JOURNALOFVIROLOGY, June 1993,p. 3639-3643 0022-538X/93/063639-05$02.00/0
Copyright©1993, AmericanSocietyfor Microbiology
Association of
Human
Immunodeficiency
Virus
Type
1
Envelope Glycoprotein with Particles Depends
on
Interactions between the Third Variable and
Conserved
Regions of gpl20
RONALD L. WILLEY*ANDMALCOLM A. MARTIN
Laboratory of Molecular Microbiology, NationalInstituteof
Allergy
andInfectiousDiseases,Bethesda,
Maryland
20892Received 8January 1993/Accepted 2 March 1993
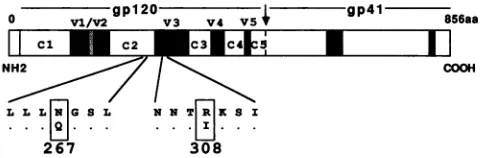
Many regions within theenvelope of humanimmunodeficiency virustype1 (HIV-1)that affect itsstructure
and function have been identified. We have
previously
reported that the interaction of the secondconserved (C2) and third variable (V3)regions ofgpl20influences theability
ofHIV-1toestablishaproductive infectionin susceptible cells. To better understand the basis for this interaction,wehave conducted structure-function analysesofenvelopeexpressedfrommolecularproviral clones ofHIV-1 containing defined mutations in C2 and
V3 that
individually
andin combinationdifferentially
affect envelope function. The substitution ofaglutaminefor an asparagine residue (Q-267) at a potential asparagine-linked glycosylation site in C2, which severely impairsvirus
infectivity,
reduces intracellular processing of gpl60intogpl20, the association ofgpl20withvirions, and the
ability
ofgpl20to bindtotheH1V-1cell surface receptorprotein, CD4. The change ofan argininetoanisoleucine codon inV3 (I-308),inthepresence of the Q-267 mutation,restoresvirusinfectivity
tonearwild-typelevelsby increasing theamountofgpl20 associated with virionsascompared with the Q-267
mutantbut doesnotcompensatefor theQ-267-induced processing defect. The I-308change in thecontextof
the
wild-type
HIV-1 hasnoaffectonprocessing, association,orCD4 binding. Theseresults indicate that theimpaired
infectivity
ofthe Q-267mutantvirus is duetoamarked reduction intheamountof viriongp120and suggest that theinteraction ofC2 and V3stabilizes the association of gp120withgp4l.The envelope of human immunodeficiency virus type 1 (HIV-1) consists of two glycoprotein subunits, gp120 and gp4l (2, 29). Both proteins are derived from a 160-kDa
precursor polyprotein (gpl60) which is cleaved during its intracellular transport through the endoplasmic reticulum
and Golgi complex (7, 28, 30). The mature gp120and gp4l cleavage products associate noncovalently (17) in virions,
and together they mediate viral attachment and entry into
CD4+ cells. Duringthepast severalyears, mutational anal-yses have been used to identify functional domains within bothgpl20andgp4l (3, 6, 8, 9, 11-13, 15-18, 20, 24,31, 33, 34). The cumulative results from these studies indicate that manyregionswithin bothgpl20 andgp4l influence specific envelopefunctions. Forexample, mutationsintroduced into four conservedregionsofgp120 impairitsbindingtothe cell surface viralreceptormolecule, CD4 (6, 24).The abilityof
gp4ltopromotefusion of the viral and host cellmembranes is reduced not only bymutations located within its amino-terminal and transmembrane regions (3, 12, 16) but alsoby changes in the third variable (V3) region ofgp120 (3, 13).
These resultsarenotparticularly surprising,sinceextensive disulfide bonding within gp120 (19) and the presence of oligomeric forms for both gp120 and gp4l (10, 25, 27) position nonadjacent regionsof theenvelope glycoproteinin closeproximity in HIV-1particles.
