AND ITS APPLI CATI ON TO
DI RECTI ONAL SELECTI VI TY IN THE RETINA
BY
ROMAN RICHARD POZNANSKI, M. S c . (
M
onash )
(
jBfvtran ‘vj
A THESIS SUBMITTED FOR THE DEGREE OF
D
octor
of Ph i l os o p h y
of
The Au s t r a l i a n
Na t i o n a l
U
n i v e r s i t y
STATEMENT
i
ACKNOWLEDGEMENTS
ii
PREFACE
iii
ABSTRACT
v
NOMENCLATURE
viii
LIST OF FIGURE LEGENDS
xiv
CHAPTER 1
MEMBRANE VOLTAGE
CHANGES
IN PASSIVE
DENDRITIC
TREES: A GENERAL
TAPERING EQUIVALENT
CABLE
MODEL
1.1
Inroduction
1
1.2
Derivation of the governing equation
6
1.3
Condition for reduction to a tapering
9
equivalent cable
1.4
Condition for reduction to a tapering
16
equivalent cable with electrical and
geometrical nonuniformities
TAPERING EQUIVALENT CABLE MODEL
2 . 1 G r e e n ’s function for a tapering equivalent 28
cable with sealed ends
2 . 2 Time course to a 6-current pulse 34
2.3 Asymptotic solutions at relatively short 41 and long time periods
2.4 Time course to a long-lasting current 47 pulse
2.5 Transient voltage decrement from a clamped 56 soma and steady-state somatopetal voltage
decay
2 . 6 Estimation of the membrane time constant 61
(t ) and electrotonic length parameter (L) m
CHAPTER 3 ANALYSIS OF A POSTSYNAPTIC SCHEME BASED ON A SINUSOIDALLY TAPERING EQUIVALENT CABLE MODEL
3.1 Introduction 70
3.2 Synaptic conductance change and 71
Volterra expansion
3.4
Interaction between excitation and
84
shunting inhibition
3.5
Application of the postsynaptic scheme to
89
directional selectivity in the retina
CHAPTER 4
MODELLING THE ELECTROTONIC STRUCTURE OF A
STARBURST AMACRINE CELL IN THE RABBIT
RETINA
4.1
Introduction
95
4.2
Experimental methods
99
4.3
Results and discussion
106
4.4
Directional selectivity in the retina:
111
a presynaptic cotransmission model
BIBLIOGRAPHY
119
APPENDIX I
129
This
thesis
contains
no
material
which
has
been
accepted for the award of any other degree or diploma
in any university, and to the best of my knowledge and
belief,
contains
no
material
previously
published
or
written by another person, except when due reference is
made in the t ext.
Roman R Poznanski
Canberra, Australia
A C K N O W L E D G E M E N T S
I am extremely indebted to my supervisor Professor
W.R. Levick,FRS, for his guidance and assistance throughout
the duration of this work.
I am additionally indebted to Dr I.G. Morgan for
his role as Chair of the Supervisory Committee and his
constant help and guidance especially in the final stages
of my research
I am also very grateful to Professor G.A. Horridge,
FRS,
and Professor A.W. Snyder,FRS, for their encouragement
which they have given me over the
last three years.
I would
like to thank Professor S.J.
Redman who
provided the initial idea behind the tapering equivalent
cable model developed in this thesis, and who acted as
an advisor during the early stages of my research.
I would also like to thank Dr D. Osorio for his advise
on the photographic techniques used in this thesis and
to Dr D.I. Vaney (University of Queensland) for kindly
providing the histological material used in this thesis.
Financial support has been provided by an Australian
P R E F A C E
My candidature as a Ph.D. student formally commenced
in February 1988. Since this time presentations have been
made at the Australian Neuroscience Society meetings in
Brisbane
(Australia)
and
Dunedin
(New
Zealand).
In
conjunction with these presentations the following papers
have also been published:
1. Poznanski, R.R. (1988) Membrane voltage changes
in
passive
dendritic
trees:
a
tapering
equivalent
cylinder
model.
IMA
Journal
of
Mathematics Applied in Medicine and Biology.
5
,113-145.
2. Poznanski, R.R. (1990) Analysis of a postsynaptic
scheme based on a tapering equivalent cable
model.
IMA
Journal
of
Mathematics
Applied
in Medicine and Biology.
7
,175-197.
3.
Poznanski,
R.R.
(1991)
A generalized
tapering
equivalent cable model for dendritic neurons.
accepted for publication:
4. Poznanski, R.R. (1992) Modelling the electrotonic
structure of starburst amacrine cells in the
rabbit
retina:
a
functional
interpretation
of
dendritic
morphology.
Bulletin
of
Mathematical Biology.
The content of this thesis has been contructed from the
above mentioned material, with the policy
that the thesis
was
written
first,
and
thereby
avoiding
reference
to
the above papers in the text of this thesis.
Roman R. Poznanski
Canberra (Australia)
A B S T R A C T
A new mathematical model has been developed which
collapses a dendritic neuron of complex geometry into
a single electrotonically tapering equivalent cable. The
modified
cable
equation
governing
the
transient
distribution
of
subthreshold
membrane
potential
in
a
branching
tree
was
transformed,
becoming
amenable
to
analytic
solution.
This
transformation
resulted
in
a
Riccati
differential
equation
whose
six
solutions
(expressed
in
terms of elementary
functions)
express
the amount and degree of taper found in the equivalent
cable model.
The Laplace
transform method
was used
to obtain
analytic expressions for the Green’s function corresponding
to an
instantaneous pulse
of current
injected
at a single
point
along a tapering
equivalent
cable with
sealed
ends .
The time course
of
the voltage in
response to
an arbitrary input was computed using the Green’s function
in a convolution integral. Infinitesimally brief (Dirac
6-function) pulses and
step pulses were considered. It
has
been demonstrated that
inputs located on a
tapering
equivalent
cable
are
more
effective
at
the
cylinder. Asymptotic solutions were also derived to enable
the voltage response behaviour over both relatively short
and
long time periods to be analysed.
Semilogarithmic
plots of these solutions provided a basis for estimating
the membrane time constant x from experimental transients.
m
A formula was derived which showed that tapering tends
to
increase
the
estimate
of
the
electrotonic
length
parameter L. Transient voltage decrement from a clamped
soma revealed that tapering tends to reduce the error
associated
with
inadequate
clamping
of
the
dendritic
membrane.
An analytic solution of the modified cable equation
with reversal potentials was used to explore nonlinear
synaptic effects in passive dendritic trees of arbitrary
geometry. To illustrate the theory, a sinusoidal equivalent
cable representing the dendritic arbor of
a cat retinal
6-ganglion cell was used to show that shunting inhibition
can be effective when located off the direct-path between
the excitation and the soma. In particular it was shown
that a peripherally placed excitatory input juxtaposed
with
a
shunting
inhibitory
input
could
produce
a
voltage-peak minimum at the soma in order to suppress
the initiation of an action potential at the axon hillock.
