0022-538X/89/031194-09$02.00/0
Copyright ©1989,American Society forMicrobiology
Mapping of RNA-
Temperature-Sensitive Mutants of Sindbis Virus:
Complementation Group F Mutants Have Lesions in nsP4
YOUNGS. HAHN,' ARASH GRAKOUI,2 CHARLES M. RICE,2 ELLEN G. STRAUSS,' ANDJAMES H.
STRAUSS'*
Division of Biology, CaliforniaInstitute of Technology, Pasadena, California 91125,1 andDepartment of Microbiology and
Immunology, Washington
University,
St. Louis, Missouri 631102Received 24 October 1988/Accepted 28 November 1988
Temperature-sensitive (ts)mutantsof Sindbis virusbelongingtocomplementationgroupF, ts6,tsllO, and ts118,aredefectivein RNAsynthesisatthenonpermissivetemperature. cDNA clones of thesegroupFmutants, aswellasofts+revertants, have beenconstructed. To assign thetsphenotype toaspecific regioninthe viral
genome,restriction fragmentsfromthemutantcDNA cloneswereused toreplacethe corresponding regions
ofthefull-lengthcloneTotol101 of Sindbis virus. Thesehybridplasmidsweretranscribed in vitroby SP6 RNA polymerasetoproduceinfectious transcripts, andthe virusrecoveredwastested fortemperature sensitivity. After thetslesion of eachmutantwasmappedtoaspecific regionof 400 to 800nucleotides by thisapproach,
thisregion ofthecDNA clonesofboth thets mutantandts+ revertantswassequencedin order todetermine
the precisenucleotide changeand amino acid substitution responsible for each mutation. Rescuedmutants,
which have a uniform background except for one or two defined changes, were examined for viral RNA synthesis and complementationtoshow that thephenotypesobservedwerethe resultof the mutationsmapped.
ts6andtsllOhadasinglebasesubstitution innsP4,resultinginreplacementofGlybyGlu atposition 153or position 324, respectively.It is of interestthat nsP4contains theGly-Asp-Aspmotifcharacteristic ofanumber ofviralreplicases, andthis, togetherwith the fact that all RNAsynthesis ints6-infected cellsand,toalesser
extent, in tsllO-infected cells shut off when the cells were shifted from a permissive to a nonpermissive
temperature, suggeststhatnsP4 isthe viruspolymerase.ts118was adouble mutant. It containedasinglebase
substitution in nsP2, resulting inreplacementofValbyAla atposition425 thatresulted in theformation of minute plaques, but not in a reduction in the plaque number atthe nonpermissive condition. The second change, asubstitution of Glnby Argints118atresidue 93 in nsP4, had littleapparentphenotype onitsown,
but incombination with thechangeinnsP2 led toatsphenotype. Thus,in eachcasethe mutationresponsible forthe temperaturesensitivityof the three knowncomplementationgroupF mutantslayin nsP4. Inaddition, theresult withtsll8suggeststhatnsP2and nsP4mayinteractwitheachother inacomplex.
Sindbis virusis a well-studied memberofthe alphavirus family. Its genome isa single molecule of plus-strand RNA 11,703nucleotides in length thatis capped at the 5' end and
polyadenylated at the 3' end (35). During replication, the
parental49Splus-strand RNA istranscribed into a comple-mentary minus strand which serves as a template for the
synthesis of both 49S plus-strandgenomic RNA and a 26S subgenomic RNA. Nonstructural polypeptides are trans-latedfromthe genomic 49S RNA as two polyprotein precur-sors that are processed by cotranslational or
posttransla-tionalcleavage into fournonstructuralproteins, callednsPl,
nsP2, nsP3, and nsP4, which are required for RNA
replica-tion (11). Three structural polypeptides are produced by
processing of a polyprotein precursor translated from the
subgenomic26S mRNA.
Large numbers oftemperature-sensitive (ts) mutants of the HR strain of Sindbis virus have been isolated and characterized (3, 30, 34). Mutants may be defective in RNA
replication (RNA- mutants) or in the production of the structuralproteins
(RNA'
mutants) andhave been groupedby complementation into four RNA- groups (A, B, F, and
G) and three RNA' groups (C, D, and E) (4, 30). Repre-sentative mutant-revertant pairs from RNA' groups have been analyzed by sequence analysis, and there is an excel-lent correlation between specific nucleotide changes and phenotypes (1, 10, 15). None of the RNA- mutant defects
*Corresponding author.
has been rigorously assigned to specific nonstructural
pro-teinsorRNA sequences.Thesemutantspresumably contain
tslesions in the viral nonstructural proteins whichfunction to replicateviral RNA.
A full-length cDNA clone of Sindbis virus has been
constructed that can be transcribed in vitro by SP6 RNA
polymerasetoproduceinfectiousfull-length transcripts (22).
Viruses produced from in vitro transcripts are identical to Sindbis virus and showstrain-specificphenotypesreflecting
the source ofRNAusedforcDNA synthesis (17, 22). This full-lengthclonecanbe usedtostudyinterestingphenotypes
ofSindbis virus.
Wehaveusedthisapproachtodefineprecisely the
muta-tions responsible for the ts phenotypes of Sindbis virus
complementation
groupF mutants. Mutants ts6, tsllO, and tsll8 of complementation group F are defective in RNAsynthesis atthenonpermissivetemperature. The
best-char-acterized member, ts6,ceases all viral RNAsynthesis after a shiftfrom permissive to nonpermissiveconditions, and it
has been postulated that ts6 has a defect in theelongation activity of the replicase (2, 14, 26). cDNA clones of these mutants,aswellasof ts+ revertants, have been constructed, and restriction fragments from the mutant cDNA clones were used to replace the corresponding regions of a
full-length clone of Sindbis virus. These plasmids were tran-scribed in vitroby SP6 RNA polymeraseto produce infec-tioustranscripts, which were then testedfor ts phenotype. The viruses recovered from these transcripts have been
1194
on November 10, 2019 by guest
http://jvi.asm.org/
characterized.Together with sequence analysis of the cDNA
clones, these experiments have defined the mutations re-sponsible for the group F mutants.
MATERIALSAND METHODS
Virus stocks, growth, and purification. Mutant ts6,
ob-tained originally from B. Burge, was isolated from the HR strain of Sindbis virus following mutagenesis with
nitroso-guanidine(3). Mutants tsllO and sll8 were isolated froma
small-plaque strain ofHR Sindbis virus following
mutagen-esis with nitrous acid (30). Revertants were isolated by
plaquingmutantstocksat 30 and40°C. Asinglevirus plaque
of a ts+ revertant was picked from the 40°C plate, and the
virus was eluted into 1 ml of Eagle medium containing 5% fetal calf serum. This revertant plaque was used to infect primary chicken cells at 40°C, and the resulting stocks, following plaqueassay at 30and40°C,wereused asinfecting stocks forRNApreparation. Viruseswere grown inprimary
orsecondary chicken embryo fibroblastsandharvested10 to 20 h afterinfection, depending on the mutant. Mutants and revertants were grown at 30 or 40°C, respectively. Viral RNAwas isolated as described before (24).
cDNA cloning. cDNA
synthesis
forts6, tsllO,andtsll8,
as well as for their revertants, followed the procedure of Okayama and Berg (20). Aprimer complementary
to a sequence nearthe startcodonofcapsid protein (nucleotides
[nt]
7642 to 7661ofthegenomic RNA) wasmade;thisprimer
also contained the
recognition
site for XbaI restrictionendonuclease at its 5' end. This
primer
was used for first-strandsynthesis,andsecond-strandsynthesis
wasdonewith EscherichiacoliDNApolymerase I,E.coliRNaseH,
andE.coliDNA ligase.