We have previously reported that the second conserved domain (C2) ofgp120 is critical for HIV-1 infectivity; the substitution ofaglutamineforanasparagineresidue(Q-267)
at apotential asparagine-linked glycosylationsite within C2 severely impairedtheabilityof HIV-1toestablisha
produc-*Correspondingauthor.
tive infectioninCD4+ T cells(33, 34). During these studies, however, spontaneous revertant viruses that were able to replicateto nearwild-typelevels emerged. Molecular
clon-ing and sequence analyses of the revertantproviral DNAs revealed that amino acid changes within the Cl and V3 regions ofgpl20 functionally compensated for the original glycosylation site mutation(33, 34). Since thisQ-267 muta-tiondidnotimpairthe abilityof solublegp120tobind CD4 (34) and theV3regionhas been reportedtoaffect a
postad-sorption step of the infectious cycle (3, 13), we suggested
that the structural interaction of Cl, C2, and V3 could influence the fusion of viral and host cell membranes(33).
To better understand the interaction of the C2 and V3
regions, we examined envelope structure and function by usingthewild-type proteinandQ-267,
Q-267/I-308,
andI-308 envelope mutants of HIV-1 (Fig. 1). Immunoblot analyseshad previously indicated that gp120 production is not af-fectedbytheQ-267substitution(34).We have also observed that theI-308change doesnotaffectprocessing and release ofgpl20 (data not shown). Since the
Q-267/I-308
revertant had not been examined in this regard (33), quantitative pulse-chase analyses of envelope processing were carried out toevaluate theamountofgp120 producedandreleased from cells overtime. ThepNL4-3, Q-267, andQ-267/I-308
proviral plasmidsweretransfected into HeLa cellsby using
calcium phosphate-precipitated DNA (31). At 18 to 20 h posttransfection,the cellswerepulse-labeledfor 30minwith [35S]methionine in methionine-free RPMI 1640 medium and then chased for 2, 4, and 6 h in complete RPMI medium
lacking radioactive label as previously described (31). Cell
pellets and the corresponding cell-free supernatants were
collectedimmediatelyafter thepulseand at each chasetime 3639
Vol. 67, No. 6
on November 9, 2019 by guest
http://jvi.asm.org/
3640 NOTES
gpl20
V5 gp4l 856aa0 v1/v2 V3 V4 v 5a
C C2cc4ci
IC
1 l+
N/H \ COOH
L L L N G B L N N T R x 8 I
...Q . . ...I ..
267 308
FIG. 1. Schematic representation of the HIV-1 gpl60envelope glycoprotein indicatingconserved(CltoC5)and variable(Vi toV5) regionswithingpl20.Boxesspecifyamino aciddifferences between thewild-typeandmutantproviralclones. aa,aminoacids.
point and then subjectedto detergent lysis (31). Thegpl60
andgpl20 envelope proteinswereimmunoprecipitatedwith
apolyclonalrabbit antiserum directed againstgpl20,
resus-pendedinsamplebuffer(2%sodium dodecylsulfate[SDS], 1%2-mercaptoethanol, 1%glycerol,65 mM
Tris-hydrochlo-ride [pH6.8]), separatedon8% SDS-acrylamide-AcrylAide gels, and visualized by fluorography (31). The amount of labeled gpl60 and gpl2O recovered from the cells and mediumatthe various timepointswasquantitatedbyusing aFujixBas 2000Bio-Image Analyzer (Fuji).
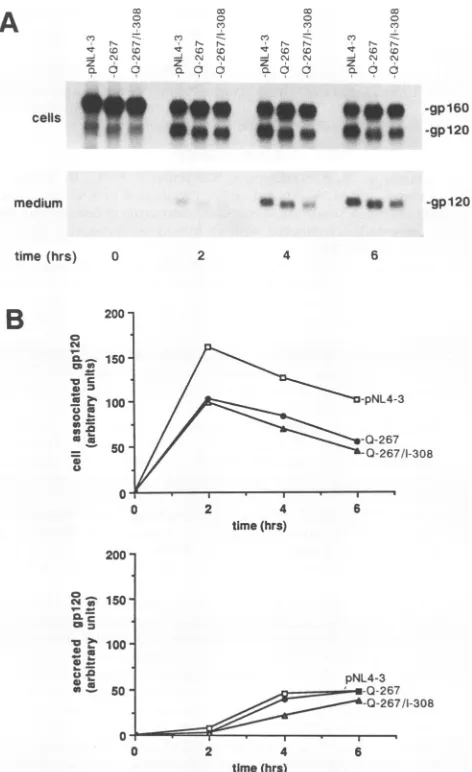
The results of the pulse-chase analysis (Fig. 2) indicated that although comparable amountsofgpl60wereproduced
followingthepulse-labeling,theamountofprocessedgpl20
presentafter 2 hof chasewasreducedinboth theQ-267-and
Q-267/I-308-transfected cells (36 and 38%, respectively) compared with cells transfected with pNL4-3 (wild type).