Such a postsynaptic scheme was shown to be applicable to
F i n a l l y , a d e t a i l e d m o r p h o m e t r i c a n a l y s i s of a
L u c i f e r y e l l o w - f i l l e d Cb a m a c r i n e c e l l u n d e r t a k e n to
p r o v i d e r a w d a t a fo r the c o n s t r u c t i o n of a t a p e r i n g c a b l e
m o d e l w i t h b o t h e l e c t r i c a l a n d g e o m e t r i c a l n o n u n i f o r m i t i e s
w a s e m p l o y e d to d e t e r m i n e w h e t h e r d i s t a l i n p u t - o u t p u t
r e g i o n s of d e n d r i t e s w e r e e l e c t r i c a l l y i s o l a t e d f r o m t he
s o m a a n d e a c h o t h e r . C a l c u l a t i o n s of s t e a d y - s t a t e
e l e c t r o t o n i c c u r r e n t s p r e a d s h o w e d t h a t d e n d r i t e s of
s t a r b u r s t a m a c r i n e c e l l s p r o c e s s i n f o r m a t i o n
e l e c t r o t o n i c a l l y w i t h a b i a s t o w a r d s the s o m a t o f u g a l
d i r e c t i o n a n d f or a p a r t i c u l a r c h o i c e of m e m b r a n e
r e s i s t a n c e R v a l u e s t he v o l t a g e a t t e n u a t i o n in the m
s o m a t o p e t a l d i r e c t i o n r e v e a l e d t h a t the a c t i o n of t h e s e
d e n d r i t e s c o u l d be c o n f i n e d l o c a l l y . A f u n c t i o n a l
i n t e r p r e t a t i o n of t h e s e r e s u l t s f a v o u r s a p r e s y n a p t i c
v e r s i o n of t h e c o t r a n s m i s s i o n m o d e l w h i c h a t t e m p t s to
e x p l a i n h o w t he n e u r a l n e t w o r k of s t a r b u r s t a m a c r i n e c e l l s
c o u l d a c c o u n t f or d i r e c t i o n a l l y s e l e c t i v e r e s p o n s e s
N O M E N C L A T U R E
The following symbols are used throughout the whole
thesis, unless stated otherwise.
V
=
V -E
m
r
Electrotonic
potential,
as
deviation
of
membrane potential from resting value (mV).
V .
reV =
E .
-J
J
-E
Synaptic reversal potential, as deviation
of the synaptic equilibrium potential from
resting value (mV).
G
r
Resting membrane conductance per unit area
(fi_ 1cm"2 ) .
R.
l
Resistivity of the intracellular material
( ftcm ).
R
=G~ 1
m
r
Resistance across a unit area of membrane
(ft cm2 ) .
C
m
Capacitance per unit area of membrane
( F cm 22 ) .
I
I
m Total membrane current per unit area
(A cm ^ ) .
t Time (m s ).
T =C /G Resting membrane time constant (ms),
m m r v '
T=t/x Dimensionless time variable,
m
G. Input conductance (fi ).
d .
J Diameter of the j ^ branch element, assumed
to be constant (cm).
G .
J Synaptic conductance per unit area
,0 -l
( u cm ) .
Ax
j Distance increment for each different branch
element encountered along the path to the soma (c m ) .
= 1 Ax . j
R
.
mj R e s i s t a n c e a c r o s s a u n i t a r e a o f m e m b r a n e o f t h e j ^ b r a n c h e l e m e n t
(
ß
c m ^ ).
V {<Rmj V /
4
Ri}i
C h a r a c t e r i s t i c l e n g t h p a r a m e t e r f o r e a c h d i f f e r e n t b r a n c h e l e m e n te n c o u n t e r e d a l o n g t h e p a t h t o t h e
s o m a , a s s u m e d t o c h a n g e a t e a c h b r a n c h p o i n t ( c m ) .
Z = £ ( A x
j
/ Aj ) - / ( d c / X J.) jE l e c t r o t o n i c d i s t a n c e f r o m t h e s o m a ( Z = 0 ) i s t h e s u m o f t h e A x . ,
e a c h d i v i d e d b y t h e A. f o r t h a t b r a n c h J
e l e m e n t ( d i m e n s i o n l e s s ) .
n N u m b e r o f b r a n c h e l e m e n t s a t a n e l e c t r o t o n i c d i s t a n c e Z f r o m t h e s o m a ( d i m e n s i o n l e s s ) .
n
0 N u m b e r o f p r i m a r y t r u n k s ( d i m e n s i o n l e s s ) .
D = ( I d . 3 / 2 ) 2 / 3 t a p e r ^ j
j = l
n o
D = ( [d.
3
/2^273
j=lD i a m e t e r of a u n i f o r m e q u i v a l e n t cable, with D=d^ if there is
only a s i n g l e p rimary trunk (cm).
F = D
3/2
-3/2
taper G e o m e t r i c ratio factor ( d i m e n s i o n l e s s ) .
F = D 3 ^ 2 D 3//2 taper
x {
I
R . 1/2 /I
R . 1/2L
mjL
mjj=l j = l
E l e c t r o g e o m e t r i c ratio factor ( d i m e n s i o n l e s s ) .
\ a p e r = ^ ^ m ^ t a p e r )/^ R i } 2 C h a r a c t e r i s t i c length p a r a m e t e r of a t a p e r i n g e q u i v a l e n t cable
( cm ) .
A = tt D A.
taper taper Unit area of m e m b r a n e
(
2
.
(cm ) .
A= [ ( Rm D )/4R_^ ]
*
C h a r a c t e r i s t i c l e n g t h p a r a m e t e r of a u n i f o r m e q u i v a l e n t cable ( cm ) .r = 4 R . / ( 7T a , t a p e r 1
2
D ^ ) I n t r a c e l l u l a r r e s i s t a n c e p e r t a p e r
u n i t l e n g t h o f a t a p e r i n g e q u i v a l e n t c a b l e ( f i c m ^ ).
r = 4 R . / (tt D 2 )
a l I n t r a c e l l u l a r r e s i s t a n c e p e r u n i t l e n g t h o f a u n i f o r m e q u i v a l e n t c a b l e ( f i c m ^ ).
Q o C h a r g e ( C ) .
I A p p l i e d C u r r e n t ( A ) .
I
o M a g n i t u d e o f a p p l i e d c u r r e n t a t t h e s o m a ( A ).
L E l e c t r o t o n i c l e n g t h o f a t a p e r i n g e q u i v a l e n t c a b l e ( d i m e n s i o n l e s s ) .
Z . J
L o c a t i o n o f s y n a p t i c i n p u t
( d i m e n s i o n l e s s ) .
^(Zp’Zj;T)=Kpj(T)
G r e e n ' s f u n c t i o n d e f i n e d a s t h e v o l t a g e t r a n s i e n t a t Z = Z a n dP
t i m e T i n a t a p e r i n g e q u i v a l e n t c a b l e a s a r e s u l t o f a v e r y b r i e f
( D i r a c d e l t a - f u n c t i o n ) c u r r e n t p u l s e d e l i v e r e d a t Z = Z^. ($7).
U n E i g e n v a l u e s o f t h e G r e e n ' s f u n c t i o n .