Phosphorylated
EcoRIlinkerswereligated
to the double-stranded cDNA to facilitate laterdigestion
with XbaI (see below). The double-stranded cDNA wasdivided into two
portions
forcloning.
For the 5'library,
the AccI (nt83)-SacII (nt2771)fragment
of thecDNA,
which encodes nsPl and the N-terminal half ofnsP2,
was cloned into Kahn5. Kahn5 isaplasmid
containing
acDNA copyofthe 5' terminus ofthe Sindbis virus genome in Proteus
1,
a vector consisting ofthe replicon andP-lactamase
genes of pBR322 and an SP6 RNApolymerase
promoter(22;
H. V. Huang and C. M. Rice,unpublished).
The 3'library
wasconstructed by cloning the
BglII
(nt2268)-XbaI (nt7662)
fragment ofthecDNA,
encoding
the C-terminal half ofnsP2 and allofnsP3andnsP4, intoplasmid pMT21,
anampicillin-resistant
cloning
vectorderivedfrompBR322 (the
XbaI site is notpresent in this viral RNAbut was introducedby
theprimer, as notedabove).
Construction of
hybrid
genomes.Hybrid
genomes wereproduced by
replacing
restrictionfragments
inSindbisvirus clone TotollOl(22)
with thecorresponding
regions
fromcDNA clones derived from the mutantsor theirrevertants
(17). Details of restriction sites used are included in the
figure legends.
Full-length
hybrid
plasmids
that containedoneofthree
nonoverlapping
intervals(A,
B,
andC)
fromthe mutantssubstituted intoTotollOl werefirstconstructed forgross mapping. Plasmids with interval A contained the sequence of the mutant from the
SspI
(ntSO4)
to the ClaI (nt2713)site inTotollOl.(TotollO1
containsapproximately
13,638nucleotides;
numbering
begins
fromthe first nucleo-tideoftheSindbis virusgenome.)
Since theSspI
site is notunique, ashuttle vector,
irnsP12
(S.
A.ChervitzandC. M.Rice,
unpublished),
containing
the Sacl(ntl3552;
a site in the vectorupstreamofthe SP6promoter)
toEcoRV(nt2750)
region ofTotollOl
cloned inIAN7
(18)
wasdigested
withSspI (nt504)
and ClaI(nt2713)
andligated
with thecorre-sponding fragment
of 5' cDNA of the ts mutant. TheSacI-ClaI
fragment
oftheresulting
clone was then cloned into TotollOl which had been cut with Sacl and ClaI and treated with calf intestinal alkalinephosphatase.
Interval Bplasmids
contained the sequence ofthe mutant from ClaI(nt2713)
toSpeI
(nt5262)
inTotollO1 andwasconstructedby
replacing
thisfragment
inTotollO1 with thecorresponding
fragment
fromthe3' cDNAlibrary
ofthetsmutant. Intervalregion
Cplasmids
contained theSpeI
(nt5262)
to AatII(nt7999)
region
of the ts mutant in TotollOl and wasconstructed
by digesting
the 3' cDNAlibrary
of the ts mutantwithSpeI
(nt5262)
and BamHI(nt7334)
andcloning
into shuttle vector
TnsP34,
which is anAN7
derivativecontaining
the PvuII(nt5160)
to NcoI(nt8038)
fragment
of TotollOl(obtained
from H. V.Huang).
TheSpeI
toAatIIfragment
of theresulting
cloneswasthen usedtoreplace
thecorresponding fragment
ofTotollOl.For fine
mapping
of the Bregion, plasmids
werecon-structed that contained three
overlapping
subregions
re-ferred to as
Bi,
B2,
and B3.Subregions
Bi,
covering
theregion
ClaI(nt2713)-AvrII
(nt4280),
andB3,
covering
the AvrII(nt4280)-SpeI
(nt5262) region,
were cloneddirectly
into TotollOl.Subregion B2,
covering
the AvaI(nt3546)-BamHI
(nt4633)
region,
wasobtainedfrom theshuttlevectorKahn5B,
consisting
of the ClaI(nt2713)
to EcoRI(nt5869)
fragment
of Sindbis virus subcloned into Kahn5. Three clonescontaining
overlapping
subregions
Cl, C2,
and C3 were constructed for finemapping
of the Cregion
withlTnsP34.
Fragment
SpeI
(nt5262)-HindIII
(nt6267),
PstI(nt5824)-HpaI
(nt6919),
orNsiI(nt6461)-BamHI
(nt7334)
ofthetsmutantwascloned into
TrnsP34,
and theSpeI
(nt5262)-AatII(nt7999)
fragment
wasusedtoreplace
thecorrespond-ing
fragment
inTotollOl.
In vitro
transcription
and transfection. RNAtranscripts
weresynthesized
in vitro withSP6
RNApolymerase,
using
supercoiled
plasmid template
orplasmid
DNAdigested
withthe
appropriate
restrictionendonuclease fortheproduction
of runoff
transcripts,
as describedpreviously
(22).
Theresulting
transcripts
were transfected into confluentmono-layers
ofsecondary
chicken cells in 35-mm multiwell tissueculture
plates,
andthephenotype
oftherecoveredviruswas tested.Plaques
werequantitated by overlaying
themonolay-erswith 2mlof1% agarose in
Eagle
mediumcontaining
2%fetal calf serum, followed
by
incubation at 30 and40°C.
Plaques
were visualizedby
staining
with neutral red orcrystal
violet after incubation for36to40 hat40°C
orfor60to72 h at
300C.
Analysis
of viralRNAsynthesis.
Chickenembryo
fibroblastmonolayers (60-mm
plate)
wereinfected with Sindbis virus HRor tsmutantsorrecombinantvirusesrecoveredfrom thehybrid
cDNA clones at amultiplicity
of 50 PFU/cell inphosphate-buffered
saline(PBS)
(6)
containing
1%fetal calfserumand
dactinomycin
(ActD)
(1
,ug/ml)
andincubated at 30or40°C
for 1 h. At the end of theadsorption
period,
theinocula were
removed,
and the cells were washed withwarmed medium and incubated at 30 or
40°C
inEagle
mediumcontaining
3%fetal calfserumand ActD(1
,ug/ml).
For theshiftfrom 30to400C,
at3.5 hpostinfection
(p.i.)
oneset of
30°C
plates
was washed once with warmed mediumlacking
ActD;
warmed mediumcontaining
ActD(1
,g/ml)
wasthen
added,
and theplates
wereshiftedto40°C.