Thisfindingis consistent withourpreviousobservation that
amutationaffectingtheadjacent266 codonimpairs process-ing by preventprocess-ingtransportofgpl60outof theendoplasmic
reticulum (31). Themigration ofboth the gpl60and gpl20 mutant proteins in the Q-267- and Q-267/I-308-transfected
cells was slightly faster than that of the corresponding
pNL4-3wild-type species (Fig. 2), suggestingthattheQ-267
mutation eliminatedanN-linkedglycosylationsite. Itshould be noted that although the I-308 revertantchange restores infectivity (Table 1), this substitution failed to compensate for the Q-267-induced processing defect. Thus, reduced
gpl20 production alone in the Q-267 mutant does not
ac-countfor thenoninfectiousphenotypeof thisvirus(Table 1).
The diminished gpl60 processing in the Q-267-transfected
cells did not cause a reduction in the amount of gpl20
released into the medium. Infact, proportionately moreof theQ-267cell-associatedgpl20wassecretedcomparedwith the wild-type envelope, since identical amounts of both
gpl2Oswerepresentin themediumby6 h ofchase(Fig.2).
Thiswasnotobserved for cellstransfected withthe replica-tion-competent Q-267/I-308 mutant; somewhat less gpl20
wasreleasedduring thesametime period.
Since a larger proportion of the gpl20 produced in the
Q-267-transfected cells wasreleased into the medium
com-paredwiththe transfection withthewild-type provirus, the possibilitythatthismutationcausedgpl20todissociate from gp4lwasexamined. Such aputative association defect (15,
17) would be expected to reduce the amount of
virion-associated gpl20. To examine this possibility, HeLa cells were transfectedwith thepNL4-3, Q-267, Q-267/I-308, and I-308 proviral plasmidDNAsandthen labeled for 24 hwith [35S]methionine aspreviouslydescribed (31). Cell-free viri-onsgenerated duringthisperiodwerepelleted by
ultracen-trifugation (31), resuspended in complete RPMI 1640
[image:2.612.51.292.79.158.2]me-dium, and then assayed for reverse transcriptase activity
TABLE 1. Virusdesignationsandrelativeinfectivity
Virus Infectivitya Reference(s)
pNL4-3 (wildtype) +++ 1, 33, 34
Q-267 33,34
Q-267/I-308 ++ 33
1-308 +++ 33
a ++, a 2-day delay in infection kinetics compared with thepNL4-3and I-308viruses.
(34).
Comparableamountsofprogeny virions(as determined by reverse transcriptase activity) were subjected to deter-gent lysis, and the labeled viral proteinswereimmunopre-cipitated with HIV-1 antibodies present in a mixture of AIDS patient sera. The HIV-1proteins wereseparated by reducing SDS-polyacrylamide gel electrophoresis (PAGE) (10% poly-acrylamide gel) and visualized by fluorography. This analy-sis (Fig. 3) demonstrated that the Q-267 mutation markedly reduced the amount of particle-associated gpl20 compared with that found in wild-type pNL4-3 virions. The
I-308
change, in the context of the
Q-267/I-308
virions, effectedasubstantial increase in the amount of particle-associated gp120 compared with that present in the Q-267 virions, suggesting that the restored infectivity associated with the Q-267/I-308 revertant (Table 1) is due to this increase in virion
gpl20.
Itis alsointeresting
to note thatvirtually
nogp4l was detected in the Q-267 virions
(Fig.