E i g e n f u n c t i o n s c o r r e s p o n d i n g t o t h e e i g e n v a l u e s .
g •
J »m a x M a x i m u m a m p l i t u d e ( p e a k ) o f s y n a p t i c c o n d u c t a n c e (fi ^ ) .
T 1
T . = a .
J ,m a x j T i m e t o p e a k o f t h e s y n a p t i c c o n d u c t a n c e c h a n g e
( d i m e n s i o n l e s s ) .
8 j ( T ) = g j , m a x a j T e x p ( 1 '-otjT) T i m e c o u r s e o f t h e s y n a p t i c c o n d u c t a n c e c h a n g e ( f 2 ^ ) .
L I S T
OF
F I G U R E
L E G E N D S
1.1 A schematic illustration of two single dendrites showing a wide range of profuseness and paucity in their branching pattern. The dashed line divides each dendrite at points of equal electrotonic distance mapping onto a sinusoidal equivalent "dendrite" cable.
1.2 Longitudinal sections of equivalent cables derived from Table 1.1 whose inhomogeniety is governed by the decline of the factor F(Z) with electrotonic distance Z. The diameters of the equivalent cables obey equation (1.20) and are simply two-thirds root of the combined DTP. Parameter v a l u e s : ot=l . 5 and u)=-1.047, except for the sinusoidal cable where the chosen values were ot = 0.15 and
I co I =1 . 1 63 .
2.1 Time course of voltage in response to a very brief (Dirac delta function) current pulse delivered at various distances along an open-circuit finite equivalent cable, whose diameter decreases exponentially at a rate determined by the constant k:(a) k = 0.0; (b) k = — 1.1; (c) k = — 1.8 ; (d) k = -4.0. The peak response for input injected at the soma
to a very brief (Dirac delta-function) current pulse, delivered at various distances from the soma, of an open-circuited finite equivalent cable, whose diameter decreases exponentially at a rate determined by the constant k:(a) k=0.0; (b) k=-l.l;(c) k=-1.8;(d) k = - 4 .0. The responses have been normalized in terms of their peaks and the abscissa begins at the peak time in each case. The continuous lines correspond to an electrotonic length L of the equivalent cable equal to 1.0, while the curves shown by the interrupted lines correspond to L=°°.
2. 4 T r a n s i e n t v o l t a g e d e c r e m e n t in a n o n t a p e r i n g e q u i v a l e n t
c a b l e ( --- ) a n d a t a p e r i n g e q u i v a l e n t c a b l e ( --- ). The
t a p e r i n g is g o v e r n e d by (a) e x p ( - l . l Z ) , (b) e x p ( - 1 . 8 Z ) ,
a n d (c) e x p ( - 4 . 0 Z ) . V a l u e s w e r e c o m p u t e d f r o m e q u a t i o n
( 2 . 2 7 ) w i t h L = 1.0; T = 0.1, 0.2, 0.5, a n d a n d n v a l u e s
f r o m 1 to 50. R o o t s of t he t r a n s c e n d e n t a l e q u a t i o n ( 2 . 2 9 )
w e r e e v a l u a t e d n u m e r i c a l l y u s i n g a N e w t o n - R a p h s o n i t e r a t i o n
s c h e m e .
2 .5 A p l o t of ß v e r s u s L for a c u r r e n t s t e p r e c o r d e d and
a p p l i e d at the s o m a of an e x p o n e n t i a l l y t a p e r i n g e q u i v a l e n t
c a b l e .
3.1 T h e F f a c t o r b a s e d on the b r a n c h i n g p a t t e r n of a cat
r e t i n a l d e l t a - g a n g l i o n c e l l v e r s u s the e l e c t r o t o n i c
d i s t a n c e . T h e s i n u s o i d a l e q u i v a l e n t c a b l e r e f e r s to the
p r o f i l e of t h i s c u r v e . A d e l t a - c e l l , t a k e n f r o m B o y c o t t
a n d W a s s l e ( 1 9 7 4 ) , is s h o w n in t he t o p c o r n e r .
3 .2 T i m e c o u r s e of t he e x c i t a t o r y an d i n h i b i t o r y
c o n d u c t a n c e c h a n g e r e p r e s e n t e d by e q u a t i o n g. oi.T e x p ( l
-1 , IT-13 X 1
a . T ) a n d g 0 a 0 T e x p ( l - a 0T), r e s p e c t i v e l y , w i t h a = 1 . 5
1 Z ,max Z Z i
a n d a 0 = 1 . 2 5 . T h e o r d i n a t e is e x p r e s s e d in t e r m s of g,
Z 1 , ma x
w h i c h is a s s u m e d to v a r y in t h e a n a l y s i s , w h i l e the
a b s c i s s a r e p r e s e n t s the t i m e s c a l e in u n i t s of the
3.3 T h e i n p u t c o n d u c t a n c e G ^ n a l o n g the s i n u s o i d a l
e q u i v a l e n t c a b l e is o b t a i n e d f r o m e q u a t i o n ( 3 . 1 0 ) and
e x p r e s s e d as the d i m e n s i o n l e s s v a r i a b l e r AG. a m
3 . 4 E l e c t r i c a l e q u i v a l e n t c i r c u i t f or a s m a l l p a t c h of
s y n a p t i c m e m b r a n e of a d i r e c t i o n a l l y s e l e c t i v e g a n g l i o n
c e l l r e c e i v i n g t wo d i s t i n c t , c l o s e l y a d j a c e n t s y n a p s e s
(a d y a d ) . T h e e x c i t a t o r y b a t t e r y h a s i t s p o s i t i v e p o l e
f a c i n g the i n t e r i o r of the p o s t s y n a p t i c m e m b r a n e b e c a u s e
the e q u i l i b r i u m p o t e n t i a l for e x c i t a t i o n is p o s i t i v e i n s i d e
(i.e. E e = 1 0 mV); t he i n h i b i t o r y a n d r e s t i n g m e m b r a n e
b a t t e r i e s h a v e t h e i r n e g a t i v e p o l e f a c i n g the i n t e r i o r
of t he p o s t s y n a p t i c m e m b r a n e b e c a u s e t h e i r e q u i l i b r i u m
p o t e n t i a l s a r e n e g a t i v e i n s i d e (i.e. E ^ - E - - 7 0 m V ) . Th e
c o n d u c t a n c e f o r e x c i t a t i o n a n d i n h i b i t i o n h a v e a r r o w s
i n d i c a t i n g t h a t t h e y c a n be c h a n g e d , but the r e s t i n g
m e m b r a n e c o n d u c t a n c e r e m a i n s f i x e d .
3 .5 T h e i m p u l s e r e s p o n s e f u n c t i o n K ( Z , Y ; T ) / r A as a r e s u l t a
of a v e r y b r i e f ( D i r a c d e l t a - f u n c t i o n ) c u r r e n t p u l s e
d e l i v e r e d at v a r i o u s l o c a t i o n s a l o n g the s i n u s o i d a l
e q u i v a l e n t c a b l e w a s c o m p u t e d f r o m e q u a t i o n ( 3 . 1 2 ) w i t h
n v a l u e s f r o m 1 to 50. T h e e i g e n v a l u e s w e r e n u m e r i c a l l y
scheme. The value of Y associated with each curve is shown.