At 10 hp.i.
(30°C),
6 hp.i. (400C),
or8 hp.i.
(after
theshiftto40°C),
cells wereharvested.Theamountof viral RNA
present
wasquantitated
by
thecytoplasmic
dothybridization
method ofWhite and Bancrofton November 10, 2019 by guest
http://jvi.asm.org/
NONSTRUCTURAL PROTEINS
I _ I _ rl%et I n MA IL
ns P2
II
CIa Ava AvrII
2713 3546 4280
3,
I ns P3 I ns F4 _
IT I I I I I I
BamHISpe I Pst Hind IIINsi HpaI BamHI
4633 5262 5824 6267 6461 6919 7334
l Toto:ts6A, Toto:tsl1 OA
I Toto:ts6B, Toto:ts11 OB 7777I| Toto:ts6C, Toto:tsl 1OC
Toto:ts6Cl, Toto:tsl1OC1
Toto:ts6C2, Toto:tsl10C2 Toto:ts6C3, Toto:tsl10C3
Toto:tsl 18A Toto:tsl18B Toto:tsl18C Toto:tsl18B1 Toto:tsl 18B2 Toto:tsl 18 B3 Toto:tsl18BC Toto:tsl18BC1 Toto:tsl18 BC2 Toto:tsl18BC3
FIG. 1. Construction ofhybridgenomes.Aschematic of thenonstructural-protein-coding regionofSindbisvirus cDNAcloneTotollOl (22) is shown together with anumberof restriction sites and theirpositions (nt) inthe Sindbis virusgenome numbered fromthe 5' end accordingtoStraussetal.(33). Translated regionsareshownastheopenboxes,in which thenamesof thevariousproteinsareindicated. Nontranslated regionsareshownas asingle line. The hatched boxes show the location of restriction fragmentsincloneTotollOl thatwere replaced with thecorrespondingrestriction fragments from thetsmutantsortheirrevertants.Thenamesusedtorefertothesehybridclones
areshownattheright.
(37). EqualnumbersofcellswerewashedwithcoldPBSand then lysed with 1% Nonidet P-40in TE buffer(10mM Tris chloride[pH 7.5],0.1 mMEDTA).Nucleiwerepelleted,and the supernatantwastreated with 14.8%formaldehyde in 1x
SSC (150 mM sodium chloride, 15 mM sodium citrate) at 60°Cfor 15min.RNAsamples(10to20,ul)wereblottedonto the nitrocellulose membranes and probed with 32P-labeled minus-strand RNA from the region of thegenome (the 26S region) encoding the structural proteins. This probe was transcribedwith SP6 RNA polymerasefromacDNA clone
ofSindbisvirusthatcontained the structural protein region onlyinsertedinan invertedsense downstream froman SP6 promoter. Relative amounts of RNAwere determined bya beta-scanning counter. All results were corrected for the amount ofincorporation into mock-infected plates, which was between 0.5 and 1% of the incorporation into cells infectedwiththe parental strain of Sindbis virus.
Alternatively, RNA synthesis following the shift was
assayed byexamining the incorporation of [3H]uridine into infectedcells. Following infectionat30°C, cellswereshifted at3hp.i.to40°C andlabeled with [3H]uridine(20,uCi/ml) in thepresence ofActD from 3.5 to8 h p.i. Monolayers were
then washedwith PBS and lysed with 0.5 ml of 2% sodium dodecyl sulfate, 50 ,ul wasprecipitated with trichloroacetic
acid,and the incorporated radioactivity was quantitated by
liquid scintillation counting.
Complementation analysis. Complementation tests were
performed as described by Strauss et al. (30) but 35-mm multiwellplatesandamultiplicityofinfectionof 20 PFU/cell for each mutant were used. A complementation index was
calculated as theyieldfrom the mixed infection dividedby the sum of the yields following infection by each parent
alone. A complementation index was calculated separately foreach mutant intests in which the two mutants differed markedly in plaque size. The absolute magnitude of the complementation index isdependentontheyieldofparental viruses (i.e., the extent of leakage of the parents), as
complementationisalwaysinefficient(4, 5, 30), not exceed-ing1 to10%of the wild-typeyield, andin the case oftsll8
only one-way complementation couldbe demonstrated
be-causeofrelativelyhighyields of tsll8at40°C. RESULTS
Construction of recombinantplasmids. Inordertolocalize thetsmutations ofts6, tsllO,andtsl18,weconstructedand
tested a number ofrecombinant plasmids. The constructs
areillustrated inFig. 1. Ineachcasesmall(873 to 2,584 nt) 5'
Sspl 504
I
I
17,7777.77,771
V/77777TI
V,17122=1
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.75.552.79.414.2]Constructs tested for phenotype nonpermissivetemperature
Recombinant Fragment Phenotypeb Location of clonea replaced(nt) mutation(nt)
Toto:ts6A 504-2713 wt 5824-6267
Toto:ts6B 2713-5262 wt
Toto:ts6C 5262-7334 ts
Toto:ts6Cl 5262-6267 ts
Toto:ts6C2 5824-6919 ts
Toto:ts6C3 6461-7334 wt
Toto:tsllOA 504-2713 wt 6461-6919
Toto:tsllOB 2713-5262 wt
Toto:tsllOC 5262-7334 ts
Toto:tsllOCl 5262-6267 wt
Toto:tsllOC2 5824-6919 ts
Toto:tsllOC3 6461-7334 ts
Toto:tsll8A 504-2713 wt 2713-3546,
Toto:tsll8B 2713-5262 (ts) 5824-6267
Toto:tsll8Bl 2713-4280 (ts) Toto:tsll8B2 3546-4633 wt
Toto:tsll8B3 4280-5262 wt
Toto:tsll8C 5262-7334 wt
Toto:tsll8BC 2713-7334 ts
Toto:tsll8BC1 2713-6267 ts
Toto:tsll8BC2 2713-5262, ts 5824-6919
Toto:tsll8BC3 2713-5262, (ts) 6461-7334
aSeeFig. 1.
bRNAtranscriptsweretransfectedontocellsat30or40°C,and the plaque titerwasdeterminedasdescribedin Materials and Methods.wt,Wildtype.
(ts),Partiallyts,inthatplaquesizebutnotplaquenumber is reducedat40°C.
restriction fragments in the Sindbis virus cDNA clone TotollO1, from which infectious RNAcanbetranscribed in vitro with SP6 RNA polymerase (22), were replaced with
cDNA from a mutant or its revertant. In this figure, the restriction sites used to construct the hybrid genomes and
theirnumbering from the 5' end of the RNA (33) are also
shown. The genomic region encoding the nonstructural proteins was first divided into three large nonoverlapping regions, A, B, and C, forgrossmapping. For fine mapping, regionsBand Cwereeachsubdivided into three overlapping
regions (Bi, B2, and B3 andCl, C2, and C3). The 5' 444 nt and the 3' 265 nt of the coding region for the nonstructural proteins, which were not covered by hybrid genome con-structions, were sequenced in each case to ensure that no changes hadoccurred within these regions.