3). While this could be due to the reduced processing of the Q-267gp160 (Fig. 2), it might also reflect a requirement for gp41 to associate with gp120 intracellularly in order to facilitate efficientincorporation
ofgp4l into particles.We
previously reported
that theQ-267
mutation alone did not affect CD4 binding, as judged from the use of solublegp120
in thebinding
assay(34). However,
it has recently been shown that thebinding
affinity
of virion-associatedgp120
for CD4 is less thanthat offreegp120
(22).
To examineCD4
binding
to virus-associatedgp120,
HeLa cells weretransfected
with thepNL4-3,
Q-267/I-308,
or I-308plasmid
DNA, and
radioactively
labeledvirions
werepelleted
from the medium aftersteady-state
labeling of the cells with[35S]methionine
asdescribed earlier. Theparticle-associated
gp120 of the Q-267mutantcouldnotbe
analyzed
because of theextremely
low levels ofvirion
gpl20(Fig. 3)
causedby
themutation. Each of the virion
pellets
wasresuspended
incomplete
RPMI 1640medium,
and100-,ul
aliquots
weretransferred into 1.5-ml screw-cap
Eppendorf
tubes(Sarstedt).
TheCD4-immunoglobulin
G(IgG)
form of CD4(4, 5) (generous
gift
ofGenentech, Inc.,
SouthSan
Fran-cisco,
Calif.)
wasaddedtothree tubesatfinal concentrations
of
500, 50, and 5 nM, while
afourth sample
served as an untreated control. All tubes wereincubated on ice for 1 h, and thevirionswerepelleted
at14,000
rpminarefrigerated
(4°C) Eppendorf microcentrifuge (Brinkman)
for 1 h(32);
supernatants were
collected
and transferred to new tubes.Detergent
lysates
of the viral pellets and corresponding supernatant samples were prepared by adding 1 ml of lysis buffer(50
mMTris-hydrochloride [pH
7.4], 300 mM NaCl,0.1%
TritonX-100)
to each sample and then subjecting thesamples
to vortexmixing.
Protein A-agarose beads(Be-thesda Research
Laboratories)
were then added to thelysates
ofthe viralpellets
toprecipitate thegpl20
which was associatedwithCD4-IgG,
since the IgGportion of CD4-IgG allows it to bindprotein
A(5).
After 1 h ofmixing
at 4°C, beadscontaining CD4-gpl2O complexes
werepelleted,
and J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.306.549.90.148.2]NOTES 3641
co x cc
0 0 0
N N N N N NK
C D v C-DC
-j N - N N N N
a C) - C- a a 0 c
cells 090
.0
000
*so
*a_s -gpl2O
gp120 0
p66- _ _ _ _
Ip55-
IV
_ Mrgp4 j ;
p39-B
200' Cco
150-100
co 6-=~ 50
-S s
p24
0 2 4 6
time(hrs)
a
0.-0
I..
200-
150-100
-50
-pNL4-3
:_-a-267/~
O2-0-267/i-308
0 2 4 6
time(hrs)
FIG. 2. Intracellularprocessingandrelease of thewild-typeand
mutant envelope glycoproteins. (A) HeLa cells were transfected
with the pNL4-3, Q-267, or Q-267/I-308 proviral plasmid DNA, pulse-labeled for 30minwith [35S]methionine,and chasedforupto
6 h in the absence of radioactive label. Polyclonal anti-gp120 antibodieswereusedtoimmunoprecipitatelabeledgp160andgp120 envelope glycoproteins present in the cells and released into the
mediumatthe indicated times. Theprecipitated glycoproteinswere
resolvedbySDS-PAGE and visualizedbyfluorography. The faint band around 120 kDaatthe 0 timepoint representsanendoplasmic
reticulum-derived envelope protein not producedfromproteolytic processingofgp160(31). (B) Quantitationof thegp120bands shown inpanelAasdescribed in thetext.
the remaining supernatants were transferred to protein A
beadspretreatedwith the AIDS patient serumforasecond round ofprecipitationtomeasureunbound
gpl20.