In (a) the impulse response function is measured at the
same position as the input. The ordinate in (b) is 1/200
of the value shown in (a). The peak response at Z
=
Y
=
L
is approximately 5335. The ordinate is in dimensionless
units, while the abscissa is the time in units of the
dimensionless time variable T.
3.6 Effect of the peak synaptic conductance strength on
the peak somatic depolarization caused by the excitatory
input being activated at (a)
the soma, (b) the centre,
and (c) the distal tip of the sinusoidal equivalent cable.
Parameter values: r A=50 Mf2; V, rev =
90 mV.
a
1
3.7
Deviation of
the somatic
potential from an initial
voltage of 17 mV caused by the activation of a shunting
inhibitory conductance change at three distinct locations
along
the sinusoidal
equivalent
cable.
The excitatory
and
inhibitory
conductance
changes
are
activated
simultaneously
(as expected for stimuli moving in the
null
direction) at (a) the soma, (b) the centre, and (c)
the
distal
tip. The dashed
line
corresponds to
the
threshold level for action potential (spike) initiation.The
abscissa represents the time in units of the dimensionless
time variable
T.
Parameter
values:
(a)
g.
=10nS,
(b)
8 2 , 8
m a x
1 , m a x
= 2 0 n S .
1 5 nS , ( c ) g 1 = 1 8n S ; r A=50Mft; V , r e v = 9 0 mV;
1 , m a x a 1
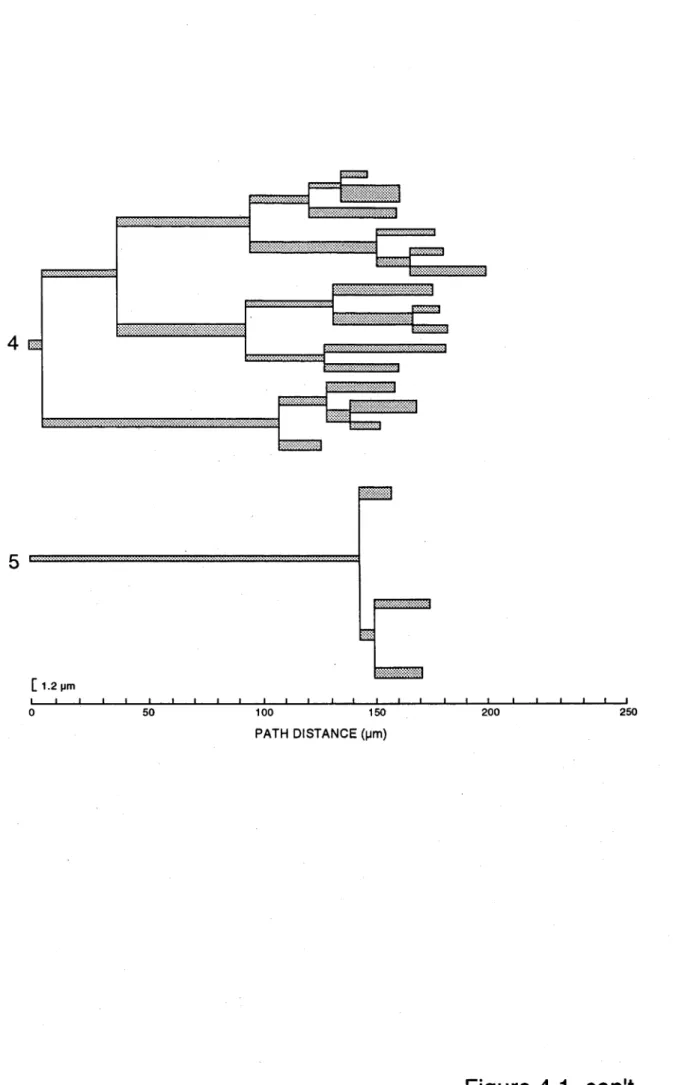
4.1 D e n d r o g r a m s of f i v e d e n d r i t e s f r o m the Cb s t a r b u r s t
a m a c r i n e c e l l p l o t t e d in t e r m s of a n a t o m i c a l p a t h d i s t a n c e
f r o m the c e l l b o d y ( a b s c i s s a ) . E a c h h o r i z o n t a l l i n e
r e p r e s e n t s a d e n d r i t i c b r a n c h . T h e t h i c k n e s s of e a c h
h o r i z o n t a l l i n e r e p r e s e n t s the a v e r a g e d i a m e t e r of the
b r a n c h ( c a l i b r a t i o n ba r on l o w e r l e f t r e p r e s e n t s 1.2 ym).
4 . 2 A c u r v e s h o w i n g t he a s s u m e d s i g m o i d a l R i n c r e a s e m
( l e f t o r d i n a t e ) a n d the c u m u l a t i v e m e m b r a n e a r e a ( r i g h t
o r d i n a t e , s c a l e d as % of t o t a l a r e a ) b o t h p l o t t e d as
f u n c t i o n s of d e n d r i t i c p a t h d i s t a n c e ( a b s c i s s a ) . H i s t o g r a m
of m e m b r a n e a r e a s in s u c c e s s i v e 10 ym b i n s is s h o w n at
b o t t o m ( C a l i b r a t i o n b ar on l o w e r r i g h t r e p r e s e n t s 90 y m 2 ).
4 . 3 T h e p a r a m e t e r (F) of the s t a r b u r s t a m a c r i n e c e l l
p l o t t e d as a f u n c t i o n of the e l e c t r o t o n i c d i s t a n c e (Z)
as c a l c u l a t e d f r o m th e d e t a i l e d g e o m e t r y of t he d e n d r i t i c
t r e e a n d a s s u m i n g a n o n u n i f o r m ' s i g m o i d a l ' R m d i s t r i b u t i o n
w i t h a s p e c i f i c a x i a l r e s i s t i v i t y R_^ of 300 f2cm. T h e d a s h e d
c u r v e r e p r e s e n t s an a p p r o x i m a t i o n of the p a r a m e t e r by
state voltage along a tapering (exponentially) equivalent
cable model plotted as a function of electrotonic distance
(Z) with Z=0 and Z=0.8 representing the soma and dendritic
terminal,
respectively.
Arrowheads denote direction of
current flow.
