous support, K. Lechte for technical asistance, and R. Stute for supplying HBV-containing plasma.
Thisworkwaspartiallysupportedby theDeutsche Forschungsge-meinschaft.
LITERATURE CITED
1. Alberti, A., S. Diana, G. H. Scullard, A. L. W. F. Eddlestone, and R. Williams. 1978. Detection of a new antibody system reacting withDane particles in hepatitis Bvirus infections. Br.
Med. J. 2:1056-1058.
2. Cattaneo, R., H. Will, N. Hernandez, and H. Schaller. 1983.
Signals regulating hepatitis B surface antigen transcription. Nature (London)305:336-338.
3. Clegg, J. C. S. 1982. Glycoprotein detection in nitrocellulose
transfers ofelectrophoretically separated protein using
conca-navalinAand peroxidase. Application toarenavirus and
flavo-virus proteins. Anal. Biochem. 127:389-394.
4. Elder, J. H., and S. Alexander. 1982.
Endo-p-N-acetylglucosa-minidaseF: endoglycosidasefrom Flavobacterium
meningosep-ticum that cleaves both high-mannose and complex glycopro-teins. Proc. Natl. Acad. Sci. U.S.A. 79:4540-4544.
5. Fattovich,G., A.Alberti, C. Crivellaro, P. Pontisso, F.Noventa,
andG. Realdi. 1983. Cellular immunitytothehepatitisBvirion
inacutehepatitistype B.Clin. Exp. Immunol. 53:645-650.
6. Feitelson,M. A., P. L. Marion, and W.S. Robinson. 1983. The nature of polypeptides larger in size than the major surface
antigen components of hepatitis and like viruses in ground
squirrel, woodchuckand ducks. Virology 130:76-90.
6a.Galibert,F., E. Mandart, F. Fitoussi, P. Tiollais, and P.
Char-nay. 1979. Nucleotide sequence of the hepatitis B genome (subtypeayw) cloned inE. coli.Nature (London)281:646-650. 7. Gerlich, W. H., U. Goldmann, R. Muller, W. Stibbe, andW. Wolff. 1982.Specificityand localizationof the hepatitisB
virus-associated proteinkinase. J. Virol. 42:761-766.
8. Gerlich, W. H., and W. Luer. 1979. Selective detectionof IgM-antibody against core antigen of the hepatitis B virus by a modified enzyme immune assay. J. Med. Virol. 4:227-238. 9. Gerlich, W. H., and R. Thomssen. 1975. Standardized detection
of hepatitis B surface antigen: determination of its serum concentration in weight units per volume. Dev. Biol. Stand. 30:78-87.
10. Kearney, J. F., A. Radbruch, B. Liesegang, andK. Rajewsky. 1979. A new mouse myeloma cell line that has lost immunoglob-ulin expression but permits the construction of antibody-secret-ing hybrid cell lines.J. Immunol. 23:1548-1550.
11. Koistinen, V. U. 1980. Hepatitis Bsurface antigenpolypeptides: artifactual bands in sodium dodecyl sulfate-polyacrylamide gel electrophoresis caused by aggregation. J. Virol. 35:20-23. 12. Kozak, M. 1983. Comparison of initiation of protein synthesisin
procaryotes, eucaryotes, and organelles. Microbiol. Rev. 47:1-45.
13. Laub,O.,L. B. Rail, M. Truett,Y. Shaul, D. N. Standring, P. Valenzuela, and W. J. Rutter. 1983. Synthesis of hepatitis B surface antigen in mammalian cells: expression of the entire gene and the coding region. J. Virol. 48:271-280.
13a.Machida, A., S. Kishimoto, H. Ohnuma, K. Baba,
Y.
Ito, H. Miyamoto, G. Funatsu, K. Oda, S. Usuda, S. Togami, T. Nakamura, Y. Miyakawa, and M. Mayumi. 1984. Apolypeptide containing 55 amino acid residues coded by the pre-s region of hepatitis B virus deoxyribonucleic acid bears the receptor for polymerized human as well aschimpanzee albumins. Gastroen-terology 86:910-918.14. Machida, A., S. Kishimoto, H. Ohnuma, H. Miyamoto, K. Baba, K. Oda, T. Nakamura, Y. Miyakawa, and M. Mayumi. 1983. A hepatitis B surface antigen polypeptide (P31) with the receptor for polymerized human as well as chimpanzee albumins. Gas-troenterology 85:268-274.