TheAIDS patientserumwasalso used toimmunoprecipitateanygpl20
which was present in the supernatant samples following
virus pelleting in the microcentrifuge to detect
gpl20
re-leased from thevirions as aresult of CD4binding (14, 21, 23).Allprecipitateswerewashed twice with thelysis buffer, resuspendedinsample buffer, and resolvedbySDS-PAGE. Labeled gpl20 was visualized by fluorography, and the
amountofgp120in the variousfractions(boundtoCD4,not
[image:3.612.70.306.77.463.2]bound,andreleased)wasquantitated by usingtheFujixBas
FIG. 3. Incorporation of envelope glycoproteins into virions. HIV-1 virions present in supernatants from metabolically labeled HeLa cells transfected with the wild-type pNL4-3 or the Q-267,
Q-267/I-308,or1-308mutantproviral DNAwerepelleted by ultra-centrifugation. The virus pellets were resuspended in RPMI 1640 medium, and lysates from comparable amounts of virions were
immunoprecipitated with protein A-agarose beads pretreated witha
mixture of AIDS patient sera. The precipitates were resolved by
SDS-PAGE and visualizedby fluorography. The virion-associated gp120 and gp4lenvelope glycoproteinsaswellasthe virus-encoded
p66 reversetranscriptase and p55, p39, and p24 Gag proteinsare
indicated. Theorigin of the protein bandsimmediately above p24 is unknown.
2000Bio-ImageAnalyzer. The relative binding of the differ-entgpl20swasdeterminedbycalculating thepercentgpl20
which had bound to CD4-IgG as follows: [(bound +
re-leased)/(bound
+ notbound + released)] x 100.AtthehighestconcentrationofCD4-IgG (500 nM), 12% of
the
Q-267/I-308
gpl20 was bound to CD4-IgG, comparedwith39 and42% for the pNL4-3 and I-308gpl20s,
respec-tively (Fig. 4 and Table 2). These values include the small amountof the virion-associated gp120 (6to7% of the total) whichwasreleased from the pNL4-3 and 1-308 virionsas a
result of the CD4-IgG binding. Only 2%of the
Q-267/1-308
gpl20bound toCD4 when the CD4-IgGconcentrationwas
reduced 10-foldto50 nM, while 12% of the pNL4-3 and9%
of the I-308 gpl20s were associated with CD4-IgG. The reducedCD4bindingof theQ-267/I-308gpl20aswellasthe
previously demonstrated reduced level of virion-associated
gpl20 (Fig. 3)mayboth contribute tothe delayedinfection kineticsdisplayedbythis virus(Table 1). AlthoughtheI-308 change alone did not affect gp120 binding to CD4, this substitutionmightstillplayarole in thecontextof the
Q-267
mutation,inview ofarecentreportindicatingthatchanges
inV3mayinfluence the interaction ofgp120with CD4(35).
Given the results of the current study, our previous conclusion that the
Q-267
glycosylation site mutation im-pairs envelopefunctionatapost-CD4bindingstep(34)mustbe amended since the N-to-Q substitution at position 267 affects multiple aspects of HIV-1 envelope structure and function. The reducedassociation of
gp120
with virionsas aconsequence of the
Q-267
mutation would appear to beprimarily responsiblefor theinabilityoftheQ-267virionsto
establishaproductive infection, inview of thehigherlevels
of gpl20 present on the infectious wild-type,
I-308,
andQ-2671/-308
virusparticles. It should be noted thatanaminoA
~00
-gpl60
-gp 120
0
4 N rN
l-
6D
z C
IC o a
medium
time(hrs) 0
0
2
VOL.67, 1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.379.498.81.263.2]3642 NOTES
viriongpl20 viriongp120 gpl20released boundtoCD4 notboundtoCD4 fromvirions
pNL4-3
Q-267/1.308
1-308
4W0
....