4.5
Tip-to-tip
signal
decrement
along
the
starburst
amacrines dendritic arbor shown as a percentage of voltage
plotted
against
the
maximum
value
of
the
spatially
nonuniform R
distribution for two different values of
m
R. shown labelled.
l
4.6 Somatofugal(--- )
and somatopetal(----)
attenuation
of
steady-state voltage along
the starburst amacrines
dendritic arbor represented as an exponential equivalent
dendrite having a specific axial resistivity (R^) of 300
ficm plotted against the maximum value of the spatially
nonuniform R distribution,
m
4.7(a)
The model predicts that a single amacrine cell
provides
excitatory
input
to
directionally
selective
subunits from 4 different groups of ganglion cells whose
preferred directions are anisotropic with respect to each
w i t h i n a s i n g l e s u b u n i t t h a t is r e d u p l i c a t e d 9 or m o r e
t i m e s t 0 c o v e r the t o t a l a r e a of a p a r t i c u l a r r e c e p t i v e
f i e l d in a g r e e m e n t w i t h the p h y s i o l o g i c a l o b s e r v a t i o n s
of B a r l o w a n d L e v i c k ( 1 9 6 5 ) . T h e r e c e p t i v e f i e l d of e a c h
d i r e c t i o n a l l y s e l e c t i v e u n i t is s h o w n to p o s s e s s
p r e f e r r e d d i r e c t i o n t h a t i s r e s t r i c t e d to on e of
n o n o v e r l a p p i n g d i r e c t i o n s in a c c o r d a n c e w i t h O y s t e r
o b s e r v a t i o n s ( O y s t e r a n d B a r l o w , 1 967; O y s t e r , 1 9 6 8 ) . E a c h
i n d i v i d u a l d i r e c t i o n - s e l e c t i v e g a n g l i o n c e l l is s h o w n
to h a v e th e s a m e a x i s of p r e f e r e n t i a l r e s p o n s e ( i n d i c a t e d
by a r r o w ) t h r o u g h o u t its r e c e p t i v e f i e l d in a c c o r d a n c e
w i t h th e e x p e r i m e n t a l r e s u l t s ( B a r l o w a nd H i l l , 1963; B a r l o w
et al . , 1 9 6 4 ) .
4 . 7 ( b ) T h e m o d e l p r o p o s e s t h a t a n y c h o s e n l o c a l r e g i o n
( s u b u n i t ) in t he r e c e p t i v e f i e l d of a s i n g l e d i r e c t i o n a l l y
s e l e c t i v e u n i t r e c e i v e s i n p u t f r o m the d i s t a l d e n d r i t e s
of s t a r b u r s t a m a c r i n e s . T h e d e n d r i t e s of s t a r b u r s t
a m a c r i n e s w i t h s o m a t a s h o w n as u n f i l l e d c i r c l e s p r o v i d e
c h o l i n e r g i c i n p u t to t he c h o s e n s u b u n i t , w h i l e the
d e n d r i t e s w h i c h m e d i a t e th e r e l e a s e of G A B A a r e s p a t i a l l y
a r r a n g e d so as to p r o d u c e a " s i l e n t " i n h i b i t o r y s u r r o u n d
( s h o w n by the d a s h e d l i n e ) t h a t is s i m i l a r in a p p e a r a n c e
to the c a r d i o i d s h a p e o b s e r v e d e x p e r i m e n t a l l y by W y a t t
t he l a t e r a l e x t e n t of t h i s i n h i b i t o r y a r e a is j u s t u n d e r
h a l f the s i z e of t he r e c e p t i v e f i e l d d i a m e t e r of the
d i r e c t i o n a l l y s e l e c t i v e g a n g l i o n c e l l a n d i t can a l s o
p r o j e c t s o m e d i s t a n c e i n t o t h e s u r r o u n d of the r e c e p t i v e
f i e l d , d e p e n d i n g on the p o s i t i o n of t h e l o c a l r e g i o n
( s u b u n i t ) in the r e c e p t i v e f i e l d ( W y a t t a n d D a w , 1975).
F u r t h e r m o r e , g i v e n t h a t the d i r e c t i o n a l l y s e l e c t i v e u n i t
h a s a r e c e p t i v e f i e l d s l i g h t l y g r e a t e r t h a n the d e n d r i t i c
f i e l d of a s t a r b u r s t a m a c r i n e c e l l at e a c h e c c e n t r i c i t y ,
t he s i l e n t - i n h i b i t o r y s u r r o u n d s h o u l d e x t e n d for o n l y
h a l f the w i d t h of a s t a r b u r s t a m a c r i n e c e l l ( V a n e y et
a l . , 1 9 8 9 ; V a n e y , 1 9 9 0 ) . T h e s i z e of t h e s i l e n t i n h i b i t o r y
s u r r o u n d w o u l d be d e p e n d e n t on t h e e c c e n t r i c p l a c e m e n t
of the s t a r b u r s t a m a c r i n e c e l l w i t h r e s p e c t to the v i s u a l
s t r e a k in r a b b i t or a r e a c e n t r a l i s in c a t . For e x a m p l e ,
in the v i s u a l s t r e a k t he d e n d r i t i c f i e l d d i a m e t e r s are
in the r a n g e of 2 5 0 jjm > w h i l e in t he p e r i p h e r a l r e t i n a
t h e y r a n g e up to 8 00 ym, d e m o n s t r a t i n g a l a r g e v a r i a t i o n
w i t h r e t i n a l e c c e n t r i c i t y ( T a u c h i a n d M a s l a n d ,1984) . T he
p r e f e r r e d d i r e c t i o n of the d i r e c t i o n a l l y s e l e c t i v e u n i t
e n c o m p a s s e d by t he n e t w o r k of s t a r b u r s t a m a c r i n e c e l l s
4 . 8 T h e a n a t o m i c a l s t r u c t u r e of the m o d e l c o n s i s t s of
a d i r e c t i o n a l l y s e l e c t i v e g a n g l i o n c e l l w i t h a r e c e p t i v e
f i e l d s u b t e n d i n g 4^° a n d s e q u e n c e - d i s c r i m i n a t i n g s u b u n i t s
m a d e up f r o m the d i s t a l d e n d r i t e s of s t a r b u r s t a m a c r i n e
c e l l s a c t i v a t e d by c o n e b i p o l a r c e l l s . T h e i n h i b i t o r y
m e c h a n i s m for i m p l e m e n t i n g the s e q u e n c e - d i s c r i m i n a t i o n
c o r r e s p o n d i n g to a s i n g l e s u b u n i t is s h o w n to be e x c i t a t e d
by b i p o l a r i n p u t s 1 7 ’ a p a r t , w h i l e s u c c e s s i v e s u b u n i t s
a r e s e p a r a t e d by b i p o l a r i n p u t s 12' a p a r t . T h i s is in
a g r e e m e n t w i t h th e s i n g l e - s l i t e x p e r i m e n t , w h e r e b y the
c o m p l e t e m e c h a n i s m of d i r e c t i o n a l s e l e c t i v i t y w a s f o u n d
to be c o n t a i n e d w i t h i n a s m a l l s u b u n i t 1/9 to 1 / 1 8 of
th e e n t i r e r e c e p t i v e f i e l d ( B a r l o w a n d L e v i c k , 1 9 6 5 ) . T h e
e x i s t e n c e of an " i n h i b i t i o n - f r e e " z o n e a d j a c e n t to the
e d g e of t h e r e c e p t i v e f i e l d t h a t is f i r s t c r o s s e d w h e n
m o t i o n is in t he p r e f e r r e d d i r e c t i o n e x t e n d s a p p r o x i m a t e l y
i° f r o m t h e e d g e . A s e r i a l s y n a p s e is s h o w n e n l a r g e d w i t h