RNAwastranscribedin vitro from therecombinant
plas-midswithSP6 RNApolymeraseandtransfectedonto mono-layers ofchicken cells. Monolayers were incubated under agarose at 30 or 40°C to determine whether the virus recovered ineach casewas temperature sensitive.
Localization of the mutations ints6, tsllO, andtsll8. The
results obtainedwith the constructs tested are summarized
in Table 1. For ts6,ofthe three large interval replacement clones (Toto:ts6A, Toto:ts6B, and Toto:ts6C) tested, plas-midsToto:ts6A andToto:ts6Bgaverisetovirus that exhib-ited wild-type growth at the nonpermissive temperature, while ts virus was obtained from plasmid Toto:ts6C. This localized thetsmutation tothe interval nt5262to nt7334of the genome. Plasmids Toto:ts6Cl, Toto:ts6C2, and Toto: ts6C3, containing three smaller intervals in the C region, werethenconstructedandtested. Plasmids Toto:ts6Cl and Toto:ts6C2gaverisetotsvirus,whereasToto:ts6C3 didnot
TABLE 2. Plaquemorphology and RNA synthesis by tsll8at 40 and 30°C
Relative RNA Plaquesize Efficiency synthesisb
* ~~~~~~~~of
Virus plaquinga Shift
400C 300C (40OC/30°C) 400C 300C to 400C TotollOl Large Large 2.6 x 10-1 1.00 1.00 1.00 tsll8 Minute Small 5.0 x 10-5 0.07 0.70 0.32 Toto:tsll8Bl Minute Small 4.0 x 10-1 0.28 0.99 0.58 Toto:tsll8C Large Large 5.0 x 10-1 0.71 0.93 0.84 Toto:tsll8BC Minute Small 5.0 x 10-5 0.08 0.73 0.48 Toto:tsll8BC*c Minute Small 2.6 x 10-5 0.08 0.76 0.44 tsll8R Large Large 5.4 x 10-1 0.70 1.14 0.81
a Plaquetiter at400C dividedbythatat300C.
bViral RNAsynthesis (relativeto that byTotollOl)wasassayedby dot hybridization following infectionat 40 or 30°C orafterashiftto40°C following infectionat30°C,asdescribedinMaterialsandMethods.
'Toto:tsll8BC* containsthe Bfragmentfromtsll8and the Cfragment fromtsll8R.
(Table 1). Thus, ts6 has one or more mutations in the region between SpeI (nt5262) and HpaI (nt6919); if only a single mutation is involved,itmustlie in the region of overlap ofCl andC2, between PstI (nt5824) and HindIII (nt6267), which is
located neartheN terminus of nsP4.
FortsllO, virusrecovered from plasmid Toto:tsllOC was ts, whereas virus from Toto:tsllOA and Toto:tsllOB was not. Plasmids Toto:tsllOCl, Toto:tsllOC2, and Toto:
tsllOC3werethenconstructed and testedfor fine mappingof
the intervalregionC oftsllO. ts viruses were obtained from
recombinant plasmids Toto:tsllOC2 and Toto:tsllOC3,
whereas plasmidToto:tsllOCl gave rise to wild-type virus. Fromthisweconclude that tsllO has one or more mutations in theregion between nt5824and nt7334 of the genome and that ifa single mutation is involved, it must lie between nt6461and nt6919ofthe genome.This region is also in nsP4.
Mappingoftsll8 suggestedthatitwasadoublemutantin which one mutation was located in the nsP4 region, as was thecaseforts6andtsllO,and thesecondmutationwasina different region. Viruses from plasmids Toto:tsll8A and
Toto:tsll8C were apparently wild type, whereas that from
plasmid Toto:tsll8B was partially ts (Tables 1 and 2). This
partial temperature sensitivity manifested itselfasachange
inplaque sizefromsmallplaquesat 30°C tominute plaques
at40°C, althoughthenumberofplaquesat30and40°Cwere the same. RNA synthesis at 40°C was reduced relative to that in TotollOl virus (Table 2). When construct Toto:
tsll8BC was tested, the virus once
again
formed smallplaques at 30°C, whereas at 40°C minute plaques were
formed,with theplaquenumberbeingreducedby4ordersof
magnitude (Table 2), as wasthe casefor theparentaltsll8. Furthermore, RNA synthesis at40°C was reduced to very low levels(Table 2). Thus,weconcludethattsll8 isadouble mutant in which a mutation in the B
region
results information ofminuteplaquesat
40°C
butnochangeinplaque
number and a second mutation in the C region,although
havinglittle apparentphenotypeonits own, whencombined
with the change in the B region, results in red4ction of
plaquenumber(thusbeingscored as tsin
plaque
assays).
Fine mapping of the two tsll8 mutations was doneby
constructing Toto:tsll8Bl, -B2, and -B3
(Fig. 1),
and thechange
responsible
forformation of minuteplaques
at40°C
mapped toregion
Bi
(Table 1).Similarly,
thechange
in the C regionthat,
when combined with the Bregion change,
on November 10, 2019 by guest
http://jvi.asm.org/
H R
ts6
ts6R
5' NONSTRUCTURAL PROTEINS
-i
nsPlI
nsP2I
nsP3I
nsP4I I I I
1 2 3 4 5 6 7 (kb)
425 93 153 324
Val GlnGly Gly
I
I.I
I
U
2953
II I
A G G
6046 6226 6739
3'
Gly - Glu
A Gly G
Gly-_ Glu
ts1I 10 I
--A Gly ts11 0 R
---ValI-_ Ala t s118
C
ts1 18R
Gln-*_ Arg
G
Val GIn-_ Arg
G
U G
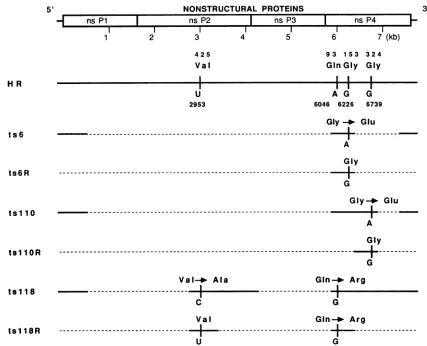
FIG. 2. Localization oftsmutations.Aschematic of thenonstructural-protein-coding regionof Sindbis virus is shown. Belowareshown sequencing schematics forHRSindbis virus (33),theparentalstrain from which thetsmutants wereisolated(34),and formutantsts6, tsllO, andtsll8andtheirrevertants. Sequencedregionsareshownassolid lines. Anychangefrom theHR sequenceonthe first line is indicated. Where nochangesareshown, the sequence is identicaltothatofHR.Nucleotidesarenumbered from the 5' end of theRNA;amino acids arenumbered fromthe Nterminus ofeachprotein.