b,AN
5e
7tnMCD4-IgG ° ¢ ° o ga 0
0 o 0
FIG. 4. Binding of CD4tovirion-associatedgpl20. HeLa cells transfected with thewild-type pNL4-3orthemutantQ-267/I-308or
I-308 proviral plasmid DNA were metabolically labeled with [35S]methionine for 24 h. Labeled virions werepelleted from the medium by ultracentrifugation and resuspended in RPMI 1640 medium, and CD4-IgGwasaddedto100-,ul aliquots ofeach virus suspensionattheindicated concentrations. Afterincubationat4°C for 1 h, virions were pelleted in an Eppendorf microcentrifuge, supernatantswereremoved, anddetergentlysates ofthepellet and supernatant sampleswere prepared. CD4-gpl20 complexes in the pellet samples (virion gpl20 bound to CD4) were recovered by adding proteinA-agarose beadswhich bind tothe IgGportion of CD4-IgG. Viriongp120notboundtoCD4wasimmunoprecipitated from thepelletlysatesbyconductingasecond round ofprecipitation with proteinA-agarose beads pretreated with a mixture of AIDS patientsera.gp120released from virionsas aresult ofCD4binding
was immunoprecipitated from the supernatant samples with the AIDSpatientserum.Allproteinswereresolvedby SDS-PAGE and visualized by fluorography. Quantitation of the data is shown in Table2.
acid
substitution
in Cl(N-128), capable
ofrestoring
infec-tivity
totheQ-267
envelope
mutantof HIV-1(34),
increases the level of virion-associatedgp120
(unpublished data).
Previous reports
have also shown that mutations
inCl
and C2disrupt
the association ofgp120
withgp41
(15, 17).
Our studies suggest that theinteraction
of theCl, C2,
and V3regions
ofgp120
stabilizes thenoncovalently linked gp120
and
gp41
(17). However, it
mustbe remembered that subse-quent to thebinding
ofvirions
to the CD4 receptor,gpl20
undergoes
conformationalchanges (26)
that may berequired
for its
dissociation
fromgp4l, thereby allowing
the lattertomediate
the fusion of viral and cellular membranes. Thus, the interaction ofCl, C2,
andV3 may be bothcomplex
anddelicately balanced. Following cleavage
from gp160, thenoncovalent association
linking gp120 and
gp41 mustbe
sufficiently stable to maintain structural integrity of this
TABLE 2. Amountof thewild-typeand mutantgpl20sbound to CD4asdetermined fromquantitation ofthegp120bandsshown
inFig. 4
%Bound' atCD4-IgGconcn(nM) of: Virus
500 50 5
pNL4-3 39 12 3
Q-267/I-308 12 2 0
I-308 42 9 1
aExpressed as
gpl20
bound +gpl20
released/totalgpl20
(bound + notbound+released).
complexyetcommensurately unstabletopermit its dissoci-ation following the binding of virion-associated
gp120
toCD4.
Wethank DimiterDimitrov forproviding the conditionsusedfor pelleting HIV-1 particles in the Eppendorf microcentrifuge and KlausStrebel for criticalreview of themanuscript.
REFERENCES
1. Adachi, A., H.E.Gendelman, S. Koenig, T.Folks, R.Willey,A. Rabson, and M. A. Martin.1986.Production ofacquired immu-nodeficiency syndrome-associatedretrovirus in human and non-human cellstransfected with aninfectious molecular clone.J. Virol. 59:284-291.
2. Allan, J. S., J. E. Colligan, F. Barin, M. F. McLane, J. G. Sodroski, C. A.Rosen, W. A. Haseltine, T. H.Lee,and M.Essex. 1985. Major glycoprotein antigens that induce antibodies in AIDS patients are encoded by HTLV-III. Science 228:1091-1094.
3. Bergeron, L., N. Sullivan, andJ. Sodroski. 1992. Target cell-specific determinants of membrane fusion within the human immunodeficiency virus type gpl20 third variableregion and gp4l amino terminus. J.Virol. 66:2389-2397.
4. Byrn, R.A., J.Mordenti, C. Lucas,D. Smith, S.A.Marsters, J. S. Johnson, P. Cossum, S. M. Chamow, F. M. Wurm, T. Gregory,J. E. Groopman, and D. J. Capon. 1990. Biological properties ofa CD4 immunoadhesion. Nature (London) 344: 667-670.
5. Capon,D.J., S. M.Chamow, J. Mordenti, S.A. Marsters,T. Gregory,H.Mitsuya, R. A. Byrn,C.Lucas, F. M.Wurm,J.E. Groopman, S.Broder, and D. H.Smith. 1989. DesigningCD4 immunoadhesions forAIDStherapy.Nature(London) 337:525-531.