(+) d e n o t i n g an e x c i t a t o r y s y n a p s e a n d (-) an i n h i b i t o r y
s y n a p s e . T h e s e r i a l s y n a p s e s a r e a r r a n g e d s u c h t h a t an
a m a c r i n e - a m a c r i n e s y n a p s e u s e s G A B A to i n h i b i t the
a m a c r i n e - g a n g l i o n c e l l s y n a p s e . W i t h the s e r i a l s y n a p s e s
o r g a n i z e d as in the d i a g r a m , a s p o t m o v i n g f r o m l e f t to
r i g h t ( n u l l d i r e c t i o n ) w i l l r e s u l t in no f i r i n g in t he
g a n g l i o n c e l l (cf. D o w l i n g , 1 9 7 0 ) . (A) d e n o t e s s t a r b u r s t
a m a c r i n e c e l l s ; (B) d e n o t e s c o n e b i p o l a r c e l l s ; (G) d e n o t e s
C H A P T E R
1
M E M B R A N E
V O L T A G E
C H A N G E S
IN
P A S S I V E
D E N D R I T I C
T R E E S :
A
T A P E R I N G E Q U I V A L E N T C A B L E M O D E L
§ 1 . 1 I n t r o d u c t i o n
A particular class of dendritic tree can be represented by a single equivalent cylinder, if several symmetry requirements hold [for a comprehensive summary see Rail ( 1977, 1989) and the lucid monograph by Jack
et al. (1975)]. The equivalent cylinder concept was formulated by Rail ( 1962a,b) over two decades ago and it provided neurophysiologists with tremendous insight into the role dendrites play in neuronal functioning (see e.g. Redman, 1976). It is based on four symmetry requirements, which restrict its application in neuronal modelling. In particular, the assumption that synaptic input must be equal at all points that are at the same electrotonic distance from the soma is a significant restriction, since local interactions between inputs on different branches cannot be investigated. In view of
the above limitation more recent models of the equivalent cylinder type have been constructed by Redman (1973) and
The 3/2 power law is based on the following two conditions, which must hold if a dendritic tree is to be transformed into an electrotonically equivalent cylinder :
(1) At every branch point, the parent branch diameter (assumed constant) raised to the 3/2 power
must equal the sum of the daughter branch
diameters (not necessarily equal, but assumed to be constant) each raised to the 3/2 power.
(2) The sum of the 3/2 power of all branch diameters at any given electrotonic distance from the soma must remain constant to the point of termination.
Implicit in these two conditions is a further symmetry requirement that all terminal branches must end at the same electrotonic distance from the soma.
As most neurons are not of the equivalent cylinder class, but instead show a decline in the dendritic trunk parameter, caused by a deviation from the 3/2 power law at branch points, completely new models not dependent on any of the symmetry requirements have been developed (see e.g. T u r n e r , 1984; Koch and Poggio, 1985; Segev et a l ., 1985; Holmes, 1986; Carnevale and L e b e d a ,1987). However, all these models have the disadvantage that analytic solutions are not directly available.
assumption can be modified by introducing the notion of taper into the model. This modification to the 3/2 power law enables a variety of different dendritic trees with a relative paucity of branching (caused by the early termination of individual branch segments) to be treated, and the symmetry requirement that terminal branches must end at the same electrotonic distance from the soma is therefore no longer required.
A dendritic tree can have many forms and still be equivalent to the same equivalent cable. An example of a dendritic tree that maps onto a sinusoidal equivalent cable is shown in Fig. 1.1. However, there is a large
difference between dendritic trees with dendrites
SOMA
Electrotonic
Distance
Sinusoidal Equivalent Dendrite
Dendrite
(X F[Z]
SOMA
Electrotonic
Distance
Sinusoidal Equivalent Dendrite
T h e a d v a n t a g e i n u s i n g t h i s m o d e l is t h a t o n l y a
s i n g l e c a b l e e q u a t i o n n e e d s to be s o l v e d an d i t is
t h e r e f o r e e a s y to i m p l e m e n t . F or e x a m p l e , the G r e e n Ts
f u n c t i o n c a n be o b t a i n e d by a n a l y t i c a l r a t h e r t h a n
c o m p u t a t i o n a l m e t h o d s , a n d as a r e s u l t t h e r e is no r i s k
of e r r o r c a u s e d by n u m e r i c a l i n v e r s i o n of the t r a n s f e r
f u n c t i o n (s e e e.g. K o c h a n d P o g g i o , 1 9 8 5 ) . A n o t h e r
i m p o r t a n t a d v a n t a g e is its a b i l i t y to a n a l y t i c a l l y
i n v e s t i g a t e t he e f f e c t s of p a r a m e t e r c h a n g e s i n c o r p o r a t e d
in the m o d e l , a n d a l s o t he l u x u r y of h a v i n g c l o s e d f o r m
s o l u t i o n s a v a i l a b l e . H e n c e , the a n a l y t i c a l t r a c t a b i l i t y
of the m o d e l p r o v i d e s t he i m p e t u s for f u r t h e r w o r k in
u n d e r s t a n d i n g the f u n c t i o n a l r o l e d e n d r i t e s p l a y in v a r i o u s
i n f o r m a t i o n p r o c e s s i n g t a s k s , s u c h as d i r e c t i o n a l
s e l e c t i v i t y in t he m a m m a l i a n r e t i n a . T h i s is t h e m a j o r
a i m of t h i s t h e s i s .
A l t h o u g h , R a i l r e c o g n i s e d t h a t h i s t h e o r y c o u l d
be e x t e n d e d to e x p o n e n t i a l l y t a p e r e d e q u i v a l e n t c a b l e s ,
he did n o t d e v e l o p t he t h e o r y to c o v e r s u c h c a s e s w i t h
e x p l i c i t s o l u t i o n s , a n d f u r t h e r m o r e the t h e o r y f or the
m o r e g e n e r a l c l a s s of t a p e r i n g e q u i v a l e n t c a b l e s r e m a i n s
u n p u b l i s h e d (but s e e S c h i e r w a g e n , 1 9 8 9 ) .
In o r d e r to u n d e r s t a n d th e r o l e c h a n g i n g g e o m e t r y
p l a y s in c o n t r o l l i n g n e u r o n a l a c t i v i t y t h e o r e t i c a l w o r k s
d e a l i n g w i t h t a p e r s h a v e b e e n p u b l i s h e d by s e v e r a l a u t h o r s
d e n d r i t i c b r a n c h e s p a r a m e t e r is e q u a l to u n i t y (s e e
G o l d s t e i n a n d R a i l , 1974; S t r a i n a n d B r o c k m a n , 1975; K e l l e r
a n d Lai, 1976; B r o c k m a n , 1981; E l l i a s a n d S t e v e n s , 1983;
R o s e a n d D a g u m , 1 9 8 8 ) .