15. Mandart, E., A. Kay, and F. Galibert. 1984. Nucleotide se-quence of a cloned duck hepatitisB virus genome: comparison with woodchuck and human hepatitis B virus sequences. J. Virol. 49:782-792.
16. Merill, C. R., D. Goldmann, S. A. Sedmann, and M. H. Ebert. 1981. Ultrasensitive stain for proteins in polyacrylamide gels shows regional variations in cerebrospinal fluid proteins. Sci-ence 211:1437-1438.
16a.Neurath, A. R., S. B. H. Kent, and N. Strick. 1984. Location and chemical synthesis of a pre-S gene coded immunodominant epitope of hepatitis B virus. Science 224:392-394.
17. Pasek, M., T. Goto, W. Gilbert, B. Zink, H. Schaller, P. McKay, G. Leadbetter, and K. Murray. 1979. Hepatitis B virus genes and their expression in E. coli. Nature (London) 282:575-579. 18. Peterson, D. L. 1981. Isolation and characterization of the major
protein and glycoprotein of hepatitis B surface antigen. J. Biol. Chem. 256:6975-6983.
19. Peterson, D. L., J. M. Roberts, and G. N. Vyas. 1977. Partial amino acid sequence of two major component polypeptides of hepatitis B surface antigen. Proc. Natl. Acad. Sci. U.S.A.
74:1530-1534.
20. Robinson, W. S. 1977. The genome of the hepatitis B virus. Annu. Rev. Microbiol. 31:357-377.
21. Sanchez, Y., I.Ionescu-Matiu,G. R. Dreesman, F. B. Hollinger, and J. L. Melnick. 1981. Evidence for the presence of repeating antigenic determinants in the major and minor polypeptides derived from hepatitis B surface antigen. Virology 114:71-80. 22. Skelly, J. C., C. R. Howard, and A. J. Zuckerman. 1978. The
labelling of galactose residues in hepatitis B surface antigen glycoprotein. J. Gen. Virol. 41:447-457.
22a.Standring, D. N., W. J. Rutter, H. E. Varmus, and D. Ganem. 1984. Transcription of the hepatitis B surface antigen gene in cultured murine cells initiates within the presurface region. J. Virol. 50:563-571.
23. Stibbe, W., and W. H. Gerlich. 1982. Variable protein composi-tion of hepatitis B surface antigen from different donors. Virolo-gy 123:436 442.
24. Stibbe, W., and W. H. Gerlich. 1983. Structural relationships between minor and major proteins of hepatitis B surface anti-gen. J. Virol. 46:626-628.
on November 10, 2019 by guest
http://jvi.asm.org/
402 HEERMANN ET AL.
25. Stibbe, W., and W. H. Gerlich. 1983.Characterization ofpre-s geneproducts in hepatitis B surface antigen. Dev. Biol. Stand. 54:33-43.
26. Symington,J., M.Green, and K. Brackmann. 1981. Immunoau-toradiographicdetection ofproteinsafterelectrophoretic
trans-ferfromgels to diazo-paper: analysis of adenovirus encoded proteins. Proc. Natl.Acad. Sci. U.S.A. 78:177-181.
27. Thomssen, R.,W.H, Gerlich,U. Bottcher, K.Legler,S.Ritter,
W. Stibbe, W. Weinmann, 0. Klinge, and U. Pfeifer. 1983.
Safety andpotencyaspectsinthe preparation ofan experimen-tal HBsAg vaccine. Dev. Biol. Stand. 54:23-31.
28. Tiollais, P., P. Charnay, and G. N.Vyas. 1981. Biologyofthe hepatitisB virus. Science 213:406-411.
29. Valenzuela, P., M. Quiroga, J. Zaldivar, P. Gray, and W.J. Rutter. 1981. The nucleotide sequenceof the hepatitis Bviral
genomeandtheidentificationof the major viralgenes,p.57-70T
In B. Fields, R. Jaenisch, and C. F. Fox (ed.), Animal virus genetics. Academic Press, Inc., NewYork.
J. VIROL.