6. Cordonnier, A.,Y.Riviere, L.Montagnier, and M. Emerman. 1989. Effects of mutations in hyperconserved regions of the extracellular glycoprotein of human immunodeficiency virus type1 on receptorbinding. J. Virol. 63:4464-4468.
7. Dewar, R. L., M. B.Vasudevachari,V. Natarajan,and N. P. Salzman.1989.Biosynthesis and processing of human immuno-deficiency virustype 1envelope glycoproteins: effects of
mon-ensinonglycosylation andtransport.J. Virol.63:2452-2456. 8. Dubay, J. W., S. J.Roberts, B. Brody, and E. Hunter. 1992.
Mutations in the leucinezipper of the humanimmunodeficiency virus type 1 transmembrane glycoprotein affect fusion and infectivity.J. Virol. 66:4748-4756.
9. Dubay, J. W., S. J. Roberts, B. H. Hahn, and E. Hunter. 1992. Truncation ofthehumanimmunodeficiencyvirus type 1 trans-membraneglycoprotein cytoplasmic domain blocks infectivity. J. Virol. 66:6616-6625.
10. Earl, P. L., R. W. Doms, and B. Moss. 1990. Oligomeric structureof the humanimmunodeficiency virustype1envelope glycoprotein. Proc.NatI.Acad.Sci. USA 87:648-652. 11. Freed, E. O., D. J. Myers, and R. Risser. 1989. Mutational
analysis of the cleavage sequence of the human immunodefi-ciency virustype 1envelope glycoprotein precursorgpl60. J. Virol. 63:4670-4675.
12. Freed, E.O., D. J. Myers, and R. Risser. 1990.Characterization of the fusion domain of the humanimmunodeficiencyvirus type 1 envelope glycoprotein gp4l. Proc. Natl. Acad. Sci. USA 87:4650-4654.
13. Freed, E. O., D. J. Myers, and R Risser. 1991. Identification of the principal neutralizing determinant of human immunodefi-ciencyvirus type 1 as afusiondomain. J. Virol.65:190-194. 14. Hart, T. K., R. Kirsh, H.Ellens,R. W. Sweet,D. M. Lambert,
S. R. Pettaway, J. Leary, and P. Bugelski. 1991. CD4-HIV-1 interactions:binding of soluble CD4 (sT4) to HIV-1 and HIV-1 infectedcells inducessheddingof envelope gpl20.Proc. Natl. Acad. Sci. USA88:2189-2193.
15. Helseth, E., U. Olshevsky, C. Furman, and J. Sodroski. 1991. Human immunodeficiency virustype 1gpl20 envelope glyco-protein regions importantforassociation with the gp4l trans-membraneglycoprotein.J.Virol. 65:2119-2123.
16. Helseth, E., U.Olshevsky,D.Gabuzda, B. Ardman, W. Hasel-J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
NOTES 3643
tine, and J. Sodroski. 1990. Changes in the transmembrane regionofthe humanimmunodeficiencyvirus type 1gp4l enve-lope glycoprotein affect membrane fusion. J. Virol. 64:6314-6318.
17. Kowalski, M., J. Potz, L. Basiripour, T. Dorfman, W. C. Goh, E. Terwilliger, A. Dayton, C. Rosen, W. Haseltine, and J. Sodroski. 1987.Functional regions of the envelope glycoprotein of human immunodeficiencyvirus type 1.Science 237:1351-1355. 18. Lasky, L. A., G. M. Nakamura, D. H. Smith, C. Fennie, C.
Shimasaki,E. Patzer, P. Berman, T.Gregory, and D. J. Capon. 1987. Delineation ofaregion of the human immunodeficiency virustype1gpl20 glycoprotein critical for interactionwith the CD4 receptor. Cell 50:975-985.
19. Leonard, C., M. Spellman, L. Riddle, R. Harris, J. Thomas, and T.Gregory. 1990.Assignment of intrachain disulfide bonds and characterization of potential glycosylation sites of the type 1 human immunodeficiencyvirusenvelope glycoprotein (gpl20) expressed in Chinese hamster ovary cells. J. Biol. Chem. 265:10373-10382.