T h e c l a s s i c a l w o r k of G o l d s t e i n a n d R a i l ( 1 9 7 4 ) d e a l t
w i t h the e f f e c t s of c h a n g e s in the s h a p e a n d v e l o c i t y
of i m p u l s e s as t h e y p r o p a g a t e d a l o n g n o n c y l i n d r i c a l a x o n s
w h i c h t a p e r e d e x p o n e n t i a l l y w i t h d i s t a n c e . H o w e v e r , the
f i r s t m a j o r w o r k to a p p e a r in t he l i t e r a t u r e w h i c h i n v o l v e d
n o n a x o n i c n e r v e p r o c e s s e s w i t h g e o m e t r i c a l i n h o m o g e n e i t i e s
in t he f o r m of t a p e r i n g w a s by S t r a i n a n d B r o c k m a n ( 1 9 7 5 ) .
T h e y c a l c u l a t e d t he s t e a d y - s t a t e v o l t a g e d e c a y in a p a s s i v e
n e r v e c y l i n d e r for t h r e e d i f f e r e n t g e o m e t r i e s by
n u m e r i c a l l y i n t e g r a t i n g the d i f f e r e n t i a l e q u a t i o n . F or
a l i n e a r t a p e r , t h e d e c a y of t he v o l t a g e w a s f o u n d be
s l o w e r t h a n t h a t of a u n i f o r m c y l i n d e r . U n f o r t u n a t e l y ,
a n a l y t i c a l s o l u t i o n s f or t he t i m e - v a r i a n t c a s e w e r e not
o b t a i n e d , b u t n u m e r i c a l m e t h o d s h a v e b e e n u t i l i z e d t h a t
e n a b l e t h e t r a n s i e n t r e s p o n s e in g e o m e t r i c a l l y
i n h o m o g e n e o u s n e r v e c y l i n d e r s to be d e t e r m i n e d (s e e e.g.
K e l l e r a n d Lai, 1976; B r o c k m a n , 1981; E l l i a s a n d S t e v e n s ,
§1.2 Derivation of the governing equation
The formulation given here of the equation associated with a tapering equivalent cable in the presence of
synaptic input proceeds along different lines from that previously presented by Jack et al.(1975). All the quantities are expressed in terms of the generalized electrotonic distance Z, under the assumption that all branch elements have constant but not identical diameters. In addition, the factor F which imposes a nonuniformity on the equivalent cable is incorporated into the equation, together with effects of synaptic action modelled by a change in postsynaptic membrane conductance.
In a nonuniform structure, the membrane current per unit area is given by
-1 3 la
Im = [ttADF(Z) ] { } 3Z
(
1
.
1
)
where the total axial current is expressed as
la = [ra,taper \ a p e r ] { }
(
1
.
2
)
3Zy i e l d s t he f o l l o w i n g r e s u l t
Im =
-1
F dF av 3 V dZ 3Z 3 Z 2
(1.3)
T h e t o t a l m e m b r a n e c u r r e n t per u n i t a r e a of the
s y n a p t i c m e m b r a n e c a n be w r i t t e n as the s u m of the i o n i c
a n d c a p a c i t i v e c u r r e n t s , t h a t is,
av
rev
rev
Im = C m — + G r V + G e ( V - V e ) + G i ( V - V i ) (1.4) 31
w h e r e it is a s s u m e d t h a t s y n a p t i c a c t i v i t y a s s o c i a t e d
w i t h e x c i t a t i o n or i n h i b i t i o n on a d e n d r i t i c t r e e o c c u r s
on a l l of t h e b r a n c h e l e m e n t s t h a t a r e at the s a m e
e l e c t r o t o n i c d i s t a n c e f r o m t he s o m a . It is i m p o r t a n t to
s t r e s s t h a t t he m e m b r a n e r e s i s t a n c e a c r o s s a u n i t a r e a
of s y n a p t i c m e m b r a n e is no l o n g e r p a s s i v e b e c a u s e it is
a l s o d e p e n d e n t on t he s y n a p t i c c o n d u c t a n c e c h a n g e s , t h a t
is
R m
*
+ G + G. )
e l ( 1.5)
but, if e a c h s y n a p t i c c o n d u c t a n c e c h a n g e is a s s u m e d to
o c c u p y an i n f i n i t e s i m a l r e g i o n r e p r e s e n t e d s y m b o l i c a l l y
p r o p e r t i e s will not c h a n g e (Poggio and Torre, 1978). By p a s s i v e it is a s s u m e d that for a unit area of m e m b r a n e the c a p a c i t a n c e and the r e s i s t a n c e in p a r a l l e l are i n d e p e n d e n t of both time and voltage.
E q u a t i n g e q u a t i o n s (1.3) and (1.4), and m u l t i p l y i n g both s i d e s by , a m o d i f i e d cable e q u a t i o n d e s c r i b i n g
m e m b r a n e v o lt a g e c h a n g e s in the p r e s e n c e of s y n a p t i c input is o b t a i n e d :
av = a_^_v +
xdF av
3T
az2
dz az
2
v ,
l
r „ A 6 ( Z - Z .)g.(T) V +L
a , t ap e r taper j jj = l
x [V . rev - V] (1.6)
w h e r e the s u m m a t i o n s u b s c r i p t j = l , 2 d e n o t e s e x c i t a t i o n and i n h i b it i o n , r e s p e c t i v e l y , and time has been scaled, or cast in d i m e n s i o n l e s s form, in terms of the m e m b r a n e time c o n s t a n t . Note that the s y n a p t i c c o n d u c t a n c e c h a n g e s per unit area G and G.
e l have been c o n v e r t e d into g^ = AGe and 8 2 = A G . , r e s p e c t i v e l y , w h ere A is the unit areal o f
m e m b r a n e . In the a b s e n c e of s y n a p t i c input the last term on the right hand side of e q u a t i o n (1.6) v a n i s h e s and
the e q u a t i o n becomes:
a2v
az2
- l
av
3T
V + F dF
av
dz az
It is emphasized that equation (1.7) also arises in circuit theory as an equation representing an inhomogeneous (nonuniform) RC network or transmission line. As a result, similar solutions have previously been derived by Kelly and Ghausi (1965).
§1.3 Condition for reduction to a tapering equivalent
cable
The reduction of a dendritic tree to a single tapering equivalent cable is analogous to the approach used by Rail (1962a) in constructing an equivalent cylinder representation of the dendritic tree, with the exception that the so-called 3/2 power law at branch points need not hold.
The following parabolic partial differential equation describes passive membrane potential distribution in a dendritic tree with noncylindrical branches (Rail, 1962a, equations 20 and 23; Jack et al., 1975, equation 7.42):
9 2 V 9Z 2
3V
3Z
[(—
)
-1
d x
d_ d x
3/2
ln{ r n
[i+ (—
>2 ]4} ]
d x(
1.
8)
3V_ 9T
where the radius (r) of all branch segments and the number of dendritic branches (n) are both functions of actual distance (x) from the soma, and
Z
X
/ d?/A
0
taper (1.9)
defines the electrotonic distance for situations where there is a continuously changing characteristic length parameter (A ) :
13 p g r
taper (r/ro ) [1 + ( — ) 2
d x
- i
(
1.