resulted in reduction in plaque count was mapped to the
region ofoverlap in Cl and C2 between nt5824 and nt6267. Sequence analysis of ts6, tsllO, and tsll8 and their rever-tants. Inordertodefine thetslesionsofts6,tsllO,and tsll8,
regionsshownby themapping experimentstocontainthets lesions were sequenced by the chemical method ofMaxam and Gilbert (19) as modified by Smith and Calvo (29), with cDNAclones ofmutants aswellasof theirrevertants. The sequences obtained are shown inFig. 2.
ts6hadasingle base substitutionin theregion sequenced. Comparingthe ts6 sequence with that of its revertant, and
from the results in Table 1, we found that the mutation responsible for temperature sensitivity in ts6 was a change of GtoAatnt6226,which led to the replacement of Gly (GGG) atposition 153 ofnsP4by Glu (GAG). Inthe revertant, the
changed nucleotide reverted to the original nucleotide, re-storing the parental amino acid. tsllO also had only one
changethroughout the sequenced region. The change was G toA at nt6739, resulting in the change of Gly (GGG) to Glu
(GAG) atposition 324 of nsP4. In the tsllO revertant, this
nucleotidereverted to the original nucleotide.
tsll8 had asinglebasesubstitution in the region between nt2713 and nt3546. The change was U to C at nt2953,
resulting in the change from Val (GUG) to Ala (GCG) at
position 425 of nsP2. As discussed earlier, this change
resulted inreductioninplaque sizebutnotnumberat
40°C.
Inthe tsll8 revertant,this nucleotide revertedtothe
original
nucleotide.
Thesecondchangeints118wasfoundtobeachange ofA toGatposition 6046, resultinginaGlntoArgsubstitutionat
position93of nsP4. Thischange, combinedwith thechange
innsP2, rendered the virus tsin thatplaquenumberaswell asplaquesizewerereducedat40°C.Therevertantselected, tsll8R,retained thechangeinnsP4, which,asnoted
earlier,
had little apparent phenotype on its own. An additionalconstruct was made and tested in order to show that this
change was in fact
responsible
in part for temperaturesensitivity. The tsll8RC region was combined with Toto: tsll8BtoproduceToto:tsll8BC*, whichupon assaywasts,
formingtiny plaques at40°C in reduced numbers (Table 2).
Characterization ofthe tsmutations rescuedfromts6,tsllO, and tsll8. As a control to establish that the mutations
mapped and defined here were theonesresponsible for the
phenotypesdescribedpreviouslyfor these mutations, and in order to establish the phenotype of these mutations in a uniform background, recombinant viruses containing a de-fined region from each of the ts mutants in a TotollOl backgroundwerestudied.Monolayersweretransfected with dilutions of RNA transcribed from recombinant plasmids Toto:ts6C2, Toto:tsllOC2,
Toto:tsll8Bl,
Toto:tsll8C,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.94.521.71.417.2]TABLE 3. Efficiency ofplaqueformation and RNA synthesis by ts6 and tsllOat40and 30'C'
Titer Relative RNA
(PFU/ml) Efficiency synthesis
Virus ofplaquing Shift
400C 300C (40OC/30°C) 40°C 30'C to
400C
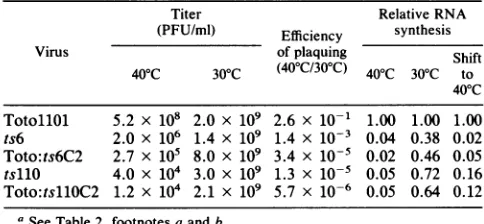
TotollOl 5.2 x 108 2.0 x 109 2.6 x 10-1 1.00 1.00 1.00 ts6 2.0 x 106 1.4 x 109 1.4 x 10-3 0.04 0.38 0.02 Toto:ts6C2 2.7 x 105 8.0 x 109 3.4 x 10-5 0.02 0.46 0.05
tsllO 4.0 x 104 3.0 x 109 1.3 x 10-5 0.05 0.72 0.16
Toto:tsllOC2 1.2 x 104 2.1 x 109 5.7 x 10-6 0.05 0.64 0.12
aSeeTable2, footnotesaand b.
Toto:tsll8BC, and Toto:tsll8BC* and incubated at 30and 40°C, withresults similartothose shown in Table 1.Asingle plaque of each viruswas isolated from the 30°Cplate and a
stockgrownat30°C, and these plaque-purified virus stocks
were characterized further. To start, titers of these
recom-binantviruses weredeterminedat 30and 40°C to ascertain the relative efficiency of plaque formation, and the results areshown in Tables 2 and 3. The virus stocks derived from
these infectious transcripts clearly showed thetemperature sensitivity of the parentalmutantin thecaseofts6andtsllO,
and theefficiencyofplaqueformationat40°C relativetothat at 30°C was low (Table 3) (the relatively high apparent reversionrateof ts6 is duetouseofastock thathadnotbeen recently plaque purified; during passage, revertants are am-plified inmost tsmutant stocks). However, asnotedearlier,
the virus recovered from Toto:tsll8Bl formed minute plaques at40°C, and the number of PFU was only slightly reduced (Table 2). The amino acid change of Valto Alain nsP2 isthusresponsible for the minute-plaque phenotype at
thenonpermissivetemperaturebutnotfor thereduced
num-berofplaques.ThemutationinnsP4 (constructToto:tsll8C) had littleeffectonitsown,butincombination with the nsP2 change(constructsToto:tsll8BC andToto:tsll8BC*) it led toapronounced decrease in theefficiency of plaque
forma-tionat40°C (Table 2).
To examine these viruses further, RNA synthesis was analyzed afterinfectionat30°C, afterinfection at40°C, and at40°Cfollowingashift from30°C.Theparentalmutantsts6, tsllO,and tsll8wereincluded,as wasvirusrecovered from
cloneTotollO1. TotalviralRNAsynthesiswasanalyzed by
thecytoplasmic hybridizationmethodofWhiteandBancroft (37) using 32P-labeled minus-strand RNA transcribed from the structural protein region as aprobe. Thevalues deter-mined, relative to those forTotollO1 virus, are shown in Tables 2 and 3. The synthesis of minus-strand RNA was significantlylessthan thatofplus-strand RNA,andtherefore the amount ofplus-strand RNA detected by hybridization wasassumedto bethetotal viralRNA. Followinginfection at 40°C, RNA synthesis by the viruses recovered from Toto:ts6C2 andToto:tsllOC2wasreducedtoalevel similar to that seen after infection by their respective parents.