20. McCune, J. M., L. B. Rabin, M. B. Fienberg, M. Lieberman, J. C.Kosek, G. R. Reyes, and I. L. Weissman. 1988. Endopro-teolytic cleavage of gpl60 is required for the activation of humanimmunodeficiencyvirus. Cell53:55-67.
21. McKeating,J. A., A. McKnight, and J. P. Moore. 1991. Differ-ential loss of envelope glycoprotein gpl20 from virions of humanimmunodeficiency virustype 1isolates:effecton infec-tivity and neutralization. J.Virol. 65:852-860.
22. Moore, J. P., J. A. McKeating, Y. Huang, A. Ashkenazi, and D. D. Ho. 1992. Virions of primary humanimmunodeficiency virus type 1isolatesresistanttosolubleCD4 (sCD4) neutraliza-tion differ in sCD4 binding and glycoprotein retention from sCD4-sensitiveisolates.J.Virol. 66:235-243.
23. Moore, J. P., J. A. McKeating, R. W. Weiss, and Q. J.Sattentau. 1990. Dissociation of gpl20 from HIV-1 virions induced by soluble CD4. Science 250:1139-1142.
24. Oldshevsky,U., E. Helseth,C.Furman, J. Li, W. Haseltine, and J.Sodroski. 1990. Identification of individual human immuno-deficiencyvirus type 1 gpl20 amino acids important for CD4 receptorbinding. J.Virol. 64:5701-5707.
25. Pinter,A., W. J. Honnen, S. A.Tilley,C.Bona,H.Zaghouani,
M. K.Gomy,andS. Zolla-Pazner. 1989.Oligomericstructure of gp4l,the transmembrane protein of humanimmunodeficiency virustype1. J.Virol. 63:2674-2679.
26. Sattentau,Q. J., and J. P. Moore. 1991. Conformationalchanges in the human immunodeficiencyvirus envelope glycoproteins bysoluble CD4binding.J. Exp.Med. 174:407-415.
27. Schwaller, M.,G.E. Smith, J. J.Skehel,and D. C.Wiley. 1989. Studieswithcrosslinkingreagents on theoligomericstructure of the envglycoproteinofHIV. Virology 172:367-369.
28. Stein,B.S.,andE. G. Engleman. 1990.Intracellular processing of the gpl60 HIV-1 envelope precursor. J. Biol. Chem. 265: 2640-2649.
29. Veronese, F. D., A. L. DeVico,T. D. Copeland, S.Oroszlan, R. C.Gallo,and M. G.Sarngadharan.1985.Characterizationof gp4l as the transmembrane protein coded by the HTLV-III/ LAVenvelopegene. Science 229:1402-1405.
30. Willey,R.L., J.S.Bonifacino, B.J. Potts,M.A. Martin,and R. D.KIausner. 1988.Biosynthesis, cleavage, and degradation of the human immunodeficiency virus type 1envelope glyco-proteingpl60.Proc. Natl. Acad. Sci. USA 85:9580-9584. 31. Willey, R.L.,T.Klimkait,D.M.Frucht,J. S.Bonifacino, and
M. A.Martin. 1991. Mutations within the human immunodefi-ciencyvirustype1gp160 envelope glycoproteinalter its intra-cellular transportandprocessing. Virology184:319-329. 32. Willey,R.L., M. A. Martin, and K. Peden.Unpublisheddata. 33. Willey, R.L.,E. K.Ross,A.J.Buckler-White,T.S.Theodore, and M.A.Martin.1989. Functional interaction ofconstantand variable domains of human immunodeficiency virus type 1 gpl20. J. Virol. 63:3595-3600.
34. Willey, R. L., D. H.Smith,L.A.Lasky, T. S.Theodore,P. L. Earl, B. Moss,D.J. Capon,andM. A. Martin. 1988. In vitro mutagenesisidentifiesaregion withintheenvelopegeneofthe humanimmunodeficiency virus that is critical for infectivity.J. Virol.62:139-147.
35. Wyatt, R., M.Thali, S.Tilley,A.Pinter,M.Posner,D.Ho, J. Robinson, and J. Sodroski. 1992. Relationship of the human immunodeficiency virus type 1 gpl20 third variableloop to a component of the CD4 binding site in the fourth conserved region.J.Virol. 66:6997-7004.
VOL. 67,1993