1 0)
where A =[R r / 2 R . 1 2 is the characteristic length parameter o m o l
for a cylinder with radius rQ (taken as the initial radius at x=0). The generalized electrotonic distance and
characteristic length Z and A respectively, are
L cL p 0 IT
concepts first introduced by Rail ( 1962a,b) and illustrated in some detail for an exponentially tapering core conductor by Goldstein and Rall(1974).
single one-dimensional cylinder. The following condition between r and n must hold for such a branching pattern
(see also Jack et al., 1975, equation 7.43):
3/2
2
i
nr [1 + ( — — ) ] = constant (1.11) d x
However, since our aim is to determine combinations of tapering and branching which will allow the dendritic
tree to be electrotonically equivalent to a tapering cable [represented by equation (1.7)], the following condition between r and n must hold (cf. Rail, 1962a, equation 21; and also Jack et al., 1975, equation 7.59 for the special case of exponential taper):
3/2 ckr 2 i 3/2
n r [ l + ( ) ] = n r F(Z;k) (1.12)
a „ o o
where n is the number of branches at x=0 and F is a factor o
which imposes taper on the equivalent cable. As a result, the coefficient of 3V/3Z in equation (1.8) simplifies to F 1(dF/dZ) and the equation describing passive membrane
electrotonus in a branching dendritic tree (with
It has been shown that the class of dendritic trees which are represented as equivalent cylinders , satisfy the property that the rate of change of dendritic surface ar ea , with respect to x, remains proportional to the rate of change of electrotonic d i s t a n c e , with respect to x
( R a i l , 1962a, equation 25 ; Jack et al. ,1975 , equation 7.45):
d A dZ
oc (1.13)
d x d x
However, for the larger class of dendritic trees represented by tapering equivalent cables, the following property must hold:
dA dZ
- F (Z ;k )[ ] (1.14)
d x d x
or alternatively
d A
« F (Z ;k ) (1.15)
dZ
If for convenience, the analysis can be restricted to the case where each dendritic branch is represented by a cylinder of constant diameter [dr/dx=0] and each individual branch at any given value of x or Z is characterized by a different diameter, then after replacing radius with diameter, together with the above assumptions, the condition on r and n governed by equation (1.12)
becomes (cf. Rail, 1962b. p.149 for the special case of exponential taper):
[ X d j 3/2 ] F(Z;k) j = l
n ( x )
: I d
j=l
3/2
(1.16)
which defines the combined DTP, where d^. represents the diameter of the branch at distance x from the soma, with Z=0 when x=0. Alternatively, if x represents the actual distance measured along successive branch points and branching occurs at distances 0 =x o < #,,<Xp with n^ branches between x. and x . (1, where x.^x^x..,, then an
l l+l l l+l
equivalent representation of equation (1.16) in more standard notation is (Goldstein and Rail, 1974, equation 32) :
F (Z ;k ) =
I d± .3/2
[I
do j 3/2 ] 1 i=0,l,...,p (1.17)Equation (1.17) simply states that the combined DTP at any given electrotonic distance from the soma is proportional to the factor F. It should be emphasized that the assumption [dr/dx=0] imposes a physical limitation in interpreting equation (1.17), because F is assumed to decrease continuously with Z rather than in discrete steps at each branch point (cf. Jack et al., 1975, Fig. 7.11). However, the theory can be extended to branches which taper at a steady rate and satisfy the condition [d r/d x ]2« i , but only if all the branches are assumed to be equal in diameter (see Jack et al., 1975, p.156).
If F is unity then the combined DTP is constant, permitting a dendritic tree to be transformed into an equivalent cylinder that has a diameter equal to
n
D = [
l
o J 3/2 ]2/3 d (1.18)j=l
or D=dQ^ if there is only a single dendrite emanating from the soma. Also, the characteristic length parameter
is given by
A [R D/4R. ]
m l
(1.19)
of the equivalent cable D changes continuously with
l0. p 0 r electrotonic distance from the soma
Dta per=D[F(Z;k)l2/3=[^d ij3/2]2/3
i = 0 ’1 .... P
(1-20)
j-1
and the characteristic length parameter, reduces from equation (1.10) to become
X
taper
[ R D
m taper/4iq J
*
(
1
.
2 1
)
If F is greater than unity the branching pattern will exhibit a wide range of profuseness, and if F is less than unity there will be a relative paucity of branching in comparison with the branching pattern of a dendritic tree which can be reduced to an equivalent cylinder. Note that at Z=0, equation (1.20) reduces to D =D, and
18 p g r
§1.4 Condition for reduction to a tapering equivalent cable with electrical and geometrical nonuniformities
The aim of this section will be to generalize the condition for reduction to a geometrically tapering equivalent cable, to include electric nonuniformities such as a spatially nonuniform R distribution, which
m
has recently attracted a lot of attention from several workers (see Rail, 1982; Redman et al . , 1987; Fleshman et al.,1988). It should be mentioned that numerical methods employed in compartmental models of neurons with different Rm values in each branch segment is an alternative approach that will not be discussed here [see Rail (1990) for a critical analysis of this technique with regard to the problem of nonuniqueness].
The derivation will be carried out in the steady- state domain, but extension to the time-dependent domain can be carried out along the same lines as sketched by Leibovic (1972) for a symmetrical tree.
[r ( x )R m (x ) / 2Ri ]
ll +
II (II) :I_
dx d Z 2 dZ dx dxIn [ r 2 ( x )n( x )— ]} = d x
V
(
1
.
2 2
)
w h e r e t he r a d i u s (r) of a ll b r a n c h s e g m e n t s , the n u m b e r
of d e n d r i t i c b r a n c h e s (n) a n d R a r e a ll f u n c t i o n s of m
a c t u a l d i s t a n c e (x) f r o m the s o m a ( a s s u m e d to be the p o i n t
x = 0 ) . T h e v a r i a b l e Z is d e f i n e d by e q u a t i o n ( 1 . 9 ) , e x c e p t
t h a t t he c h a r a c t e r i s t i c l e n g t h p a r a m e t e r ^ t a p e r ^ as the
f o l l o w i n g f o r m :
t a p e r [ R ( x ) r ( x ) / 2R m l ] ( 1 . 2 3 )
w h e r e R^ is the a x i a l r e s i s t i v i t y a s s u m e d to be c o n s t a n t .
L i k e w i s e , t he g e n e r a l e q u a t i o n for s t e a d y - s t a t e
v o l t a g e d i s t r i b u t i o n in a s i n g l e o n e - d i m e n s i o n a l u n i f o r m
c a b l e h a s the f o l l o w i n g f o r m ( J a c k et al., 1975, p . 1 4 8 ) :
( 1 . 2 4 )
T h e a i m w i l l n o w be to r e l a t e e q u a t i o n ( 1 . 2 4 ) w i t h
e q u a t i o n ( 1 . 2 2 ) by s o m e k i n d of p o w e r law. I n t e g r a t i n g
e q u a t i o n (1 . 9 ) w i t h r e s p e c t to x a n d s u b s t i t u t i n g t he
r e s u l t i n t o e q u a t i o n ( 1 . 2 2 ) y i e l d s th e f o l l o w i n g