However, the virus recoveredfromToto:tsll8Bl showed a higherlevelof RNAsynthesisthan theparentaltsll8 atthe nonpermissivetemperature(Table 2).Thisisconsistent with theobservationthat the mutation in nsP2givesrise tominute plaques at 40°C but does not reduce plaquing efficiency. RNAsynthesis by virus fromToto:tsll8C, containingonly thechangeinnsP4,wasonly slightlyreducedat40°Crelative to that by TotollO1 virus and was the same as that by tsll8R. However, RNA synthesis by the double mutants Toto:tsll8BC and Toto:tsll8BC* at 40°C was low and
TABLE 4. Complementation analysis' Complementation index
Virus Group A GroupB Group G GroupF (ts24) (tsll) (tsl8) ts6 tsllO tsll8
Toto:ts6C2 53 48 55 0.4 b
Toto:tsllOC2 67 42 42 3 0.3
-Toto:tsll8B1 12 7 7 4 0.2
Toto:tsll8BC 16 104 4 1 0.2
aComplementation indicesshown are theyieldfrom mixedly infected cells divided by the sum of the yields from singly infected cells. For tsll8 constructs,thecomplementation indicesare one way, asdescribedin Mate-rialsand Methods.
b-, Notdetermined.
identical to that of the parental tsll8, and thus the nsP4 mutationatposition93 markedly reduced RNA synthesisat thenonpermissive temperature whenit was combined with
the nsP2 change.
Themutantsand the constructs werealso tested for RNA
synthesis afterestablishing infectionat30°C andshifting to 40°C, for comparison with the results of Keranen and Kaariainen (14) and Sawicki et al. (26) for ts6 (see also
reference2). ts6 makes verylittle RNA after a shift to 40°C, asfoundby Sawickietal. (26), and thevirus recoveredfrom
Toto:ts6C2 exhibited the same phenotype (Table 3). tsllO also made little RNA after a shift to 40°C, as did Toto:
tsllOC2(Table 3). Fromamoredetailedstudyofthekinetics of cessation of RNA synthesis in ts6-infected cells after a
shift up (14, 26), and from studies of ts6 replication com-plexes in vitro (2), it was concluded that the elongation of
RNA chains, as opposed to initiation, was ts in ts6, and it waspostulatedthat the F groupfunction, here shown to lie in nsP4, definedthe RNA polymerase. Thetsll8 constructs
such as Toto:tsll8BC made significant amounts of RNA
after a shift, much more RNA than when infection and
incubationweredone at40°C continuously(Table2).Thus,
thereplicase complexes of tsll8, onceformedat 30°C,are
active afterashift to 40°C,in contrast to those specified by
tsllO and ts6.
It is worth noting that the assay used for these shift experiments examines total virus plus-strand RNA in the infectedcellsat8 hp.i. afterashiftat3.5 h. The results make clear that the RNA present at 3.5 h p.i. at 30°C does not
contribute
significantly
tothe RNApoolat8h,althoughthereplicase enzymes needed for an essentially full yield of virus RNA are present. As a control for these results, we
repeatedthe shift experiments and examined labeled RNA made between 3.5and 8 h p.i. in the presenceofActD and
[3H]uridine.
The results were similarto thosepresented
in Tables 2and 3.Complementation analysis of rescued mutants. We also
examined the ability ofthese virusesto
complement
repre-sentative ts mutants from the three othercomplementation
groups of RNA- mutants. Thecomplementation
indices shown in Table4demonstrate that theviruses derivedfromToto:ts6C2
andToto:tslIOC2complementedtheotherthreecomplementation
groups of RNA- mutants, as did theparental viruses, but did not
complement
theirparental
viruses or each other, in agreement withprevious
comple-mentation results (30).Interpretation of results with virus derived from Toto:
tsll8B1
and Toto:tsll8BC iscomplicated
because ofthe high titer of virusproduced
at40°C (the
virus from Toto:tsll8B1
was onlymarginally
ts, asnotedearlier,
and viruson November 10, 2019 by guest
http://jvi.asm.org/
1200 HAHN ET AL.
from Toto:tsll8BC also leaked at 40°C compared with the other mutants studied). In these cases thecomplementation indices shown are one-way indices. Because viruses from Toto:tsll8Bl and Toto:tsll8BC formed minute plaques, it waspossible to distinguish these plaques from thoseformed by the other ts mutants used. The complementation indices shown are the yield of large plaques in mixed infection divided by the yield of the large-plaque parents during single infection. Even so, complementation by Toto:tsll8Bl was marginal, and we cannot assign it to a complementation group, although it did seem tocomplement all of the other mutants tried. Analysis of the doublemutant Toto:tsll8BC showed that it did not complement its parental virus or ts6 andonly poorly complemented ts18 in groupG,but
comple-mented better the A mutant ts24 andquite well the Bmutant tsll. Thus, except for the marginal complementation of Toto:tsll8BC with tsl8, the resultswith therescued muta-tions are in good agreement withpreviouscomplementation
results (30).
DISCUSSION
Inthis report we have localized themutationsresponsible for the ts phenotype ofcomplementation group F mutants. The mutations in ts6 andtsllOwere mapped to nsP4. Each of them had a single base substitution, resulting in replace-ment of Gly by Glu at positions 153 and 324 of nsP4, respectively. Analysis of these mutations in a TotollO1
backgroundin avariety of ways,including RNA synthesisat
40°Candabilitytocomplement other RNA- mutants, dem-onstrated thatthese mutationsare infactthose responsible forthe tsphenotype and the mutantphenotypes previously
described in the literature. tsll8 turned out to bea double mutant. It had a defect in nsP2 (a Val to Ala change at
position425)whichonlypartiallydisabled itat40°C,
result-ing in production of small plaques and reduced RNA
syn-thesis, although the plaque numberwas unchanged. A sec-ond mutation in nsP4 combined with this change in nsP2 resulted in true temperature sensitivity, in that the plaque titer as wellas plaque size was reduced at 40°C and RNA synthesis after infection at 40°C was reduced to the level
characteristic of RNA- mutants. It is of interest that this
change in nsP4 alone had little apparent phenotype, but
becauseit is the one responsible for temperature sensitivity,
leadingtoreduced plaque titer in combination with the nsP2
change,thedouble mutant ts118 complements ts6 andtsllO
asagroup F mutant. Thus, mutations in nsP4 lead to group Fcomplementation behavior.
The experiments of Fuller and Marcus (8) in which the complementation groups were ordered by the relative rate of UVinactivationof their ability to complement gave an order for the complementation groups of NH2-G-A-B/F-COOH. Mutations in B and Fcould not be precisely localized, but both occurred downstream of group G and group A. Thus, the UV mapping data are consistent with our results that group F mutants contain defects innsP4. However, mapping of the other complementation groups of Sindbis virus ts RNA- mutants, currently being carried out, indicates that the Fuller and Marcus (8) order is not correct (Y. S. Hahn, unpublished observations).
Keranen and Kaariainen (14) and Sawicki et al. (26)
demonstrated that ts6 ceased genomic, subgenomic, and
minus-strand RNA synthesis upon ashift from the permis-sive to nonpermispermis-sive temperature and postulated that there wasa ts lesion in the elongationcomponent of the replicase. Recently, Barton et al. (2) extended such studies to in vitro
SIN SF RR ONN MID
SIN SF RR
ns P4 E:ts6
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~I
170NYPTVASYQITDEYDAYLDMVDGTVACLDTATFCPAKLRS ---SDS---R---C ---SES ---R---C ---V---SES---R---N-S----
---T---SES---R-A---S----E:tsll0
ns P4 t 336
PGTKHTEERPKVQVIQAAEPLATAYLCGIHRELVRRLTAV
---N--
---K--ONN
---N--MID
---N--ns P2 A:tsll8
SIN QKVNENPLYAITSEHVNVLLTRTEDRLVWKTLQGDPWIKQ SF ---PA---A---V RR
ONN
---PS---N---S---V ---P---GK-T----S---I
ns P4 R:tsll8 106
;* 106~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
SIN KVENQKAITTERLLSGLRLYNSATDQPEC YKITYPKPLY SF ----M--TVVD--T--A---TG-DVGRIPT-AVR--R-V-RR ----M--VIID--KD-A-T-LTEQSEKIPT-VSK--R-V-ONN ----M--TIIH--KE-C---LASETPRVPS-RVT--A-I-MID ----M--EVID--LG-AK-FVTP-TDCRY
VTHKH---M-FIG. 3. Aminoacidsequencesofproteins from fivealphaviruses in theregionnearthe group F mutations.Sequencedataarefrom the followingsources: Sindbis virus(SIN), Straussetal. (33); Semliki
Vorest
virus(SF), Takkinen (36);RossRivervirus (RR),Faragheret al.(7)andStraussetal.(31);O'Nyong-nyong virus (ONN), Straussetal.(31)andunpublished data; Middelburg virus (MID), Strausset al.(32).
studies of replication complexes isolated from ts6-infected cellsandconcludedthatsuch transcription complexeswere ts inelongation. In contrast,othermutantsexaminedretain activity at 40°C ifreplication complexes areformed first at
30°C (14). Theseresults, togetherwith theresultspresented here, suggest that nsP4 is an RNA polymerase or the
elongation component of the alphavirus replicase. In this regard, it is of considerable interest that nsP4 contains the
Gly-Asp-Aspsequenceandsurrounding hydrophobicamino acids that have been found to be present in the replicase
proteins of several other RNA viruses (13, 23).
Themutations in ts6,tsllO, and tsll8 are shown in Fig. 3, in which the sequences of up to five alphaviruses are
compared in the regions affected. It is readily evident that theglycines affected in nsP4 of ts6 and tsllO are conserved in allalphaviruses sequenced to date and that each is found within a long stretch of highly conserved amino acids. In view of this conservation in sequence, it is evident that
changes in the sequence, such as in ts6 and tsllO, might affect function. The nsP4 polypeptide is composed of 610 amino acids and is onaverage 71 to 74% conserved among
alphaviruseswhich havebeen examined (31). However, this
conservation is not uniform throughout the protein; amino acids1 to125from theNterminus(including the location of thechangeintsll8)are lesshighlyconserved, as are amino J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.322.560.75.399.2]acids 550 to 604 near the C terminus. The canonical se-quence Gly-Asp-Asp, flanked by
hydrophobic
aminoacids,
is found at residues 464 to 466. This sequence, which has been found in a number ofRNA-dependent RNAreplicases
(13; see alsodiscussioninreference23),is wellseparated (on
thelinear sequence) from either the ts6 or tsllO lesion. The change ofVal to Ala in nsP2 of tsll8 is found in a domain thatis wellconservedamongalphaviruses, although conservation is not absolute. In
particular,
the Val affected in ts118 is not totally conserved, being replaced by Thr inO'Nyong-nyong virus. The Gln to Arg
change
in nsP4 of ts118affectsanamino acidin adomainthatis notconserved (Fig. 3). The Gln isreplaced by Gly, Glu, Pro,
orAsp
indifferent alphaviruses. Thus, the failure of this
change
to have much effect by itself is perhaps notsurprising.
Thepronounced effect of this substitution in combination with
thechangein nsP2 suggests that nsP2 and nsP4 mayinteract
to form a functional complex, although other
explanations
for such a synergistic effectare
possible.
In Sindbisvirus, translation ofnsP4
requires
readthrough
ofanopal termination codonsothat nsP4is
underproduced
relativetonsPl, nsP2, and nsP3(11,
16, 32).
Theactive form ofnsP4 may be thepolypeptide
nsP34, which accumulatesduring
Sindbis virusinfection,
whereas little or no nsP4 is detected. Thefinding
that nsP4 may be the viral RNApolymerase, basedonresultswith
complementation
group F mutants and the presence of motifs within thisprotein
that are shared with other RNApolymerases
ofanimal viruses(13), is then reminiscent ofthe control ofvirus
replicases
inother systems in which
readthrough
ofaterminationcodonis required to produce the
polymerase.
In tobacco mosaic virus,readthrough ofanambercodonisrequired (9, 21),
andthe
readthrough
portion
oftheprotein
ishomologous
tothatofthe Sindbis virus
protein (12).
Similarly,
in theretrovi-ruses, translation of the reverse
transcriptase
requires
readthrough ofanambercodonorframeshifting
toeliminatean amber codon(25, 27,
28).
Wepresume thatregulation
of the amount of RNApolymerase
produced
isimportant
during
theviral lifecycle.
Ashasbeenpointed
out,however,
Semliki Forest virus and
O'Nyong-nyong
virus lack thistermination codon(31,
36),
andregulation
oftheactivity
of thepolymerase seems tobedifferent forthesealphaviruses.
ACKNOWLEDGMENTS
The expert technical assistance ofE. M. Lenches is
gratefully
acknowledged. We thank H. V. Huang and S. A. Chervitz for plasmid DNAs.This work was supported by Public Health Service grants Al 10793, Al 20612, and Al 24134 from the National Institutes of Health. C.M.R. is aPewscholarin the biomedical sciences.
LITERATURE CITED
1. Arias, C., J. R. Bell, E. M.Lenches, E. G. Strauss,and J.H. Strauss.1983.Sequenceanalysisoftwomutantsof Sindbis virus defective in theintracellular transport of their
glycoproteins.
J. Mol. Biol. 168:87-102.2. Barton, D.J., S. G.Sawicki,and D. L. Sawicki. 1988. Demon-stration in vitrooftemperature-sensitive
elongation
of RNA in Sindbisvirusmutantts6. J. Virol. 62:3597-3602.3. Burge, B. W., and E. R. Pfefferkorn. 1966. Isolation and characterization of conditional-lethal mutants of Sindbis virus. Virology 30:204-213.
4. Burge, B. W., andE. R. Pfefferkorn. 1966.
Complementation
betweentemperature-sensitivemutants of Sindbisvirus. Virol-ogy30:214-223.5. Burge, B. W., and E. R. Pfefferkorn. 1967.
Temperature-sensitive mutants of Sindbis virus: biochemical correlates of
complementation.
6. Dulbecco, R., and M.
Vogt.
1954. One stepgrowth
curve ofWestern
equine encephalomyelitis
virusonchickembryo
cells grown in vitroandanalysis
of virusyields
fromsingle
cells. J.Exp. Med.99:183-189.
7. Faragher, S. G., A.D. J. Meek, C. M. Rice, and L. Dalgarno. 1988. Genome sequences ofa mouse-avirulent and a
mouse-virulentstrain ofRossRivervirus.
Virology
163:509-526. 8. Fuller, F. J., and P. I. Marcus. 1980. Sindbis virus. I. Geneorder oftranslation in vivo.
Virology
107:441-451.9. Goelet, P.,G.P.Lomonssoff,P.J.G.Butler,M. E.Akam,M.J.
Gait,andJ.Karn.1982.Nucleotidesequenceof tobaccomosaic virusRNA. Proc. Natl. Acad. Sci. USA 79:5818-5822. 10. Hahn,C. S., E.G.Strauss, andJ. H.Strauss. 1985.
Sequence
analysis
of threeSindbis virusmutantstemperature-sensitive
in thecapsid
autoprotease. Proc. Natl. Acad. Sci. USA 82:4648-4652.11.
Hardy,
W.R.,andJ.H.Strauss.1988.Processing
thenonstruc-tural
polyproteins
of Sindbis virus:study
ofthekineticsinvivoby
using
monospecific
antibodies. J. Virol. 62:998-1007. 12. Haseloff, J.,P. Goelet, D. Zimmern,P.Ahlquist,
R.Dasgupta,
and P. Kaesberg. 1984.
Striking
similarities in amino acid sequence among nonstructuralproteins
encodedby
RNAvi-ruses that have dissimilar
genomic
organization.
Proc. Natl.Acad. Sci. USA81:4358-4362.
13. Kamer,G.,and P. Argos. 1984.
Primary
structuralcomparison
ofRNA-dependent polymerases
fromplant,
animal and bacte-rial viruses. Nucleic AcidsRes. 12:7269-7282.14. Keranen, S., and L. Kaariainen. 1979. Functional defects of RNA(-) ts mutants of Sindbis and Semliki Forestviruses. J. Virol.32:19-29.
15.
Lindqvist,
B.H.,J.DiSalvo,C. M.Rice,J.H.Strauss,and E. G. Strauss. 1986. Sindbis virus mutant ts2O ofcomplementation
group E contains a lesion in
glycoprotein
E2.Virology
151:10-20.
16. Lopez,S.,J.R.Bell,E.G.Strauss,andJ.H.Strauss. 1985. The nonstructural
proteins
of Sindbis virus as studied with anantibody
specific
for the C terminus of the nonstructuralreadthrough
polyprotein. Virology
141:235-247.17.
Lustig,
S., A. C. Jackson, C. S. Hahn, D. E. Griffin, E.G.Strauss, and J.H. Strauss. 1988. Molecular basis of Sindbis virusneurovirulence inmice. J. Virol. 62:2329-2336.
18. Lutz, C. T., W. M. Hollifield,B.Seed, J.M. Davie,and H. V.
Huang. 1987.
Syoinx
2A: animproved
Xphage
vectordesigned
for
screening
DNA librariesby
recombination in vivo. Proc. Natl. Acad. Sci. USA84:4379-4383.19. Maxam,A. M.,and W.Gilbert. 1980.
Sequencing
end-labeled DNAwithbase-specific
chemicalcleavages.
MethodsEnzymol.
65:499-560.20.
Okayama,
H., and P.Berg.
1982.High-efficiency cloning
offull-length
cDNA. Mol. Cell.Biol. 2:161-170.21. Pelham, H. R. B. 1978.
Leaky
UAG termination codon in tobacco virus RNA. Nature(London)
272:469-471.22. Rice, C. M., R. Levis, J. H.Strauss, andH. V. Huang. 1987. Production of infectious RNA
transcripts
from Sindbis virus cDNA clones:mapping
oflethalmutations,rescueofatemper-ature-sensitive marker, and in vitro
mutagenesis
to generate definedmutants.J. Virol.61:3809-3819.23. Rice,C.M.,E.G.Strauss,andJ.H.Strauss. 1986.Structure of the flavivirus genome, p. 35-90. In S.
Schlesinger
and M.Schlesinger
(ed.), TheTogaviridae
and Flaviviridae. PlenumPublishing Corp.,
NewYork.24. Rice, C. M., andJ. H. Strauss. 1981.
Synthesis,
cleavage
andsequenceanalysisof DNAcomplementarytothe 26Smessenger RNAofSindbisvirus. J. Mol. Biol. 150:313-340.
25. Rice,N.R.,R.M.
Stephens,
A.Burny,andR. V.Gilden. 1985. The gag and pol genes of bovine leukemia virus: nucleotide sequenceandanalysis. Virology
142:357-377.26. Sawicki,D. L.,S. G.Sawicki, S. Keranen,and L. Kaariainen. 1981.
Specific
Sindbis virus-coded function for minus-strand RNAsynthesis.
J. Virol. 39:348-358.27. Schwartz, D. E., R. Tizard, and W. Gilbert. 1983. Nucleotide sequenceofRous sarcomavirus. Cell32:853-869.
on November 10, 2019 by guest
http://jvi.asm.org/
28. Shinnick, T.M., R. A. Lerner, and J. G. Sutcliff. 1981. Nucle-otide sequence of Moloney murine leukaemia virus. Nature
(London)293:543-548.
29. Smith,D. R., and J. M. Calvo.1980.Nucleotidesequenceofthe
E. coligenecodingfor dihydrofolatereductase. Nucleic Acids
Res. 8:2255-2274.
30. Strauss, E. G., E.M. Lenches, andJ. H.Strauss. 1976. Mutants ofSindbis virus. I. Isolation and partial characterization of89
newtemperature-sensitive mutants. Virology74:154-168. 31. Strauss, E. G., R. Levinson, C. M. Rice, J. Dalrymple, and J. H.
Strauss. 1988. Nonstructural proteins nsP3 and nsP4 ofRoss Riverand O'Nyong-nyong viruses: sequence and comparison
with those of other alphaviruses. Virology 164:265-274. 32. Strauss, E. G., C. M. Rice, and J. H.Strauss. 1983.Sequence
coding forthe alphavirus nonstructural proteins is interrupted by an opal termination codon. Proc. Natl. Acad. Sci. USA 80:5271-5275.
33. Strauss, E. G., C. M. Rice, and J. H. Strauss. 1984. Complete nucleotide sequence of the genomic RNA of Sindbis virus.
Virology 133:92-110.
34. Strauss, E.G., and J. H. Strauss. 1980. Mutants ofalphaviruses: geneticsandphysiology,p.393-426.InR. W.Schlesinger(ed.),
Thetogaviruses. AcademicPress, NewYork.
35. Strauss, E. G., and J. H. Strauss. 1986. Structure andreplication of thealphavirus genome, p. 35-90. InS. Schlesingerand M.
Schlesinger (ed.), The Togaviridae and Flaviviridae. Plenum Publishing Corp., New York.
36. Takkinen, K. 1986. Complete nucleotidesequenceof the
non-structural proteingenesof Semliki Forestvirus. Nucleic Acids Res.14:5667-5682.
37. White, B. A., and F. C. Bancroft. 1982. Cytoplasmic dot hybridization. Simple analysis of relative mRNA levels in multiple small cell or tissue samples. J. Biol. Chem. 257: 8569-8572.